

Abstract
Sorting of regulated secretory proteins in the TGN to immature secretory granules (ISG) is thought to involve at least two steps: their selective aggregation and their interaction with membrane components destined to ISG. Here, we have investigated the sorting of chromogranin B (CgB), a member of the granin family present in the secretory granules of many endocrine cells and neurons. Specifically, we have studied the role of a candidate structural motif implicated in the sorting of CgB, the highly conserved NH2-terminal disulfide– bonded loop. Sorting to ISG of full-length human CgB and a deletion mutant of human CgB (Δcys-hCgB) lacking the 22–amino acid residues comprising the disulfide-bonded loop was compared in the rat neuroendocrine cell line PC12. Upon transfection, i.e., with ongoing synthesis of endogenous granins, the sorting of the deletion mutant was only slightly impaired compared to full-length CgB. To investigate whether this sorting was due to coaggregation of the deletion mutant with endogenous granins, we expressed human CgB using recombinant vaccinia viruses, under conditions in which the synthesis of endogenous granins in the infected PC12 cells was shut off. In these conditions, Δcys-hCgB, in contrast to full-length hCgB, was no longer sorted to ISG, but exited from the TGN in constitutive secretory vesicles. Coexpression of full-length hCgB together with Δcys-hCgB by double infection, using the respective recombinant vaccinia viruses, rescued the sorting of the deletion mutant to ISG. In conclusion, our data show that (a) the disulfide-bonded loop is essential for sorting of CgB to ISG and (b) the lack of this structural motif can be compensated by coexpression of loop-bearing CgB. Furthermore, comparison of the two expression systems, transfection and vaccinia virus–mediated expression, reveals that analyses under conditions in which host cell secretory protein synthesis is blocked greatly facilitate the identification of sequence motifs required for sorting of regulated secretory proteins to secretory granules.
Specialized eukaryotic cells, such as endocrine cells and peptidergic neurons, contain a regulated secretory pathway in addition to the constitutive pathway present in all cells (Burgess and Kelly, 1987). Proteins specific for either of the two pathways are segregated from each other at the level of the TGN. Morphological and biochemical data indicate that a key feature in this sorting is the formation of aggregates of regulated secretory proteins in the TGN from which constitutively secreted proteins are excluded (Tooze et al., 1993). A paradigm for the aggregation-mediated sorting are chromogranin A (CgA),1 chromogranin B (CgB, secretogranin I), and secretogranin II (SgII), the three classical members of the granin family with widespread occurrence in secretory granules of neuroendocrine cells (Huttner et al., 1991; Rosa and Gerdes, 1994). It was shown that their selective aggregation is triggered by an acidic pH and millimolar concentration of calcium in vitro (Gerdes et al., 1989; Gorr et al., 1989; Yoo and Albanesi, 1990) and in the TGN (Chanat and Huttner, 1991). This low pH/calcium-induced aggregation is thought to be mediated by structural features common to all granins, in particular, an abundance of acidic residues distributed along the polypeptide chain (Gerdes et al., 1989). Aggregates of regulated secretory proteins exit the TGN in immature secretory granules (ISG), a short lived intermediate in the biogenesis of mature secretory granules (Tooze and Huttner, 1990; Tooze et al., 1991). During maturation of ISG, which includes further condensation of the soluble content and removal of excess membrane, budding of ISG-derived vesicles has been proposed (Tooze et al., 1991). Such vesicles are thought to mediate the constitutive-like secretion of proteinaceous material that remained soluble in the ISG (Arvan and Castle, 1992; Kuliawat and Arvan, 1992).
During exit from the TGN into ISG, regulated secretory proteins are thought to interact with specific components in the membrane of the TGN. Recently it has been proposed that carboxypeptidase E is a membrane-associated binding site, recognizing regulated secretory proteins in the TGN (Cool et al., 1997). The structural features of regulated secretory proteins involved in membrane recognition, often referred to as sorting signals, are poorly understood. To date, no such sorting signals have been unequivocally identified. One dilemma could lie in the fact that it is not single molecules, but rather aggregates, which are sorted to ISG. Therefore, mutated regulated secretory proteins lacking the sorting signal when analyzed by transfection might not be missorted if the mutant protein can coaggregate with endogenous regulated secretory proteins carrying a sorting signal. Coaggregative sorting could explain why mutated regulated secretory proteins were not seen to be affected in their sorting to secretory granules (Burgess et al., 1987; Powell et al., 1988; Chu et al., 1990; Roy et al., 1991; Castle et al., 1992; Chevrier et al., 1993). In the few cases in which effects of mutations have been detected, the analysis was not performed at the level of the TGN, but rather at the level of mature secretory granules (Sevarino et al., 1989; Stoller and Shields, 1989; Cool and Peng Loh, 1994; Cool et al., 1995), and thus it remained open whether sorting to, or storage in, secretory granules was effected. To overcome these two limitations of previous studies we pursued the following strategy. First, we surmised that the identification of sorting signals by a mutagenesis approach would necessitate expression of the mutants under conditions in which there is no synthesis of endogenous regulated secretory proteins, thus precluding coaggregative sorting. Second, to analyze sorting at the level of the TGN, we monitored the exit of proteins from the TGN into ISG by pulse-chase labeling in combination with subcellular fractionation.
Here we study the sorting of human CgB (hCgB), which contains near the NH2 terminus a stretch of 22–amino acid residues that is highly conserved between CgA and CgB in many species. These amino acids form a loop stabilized by a disulfide bond, and are encoded in both proteins by a separate exon, exon 3 (Benedum et al., 1987; Pohl et al., 1990; Iacangelo et al., 1991; Wu et al., 1991; Mouland et al., 1994). It has been shown previously that reductive cleavage of the disulfide bond with DTT in vivo, results in the constitutive secretion of CgB (Chanat et al., 1993), without affecting its aggregative properties (Chanat et al., 1994), suggesting a role of the disulfide bond in the sorting of CgB to the regulated pathway of secretion. Although DTT-induced missorting was specific for CgB and not observed for SgII, which lacks cysteins, it cannot be entirely excluded that the effect of DTT was not due to its reduction of the disulfide bond of CgB, but reflected an effect on a component necessary for the sorting of CgB and not for SgII. In this report, we analyze the sorting of hCgB and of a deletion mutant of hCgB lacking the highly conserved disulfide-bonded loop at the level of the TGN as well as at the level of the mature secretory granule. Recombinant proteins were expressed in the neuroendocrine cell line PC12, either by transfection, i.e., in the presence of endogenous granin synthesis, or by recombinant vaccinia viruses, i.e., in the absence of endogenous granin synthesis.
Materials and Methods
Materials
Hydroxyurea was purchased from Boehringer Ingelheim Bioproducts (Heidelberg, Germany), sodium butyrate from Merck (Darmstadt, Germany), radiochemicals from Amersham Intl. (Buckinghamshire, UK), and cell culture reagents from GIBCO BRL (Gaithersburg, MD).
Cells and Viruses
RK13 (rabbit kidney cells ATCC CCL 37) and Hu143 Tk− (ATCC CRL 8303) were cultured in Eagle's minimal essential medium supplemented with 10% fetal calf serum at 37°C in 5% CO2. PC12 cells (rat pheochromocytoma cells, clone 251; Heumann et al., 1983) were grown in DME supplemented with 10% horse serum and 5% fetal calf serum as previously described (Tooze and Huttner, 1990). Vaccinia virus, wild type (strain vv-WR), and a temperature-sensitive mutant, Ts7 (strain Copenhagen), were kindly provided by H. Stunnenberg (University of Nijmegen, The Netherlands).
Antibodies
The hybridoma supernatant monoclonal antibody 67-C7-2 was used for hCgB detection (Rosa et al., 1992). A polyclonal antibody was used for detection of rat CgB (rCgB; Rosa et al., 1992). For rat SgII (rSgII) detection, a rabbit antiserum against the NH2-terminal peptide (-QRNQLLQKEPDL RLENV-) of rSgII was raised. Anti-myc ascites was a gift from E. Karsenti (European Molecular Biology Laboratory [EMBL], Heidelberg, Germany). A polyclonal anti–human α1-antitrypsin antibody was kindly provided by J.L. Brown (Denver, CO) and bought as IgG fraction from Sigma Chemical Co. (St. Louis, MO).
Expression Vector Cloning
wt-hCgB.
The cDNA of hCgB was released from vector pDS6/SgI (Benedum et al., 1987) by EcoRI/BamHI digestion, blunt-ended, and subcloned into the SmaI site of pGEM4 (Promega, Heidelberg, Germany). hCgB was subcloned into p160/10 (gift from U. Ruether, Medizinische Hochschule, Hannover, Germany) using HindIII and EcoRI as restriction sites. It was exized from p160/10 with HindIII and NotI and subcloned into pCDM8 (Invitrogen, Carlsbad, CA).
