

Abstract
The two major cadherins of endothelial cells are neural (N)-cadherin and vascular endothelial (VE)- cadherin. Despite similar level of protein expression only VE-cadherin is located at cell–cell contacts, whereas N-cadherin is distributed over the whole cell membrane. Cotransfection of VE-cadherin and N-cadherin in CHO cells resulted in the same distribution as that observed in endothelial cells indicating that the behavior of the two cadherins was not cell specific but related to their structural characteristics. Similar amounts of α- and β-catenins and plakoglobin were associated to VE- and N-cadherins, whereas p120 was higher in the VE-cadherin complex. The presence of VE-cadherin did not affect N-cadherin homotypic adhesive properties or its capacity to localize at junctions when cotransfectants were cocultured with cells transfected with N-cadherin only. To define the molecular domain responsible for the VE-cadherin–dominant activity we prepared a chimeric construct formed by VE-cadherin extracellular region linked to N-cadherin intracellular domain. The chimera lost the capacity to exclude N-cadherin from junctions indicating that the extracellular domain of VE-cadherin alone is not sufficient for the preferential localization of the molecule at the junctions. A truncated mutant of VE-cadherin retaining the full extracellular domain and a short cytoplasmic tail (Arg621–Pro702) lacking the catenin-binding region was able to exclude N-cadherin from junctions. This indicates that the Arg621–Pro702 sequence in the VE-cadherin cytoplasmic tail is required for N-cadherin exclusion from junctions. Competition between cadherins for their clustering at intercellular junctions in the same cell has never been described before. We speculate that, in the endothelium, VE- and N-cadherin play different roles; whereas VE-cadherin mostly promotes the homotypic interaction between endothelial cells, N-cadherin may be responsible for the anchorage of the endothelium to other surrounding cell types expressing N-cadherin such as vascular smooth muscle cells or pericytes.
Cadherins are a family of transmembrane glycoproteins that mediate calcium-dependent cell–cell adhesion (1, 15, 55). These adhesion receptors are expressed by virtually all cell types that form solid tissues. Cadherins are organized in junctional structures called adherens junctions. In these junctions, cadherins are clustered and connected through their cytoplasmic domain with a complex network of cytoskeletal proteins (23, 26, 28, 44). Cadherins are important in regulating morphogenesis (15, 22, 47, 55). Through their homophilic interactions, they play a role in sorting cells of different lineages during embryogenesis, establishing cell polarity, and maintaining tissue morphology and cell differentiation. Classical cadherins share a common basic structure consisting of an extracellular domain, which contains four major repeats, a transmembrane and a cytoplasmic domain. The short cytoplasmic region directly binds to three homologous proteins that belong to the “armadillo” family (β-catenin, plakoglobin, and p120). β-catenin and plakoglobin bind α-catenin, which is homologous to vinculin and mediates the binding of the cadherin–catenin complex to the actin cytoskeleton.
Comparison of different cadherin sequences shows strong homology at the amino acid level (26, 55). Conservation is particularly striking in the cytoplasmic tail where the catenin-binding region is located.
Cadherins present a certain degree of cell type specificity, for instance epithelial (E)-cadherin is preferentially expressed in cells of epithelial origin (53, 54), and neural (N)-cadherin1 in cells of the nervous tissue (17, 18), vascular smooth muscle cells, and myocytes (8, 19, 45, 60). More than one cadherin may be expressed in the same cell type, but how these molecules interact with each other and how they can contribute to the transfer of specific intracellular signals is still unknown.
The two major cadherins in the endothelium are vascular endothelial (VE)- and N-cadherin (10, 32, 49). VE-cadherin is cell specific and is strictly located at intercellular junctions of essentially all types of endothelium, both in vitro and in vivo (10, 32). Compared with the classical cadherins, the VE-cadherin amino acid sequence shows considerable differences (only 23% identity when compared with classical cadherins such as E-, N-, and placental (P)- cadherins) (5, 6).
N-cadherin is present in significant amounts in the endothelium, but surprisingly, it is not clustered at cell–cell junctions but remains diffuse on the cell membrane (50). This observation raises several questions about the mechanisms that regulate cadherin clustering at adherens junctions, and raises the possibility that a hierarchy does exist between cadherins, which regulates their functional behavior and signaling properties in a cell type–specific way.
In this report, we have investigated the mechanisms responsible for the differential membrane targeting of N- and VE-cadherin in the endothelium. We have found that VE-cadherin presents structural features that are responsible for its ability to exclude N-cadherin from cell–cell contacts. This dominant activity of VE-cadherin requires a short cytoplasmic region of the molecule, which is distinct from the previously described catenin-binding domain. Overall these observations suggest that cadherins might present different functional behaviors depending on the other cadherins expressed.
Materials and Methods
All reagents were purchased from Sigma Chemical Co. (St. Louis, MO), unless indicated otherwise.
Antibodies
Mouse mAbs against the extracellular domain of human VE-cadherin were: clone TEA 1.31 (36), clone BV9 (Hemeris, Grenoble, France) (34, 36), and clone BV6 (Bioline Diag., Torino, Italy) (34). Anti–human N-cadherin polyclonal antibodies against the whole molecule (1447; see reference 3) or the cytoplasmic domain (R156; see reference 13), and rabbit pan-cadherin antiserum against the conserved cytoplasmic sequence of all cadherins (13), were a gift from Dr. B. Geiger (Weizmann Institute, Rehovot, Israel). Mouse mAbs against the extracellular (8C11) or cytoplasmic domain (13A9) of human N-cadherin were donated by Dr. M. J. Wheelock (University of Toledo, Toledo, OH). Mouse mAbs against α-catenin, β-catenin, plakoglobin, and p120 were purchased from Transduction Laboratories (Lexington, KY). Fluorescein- and rhodamine-conjugated secondary antibodies (reactive with either mouse or rabbit IgG) were purchased from DAKOPATTS (Copenhagen, Denmark).
Goat anti–mouse IgG peroxidase–conjugated and protein A peroxidase–conjugated mAbs used for immunoblotting detection were from Pierce Chemical Co. (Rockford, IL,), and anti-phosphotyrosine HRP-conjugated mAb (RC20) was purchased from Transduction Laboratories.
Cells
Endothelial cells (ECs) were harvested from human umbilical veins and cultured as previously described (34). CHO cells were routinely maintained on culture dishes in DME containing 10% FCS, and kept at 37°C in 5% CO2. Transfection of CHO cells with the full-length cDNA of human N-cadherin (CHO N), full-length cDNA of human VE-cadherin (CHO VE), and cytoplasmic truncated mutant cDNA of human VE-cadherin (CHO tVE) was previously described (6, 41, 50).
Sterile plastic ware was from Falcon (Becton Dickinson Labware, Lincoln Park, NJ) whereas both culture medium and serum were from GIBCO BRL (Paisley, Scotland).
Plasmids Construction
Vectors containing the entire coding sequence of human VE-cadherin and N-cadherin have been described elsewhere (6, 50).
Cytoplasmic truncated mutant cDNA of human VE-cadherin (tVE) was generated by removing the coding region for the last 82 amino acids (aa) of the cytoplasmic domain as described previously (41).