Δcys-hCgB.
The deletion mutant of hCgB was constructed using a oligonucleotide-directed mutagenesis system (Amersham Intl., version 2.1). In short, the hCgB cDNA was subcloned from pGEM4 into M13mp19 using HindIII and EcoRI restriction sites. The single-stranded antisense clone was used as a template for mutagenesis. The mutagenic primer, TTCAGGACTTGGCGGCGAGTCACCATTC, was used to obtain the desired deletion. After mutagenesis, double-stranded phage DNA was isolated and screened by digestion with HaeI. Three out of five analyzed phage plaque DNAs showed the expected restriction fragment pattern. The DNA sequence of the deletion mutation from one phage plaque was confirmed by sequencing. DNA from this phage was digested with SalI and XhoI and the restriction fragment containing the deleted region was used to replace the corresponding wild-type restriction fragment in pCDM8/wt-hCgB.
AT.
By HindIII/XbaI digestion of pECE/A1PiTS (Leitinger et al., 1994) the α1-antitrypsin cDNA containing the tyrosine sulfation site (AT) of rat cholecystokinin precursor was released and subcloned into pRC/CMV.
For the construction of wt-hCgB and Δcys-hCgB containing a myc epitope, a ClaI and a KpnI site (underlined) were introduced at the COOH terminus of hCgB by PCR using a T7-primer and as a reverse primer oligonucleotide 5′CCGATCGATCATGGGTACCCCCCTTTGGCTGAATTTC. pGEM4/hCgB served as a template. The amplified fragment was subcloned into pSP73 (Promega) using XhoI and ClaI sites. The resulting vector was digested with ClaI and KpnI. Next, two complementary oligonucleotides, 5′CCATGGAACAAAAACTCATCTCAGAAGAGGATCTGGGAGGATAAT and 5′CGATTATCCTCCCAGATCCTCTTCTGAGATGAGTTTTTGTTCCATGGGTAC encoding for the amino acid sequence MEQKLISEEDLGG of the myc epitope and a COOH-terminal stop codon (underlined) were annealed and inserted. The XhoI–EcoRV restriction fragment was subcloned into pBluescript (Stratagene, La Jolla, CA). The XhoI–EcoRI restriction fragment of the resulting vector containing the sequence encoding for the myc epitope was used to replace the corresponding wild-type restriction fragments in pCDM8/wt-hCgB and pCDM8/Δcys-hCgB, respectively. To obtain the expression vector constructs used for transfection, wt-hCgB and Δcys-hCgB were subcloned into pRC/CMV using XbaI sites.
Preparation of Recombinant Viruses
vv:wt-hCgB.
A plasmid transfer vector with a 7.5K vaccinia promoter flanked by viral thymidine kinase sequences (Chocran et al., 1985), modified by Bujard and co-workers (von Brunn et al., 1988) was linearized with BamHI and ClaI and blunt-ended. The cDNA of hCgB (2.5 kb) was released from vector pDS6/SgI (Benedum et al., 1987) by EcoRI/BamHI digestion, blunt-ended, and subcloned into the plasmid transfer vector under control of the 7.5-K promoter.
Recombinant virus was prepared using Hu143 TK− cells infected at 33°C for 2 h with Ts7 virus and transfected with a mixture of wt vaccinia virus DNA and recombinant plasmid essentially as described (Hänggi et al., 1986). The transfected cells were incubated for 2 d at the restrictive temperature (39.5°C) for Ts7 virus. Recombinant viruses were then further selected and plaque-purified on Hu143 TK− cells in the presence of 100 μg/ml 5-bromodeoxyuridine. Plaque-pure recombinants were tested for expression of hCgB on RK13 cells by immunofluorescence analysis with anti-hCgB antibody and metabolic labeling (see below).
vv:Δcys-hCgB, vv:AT, vv:wt-hCgB-EGFP.
A dominant selection system for recombinant viruses based on the xanthine-guanine phosphoribosyltransferase (gpt) gene of Escherichia coli was used (Falkner and Moss, 1988). Therefore, a gpt gene was introduced into the PstI site of the plasmid transfer vector (constructed by von Brunn et al., 1988). The transfer vector was linearized with PstI and blunt-ended. A DNA fragment encoding the gpt gene was obtained from the pEMBL-I3-gpt plasmid (kindly provided by H. Stunnenberg) after ClaI/EcoRI digestion. The fragment was blunt-ended and ligated into the prepared transfer vector, such that the 7.5-K promoter and the gpt promoter were oriented in opposite directions. In addition, a SalI site in the NH2-terminal region of the gpt gene was removed by partial digestion, treatment of the recessive ends by Klenow-polymerase, and religation. The resulting plasmid transfer vector, termed pAB, was used to generate three recombinant viruses. The vector was linearized with XhoI and SalI and blunt-ended. Δcys-hCgB was released from pCDM8/Δcys-hCgB by SalI/EcoRI digestion, α1-antitrypsin containing the tyrosine sulfation site of rat cholecystokinin precursor was released from pECE/A1PiTS (Leitinger et al., 1994) by HindIII/XbaI digestion and hCgB-EGFP (Kaether et al., 1997) was released from pSP73 by restriction with HindIII/EcoRI. Each fragment was blunt-ended and subcloned into pAB. Generation of recombinant virus and gpt+ selection was carried out as described (Falkner and Moss, 1988) except that recombinant viruses were generated in HuTK− cells and gpt+ selection was carried out with RK13 cells.
Viral Amplifications and Titrations
Vaccinia virus–infected RK13 cells were scraped into the medium 2.5 d post infection (PI), pelleted for 1 h at 31,000 g, resuspended in 10 mM Tris, pH 9.0, and lysed by Dounce homogenization. After low-speed centrifugation to remove nuclei, the virus was sedimented through a 36% sucrose cushion (10 mM Tris, pH 9.0) in an SW28 rotor (Beckman Instruments, Inc., Fullerton, CA) at 15,000 rpm for 1 h. The virus pellet was resuspended in PBS, briefly sonicated, and stored in aliquots at −20°C. Titer estimations of viral stocks were performed on RK13 cells by infecting confluent dishes with dilution series in complete culture medium.
Infections
PC12 cells were plated onto laminin-coated dishes (for metabolic labeling) or on poly(L)lysine-coated coverslips (for immunofluorescence) 5 d before infection. Single infections were done at 100 plaque-forming units per cell (pfu/cell) in complete growth medium containing 5 mM hydroxyurea. For double infections, vv:hCgB-EGFP was used at 400 pfu/cell to adjust the expression level to that of Δcys-hCgB, which was used at 100 pfu/cell. Hydroxyurea was present in the inoculum and in all subsequent culture media. After 1 h at 37°C and 10% CO2 with intermittent agitation, the inoculum was aspirated, the cells were washed with growth medium, and then propagated in complete growth medium. Standard infection times before analysis were 3 h. Deviations from these standard conditions are indicated in the figure legends.
Transfections
Standard protocol: PC12 cells were trypsinized, washed with PBS, and resuspended in PBS at a density of 1.5–2.5 × 107 cells/ml. Electroporation was done with 50 μg DNA in a total volume of 0.8 ml cell suspension using a BioRad Gene Pulser at 960 μF and 250 V (Hercules, CA). Transfected cells were diluted in 15 ml growth medium and plated onto cell culture dishes. Growth medium was changed the next day and, except for the stimulated release experiments, replaced by growth medium containing 10 mM sodium butyrate 17.5 h before pulse labeling.
Metabolic Labeling
To label PC12 cells (3.5-cm dishes) with [35S]methionine, cells were depleted of endogenous methionine by incubation for 30 min at 37°C in methionine-free medium (DME lacking methionine) with 1% dialyzed horse serum and 0.5% dialyzed fetal calf serum, and, thereafter, pulse labeled for 5 min with [35S]methionine. At the end of the pulse, dishes were transferred to 4°C, the medium was aspirated, and the cells were washed twice with 1 ml ice-cold PBS supplemented with 5 mM methionine and 5 mM cysteine. Thereafter, cells were scraped off in 0.5 ml of isotonic buffer (20 mM Tris/HCl, pH 7.4 , 0.3 M sucrose, 10 μg/ml leupeptin, 10 μg/ml aprotinin, 0.25 mM PMSF), and divided into two aliquots. One aliquot was analyzed directly on SDS-PAGE (referred to as total homogenate) to estimate the amount of isolated cells by scanning Coomassie-stained cellular protein bands. From the other aliquot, a heat stable fraction highly enriched for the granins was prepared, as described previously, (Benedum et al., 1987) followed by quantitation of labeled rSgII from the fluorogram.
To label the granins present in the TGN, PC12 cells were preincubated for 30 min with sulfate-free DME, supplemented with 10% of the normal concentration of methionine and cysteine. Thereafter, they were pulse labeled for 5 min with sulfate-free DME containing 3–4 mCi/ml carrier-free [35S]sulfate. To chase the sulfate label, the normal growth medium was supplemented with twice the normal concentration of sulfate. The total amount of [35S]sulfate-labeled proteins always increased slightly over time due to a lag time of chase with [35S]sulfate, as observed previously (Baeuerle and Huttner, 1987).
Depolarization-induced Release
3 d or 3 h after transfection, PC12 cells were labeled with [35S]sulfate for 10 or 5 min, respectively. After 90 min of chase, cells were washed twice with either 5K/Ca medium (127 mM NaCl, 5 mM KCl, 2.2 mM CaCl, 0.33 mM Na2HPO4, 0.44 mM KH2PO4, 4.2 mM NaHCO3, 5.6 mM glucose, and 10 mM Hepes-NaOH; pH 7.4; for 5 mM KCl control and 55 mM KCl release) or 5K/Mg medium (5K/Ca medium in which 2.2 mM CaCl2 was replaced by 10 mM MgCl2; for 55 mM KCl plus 10 mM MgCl2 control). Thereafter, cells were incubated for 10 min in 5K/Ca medium (for 5 mM KCl control), 55K/Mg medium (5K/Mg medium in which 127 mM NaCl and 5 mM KCl were replaced by 77 mM NaCl and 55 mM KCl; for 55 mM KCl plus 10 mM MgCl2 control) or 55K/Ca medium (5K/Ca medium in which 127 mM NaCl and 5 mM KCl were replaced by 77 mM NaCl and 55 mM KCl; for 55 mM KCl release). At the end of the incubation, dishes were placed on ice, medium was removed, and cells and media were analyzed, either after immunoprecipitation or directly by SDS-PAGE and phosphoimaging.
Subcellular Fractionation
The exit of the granins from the TGN into constitutive secretory vesicles (CV) and immature secretory granules was followed by sequential velocity and equilibrium sucrose gradients according to standard procedures (Tooze and Huttner, 1990; Tooze and Huttner, 1992), except for the following changes: velocity and equilibrium gradients were fractionated into 1-ml aliquots, fractions 2–5 from the top of the velocity gradients were pooled and analyzed by equilibrium gradient centrifugation on linear 0.6– 1.6 M sucrose gradients.
Immunoprecipitation
Myc-tagged proteins.
After the indicated chase times, the medium was collected and cells were washed twice with ice-cold PBS. Cells were lysed for 10 min in 0.8 ml (6-cm dishes), or 1.2 ml (10-cm dishes) myc lysis buffer (10 mM Tris-HCl, pH 7.5, 100 mM NaCl, 1 mM EDTA, 1% Triton X-100, 10 μg/ml leupeptin, 0.25 mM PMSF). The lysate was cleared by sequential centrifugation for 10 min at 800 g and 30 min at 100,000 g diluted with 1 vol cell dilution buffer (10 mM Tris-HCl, pH 8.0, 100 mM NaCl, 1 mM EDTA, 10 μg/ml leupeptin, 0.25 mM PMSF). Chase or release media were cleared as done for cell lysates and diluted with 1 vol medium dilution buffer (10 mM Tris-HCl, pH 8.0, 50 mM NaCl, 2 mM EDTA, 1% Triton X-100, 10 μg/ml leupeptin, 0.25 mM PMSF). Samples were incubated for 4 h after which 40 μl of a 50% (wt/vol) suspension of protein A–Sepharose CL-4B (Pharmacia Diagnostics AB, Uppsala, Sweden), swollen and washed three times in PBS and a 1:1 mixture of cell lysis and cell dilution buffer, was added. After a 2-h incubation, the immunocomplexes were washed four times with myc wash buffer (10 mM Tris-HCl, pH 8.0, 100 mM NaCl, 1 mM EDTA, 0.2% Triton X-100). The final pellet was resuspended in Laemmli sample buffer (Laemmli, 1970), boiled, and subjected to SDS-PAGE. In some experiments, an aliquot of the supernatants before dilution with cell or medium dilution buffer was mixed with Laemmli sample buffer, and directly subjected to SDS-PAGE.
AT was immunoprecipitated like myc-tagged proteins with the following changes: lysis buffer: 10 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, 0.5% deoxycholic acid, 10 μg/ml leupeptin, 0.25 mM PMSF; cell dilution buffer: 50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, 0.5% deoxycholic acid, 0.5% SDS, 1 mg/ml BSA, 0.5% low-fat milk powder, 10 μg/ml leupeptin, 0.25 mM PMSF; Medium dilution buffer: cell dilution buffer with 2% Triton X-100 and 1% deoxycholic acid. The antibody incubation was done overnight. Immunocomplexes were washed twice with wash buffer A (10 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1 mM EDTA, 0.2% Triton X-100) and B (10 mM Tris-HCl, pH 8.0, 500 mM NaCl, 1 mM EDTA, 0.2% Triton X-100) and once with C (10 mM Tris-HCl, pH 8.0, 0.2% Triton X-100). For immunoprecipitation, gradient fractions were diluted with 4 vol gradient dilution buffer M (12.5 Tris-HCl, pH 8.0, 125 mM NaCl, 1.25 mM EDTA, 0.63% Triton X-100) before addition of antibody.
Electrophoresis and Immunoblotting
SDS-PAGE was performed and gels were processed as described (Lee and Huttner, 1983). For the analysis of gradient fractions, equal aliquots were acetone precipitated before analysis. Western blots were probed with a monoclonal anti-hCgB (67-C7-2) followed by a goat anti–mouse antibody (Dianova, Hamburg, Germany) and 125I-protein A.
Immunofluorescence
Single and double immunofluorescence was performed following standard procedures (Rosa et al., 1989). The coverslips were mounted on glass slides with 90% glycerol containing 1 mg/ml p-phenylenediamine. Fluorescence was viewed using a conventional Zeiss Axiophot microscope (objective 63 × and 40 ×; Oberkochen, Germany) or a confocal microscope (Leica TCS 4D, Heerbrugg, Switzerland) with ∼800 nm depth for the analyzed sections.
Quantitations
Unless indicated, quantitation was done using a FUJI X-BAS 1000 phosophoimager (Tokyo, Japan). Fluorograms were quantitated by densitometric scanning using the Pharmacia-LKB ultroscan XL.
For determination of the infection rate, cells were fixed after different time periods of infection followed by immunofluorescence with anti-hCgB. For each time point, the percentage of immunopositive cells was calculated by analyzing ∼2,000 cells (Zeiss microscope, objective 40 ×).
Results
Transfected Wild Type and Δcys-hCgB Are Stored in Secretory Granules
To test whether the disulfide-bonded loop of CgB is an essential structure for its sorting to secretory granules, a deletion mutant of hCgB (Δcys-hCgB) was constructed (Fig. 1). This mutant lacks amino acids 16–37 comprising the disulfide-bonded loop of hCgB. Transfection experiments were carried out with wild-type hCgB (wt-hCgB) and Δcys-hCgB containing a myc tag at their COOH termini.
Figure 1.

Schematic representation of wt-hCgB and Δcys-hCgB. Numbers refer to the positions of amino acid residues in wt-hCgB after cleavage of the signal peptide (hatched boxes). Gray arch and box: disulfide-bonded loop; continuous line: disulfide bond between Cys 16 and 37; dotted line: deletion of disulfide-bonded loop.
Trafficking of the recombinant proteins was analyzed by biochemical and morphological methods. We (a) determined secretion kinetics and intracellular storage of recombinant proteins by pulse-chase analysis, (b) characterized the type of vesicle to which recombinant proteins were sorted upon exit from the TGN by sucrose gradient centrifugation, (c) tested the storage of recombinant proteins in secretory granules by depolarization-induced release, and (d) analyzed their subcellular distribution by immunofluorescence analysis.
First, we analyzed whether wt-hCgB and Δcys-hCgB were sorted to the regulated pathway of secretion during ongoing synthesis of endogenous regulated secretory proteins. Both proteins were transiently expressed in the neuroendocrine cell line PC12. Expression of transfected proteins was monitored by [35S]sulfate-labeling, a posttranslational modification of the TGN (Baeuerle and Huttner, 1987), followed by immunoprecipitation with an antibody against the myc tag. At the end of the 5-min pulse, wt-hCgB and Δcys-hCgB were detected as single bands in cell lysates indicating the full-length products (Fig. 2 A, bracket). In addition, after a 90-min chase, two fragments of hCgB of ∼68 kD were observed in cell lysates and media (Fig. 2 A, diamond), which could also be immunoprecipitated with an antibody against hCgB (data not shown). As a marker for constitutive secretion, recombinant human α1-antitrypsin containing a tyrosine sulfation site at the COOH terminus (AT; Leitinger et al., 1994) was expressed in PC12 cells. AT was chosen because its isoelectric point is similar to that of hCgB (Owen and Carrell, 1992; Gerdes and Huttner, 1995), but, in contrast to the latter protein, does not undergo calcium- and low pH-induced aggregation in vitro (Krömer, A., and H.-H. Gerdes, unpublished observation). By immunoprecipitation of [35S]sulfate-labeled AT from cell lysates and media a major band of ∼62 kD (Fig. 2 A, asterisk) was detected. A minor band of lower molecular weight observed in cell lysates at the end of the 5-min pulse (Fig. 2 A, arrowhead) probably reflected sulfation of incompletely sialyted AT, because this form disappeared already after short chase times (data not shown).