VEecNcyt plasmid contains a chimera formed by the extracellular (ec) domain of VE-cadherin and the cytoplasmic (cyt) region of N-cadherin. The chimera was constructed by substitution of the VE-cadherin cytoplasmic coding region with the N-cadherin cytoplasmic coding region. The human VE-cadherin cDNA cloned into the pBlueScript vector (6) was digested with BglI/BamHI to remove the coding region from the 1,519-bp position to the end of the cDNA. To preserve the VE-cadherin coding sequence from the 1,519-bp to the 1,884-bp position (comprising the transmembrane domain), we generate a PCR fragment using 5′ and 3′ end primers matching at the appropriate positions and containing the BglI/ BamHI sites for cloning. The resultant plasmid, corresponding to the extracellular and transmembrane coding region of VE-cadherin cDNA, was called pVEec. N-cadherin cytoplasmic domain coding sequence was generated by PCR using 5′ and 3′ end primers matching at the 2,701-bp and the 3,215-bp positions of N-cadherin cDNA (provided by Dr. B. Geiger, Weizmann Institute), and containing BamHI and XbaI sites, respectively. This fragment was further cloned into pVEec digested with BamHI/XbaI. The construct was subsequently checked by sequence analysis using the dideoxynucleotide chain termination method (52). This plasmid was then cut with KpnI and XbaI enzymes, and the coding sequence insert was subcloned into the pECE eukaryotic expression vector (12).
VEec tNcyt chimera plasmid contains VE-cadherin extracellular and transmembrane coding sequence, and the cytoplasmic domain of N-cadherin lacking the last 77 aas. N-cadherin deleted cytoplasmic coding region was generated by PCR using the 5′ end primer containing the BamHI site described above and a 3′ end primer matching at the 2,944-bp position of N-cadherin cDNA, and containing stop codon sequences and an XbaI site for cloning. The fragment encodes 83 aas (Lys746–Pro828) of the cytoplasmic domain of N-cadherin lacking the COOH terminus Gly829–Asp906 (77 aas). This fragment was cloned into pVEec and subcloned into pECE as described above.
Transfection Procedure
As expression control of chimeric molecules described in the previous section, CHO cells were transiently transfected with VEecNcyt and VEec tNcyt plasmids using lipofectin reagent (GIBCO BRL). Cells were grown on glass coverslips, fixed 60 h after transfection, and then tested for chimera expression by immunofluorescence using anti–VE-cadherin mAb BV9 and anti–N-cadherin mAb 13A9, or polyclonal antibody R156.
For obtaining double transfectants, CHO N cells were stably transfected with VE-cadherin. Cells were plated at 3–4 × 106 cells per 100-mm petri dish in DME with 10% FCS. 24 h later they were cotransfected by calcium phosphate precipitation method (51) using 20 μg of VE-cadherin into pECE vector and 2 μg of pBSpacΔp plasmid containing puromycin resistance (a gift from Dr. J. Ortin, Centro Nacional Biotecnología, Madrid, Spain; see reference 9). After 24 h, the DNA-containing medium was replaced by fresh DME containing 10% FCS, and then cultured for another 48 h. Cells were then trypsinized, plated at a density of 106 per 100-mm petri dish, and then cultured in selective medium with 7 μg/ml puromycin (Sigma Chemical Co.). Resistant colonies were isolated and tested for VE-cadherin expression by immunofluorescence analysis. Positive cells were subcloned and used for further studies.
A similar procedure was used to generate double transfectants CHO N/tVE (cotransfected with N and tVE-cadherins), CHO N/VEecNcyt (cotransfected with N-cadherin and VEecNcyt chimera), and CHO N/VEec tNcyt (cotransfected with N-cadherin and VEec tNcyt-truncated chimera).
Immunohistochemistry
Human tissue specimens of different origin were embedded in optimal cryopreserving tissue compound (Ames Miles Laboratories Inc., Naperville, IN), snap frozen in liquid nitrogen, and then stored at −80°C until processed. Cryostat sections were fixed in acetone for 10 min at 22°C and immunostained with the anti–VE-cadherin mAb TEA 1.31 or anti–N-cadherin polyclonal antibody 1447, using avidin–biotin–peroxidase complex technique. The specimens were then processed and analyzed as previously described (33).
Immunofluorescence Microscopy
Cells were grown on glass coverslips (for EC coated with human plasma fibronectin, 7 μg/ml), rinsed in PBS, and fixed in methanol. The cells were then rinsed and incubated for 45 min at 37°C with the relevant primary antibodies (against VE- or N-cadherins), washed three times with PBS, and then incubated for 30 min with the fluorophore-conjugated secondary antibodies. For double staining, the coverslips were incubated with mouse anti–VE-cadherin (BV6 or BV9) in combination with rabbit anti–N-cadherin (polyclonal antibody 1447), followed by TRITC-coupled, goat anti– mouse in combination with FITC-coupled, goat anti–rabbit. Coverslips were then mounted in Mowiol 4-88 (Calbiochem-Novabiochem Corp., La Jolla, CA) and examined with an Axiophot microscope (Carl Zeiss Inc., Thornwood, NY). Photographs were taken using Tmax P3200 films.
For mixed culture studies (see Fig. 6), the two types of cells (CHO N and CHO N/VE) were seeded in small aggregates and let grow until confluency. After methanol fixation, glass coverslips were processed for double immunofluorescence as described above.
Figure 6.

N- and VE-cadherin distribution in mixed cultures of CHO N and CHO N/VE. CHO N and double-transfected CHO N/VE cells were seeded separately in small aggregates and cocultured on glass coverslips. Cells were grown to confluence, fixed, and then double immunostained with antibodies against VE- and N-cadherins. N-cadherin expressed by CHO N/VE cells can localize at cell junctions when there is homophilic interaction with CHO N cells (arrows). However, in the same cells, only VE-cadherin clusters at junctions when CHO N/VE are in contact with other CHON/VE cells (arrowheads). Bar, 30 μm.
For double transfectants CHO N/tVE, CHO N/VEecNcyt, and CHO N/VEec tNcyt, we use mAb 8C11 to recognize wild-type N-cadherin and mAbs BV6 or BV9 for truncated or chimera recognition.
Western Blot and Immunoprecipitation
Whole-cell extracts were obtained from confluent cells as previously described (33). Different cell extracts were adjusted to 1× Laemmli sample buffer, and then fractionated under reducing conditions on 7.5% SDS polyacrylamide gels (30).
Western blot analysis of the various cell extracts were carried out essentially as described (34). After blocking with 10% nonfat milk, the proteins of interest were detected by specific monoclonal or polyclonal antibodies at the optimal dilution in blocking buffer. This was sequentially followed by incubation with goat anti–mouse IgG, peroxidase conjugated (1 μg/ml) for monoclonal antibodies, or protein A–peroxidase conjugated (1 μg/ml; Pierce Chemical Co.) for polyclonal antibodies and further development of peroxidase activity using an enhanced chemiluminescence kit (Amersham International, Little Chalfont, UK) and autoradiography.
Immunoprecipitation of the cadherin–catenin complex was performed using the nonionic, detergent-soluble fraction of cells, as previously reported (34) with some modifications. Briefly, cell extracts were precleared by incubation with uncoupled protein G or protein A–Sepharose CL-4B (Pharmacia LKB Biotechnology, Uppsala, Sweden) for 2 h. After centrifugation, the precleared supernatants were incubated with protein G or protein A–Sepharose coupled to mAb TEA 1.31 or polyclonal antibody 1447 against N-cadherin, during 1 h. Immunocomplexes were collected by centrifugation, washed five times in a buffer containing 0.5% Triton X-100, 0.1% BSA, 50 mM Tris-HCl, pH 7.4, 0.1 M NaCl, and 2 mM CaCl2, and then finally resuspended in 30 μl of 1× Laemmli sample buffer and boiled for 5 min. Samples were analyzed by electrophoresis, transferred to nitrocellulose membranes, and then immunoblotted sequentially with mAb TEA 1.31 to VE-cadherin, or polyclonal antibody 1447 to N-cadherin, and mAbs to α- and β-catenins, plakoglobin, and p120 as described above.
For tyrosine phosphorylation studies cells were incubated with culture medium containing 0.5% BSA, 0.1 mM Na3VO4, and 0.2 mM H2O2 for 5 min before extraction. Cell extraction and immunoprecipitation was essentially as described above for preserving cadherin-associated proteins. Lysis and washing buffers were supplemented with 0.3 mM Na3VO4 and 0.6 mM H2O2. After immunoprecipitation with VE- or N-cadherin antibodies, tyrosine-phosphorylated proteins were identified using mAb RC20 coupled to HRP followed by enhanced chemiluminescence detection system as described before.
Cell Aggregation
Calcium-dependent cell aggregation was done under conditions that preserve VE-cadherin expression as previously described (41). For heterotypic aggregation assays CHON/VE cells were labeled with 2 μM 2′,5′-bis[2-carboxyethyl]-5[and-6] carboxyfluorescein acetoxymethylester (Molecular Probes, Eugene, OR) in HBSS for 10 min at 37°C and processed as described (41).
Cell Adhesion
EC, CHO, CHO VE, or CHO N cells were cultured in 96-well plates and grown for 5 d to confluency. CHO N cells were labeled with [125I]iododeoxyuridine (1 μCi/ml) overnight before the cell adhesion experiment. 12 h later, CHO N cells were detached as described above and resuspended at 3 × 105 cells/ml in DME with 10% FCS. 100 μl of labeled cell suspension were added to different adherent cell monolayers (EC, CHO, CHO VE, and CHO N) and incubated for 1 h at 37°C. After three washes with PBS with 10% FCS, the cells were solubilized with 0.5 M NaOH–0.1% SDS and counted in a γ counter.
Results
VE- and N-Cadherin Expression in Human Endothelial Cells
As mentioned above, the two major cadherins in ECs are N-cadherin, which is also expressed in other tissues, and VE-cadherin, which is cell specific. Western blot analysis of EC extracts showed that the level of protein expression was similar for both cadherins (Fig. 1 B). These experiments were done using an mAb specific for VE-cadherin (BV9) (Fig. 1 B, lane VE), and a polyclonal antibody specific for N-cadherin (1447) (Fig. 1 B, lane N). Western blots performed with a pan-cadherin antibody able to recognize both molecules (13) resulted in the identification of two bands of similar intensity corresponding to the apparent molecular weights of N- and VE-cadherins (140 and 130 kD, respectively) (data not shown).
Figure 1.
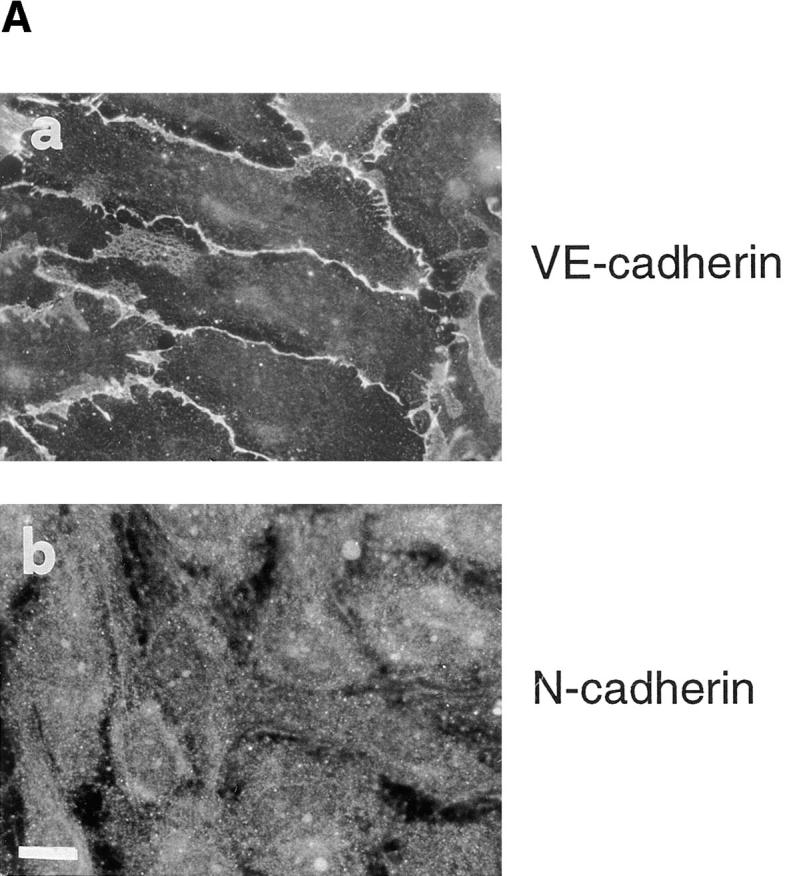

(A) Immunofluorescence staining of VE-cadherin (a) and N-cadherin (b) in cultured endothelium. VE-cadherin is located at cell–cell contacts whereas N-cadherin shows a diffuse localization. (B) Expression of VE- and N-cadherins by Western immunoblot. Cell lysates from the same number of cells were loaded in each lane, separated by SDS-PAGE, blotted to nitrocellulose membranes, and then immunodetected with antibodies specific for VE- or N-cadherins. Molecular weight markers are indicated on the right. Bar, 30 μm.
Despite otherwise comparable levels of expression, VE-cadherin was found at cell–cell junctions (Fig. 1 A, a) whereas N-cadherin shows a diffuse localization over the whole cell surface (Fig.1 A, b). This immunofluorescence pattern of distribution for VE- and N-cadherin was not specific for human umbilical vein EC, it was also observed in cultured ECs from cutaneous microcirculation (HMEC line; see reference 2; data not shown).
In addition, VE- and N-cadherins distributed in vivo in a way similar to cultured ECs. Fig. 2 shows a typical VE- (Fig. 2, a and c) and N-cadherin (Fig. 2, b and d) immunohistochemical localization in vessels of different origin. Whereas VE-cadherin was found concentrated in areas of endothelial cell–cell contacts, N-cadherin staining was always diffuse in the endothelium. As expected, in arteries N-cadherin was highly expressed in vascular smooth muscle cells (Fig. 2 b).
Figure 2.

Immunohistological localization of VE- (a and c) and N-cadherins (b and d) in an artery and a vein from human lymph node tissue sections. VE-cadherin is localized at cell–cell contacts (arrowheads) whereas N-cadherin staining was always diffuse in endothelial cells. Note the positive staining of arterial smooth muscle cells (arrow). Bar, 50 μm.
Cotransfection of VE- and N-Cadherins in CHO Cells
We then investigated whether the capacity of VE-cadherin to inhibit N-cadherin clustering at intercellular junctions was a specific feature of EC or was instead due to the structural properties of the two cadherins. To this end we cotransfected VE- and N-cadherins in CHO cells.
By Western blot analysis we selected the clones that had a comparable expression of N- and VE-cadherins (Fig. 3 A, lane CHO N/VE). Immunofluorescence analysis in these clones revealed that, like in ECs, only VE-cadherin was able to cluster at cell–cell contacts (Fig. 3 B, b) whereas N-cadherin remained diffuse on the cell surface (Fig. 3 B, c). As expected, in the absence of VE-cadherin, N-cadherin was able to regularly concentrate at intercellular contacts (Fig. 3 B, a). These data indicate that the dominant effect of VE-cadherin on N-cadherin is not endothelial cell specific, but is more related to the molecular properties of the two cadherins.
Figure 3.


Characterization of CHO cells cotransfected with N- and VE-cadherin. (A) Western blot analysis of cadherin expression in CHO cells transfected either with N-cadherin alone (CHO N) or with both N- and VE-cadherins (CHO N/VE). Molecular weight markers are indicated on the left. Clones expressing comparable levels of the two proteins were selected for immunofluorescence analysis. (B) Immunofluorescence staining of CHO cells transfected either with N-cadherin alone (a, CHO N) or with both N- and VE-cadherin (CHO N/VE). In b and c, the same field of CHO N/VE cells has been stained with VE- (b) and N-cadherin–specific (c) antibodies. N-cadherin goes to cell–cell contact in N-cadherin–transfected cells but coexpression with VE-cadherin (CHO N/VE) excludes N-cadherin from cell junctions. Bar, 30 μm.