Figure 2.

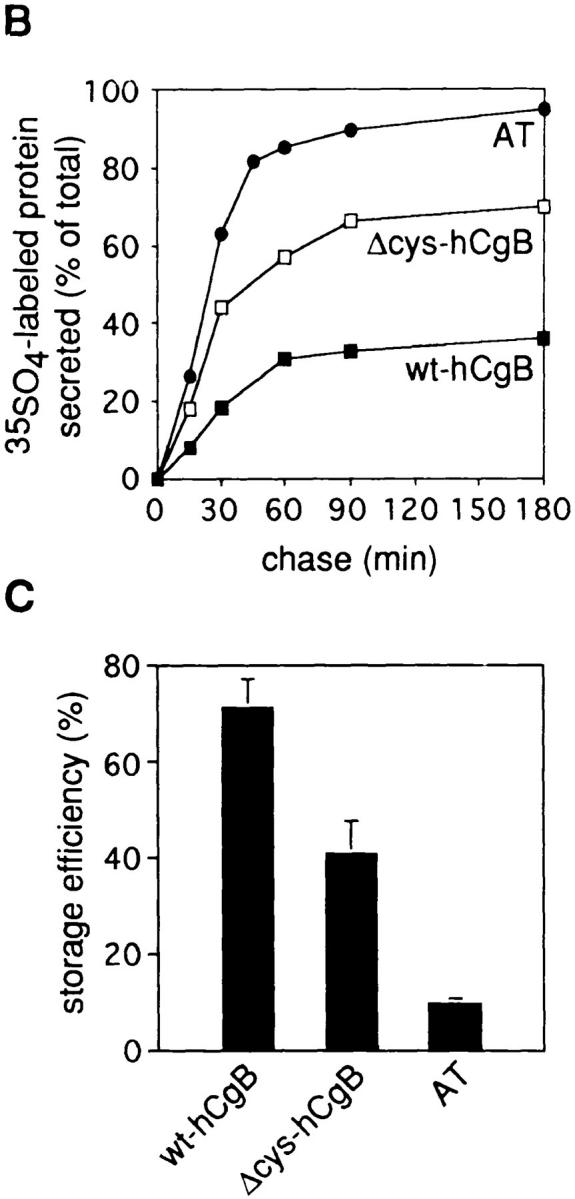
Storage efficiencies of wt-hCgB, Δcys-hCgB, and AT transiently transfected in PC12 cells. PC12 cells transfected with wt-hCgB, Δcys-hCgB, or AT were pulse labeled for 5 min with [35S]sulfate and chased for the indicated times. Equal aliquots of cells and media collected at the end of the chase were analyzed by immunoprecipitation and SDS-PAGE followed by phosphoimaging. (A) Autoradiograms of cell lysates (lanes C) and media (lanes M) at the end of the pulse (0 min of chase) and after 90 min of chase. (Bracket: hCgB; diamond: hCgB fragments; asterisk: AT). Note the slightly slower mobility of Δcys-hCgB as compared to wt-hCgB. (B) Secretion of wt-hCgB, Δcys-hCgB, and AT after different chase times. (C) Storage efficiencies were determined after 90 min of chase (four independent experiments). Error bars show standard deviation. In B and C, the labeled protein in the medium (B) and in the cell (C) is expressed as percent of total (sum of cell plus medium).
Secretion kinetics of transfected wt-hCgB, Δcys-hCgB, and of AT were determined after 5-min sulfate pulse- labeling followed by chase times for up to 3 h (Fig. 2 B). At all time points, the total amount of [35S]sulfate-labeled proteins had increased slightly (see Materials and Methods) indicating that degradation of [35S]sulfate-labeled proteins did not take place during the chase periods. Rapid secretion of all three proteins occurred during the first 60 min and approached a plateau after 90 min of chase. The relative amounts that were released into the medium differed for each protein. We determined the storage efficiency (Seff) for each protein by quantitation of the relative amount stored in the cell after 90 min of chase (Fig. 2 C). Wt-hCgB was stored with a Seff of 70.9% ± 6.0 SD. This value was lower than that determined for endogenous rSgII in transfected cells (Seff: 93.4% ± 2.7 SD), suggesting that granins of species other than rat are stored less efficiently in PC12 cells. Δcys-hCgB lacking the disulfide-bonded loop was stored to 40.8% ± 6.8 SD and the constitutive marker AT to 9.3% ± 1.2 SD. Although the reduction in storage efficiency of Δcys-hCgB as compared to wt-hCgB points to a role of the disulfide-bonded loop in sorting of hCgB, this structure is apparently not essential for its sorting after transfection.
The subcellular compartment to which wt-hCgB and Δcys-hCgB were sorted was characterized by three different approaches. First, we determined the type of vesicle into which wt-hCgB and Δcys-hCgB were sorted upon exit from the TGN. PC12 cells were transfected, pulse labeled for 5 min with [35S]sulfate, and chased for 12 min, after which the transfected protein had completely left the TGN (data not shown). Thereafter, postnuclear supernatants were prepared and subjected to an established subcellular fractionation protocol, which consists of a sequential velocity and equilibrium sucrose gradient centrifugation (Tooze and Huttner, 1990). The distribution across the equilibrium gradients of the heparansulfate proteoglycan (hsPG), a marker for CV, SgII, a marker for ISG and transfected proteins is shown in Fig. 3. Both wt-hCgB (Fig. 3, left) and Δcys-hCgB (Fig. 3, right) were found in fractions also containing rSgII indicating sorting to ISG. A minor amount of Δcys-hCgB (∼30%) comigrated with hsPG (Fig. 3, right), suggesting that it exited from the TGN in CV. The observation that nearly 100% of transfected wt-hCgB and ∼70% of Δcys-hCgB exited into ISG within 12 min but remained in the cell to only ∼70% and ∼40% after 90 min of chase (see Fig. 2 C), respectively, suggests basal release of mature secretory granules or constitutive-like secretion from ISG of the transfected proteins (see Discussion). In contrast to the gradient profiles obtained after 12 min of chase, complete codistribution of wt-hCgB and Δcys-hCgB with rSgII was observed for equilibrium gradients prepared after 90 min of chase at a density characteristic of mature secretory granules (data not shown).
Figure 3.

Transfected wt-hgB and Δcys-hCgB are sorted to immature secretory granules upon exit from the TGN. PC12 cells transfected with wt-hgB (left) or Δcys-hCgB (right) were pulse labeled with [35S]sulfate for 5 min and chased for 12 min. Postnuclear supernatants were prepared and subjected to sequential velocity and equilibrium sucrose gradient centrifugation. Equal aliquots of equilibrium gradient fractions were analyzed, either after immunoprecipitation (hCgB) or directly (hsPG, rSgII) by SDS-PAGE and phosphoimaging. The graphs show the quantitation of [35S]sulfate-labeled hCgB, hsPG, and rSgII across the equilibrium sucrose gradient (fraction 1, top). The arrows indicate the peak of constitutive secretory vesicles (CV) and immature secretory granules (ISG).
Second, we tested whether secretion of wt-hCgB and Δcys-hCgB could be induced by depolarization of transfected PC12 cells. After transfection with either wt-hCgB or Δcys-hCgB, PC12 cells were pulse labeled for 10 min with [35S]sulfate and chased for 90 min. Depolarization of cells by incubation in medium containing 55 mM KCl released 18.2% ± 1.9 SD and 16.1% ± 5.1 SD of wt-hCgB and Δcys-hCgB, respectively (Fig. 4). In contrast, incubation in control medium containing 5 mM KCl or 55 mM KCl plus 10 mM MgCl2 in the absence of calcium resulted in release of 1.6–2.5% of recombinant proteins (Fig. 4). As also shown previously (Bauerfeind et al., 1995), the transfected protein rSgII was released to a similar extent (55 mM KCl: 21.5 ± 2.1 SD, 5 mM KCl: 2.7 ± 2.6 SD, 55 mM KCl/10 mM MgCl2: 4.2 ± 1.0 SD). Consistent with its efficient secretion (Fig. 2), no stimulated release of transfected, sulfate-labeled AT was detectable. The sevenfold increase in secretion of wt-hCgB and Δcys-hCgB after stimulation provides strong evidence that both proteins were stored in secretory granules.
Figure 4.