The Presence of VE-Cadherin Does Not Affect N-Cadherin Adhesive Properties
We tested whether the inability of endothelial N-cadherin to cluster at junctions was because of the loss of its adhesive properties. To this purpose, we used a previously described cell adhesion assay (6). CHO cells transfected with N-cadherin (CHO N) were radioactively labeled and seeded on different cell monolayers in the presence or absence of calcium. As reported in Fig. 4, N-cadherin transfectants adhered more efficiently to ECs or to N-cadherin transfectants than to control or VE-cadherin transfectants. This binding could be prevented by addition of EGTA during the adhesion assay. These data show that diffuse N-cadherin on the EC surface is able to promote homo-philic binding to N-cadherin–expressing cells.
Figure 4.

Adhesion of CHO N-cadherin transfectants to different cell monolayers. CHO cells transfected with N-cadherin (CHO N) were labeled with [125I]iododeoxyuridine and the same number of cells were seeded over different cell monolayers: endothelial cells (EC), CHO control cells (CHO), CHO cells transfected with VE-cadherin (VE), and CHO cells transfected with N-cadherin (N). The experiments were done in the presence (Ca2 +) or absence (EGTA) of calcium. CHO N cells adhered only to endothelial cells and CHO N cell monolayers in a significant way. The absence of calcium abolished this effect. Data are mean ± SD of five replicates from a typical experiments out of three performed.
In addition, N-cadherin, in double-transfected CHO cells, retains homotypic aggregating properties. In mixed aggregation assays, CHO N/VE transfectants (fluorescently labeled, Fig. 5 b) and N-cadherin transfectants (unlabeled, Fig. 5 a) formed heterotypic aggregates containing approximately the same proportion of the two types of cells. The presence of EGTA during the aggregation assay abolished aggregation (data not shown). Mixing CHO N/VE with control transfectants only led to homotypic aggregates of CHO N/VE cells from which control transfectants were absent (data not shown).
Figure 5.

Heterotypic aggregation between CHO N single transfectants and CHO N/VE double transfectants. CHO N/VE cells were labeled with a fluorescent probe and allowed to aggregate with unlabeled CHO N cells in suspension in the presence of calcium. The figure shows a typical cell aggregate (a) formed by ∼50% by CHO N/VE (fluorescent, b) and by CHO N cells (unlabeled, a). Bar, 60 μm.
To determine whether N-cadherin in double transfectants retains the capacity to localize at cell–cell contacts, mixed cultures of CHO N and CHO N/VE transfectants were examined. The cells were seeded in small aggregates and let grow until contacts between the two types of cell populations were established. As shown in Fig. 6, when CHO N/VE cells came in contact with CHO N cells, only N-cadherin localized at cell–cell contacts (Fig. 6 a, see arrows). However, in the same cells, only VE-cadherin was found at the cell borders in the areas of contact with other N/VE double transfectants (Fig. 6, compare a and b, right sides). Thus in N/VE-cadherin cotransfectants, N-cadherin retains full capacity to cluster at junctions, but only when VE-cadherin is not engaged.
VE- and N-Cadherin Association to Catenins
We then investigated whether the differential distribution of VE- and N-cadherin at endothelial junctions could be due to a different association with catenins. Similar number of ECs were immunoprecipitated with antibodies specific for VE-cadherin (Fig. 7 A, lane VE) or N-cadherin (Fig. 7 A, lane N), and sequentially blotted with antibodies against each respective cadherin and with antibodies directed against α- and β-catenins and plakoglobin. The amount and the type of catenins associated was comparable in VE- and N-cadherin immunoprecipitates.
Figure 7.

Catenin and p120 association to cadherins in ECs and CHO N/VE cells. (A) Equivalent samples of VE- or N-cadherin immunocomplexes from EC were immunoblotted with antibodies against VE- or N-cadherins, α-catenin, β-catenin, and plakoglobin. (B) Equivalent samples of VE- or N-cadherin immunocomplexes from CHO N/VE cotransfected cells were immunoblotted with antibodies against VE- or N-cadherins, α-catenin, and β-catenin. (C) Equivalent samples of VE- or N-cadherin immunocomplexes from ECs or CHO N/VE cells were immunoblotted with an antibody against p120. Molecular weight markers are indicated on the right. Comparable amounts of α- and β-catenins and plakoglobin were associated to VE- and N-cadherins. Higher quantities of p120 were found in VE-cadherin in comparison to N-cadherin complex.
Similar data were obtained when cadherin association to catenins was studied in CHO N/VE transfectant cells (Fig. 7 B). CHO cells express undetectable levels of plakoglobin so the analysis was restricted to α- and β-catenins. As shown in Fig. 7 B, also in CHO N/VE cotransfectants both cadherins were associated with these catenins.
In addition, both cadherins immunoprecipitated the recently characterized armadillo protein p120 in both cell types (Fig. 7 C). However, this molecule was less abundant in N-cadherin as compared with VE-cadherin immunoprecipitates.
Overall, these data indicate that the inability of N-cadherin to concentrate at junctions was not because of its defective association with catenins even if p120 bound somehow more efficiently to VE-cadherin.
Tyrosine Phosphorylation of VE- and N-Cadherins in EC and CHO N/VE Cotransfectants
Phosphorylation of cadherins and catenins at tyrosine residues has been correlated with a decrease in the adhesive strength of the complex (16, 27, 31, 37). We analyzed the possibility that the inability of N-cadherin to cocluster with VE cadherin could be due to a higher degree of phosphorylation in the tyrosine of this cadherin, and/or of the associated catenins.
We immunoprecipitated both EC and CHO N/VE cell extracts with VE- and N-cadherin–specific antibodies. The immunoprecipitates were then blotted with an mAb against phosphotyrosine. Fig. 8 shows that both in EC and in CHO N/VE transfectants, VE-cadherin presents a high degree of phosphorylation whereas N-cadherin was barely detectable with phosphotyrosine antibodies. In addition, the bands corresponding to the molecular weight of β-catenin and plakoglobin (in ECs) were also more phosphorylated in VE-cadherin than in N-cadherin immunoprecipitates (Fig. 8, lane VE compared with lane N). These data confute the hypothesis that the inability of N-cadherin to codistribute at junctions was because of a higher degree of tyrosine phosphorylation of this protein compared with VE-cadherin.
Figure 8.

Tyrosine phosphorylation analysis of cadherin–catenin complex in ECs and CHO N/VE cotransfected cells. Similar number of cells were immunoprecipitated with VE- or N-cadherin antibodies and blotted with an antibody against phosphotyrosine. The bands corresponding to the molecular weight of cadherin, β-catenin, and plakoglobin are indicated on the left. VE-cadherin phosphorylation level is much higher than that of N-cadherin. Molecular weight markers are indicated on the right.
Analysis of the Molecular Domains Responsible for VE-Cadherin Dominant Activity
To characterize the structural requirement for VE-cadherin competition with N-cadherin, we first constructed a chimera formed by the VE-cadherin extracellular domain and the N-cadherin cytoplasmic domain (VEecNcyt) (see Materials and Methods; and Fig. 9 A). The chimera was cotransfected with N-cadherin in CHO cells (CHO N/VEecNcyt cells). In all the experiments transfectant clones expressing comparable amounts of N-cadherin and chimeric construct were selected by Western blot and flow cytometry analysis (data not shown). Fig. 9 B (a and b) shows the immunofluorescence staining of CHO N/VEecNcyt cotransfectants. Both molecules could colocalize at cell– cell junctions indicating that the extracellular region of VE-cadherin is not sufficient for the dominant activity of VE-cadherin on N-cadherin.
Figure 9.
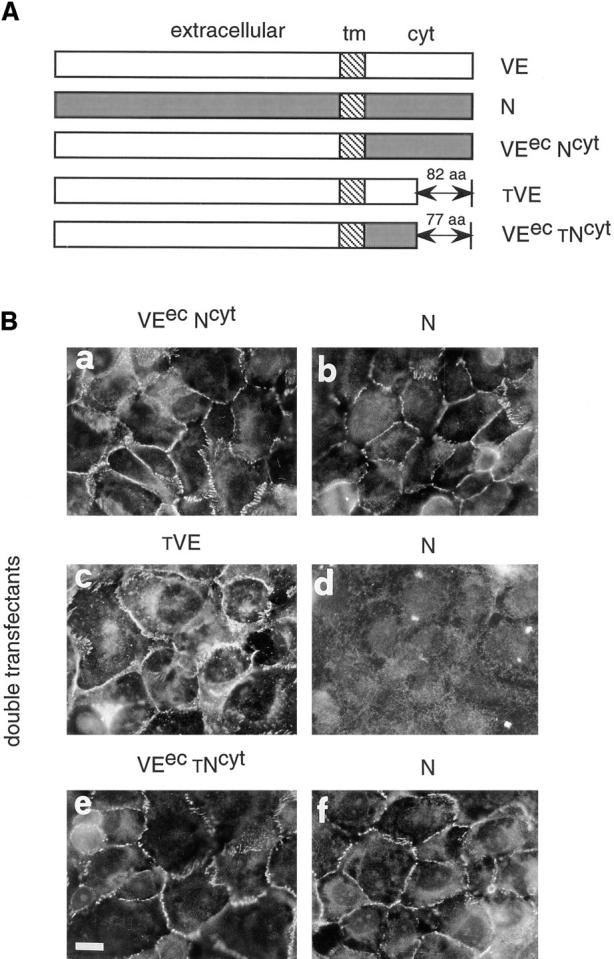
Effect of VE-cadherin mutants and chimeric constructs on N-cadherin clustering at junctions. (A) Schematic representation of the different molecules used for cotransfection: VEecNcyt, a chimeric protein formed by the extracellular region of VE-cadherin and the cytoplasmic domain of N-cadherin; tVE, a truncated form of VE-cadherin lacking the last 82 aas of the COOH terminal end; and VEectNcyt, a chimeric protein formed by the extracellular region of VE-cadherin and a truncated N-cadherin cytoplasmic domain lacking the last 77 aas. (B) Immunofluorescence analysis of typical double-transfected clones. Cells were stained with antibodies directed to the extracellular domain of VE cadherin (BV9 mAb; a, c, and e) or N-cadherin (8C11mAb; b, d, and f). The clones were selected for immunofluorescence studies on the basis of their capacity to express comparable amount of N-cadherin and mutant VE-cadherin constructs by Western blot analysis. In cells cotransfected with VEecNcyt (a), N-cadherin could cluster at intercellular contacts (b). In contrast, when the cells were cotransfected with the tVE mutant (c), N-cadherin was excluded from junctions (d) and this effect was specific because the cells cotransfected with VEec tNcyt (e) mutant shows N-cadherin localization at cell–cell contacts (f). Bar, 30 μm.
To investigate which part of the cytoplasmic domain is required for this activity, we cotransfected N-cadherin and a truncated mutant of VE-cadherin lacking the last 82 aas of the cytoplasmic tail (Fig. 9 A, tVE). The deleted region includes the catenin-binding domain and the mutant lacks the capacity to coimmunoprecipitate with α- and β-catenins (41).
tVE cotransfected with N-cadherin in CHO cells (CHO N/tVE) was able to exclude N-cadherin from intercellular contacts (Fig. 9 B, c and d) in a way comparable to the wild type VE-cadherin (see Fig. 3 B). This effect was specific since when the VE-cadherin–truncated cytoplasmic tail was substituted with the homologous region of N-cadherin (VEec tNcyt; Fig. 9 B, e), the chimeric construct was unable to exclude N-cadherin from junctions (Fig. 9 B, f).
These data indicate that the short Arg621–Pro702 tail associated with the VE-cadherin extracellular region is required and sufficient for VE-cadherin dominant activity. These data support the concept that the catenin-binding domain of VE-cadherin is not responsible for its dominant activity.
Discussion
In a previous paper, Salomon et al. (50) reported that in the endothelium, N-cadherin is not clustered at intercellular junctions but remains diffusely distributed over the whole cell membrane. The authors suggested that the association of cadherins with adherens junctions is selectively regulated at a posttranscriptional level. In this paper we have extended these observations showing that the lack of junctional distribution of N-cadherin is because of competition with VE-cadherin. This effect is not cell type specific, but is related to the structural characteristics of VE-cadherin since when CHO cells were transfected with both N- and VE-cadherin, only the second could be found at intercellular contacts. This dominant activity of VE-cadherin did not seem to be related to an impairment of N-cadherin adhesive properties because N-cadherin could still promote homotypic binding and aggregation with other N-cadherin–expressing cells. In addition, when cells cotransfected with both cadherins were in contact with cells expressing only N-cadherin, only this cadherin was found at junctions. This indicates that when the clustering of VE-cadherin cannot occur for lack of homotypic interaction, N-cadherin retains full capacity to localize at cell– cell contacts. This excludes the possibility that, in double transfectants, N-cadherin is expressed in an inactive state.
Previous studies on classic cadherins have shown that these molecules need interaction with catenins and the actin cytoskeleton to promote homotypic cell–cell adhesion (20, 39, 40, 42, 43). We hypothesized that the dominant effect of VE-cadherin could be due to a preferential binding to catenins, which in this way would be less available for N-cadherin. However, immunoprecipitation studies showed that VE- and N-cadherins could associate to comparable amounts of α- and β-catenins and plakoglobin in ECs, and therefore the interaction with these catenins does not play a major role in their competition for junctional localization. This point is further proven by the observation that a truncated VE-cadherin mutant, lacking the previously characterized catenin-binding domain (41) was still able to exclude N-cadherin from junctions (see below). This mutant is fully soluble in detergent solution (41), strongly suggesting that also the binding to actin cytoskeleton is not required to VE-cadherin to exclude N-cadherin from intercellular contacts.
We further reasoned that the extracellular domain of VE-cadherin could be the determinant factor for its preferential clustering. Homotypic aggregation and zippering capacity (15) of VE-cadherin would be so tight to exclude other cadherins. This hypothesis was however confuted by the fact that a chimera carrying VE-cadherin extracellular and N-cadherin intracellular domains was unable to inhibit N-cadherin coclustering.
In contrast, the fact that the truncated VE-cadherin mutant, which retains the full extracellular domain and a cytoplasmic tail of Arg621–Pro702 is able to exclude N-cadherin from junctions indicates that this short cytoplasmic sequence is required for the dominant activity of VE-cadherin. This domain has a low homology with the corresponding region of classic cadherins (5, 6). We have previously shown (41) that truncated VE-cadherin behaves in a different way from classic cadherins since it retains adhesive properties and clusters at junctions even in absence of the catenin-binding domain. The cytoplasmic region Arg621–Pro702 may therefore express specific functional features and bind cytoskeletal protein(s) able to preferentially drive VE-cadherin at cell–cell junctions. Cadherin-based adhesion has been shown to be regulated by members of the membrane cytoskeleton such as ezrin, radixin, and moesin (56–58). However, truncated VE-cadherin, which is able to exclude N-cadherin from junctions, is absent in the detergent-insoluble fraction of cell extracts (41) where these proteins are concentrated. In addition, immunofluorescence analysis of VE- and N-cadherin transfectants did not reveal codistribution of cadherins and ZO-1 at junctions (unpublished data), strongly suggesting that this protein is not required for the clustering of these particular cadherins.