Depolarization- induced, calcium-dependent secretion of wt-hgB and Δcys-hCgB. PC12 cells transfected with wt-hgB or Δcys-hCgB were pulse labeled with [35S]sulfate for 10 min and chased for 90 min. Cells were then incubated for 10 min, as indicated. Equal aliquots of cells and media were analyzed after immunoprecipitation by SDS-PAGE and phosphoimaging. The graph shows the quantitation of [35S]sulfate-labeled hCgB secreted during the 10-min incubation. Each value is expressed as percentage of total (sum of cells plus medium). Error bars show standard deviation of four independent experiments.
Third, the subcellular distribution of transfected proteins was analyzed by confocal double immunofluorescence. After incubation of transfected PC12 cells with cycloheximide for 90 min, both wt-hCgB (Fig. 5 B) and Δcys-hCgB (Fig. 5 D) were detected as a punctate pattern at the periphery of the cell. Punctate structures immunoreactive for wt-hCgB and Δcys-hCgB were also immunoreactive for endogenous rCgB (compare arrowheads in Fig. 5, A vs. B and C vs. D), suggesting the presence of wt-hCgB and Δcys-hCgB in secretory granules. On the other hand, the marker protein for the constitutive pathway, AT, which could be detected as a perinuclear staining without cycloheximide treatment (Fig. 5 E), was barely detectable in cells after 90 min of cycloheximide chase (Fig. 5 F), consistent with its efficient secretion during pulse-chase labeling (Fig. 2 B and C). In summary, we conclude that both wt-hCgB and Δcys-hCgB expressed by transfection are sorted to ISG after exit from the TGN, and are stored in secretory granules.
Figure 5.

Confocal immunofluorescence analysis of PC12 cells transfected with wt-hCgB, Δcys-hCgB, and AT. Single sections are shown. A–D: PC12 cells were transfected with wt-hCgB (A and B), or Δcys-hCgB (C and D), incubated with cycloheximide for 90 min, and analyzed by double immunofluorescence for endogenous rCgB (A and C) or transfected hCgB (B and D). Note the colocalization of wt-hCgB and Δcys-hCgB with rCgB (arrowheads). E and F: PC12 cells were transfected with AT, incubated without (E) or with (F) cycloheximide for 90 min, and analyzed by single immunofluorescence for AT. Micrographs E and F were obtained with the same confocal setting and exposure time. Bars, 10 μm.
Vaccinia Virus Infection of PC12 Cells Blocks Endogenous Protein Synthesis
We next analyzed sorting of the same recombinant proteins expressed in the absence of endogenous granins. Because all known cell lines bearing the regulated pathway of protein secretion express granins, we set up the vaccinia expression system for PC12 cells. Infection with vaccinia virus has been shown to inhibit host cell protein synthesis and to induce the synthesis of virus-encoded proteins (Moss, 1968).
The viral system was established by infection of PC12 cells with a recombinant virus vv:wt-hCgB which contains wt-hCgB cDNA under the control of the viral 7.5-K promoter. This promoter is active at early and late stages of viral infection. Most importantly, viral infections were performed in the presence of hydroxyurea, a drug known to inhibit the expression of late viral genes (Pogo and Dales, 1971). Thereby viral assembly, in particular, two steps of viral membrane recruitment, at the intermediate compartment (Sodeik et al., 1993) and at the TGN (Schmelz et al., 1994), are blocked, thus preventing disturbance of the secretory pathway.
Infection of PC12 cells with a recombinant virus led to a striking decrease in newly synthesized host cell proteins as determined by pulse labeling of PC12 cells with [35S]methionine. This effect was visible by comparison of total homogenates prepared from either mock-infected or infected PC12 cells (Fig. 6 A, compare total, lanes m and vv). In particular, the cessation of protein synthesis of the two major secretory proteins of PC12 cells, rSgII and rCgB, could be demonstrated after preparation of heat stable fractions (Fig. 6 A, HS). Whereas in heat stable fractions of mock-infected cells rSgII (arrowhead) and rCgB (bracket) were the most prominent bands, in infected cells their expression was reduced to a low level. Residual expression of endogenous granins was due to noninfected cells (see below). At the same time hCgB was expressed under viral control to approximately the same level as either of the two endogenous granins, rSgII and rCgB, and was already visible in total homogenates (Fig. 6 A, total and HS, lanes vv, arrows). The expression level was unchanged from 3 to 8 h PI (data not shown). The identity of hCgB was confirmed by Western blotting of total homogenates with a species-specific antibody that recognizes human, but not rat, CgB. A signal of the expected molecular weight was detected only in infected cells (Fig. 6 B, lane vv, arrow).
Figure 6.
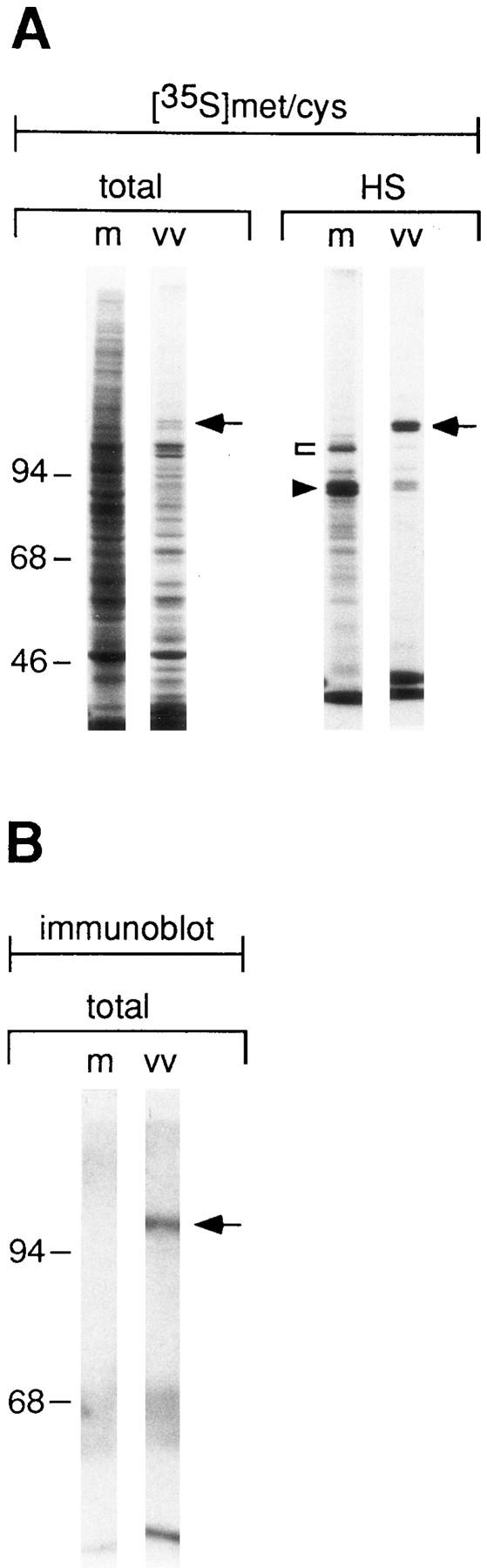
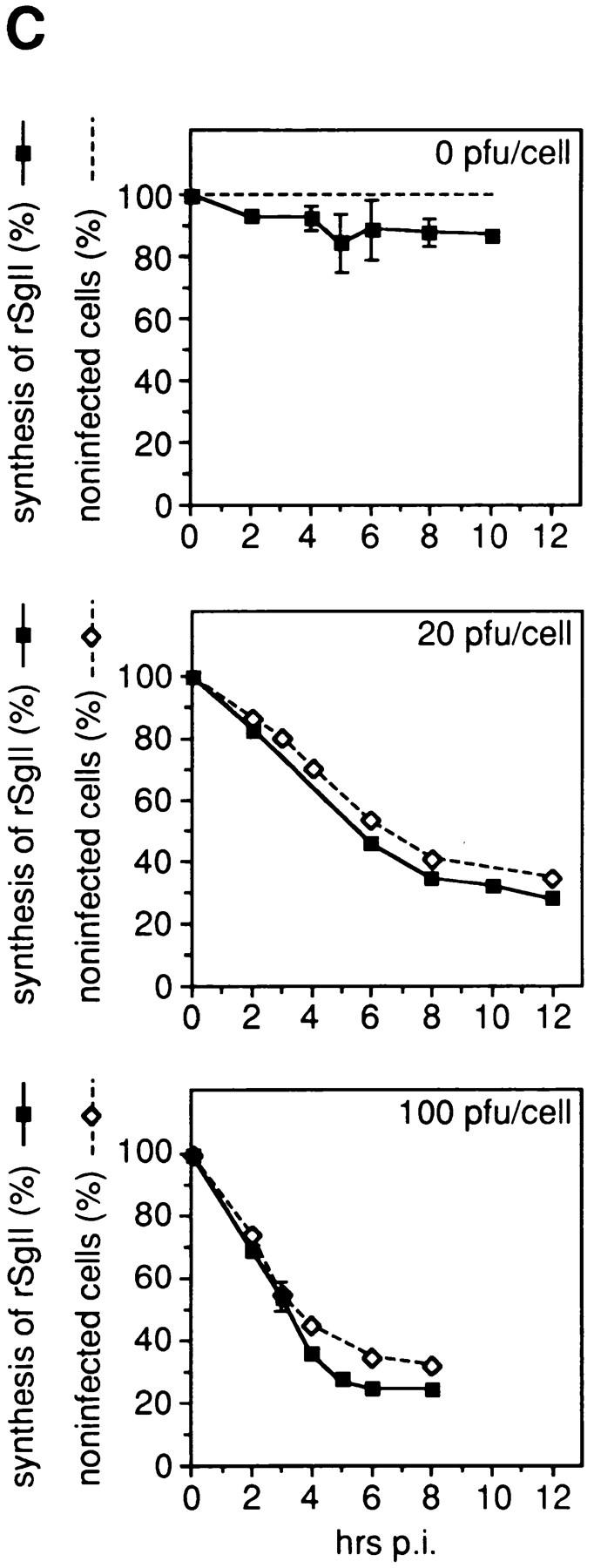