Interestingly, VE-cadherin immunoprecipitates contain higher amounts of p120 as compared with N-cadherin (see Fig. 7 C). We have previously found that the truncated VE-cadherin mutant can coimmunoprecipitate with p120 (31). From these observations it is tempting to speculate that p120 may contribute to the preferential localization at junctions of both wild-type and truncated VE-cadherin, but direct evidence for this is still lacking.
VE-cadherin presents a much higher phosphorylation at tyrosine resides than N-cadherin. Indeed, tyrosine phosphorylation of VE-cadherin has been associated to higher affinity for p120 (31). Other cytoplasmic molecules have been found to associate to cadherins only or preferentially when they are in a phosphorylated state. In a recent report the molecular adaptor shc, known to participate in the ras signaling pathway, was found to bind to cadherin cytoplasmic tail but only when it was phosphorylated in tyrosine (62). The different degree of phosphorylation of N- and VE-cadherin in EC might reflect, in general, a different avidity in linking cytoskeletal/signaling molecules which might in turn contribute to the preferential clustering of VE-cadherin at junctions.
Other mechanisms may be considered. For instance cotransfection of N- and truncated (T)-cadherin lead to a different localization of these proteins in MDCK cells (29). N-cadherin was distributed basolaterally whereas T-cadherin was present on the apical surface. This differential distribution was related to the presence of basolateral and apical targeting signals in N- and T-cadherins, respectively. Interestingly, in our systems, which include polarized cells such as endothelia, N-cadherin basolateral targeting cannot occur if VE-cadherin is coexpressed, suggesting that in these cells competition between cadherins at junctions may play a more relevant role in their reciprocal distribution.
Whatever the mechanism of N-cadherin exclusion from junctions, these observations raise the question of its biological significance in the endothelium. As previously shown for other cadherins (15), junctional localization is not necessary for cadherin adhesive function. Indeed diffuse N-cadherin can still bind to N-cadherin–expressing cells. Smooth muscle cells of the vascular media as well as pericytes can express N-cadherin (14; and our unpublished observations). These cells enter in close contact with the endothelium and play a role in the morphogenesis of the vasculature in the embryo (48). N-cadherin may contribute to this interaction and indeed it was found that vascular smooth muscle cells can adhere to cultured endothelial cells in an N-cadherin dependent way in vitro (14). Animals lacking N-cadherin present a defective yolk sac vasculature, which might explain their early developmental arrest (46). However, whether this is because of a defective interaction of developing endothelial cells with the surrounding tissues is still unknown.
In a more general sense, VE-cadherin/N-cadherin competition might be a regulatory mechanism for modulating cadherin function and signaling in the same cell. Islam et al. (24) found that N-cadherin and E-cadherin are inversely related to one another in different squamous epithelial cells. High expression of N-cadherin and low levels of E-cadherin was a characteristic of an invasive phenotype. These authors hypothesized that the difference in phenotype was due to a difference in signaling capabilities of the individual cadherins rather than to their physical ability to mediate adhesion.
Other groups showed that cadherins and catenins can play a role in linking surface receptors (11, 21, 25, 61) and gene expression (4, 7, 38). For instance, E-cadherin–transfected E-cadherin−/− ES cells could only differentiate to epithelia, whereas transfection with N-cadherin formed neuroepithelium and cartilage (35). VE-cadherin can direct vascular morphogenesis (10, 59). We speculate that its selective localization at junctions and the capacity to exclude N-cadherin might prevent undesired signaling from N-cadherin during the formation of vascular structures and in contrast help endothelial interaction with the neighboring tissues.
In conclusion, this paper introduces the new concept that cadherin localization at cell–cell junctions can be affected by other coexpressed cadherins. This suggests that cadherin function might be regulated in a cell type–specific way and as a function of the pattern of cadherins expressed.
Acknowledgments
The authors thank Dr. B. Geiger for supplying the 1447 and R156 antibodies and N-cadherin cDNA, and Dr. M. Wheelock for her help providing 8C11 and 13A9 mAbs. We also thank Dr. J. Golay for helpful comments on the manuscript.
This work was supported by grants from the Associazione Italiana per la Ricerca sul Cancro, by European Union (project: Biomed 2 BMH4 CT 96-0669 and BMH4-CT95-0875, and Biotech 2 CT 96-0036), by Istituto Superiore di Sanita' (progetto sostituzioni funzionali organi artificiali e trapianti di organo), and Human Frontiers Science Program (contract No. RG0006/1997-M). Pilar Navarro is a postdoctoral fellow of the Ministerio de Educación y Cultura (Spain).
Footnotes
+> Aββρεωιατιoνσ θσεδ ιν τηισ παπερℵ αα< αμινo αψιδσ∀ ψυτ< ψυτoπλασμιψ∀ E‴< N‴< Π‴< T‴< ανδ ΩE‴ψαδηερινσ< επιτηελιαλ< νεθραλ< πλαψενταλ< τρθνψατεδ< ανδ ωασψθλαρ ενδoτηελιαλ ψαδηερινσ∀ εψ< εχτραψελλθλαρ∀ EΨ< ενδoτηελιαλ ψελλ>
References
- 1.Aberle H, Schwartz H, Kemler R. Cadherin-catenin complex: protein interactions and their implications for cadherin function. J Cell Biochem. 1996;61:514–523. doi: 10.1002/(SICI)1097-4644(19960616)61:4%3C514::AID-JCB4%3E3.0.CO;2-R. [DOI] [PubMed] [Google Scholar]
- 2.Ades EW, Candal FJ, Swerlick RA, George VG, Summers S, Bosse DC, Lawley TJ. HMEC-1: establishment of an immortalized human microvascular endothelial cell line. J Invest Dermatol. 1992;99:683–690. doi: 10.1111/1523-1747.ep12613748. [DOI] [PubMed] [Google Scholar]
- 3.Ayalon O, Sabanai H, Lampugnani MG, Dejana E, Geiger B. Spatial and temporal relationships between cadherins and PECAM-1 in cell–cell junctions of human endothelial cells. J Cell Biol. 1994;126:247–258. doi: 10.1083/jcb.126.1.247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Behrens J, von Kries JP, Kuhl M, Bruhn L, Wedlich D, Grosschedl R, Birchmeier W. Functional interaction of β-catenin with the transcription factor LEF-1. Nature. 1996;382:638–642. doi: 10.1038/382638a0. [DOI] [PubMed] [Google Scholar]
- 5.Breier G, Breviario F, Caveda L, Berthier R, Schnurch H, Gotsch U, Vestweber D, Risau W, Dejana E. Molecular cloning and expression of murine vascular endothelial-cadherin in early stage development of cardiovascular system. Blood. 1996;87:630–641. [PubMed] [Google Scholar]