Vaccinia virus infection of PC12 cells blocks endogenous protein synthesis. In A and B, PC12 cells were either mock-infected (m) or infected with vv:wt-hCgB (vv) for 8 h with 20 pfu/ cell. (A) Cells were pulse labeled for 5 min with [35S]methionine. Equal aliquots of cell homogenates (total) and of heat stable fractions prepared therefrom (HS) were analyzed by SDS-PAGE and fluorography. Full-length hCgB, arrows; rCgB, bracket; rSgII, arrowhead. (B) Immunoblot of cell homogenates (total) for hCgB (arrow). (C) PC12 cells were either mock-infected (0 pfu/cell) or infected with vv:wt-hCgB at 20 or 100 pfu/cell. After the indicated time periods of infection (hrs p. i.), cells were either fixed and analyzed for infection by immunofluorescence for hCgB, or analyzed for the synthesis of rSgII by metabolic labeling with [35S]methionine. The percentage of noninfected cells, defined by the lack of hCgB immunofluorescence (broken lines with diamonds), or the extent of rSgII synthesis, expressed as percentage of that obtained before infection (0 hrs p. i.; continuous lines, filled squares) are shown. Data are from single experiments (0 pfu, 0, 2, 10 hrs p. i.; 20 pfu, all time points; 100 pfu, 0, 6, 8 hrs p. i.), or are the mean of duplicate experiments (0 pfu/cell, 4, 5, 6, 8 hrs p. i.), or of triplicate experiments (100 pfu, 2, 3, 4 hrs p. i.). Error bars indicate SEM or the variation of individual values from the mean.
We quantitatively correlated cessation of host cell protein synthesis with the onset of hCgB expression. Expression of hCgB at the cellular level was detected by indirect immunofluorescence with the antibody specific for the human protein. As a marker protein for regulated secretory protein synthesis of the host cell, we followed expression of rSgII. Synthesis of rSgII was monitored by pulse labeling of PC12 cells with [35S]methionine after various times of infection with 0, 20, and 100 pfu per cell. The time courses of the synthesis of rSgII and of the percentage of noninfected cells are shown in Fig. 6 C. For all conditions applied, the block of synthesis of rSgII upon infection was paralleled by the decrease of noninfected cells. By trypan blue staining, after 8 h of infection with 100 pfu/cell, <1% positive cells were detected, indicating that the decrease in rSgII synthesis was not due to cell death (data not shown). The expression of hCgB and shut-off of host cell protein synthesis, therefore, are concurrent events and the remaining expression of rSgII seen at 8 h PI is due to noninfected cells. This implies that the shut-off of host cell protein synthesis starts at the latest with the onset of early viral expression. Furthermore, the decrease in methionine-labeled or sulfate-labeled endogenous rSgII was compared after 3 h PI of either vv:wt-hCgB or vv:Δcys-hCgB at 100 pfu/cell (not shown). Because the same amount in decrease of labeled rSgII was detected for both recombinant viruses, we conclude that the shut-off of endogenous granin synthesis of vv:Δcys-hCgB was as complete as for vv:wt-hCgB.
To obtain direct evidence at the single cell level for a complete block of host cell protein synthesis in infected cells, we used confocal double immunofluorescence. Two constraints of our system have to be considered in this context. First, shut-off at the cellular level cannot be monitored by a lack of endogenous granins because PC12 cells store them in secretory granules located in the cell periphery. Second, immunoreactivity for the endogenous granins (in contrast to that for exogenous hCgB) was, for unknown reasons, not detectable at the level of the PC12 cell Golgi, not even after application of various transport blocks (data not shown). We, therefore, analyzed whether vaccinia virus–expressed hCgB colocalized with endogenous granins at the level of mature secretory granules, as has been shown to be the case for hCgB expressed by transfection (Fig. 5). Lack of colocalization would imply that virally expressed hCgB and endogenous granins were not packaged into the same secretory granule and, thus, were not present in the TGN at the same time. Infected PC12 cells were chased for 90 min with cycloheximide before fixation. In vv:wt-hCgB–infected cells (Fig. 7 A), as was the case in noninfected cells (not shown), endogenous rCgB and rSgII were largely colocalized, with most of the immunostaining being observed at the cell periphery. This result is consistent with both proteins being present in the TGN simultaneously and being copackaged into secretory granules. In contrast, vaccinia virus–expressed wt-hCgB chased to mature secretory granules (see also Figs. 9 and 10) did not colocalize with endogenous rCgB (Fig. 7 B) nor endogenous rSgII (Fig. 7 C), but was observed as distinct punctate structures distributed largely at the cell periphery and to some extent scattered in the cytoplasm. A quantitative analysis of >100 cells revealed that <5% of punctate signals were positive for both virally expressed hCgB and endogenous granins. Bearing in mind that wt-hCgB in transfected cells colocalized with endogenous granins (Fig. 5), we conclude that the synthesis of endogenous granins is shut off under vaccinia virus infection and that the secretory granules containing endogenous granins were made before shut-off, whereas secretory granules containing hCgB were made after shut-off.
Figure 7.
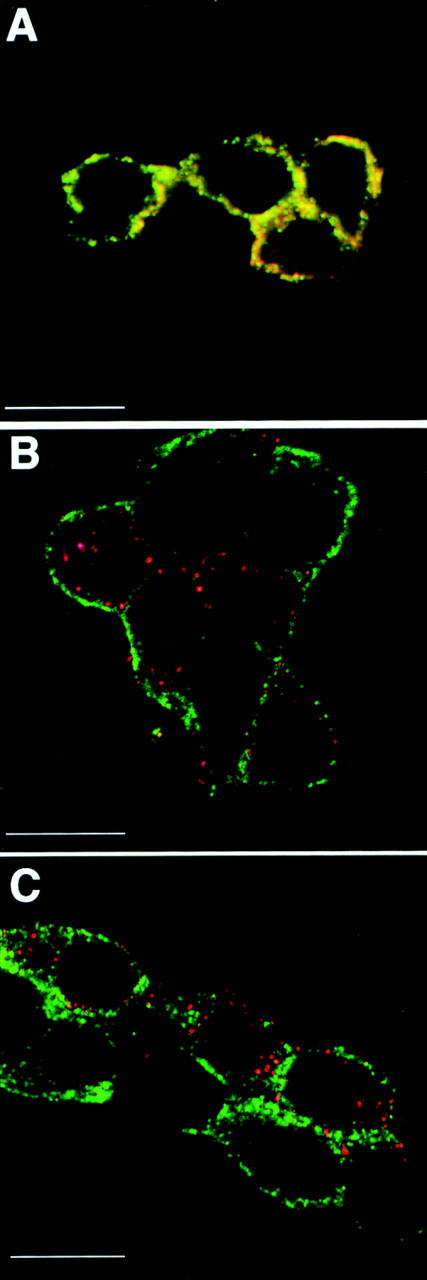
Secretory granules formed under vaccinia infection lack endogenous granins. PC12 cells were infected with vv:wt-hCgB, and then chased for 90 min with cycloheximide, and, thereafter, analyzed by confocal double-immunofluorescence. Single sections are shown. A monoclonal antibody specific for virally expressed hCgB and polyclonal antibodies specific for endogenous rCgB or rSgII were used. (A) Red: α-rCgB, green: α-rSgII. (B) Red: α-hCgB, green: α-rCgB. (C) Red: α-hCgB, green: α-rSgII. Note yellow signal in A reflecting colocalization of endogenous granins and red and green signals in B and C, reflecting distinct localization of virally expressed hCgB and endogenous granins.
Figure 9.

Depolarization- induced, calcium-dependent secretion of wt-hCgB expressed by infection of PC12 with a recombinant vaccinia virus. PC12 cells infected with vv:wt-hCgB were pulse labeled for 5 min with [35S]sulfate and chased for 90 min. Cells were then incubated for 10 min, as indicated, and analyzed for the secretion of [35S]sulfate- labeled wt-hCgB and rSgII. Equal aliquots of media and of heat stable fractions prepared from cell homogenates were analyzed by SDS-PAGE and phosphoimaging. The graph shows the quantitation of [35S]sulfate-labeled hCgB (black columns) and rSgII (gray columns) secreted during the 10-min incubation. Note that [35S]sulfate-labeled rSgII was synthesized in noninfected cells from the same dish. Values are expressed as percentage of total recovered from cells and medium. Error bars show standard deviation of three independent experiments.
Figure 10.


Wt-hCgB but not Δcys-hCgB expressed by recombinant vaccinia viruses is sorted into immature secretory granules upon exit from the TGN and stored in mature secretory granules. PC12 cells, infected with vv:wt-hCgB (left, right) or vv:Δcys-hCgB (middle) were pulse labeled for 5 min with [35S]sulfate and chased for either 12 min (left, middle) or 90 min (right). Postnuclear supernatants were prepared and subjected to sequential velocity and equilibrium sucrose gradient centrifugation. Top: Equal aliquots of the equilibrium gradient fractions were analyzed by SDS-PAGE and phosphoimaging (fraction 1, top). □, hsPG (bracket, area used for quantitation); •, full-length hCgB and fragments (curly bracket); ▵, rSgII. The identity of hCgB fragments was verified by immunoprecipitation. Bottom: The graphs show the quantitation of [35S]sulfate-labeled hCgB, hsPG, and rSgII across the equilibrium sucrose gradients. Symbols correspond to those used in the top panels. The arrows indicate the peak of constitutive secretory vesicles (CV), immature secretory granules (ISG), and mature secretory granules (MSG). Minor peak at the loading position of left panel reflects lysed ISG. Note that [35S]sulfate-labeled hsPG and rSgII were synthesized in noninfected cells from the same dish.
ER to trans-Golgi Transport Rate Is Unaffected after Prolonged Shut-off
To analyze the transport of hCgB in the early secretory pathway of infected cells, we monitored the conversion of newly synthesized hCgB to the mature form, i.e., its arrival in the trans-Golgi, by pulse-chase labeling. After a 3-min pulse with [35S]methionine, the ER form of hCgB was detected as a doublet (Fig. 8 top, arrowhead). With increasing chase time, this form shifted to a higher molecular mass form of ∼125 kD (Fig. 8 top, arrow), which comigrated with [35S]sulfate-labeled hCgB (data not shown), a marker for the mature form in the TGN (Baeuerle and Huttner, 1987). The calculated t 1/2 of transport from ER to Golgi was ∼7.5 min (Fig. 8, bottom). In uninfected PC12 cells, transport of rCgB to the trans-Golgi occurred with a t 1/2 of ∼7 min (Chanat et al., 1993). Our results indicate that host cell protein synthesis shut-off has virtually no effect on ER to Golgi transport of hCgB. Furthermore, the short t 1/2 suggests that endogenous granins synthesized before host cell protein synthesis shut-off are cleared out from the secretory pathway rapidly, and, thus, do not contribute to the cargo of newly formed secretory granules during shut-off.
Figure 8.


Time course of arrival of newly synthesized hCgB at the trans-Golgi. Top: PC12 cells infected with 100 pfu/cell vv:wt-hCgB for 6.5 h were pulse labeled for 3 min with [35S]methionine and chased for the indicated times. At the end of various chase periods, a heat stable protein fraction was prepared from the cells and analyzed by SDS-PAGE and fluorography. trans-Golgi form, arrow; rough ER form of wt-hCgB, arrowhead. Bottom: The trans-Golgi form of [35S]methionine-labeled hCgB is expressed as percentage of total (sum of rough ER and trans-Golgi forms of hCgB). Arrowhead and dashed line, t 1/2 of ER to trans-Golgi transport.
hCgB Is Sorted to the Regulated Secretory Pathway When Protein Synthesis of the Host Cell Is Blocked
The data in Fig. 6 suggest that hCgB is the only regulated secretory protein in the TGN of PC12 cells when expressed by a recombinant vaccinia virus. Sorting of hCgB to secretory granules under these conditions was tested by examining depolarization-induced secretion (Fig. 9). 3 h PI, cells were pulse labeled for 5 min with [35S]sulfate followed by a 90-min chase. Subsequent depolarization of the cells by incubation in medium containing 55 mM KCl released 21.5% ± 2.1 SD of hCgB. Under control conditions (5 mM KCl or 55 mM KCl plus 10 mM MgCl2 in the absence of calcium) 5.0% ± 2.8 SD or 4.2% ± 1.0 SD was, respectively, released. rSgII from noninfected cells was released in comparable amounts, i.e., 19.7% ± 2.9 SD in the presence of 55 mM KCl and 2.7% ± 2.6 SD or 2.1% ± 1.7 SD in control media (5 mM KCl or 55 mM KCl plus 10 mM MgCl2 in the absence of calcium, respectively). These data show that hCgB is stored in a compartment which can undergo depolarization-induced, calcium-dependent exocytosis. This result demonstrates that sorting to the regulated pathway of secretion is not abolished under shut-off conditions.
Virally Expressed wt-hCgB but Not Δcys-hCgB Is Sorted to Immature Secretory Granules Which Undergo Maturation
Having established the vaccinia virus–mediated expression system, we investigated the role of the disulfide-bonded loop in sorting. For this purpose, a recombinant virus expressing a deletion mutant lacking the disulfide-bonded loop (vv:Δcys-hCgB) was generated. To characterize the type of vesicle into which Δcys-hCgB is sorted upon exit from the TGN, sucrose gradient centrifugation analysis of a postnuclear supernatant of infected and pulse-chase labeled cells was performed as described for transfected cells (Fig. 3). wt-hCgB was analyzed in parallel. Both recombinant proteins were detected as full-length forms and in addition as two fragments of ∼68 kD, which have also been observed after transfection (compare with Fig. 2 A). After a 12-min chase period, the majority of wt-hCgB comigrated on the equilibrium sucrose gradient with the ISG marker protein, rSgII, synthesized in noninfected cells of the same dish (Fig. 10, ISG; left). In addition a minor portion codistributed with the constitutive marker protein hsPG. (Fig. 10, CV, left). In contrast Δcys-hCgB comigrated exclusively with hsPG after a 12-min chase (Fig. 10, middle) indicating that it exits from the TGN exclusively into CV.
The depolarization-induced, calcium-dependent release of virally expressed wt-hCgB suggested its presence in secretory granules (Fig. 9). To directly show that ISG in infected cells undergo maturation as has been shown for ISG in uninfected cells (Tooze et al., 1991), equilibrium gradient analysis was performed after a 90-min chase period. As shown in Fig. 10, under these conditions, the peak of wt-hCgB was shifted by one fraction towards higher density as compared to 12 min of chase (Fig. 10, compare fractions 8 and 9, left, to fractions 9 and 10, right). Notably, after 90 min of chase wt-hCgB was detected in the same gradient fractions as rSgII synthesized in noninfected cells. This suggests that maturation of ISG in infected cells occurred within a similar time window as in noninfected cells. In contrast Δcys-hCgB colocalizing with the peak of hsPG after 12 min of chase was barely detectable on equilibrium gradients after a 90-min chase (not shown) suggesting its constitutive secretion into the medium (see below). We conclude from the gradient analyses that after vaccinia virus expression (a) the majority of wt-hCgB is sorted from the TGN into ISG, which undergo maturation during a 90-min chase and (b) Δcys-hCgB exits from the TGN exclusively into CV.
Vaccinia-expressed wt-hCgB but Not Δcys-hCgB Is Stored in Secretory Granules
The differential exit of wt-hCgB and Δcys-hCgB from the TGN indicated sorting of wt-hCgB but not of the mutant to the regulated pathway of secretion. To determine the storage efficiencies of vaccinia-expressed proteins, PC12 cells were infected with either vv:wt-hCgB, vv:Δcys-hCgB, or vv:AT and analyzed by a 5-min pulse with [35S]sulfate followed by a 90-min chase. The recovery of [35S]sulfate-labeled hCgB and AT in cells and media after 90 min of chase was slightly higher than that obtained at the end of the pulse (see Materials and Methods) indicating that no degradation of sulfated proteins occurred under infection conditions. Virally expressed wt-hCgB was stored with an efficiency of 32.2% ± 1.0 SD which is ∼4.5 times higher than that of Δcys-hCgB (Seff: 7.0% ± 2.7 SD; Fig. 11). Notably, after vaccinia expression Δcys-hCgB was secreted like the constitutive marker protein AT (Seff: 8.3% ± 0.8 SD). This result was different from that obtained after transfection. Together with the data obtained by gradient centrifugation it indicates secretion of virally expressed Δcys-hCgB via CV during 90 min of chase.