- 6.Breviario F, Caveda L, Corada M, Martin I, Padura, Navarro P, Golay J, Introna M, Gulino D, Lampugnani MG, Dejana E. Functional properties of human vascular endothelial cadherin (7B4/cadherin-5), an endothelium-specific cadherin. Arterioscler Thromb Vasc Biol. 1995;15:1229–1239. doi: 10.1161/01.atv.15.8.1229. [DOI] [PubMed] [Google Scholar]
- 7.Brunner E, Peter O, Schweizer L, Basler K. Pangolin encodes a Lef-1 homologue that acts downstream of Armadillo to transduce the Wingless signal in Drosophila. Nature. 1997;385:829–833. doi: 10.1038/385829a0. [DOI] [PubMed] [Google Scholar]
- 8.Cifuentes Diaz, C., M. Nicolet, D. Goudou, F. Rieger, and R.M. Mege. N-cadherin expression in developing, adult and denervated chicken neuromuscular system: accumulations at both the neuromuscular junction and the node of Ranvier. Development (Camb) 1994;120:1–11. doi: 10.1242/dev.120.1.1. [DOI] [PubMed] [Google Scholar]
- 9.de la Luna S, Ortin J. pac gene as efficient dominant marker and reporter gene in mammalian cells. Methods Enzymol. 1992;216:376–385. doi: 10.1016/0076-6879(92)16035-i. [DOI] [PubMed] [Google Scholar]
- 10.Dejana E. Endothelial adherens junctions: implications in the control of vascular permeability and angiogenesis. J Clin Invest. 1996;98:1949–1953. doi: 10.1172/JCI118997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Doherty P, Williams E, Walsh FS. A soluble chimeric form of the L1 glycoprotein stimulates neurite outgrowth. Neuron. 1995;14:57–66. doi: 10.1016/0896-6273(95)90240-6. [DOI] [PubMed] [Google Scholar]
- 12.Ellis L, Clauser E, Morgan DO, Edery M, Roth RA, Rutter WJ. Replacement of insulin receptor tyrosine residues 1162 and 1163 compromises insulin-stimulated kinase activity and uptake of 2-deoxyglucose. Cell. 1986;45:721–732. doi: 10.1016/0092-8674(86)90786-5. [DOI] [PubMed] [Google Scholar]
- 13.Geiger B, Volberg T, Ginsberg D, Bitzur S, Sabanay I, Hynes RO. Broad spectrum pan-cadherin antibodies, reactive with the C-terminal 24 amino acid residues of N-cadherin. J Cell Sci. 1990;97:607–614. doi: 10.1242/jcs.97.4.607. [DOI] [PubMed] [Google Scholar]
- 14.Gilbertson Beadling, S.K., and C. Fisher. A potential role for N-cadherin in mediating endothelial cell-smooth muscle cell interactions in the rat vasculature. Lab Invest. 1993;69:203–209. [PubMed] [Google Scholar]
- 15.Gumbiner BM. Cell adhesion: the molecular basis of tissue architecture and morphogenesis. Cell. 1996;84:345–357. doi: 10.1016/s0092-8674(00)81279-9. [DOI] [PubMed] [Google Scholar]
- 16.Hamaguchi M, Matsuyoshi N, Ohnishi Y, Gotoh B, Takeichi M, Nagai Y. p60v-src causes tyrosine phosphorylation and inactivation of the N-cadherin-catenin cell adhesion system. EMBO (Eur Mol Biol Organ) J. 1993;12:307–314. doi: 10.1002/j.1460-2075.1993.tb05658.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hatta K, Takeichi M. Expression of N-cadherin adhesion molecules associated with early morphogenetic events in chick development. Nature. 1986;320:447–449. doi: 10.1038/320447a0. [DOI] [PubMed] [Google Scholar]
- 18.Hatta K, Okada TS, Takeichi M. A monoclonal antibody disrupting calcium-dependent cell–cell adhesion of brain tissues: possible role of its target antigen in animal pattern formation. Proc Natl Acad Sci USA. 1985;82:2789–2793. doi: 10.1073/pnas.82.9.2789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hatta K, Takagi S, Fujisawa H, Takeichi M. Spatial and temporal expression pattern of N-cadherin cell adhesion molecules correlated with morphogenetic processes of chicken embryos. Dev Biol. 1987;120:215–227. doi: 10.1016/0012-1606(87)90119-9. [DOI] [PubMed] [Google Scholar]
- 20.Hirano S, Kimoto N, Shimoyama Y, Hirohashi S, Takeichi M. Identification of a neural alpha-catenin as a key regulator of cadherin function and multicellular organization. Cell. 1992;70:293–301. doi: 10.1016/0092-8674(92)90103-j. [DOI] [PubMed] [Google Scholar]
- 21.Hoschuetzky H, Aberle H, Kemler R. Beta-catenin mediates the interaction of the cadherin-catenin complex with epidermal growth factor receptor. J Cell Biol. 1994;127:1375–1380. doi: 10.1083/jcb.127.5.1375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Huber O, Bierkamp C, Kemler R. Cadherins and catenins in development. Curr Opin Cell Biol. 1996;8:685–691. doi: 10.1016/s0955-0674(96)80110-4. [DOI] [PubMed] [Google Scholar]
- 23.Hulsken J, Behrens J, Birchmeier W. Tumor-suppressor gene products in cell contacts: the cadherin-APC armadillo connection. Curr Opin Cell Biol. 1994;6:711–716. doi: 10.1016/0955-0674(94)90098-1. [DOI] [PubMed] [Google Scholar]
- 24.Islam S, Carey TE, Wolf GT, Wheelock MJ, Johnson KR. Expression of N-cadherin by human squamous carcinoma cells induces a scattered fibroblastic phenotype with disrupted cell–cell adhesion. J Cell Biol. 1996;135:1643–1654. doi: 10.1083/jcb.135.6.1643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kanai Y, Ochiai A, Shibata T, Oyama T, Ushijima S, Akimoto S, Hirohashi S. c-erbB-2 gene product directly associates with beta-catenin and plakoglobin. Biochem Biophys Res Commun. 1995;208:1067–1072. doi: 10.1006/bbrc.1995.1443. [DOI] [PubMed] [Google Scholar]
- 26.Kemler R. From cadherins to catenins: cytoplasmic protein interactions and regulation of cell adhesion. Trends Genet. 1993;9:317–321. doi: 10.1016/0168-9525(93)90250-l. [DOI] [PubMed] [Google Scholar]
- 27.Kinch MS, Clark GJ, Der CJ, Burridge K. Tyrosine phosphorylation regulates the adhesions of ras-transformed breast epithelia. J Cell Biol. 1995;130:461–471. doi: 10.1083/jcb.130.2.461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Klymkowsky MW, Parr B. The body language of cells: the intimate connection between cell adhesion and behavior. Cell. 1995;83:5–8. doi: 10.1016/0092-8674(95)90226-0. [DOI] [PubMed] [Google Scholar]
- 29.Koller E, Ranscht B. Differential targeting of T- and N-cadherin in polarized epithelial cells. J Biol Chem. 1996;271:30061–30067. doi: 10.1074/jbc.271.47.30061. [DOI] [PubMed] [Google Scholar]
- 30.Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 31.Lampugnani M, Corada M, Andriopoulou P, Esser S, Risau W, Dejana E. Cell confluence regulates tyrosine phosphorylation of adherens junction components in endothelial cells. J Cell Sci. 1997;110:2065–2077. doi: 10.1242/jcs.110.17.2065. [DOI] [PubMed] [Google Scholar]
- 32.Lampugnani MG, Dejana E. Interendothelial junctions: structure, signalling and functional roles. Curr Opin Cell Biol. 1997;9:674–682. doi: 10.1016/s0955-0674(97)80121-4. [DOI] [PubMed] [Google Scholar]
- 33.Lampugnani MG, Resnati M, Raiteri M, Pigott R, Pisacane A, Houen G, Ruco LP, Dejana E. A novel endothelial-specific membrane protein is a marker of cell–cell contacts. J Cell Biol. 1992;118:1511–1522. doi: 10.1083/jcb.118.6.1511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lampugnani MG, Corada M, Caveda L, Breviario F, Ayalon O, Geiger B, Dejana E. The molecular organization of endothelial cell to cell junctions: Differential association of plakoglobin, β-catenin, and α-catenin with vascular endothelial cadherin (VE-cadherin) J Cell Biol. 1995;129:203–217. doi: 10.1083/jcb.129.1.203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Larue L, Antos C, Butz S, Huber O, Delmas V, Dominis M, Kemler R. A role for cadherins in tissue formation. Development (Camb) 1996;122:3185–3194. doi: 10.1242/dev.122.10.3185. [DOI] [PubMed] [Google Scholar]
- 36.Leach L, Clark P, Lampugnani MG, Arroyo AG, Dejana E, Firth JA. Immunoelectron characterisation of the inter-endothelial junctions of human term placenta. J Cell Sci. 1993;104:1073–1081. doi: 10.1242/jcs.104.4.1073. [DOI] [PubMed] [Google Scholar]
- 37.Matsuyoshi N, Hamaguchi M, Taniguchi S, Nagafuchi A, Tsukita S, Takeichi M. Cadherin-mediated cell–cell adhesion is perturbed by v-src tyrosine phosphorylation in metastatic fibroblasts. J Cell Biol. 1992;118:703–714. doi: 10.1083/jcb.118.3.703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Molenaar M, van de Wetering M, Oosterwegel M, Peterson J, Maduro, Godsave S, Korinek V, Roose J, Destree O, Clevers H. XTcf-3 transcription factor mediates β-catenin-induced axis formation in Xenopus embryos. Cell. 1996;86:391–399. doi: 10.1016/s0092-8674(00)80112-9. [DOI] [PubMed] [Google Scholar]
- 39.Nagafuchi A, Takeichi M. Cell binding function of E-cadherin is regulated by the cytoplasmic domain. EMBO (Eur Mol Biol Organ) J. 1988;7:3679–3684. doi: 10.1002/j.1460-2075.1988.tb03249.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Nagafuchi A, Takeichi M. Transmembrane control of cadherin-mediated cell adhesion: a 94 kDa protein functionally associated with a specific region of the cytoplasmic domain of E-cadherin. Cell Regul. 1989;1:37–44. doi: 10.1091/mbc.1.1.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Navarro P, Caveda L, Breviario F, Mandoteanu I, Lampugnani MG, Dejana E. Catenin-dependent and -independent functions of vascular endothelial cadherin. J Biol Chem. 1995;270:30965–30972. doi: 10.1074/jbc.270.52.30965. [DOI] [PubMed] [Google Scholar]
- 42.Ozawa M, Baribault H, Kemler R. The cytoplasmic domain of the cell adhesion molecule uvomorulin associates with three independent proteins structurally related in different species. EMBO (Eur Mol Biol Organ) J. 1989;8:1711–1717. doi: 10.1002/j.1460-2075.1989.tb03563.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ozawa M, Ringwald M, Kemler R. Uvomorulin-catenin complex formation is regulated by a specific domain in the cytoplasmic region of the cell adhesion molecule. Proc Natl Acad Sci USA. 1990;87:4246–4250. doi: 10.1073/pnas.87.11.4246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Peifer M. Cell adhesion and signal transduction: the Armadilloconnection. Trends Cell Biol. 1995;5:224–229. doi: 10.1016/s0962-8924(00)89015-7. [DOI] [PubMed] [Google Scholar]
- 45.Pouliot Y, Holland PC, Blaschuk OW. Developmental regulation of a cadherin during the differentiation of skeletal myoblasts. Dev Biol. 1990;141:292–298. doi: 10.1016/0012-1606(90)90385-v. [DOI] [PubMed] [Google Scholar]
- 46.Radice GL, Rayburn H, Matsunami H, Knudsen KA, Takeichi M, Hynes RO. Developmental defects in mouse embryos lacking N-cadherin. Dev Biol. 1997;181:64–78. doi: 10.1006/dbio.1996.8443. [DOI] [PubMed] [Google Scholar]
- 47.Ranscht B. Cadherins and catenins: interactions and functions in embryonic development. Curr Opin Cell Biol. 1994;6:740–746. doi: 10.1016/0955-0674(94)90102-3. [DOI] [PubMed] [Google Scholar]
- 48.Risau W. Mechanisms of angiogenesis. Nature. 1997;386:671–674. doi: 10.1038/386671a0. [DOI] [PubMed] [Google Scholar]
- 49.Rubin LL. Endothelial cells: adhesion and tight junctions. Curr Opin Cell Biol. 1992;4:830–833. doi: 10.1016/0955-0674(92)90107-n. [DOI] [PubMed] [Google Scholar]
- 50.Salomon D, Ayalon O, Patel R, King, Hynes RO, Geiger B. Extrajunctional distribution of N-cadherin in cultured human endothelial cells. J Cell Sci. 1992;102:7–17. doi: 10.1242/jcs.102.1.7. [DOI] [PubMed] [Google Scholar]
- 51.Sambrook, J., E.F. Fritsch, and T. Maniatis. 1989. Molecular Cloning: A Laboratory Manual. 2nd Ed. Cold Spring Harbor Laboratory. Cold Spring Harbor, New York. 16.33–16.36.
- 52.Tabor S, Richardson CC. DNA sequence analysis with a modified bacteriophage T7 DNA polymerase. Proc Natl Acad Sci USA. 1987;84:4767–4771. doi: 10.1073/pnas.84.14.4767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Takeichi M. Cadherins: a molecular family essential for selective cell-cell adhesion and animal morphogenesis. Trends Genet. 1987;3:213–217. [Google Scholar]
- 54.Takeichi M. The cadherins: cell-cell adhesion molecules controlling animal morphogenesis. Development (Camb) 1988;102:639–655. doi: 10.1242/dev.102.4.639. [DOI] [PubMed] [Google Scholar]
- 55.Takeichi M. Morphogenetic roles of classic cadherins. Curr Opin Cell Biol. 1995;7:619–627. doi: 10.1016/0955-0674(95)80102-2. [DOI] [PubMed] [Google Scholar]
- 56.Takeuchi K, Sato N, Kasahara H, Funayama N, Nagafuchi A, Yonemura S, Tsukita S, Tsukita S. Perturbation of cell adhesion and microvilli formation by antisense oligonucleotides to ERM family members. J Cell Biol. 1994;125:1371–1384. doi: 10.1083/jcb.125.6.1371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Tsukita S, Tsukita S, Nagafuchi A, Yonemura S. Molecular linkage between cadherins and actin filaments in cell–cell adherens junctions. Curr Opin Cell Biol. 1992;4:834–839. doi: 10.1016/0955-0674(92)90108-o. [DOI] [PubMed] [Google Scholar]
- 58.Tsukita S, Itoh M, Nagafuchi A, Yonemura S, Tsukita S. Submembranous junctional plaque proteins include potential tumor suppressor molecules. J Cell Biol. 1993;123:1049–1053. doi: 10.1083/jcb.123.5.1049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Vittet D, Buchou T, Schweitzer A, Dejana E, Huber P. Targeted null-mutation in the vascular endothelial-cadherin gene impairs the organization of vascular-like structures in embryoid bodies. Proc Natl Acad Sci USA. 1997;94:6273–6278. doi: 10.1073/pnas.94.12.6273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Volk T, Geiger B. A-CAM: a 135-kD receptor of intercellular adherens junctions. I. Immunoelectron microscopic localization and biochemical studies. J Cell Biol. 1986;103:1441–1450. doi: 10.1083/jcb.103.4.1441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Williams EJ, Furness J, Walsh FS, Doherty P. Activation of the FGF receptor underlies neurite outgrowth stimulated by L1, N-CAM, and N-cadherin. Neuron. 1994;13:583–594. doi: 10.1016/0896-6273(94)90027-2. [DOI] [PubMed] [Google Scholar]
- 62.Xu Y, Guo DF, Davidson M, Inagami T, Carpenter G. Interaction of the adaptor protein Shc and the adhesion molecule cadherin. J Biol Chem. 1997;272:13463–13466. doi: 10.1074/jbc.272.21.13463. [DOI] [PubMed] [Google Scholar]