Figure 11.

Wt-hCgB but not Δcys-hCgB or AT is stored in PC12 cells when expressed by recombinant vaccinia viruses. PC12 cells infected with either vv:wt-hCgB, vv: Δcys-hCgB, or vv:AT were pulse labeled for 5 min with [35S]sulfate, and chased for 90 min. Equal aliquots of cells and media collected at the end of the chase were analyzed, after immunoprecipitation (AT), preparation of heat stable fractions from cell homogenates (hCgB, cells) or directly (hCgB, media), by SDS-PAGE and phosphoimaging. Storage efficiencies of labeled proteins in the cell are expressed as percentage of total (sum of cells plus medium). Error bars show SEM of three independent experiments.
The finding that on the one hand ∼70% wt-hCgB exited the TGN in ISG (Fig. 10, left) and on the other hand the storage efficiency determined after 90 min was ∼32% (Fig. 11) suggests that constitutive-like secretion from ISG or an increased basal release of secretory granules occurred. A similar observation was made in the transfection studies (see Discussion).
Immunofluorescence Analysis of Vaccinia-mediated Expression of wt-hCgB and Δcys-hCgB
Storage of wt-hCgB but not Δcys-hCgB could also be demonstrated by immunofluorescence analysis of infected cells. 3 h PI, cells were immediately fixed or chased for 90 min in the presence of cycloheximide and subsequently fixed. Immunofluorescence analysis for hCgB is shown in Fig. 12. Without cycloheximide treatment, both wt-hCgB (Fig. 12 A, arrows) and Δcys-hCgB (Fig. 12 C, arrows) were detected as a perinuclear staining, which colocalized with TGN38 (data not shown), a marker protein of the TGN. After cycloheximide treatment, the signal in vv: Δcys-hCgB infected cells was almost completely lost (Fig. 12 D), reflecting its efficient secretion as observed by metabolic pulse-chase labeling (Fig. 11). In contrast, virally expressed wt-hCgB showed a peripheral distribution after a 90-min cycloheximide treatment (i.e., 4.5 h PI), characteristic of secretory granules (Fig. 12 B, arrowheads), that is consistent with its storage observed by pulse-chase analysis. Peripheral staining was already visible, although less pronounced, before incubation with cycloheximide (i.e., 3 h PI) in vv:wt-hCgB (Fig. 12 A, arrowheads) but not in vv: Δcys-hCgB (Fig. 12 C) infected cells. Incubation of vv:wt-hCgB infected cells with cycloheximide for up to 3 h did not significantly decrease the hCgB signal when compared to that obtained after 90 min, suggesting that wt-hCgB is neither secreted nor degraded once it has reached the mature secretory granule (data not shown).
Figure 12.
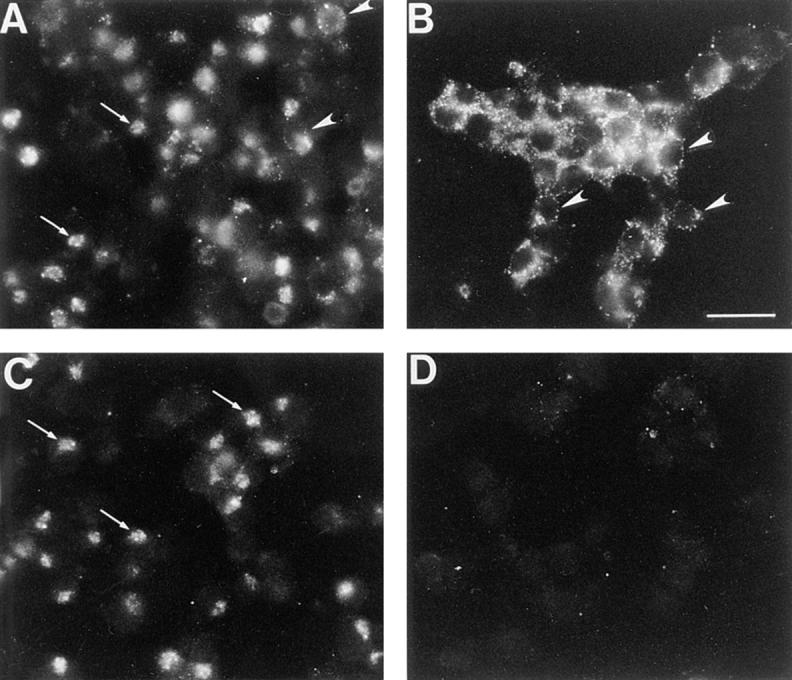
Immunofluorescence of PC12 cells expressing wt-hCgB or Δcys-hCgB by infection with recombinant viruses. PC12 cells were infected with vv:wt-hCgB (A and B) or vv:Δcys-hCgB (C and D) for 3 h, and either fixed (A and C) or incubated for an additional 90 min with cycloheximide, and fixed thereafter (B and D). The micrographs show single immunofluorescence for hCgB. Note that wt-hCgB but not Δcys-hCgB was detected in the cell after incubation with cycloheximide. Arrows, perinuclear staining; arrowheads, peripheral staining; all images are of same magnification. Bar (B), 10 μm.
Sorting of Vaccinia Virus–expressed Δcys-hCgB Is Rescued by Coinfection with a Virus Expressing Full-Length hCgB
Comparison of the sorting behavior of Δcys-hCgB after transfection or infection suggests that, in the case of transfection, the lack of the disulfide-bonded loop was compensated by coaggregation of Δcys-hCgB with loop-containing full-length CgB. If so, double infection of vv:Δcys-hCgB with a recombinant virus expressing full-length hCgB should mimic the transfection conditions and rescue sorting of the mutant to secretory granules. We investigated this issue using [35S]sulfate pulse-chase in combination with subcellular fractionation. This approach has the advantage of providing a direct analysis of protein sorting from the TGN to ISG. In contrast, the three other approaches used to analyze protein sorting to the regulated pathway of secretion (determination of storage efficiency, stimulation of secretion, immunofluorescence) are indirect in that conclusions about sorting at the level of the TGN can only be made if the protein under study that is sorted to ISG is also retained in secretory granules during their maturation.
To investigate whether loop-containing full-length CgB would rescue the sorting of Δcys-hCgB upon double infection, a recombinant virus, vv:wt-hCgB-EGFP, expressing wt-hCgB tagged with enhanced green fluorescent protein (EGFP; Kaether et al., 1997) was generated. The EGFP tag, which does not interfere with the sorting of wt-hCgB to secretory granules (Kaether et al., 1997), makes it possible to distinguish wt-hCgB from Δcys-hCgB on SDS-PAGE due to a shift in molecular weight. PC12 cells were double-infected with vv:Δcys-hCgB and vv:wt-hCgB-EGFP and subjected to sucrose gradient analysis after sulfate pulse-chase as described (Fig. 10, left, middle) to analyze for coaggregative sorting during exit from the TGN. In parallel the same analysis was performed with PC12 cells infected with vv:Δcys-hCgB only. Fig. 13 shows the distribution of sulfated proteins across the equilibrium gradient. Similar to wt-hCgB, the fusion protein wt-hCgB-EGFP is largely found in fractions 8 and 9, the position of ISG, and to a minor extent in fraction 6, the position of CV (Fig. 13 middle, bottom, ▵). In double-infected cells, at least 50% of Δcys-hCgB colocalized with wt-hCgB-EGFP at the position of ISG (Fig. 13 middle, bottom, •), whereas in single infections with vv:Δcys-hCgB, the deletion mutant was almost exclusively found at the position of CV (Fig. 13, top, bottom, □; also Fig. 10, middle). In conclusion, coexpression of Δcys-hCgB with wt-hCgB-EGFP rescues sorting of the mutant to ISG in the vaccinia system.
Figure 13.


Vaccinia coexpression of full-length hCgB rescues sorting of Δcys-hCgB to ISG. PC12 cells were either single- infected with vv:Δcys-hCgB (top, bottom) or double-infected with vv:Δcys-hCgB and vv:wt-hCgB-EGFP (middle, bottom), pulsed for 5 min with [35S]sulfate and chased for 12 min. Postnuclear supernatants were prepared and fractionated by sequential, velocity, and equilibrium sucrose gradient centrifugation. Equal aliquots of the equilibrium gradient (fraction 1, top) were analyzed by SDS-PAGE and phosphoimaging. Top: single infection; □, position of Δcys-hCgB and fragments (curly bracket, see also Fig. 10, middle). Middle: double infection; ▵, position of wt-hCgB-EGFP; •, position of Δcys-hCgB and fragments (curly bracket, see also Fig. 10, middle). Top, middle: endogenous hsPG and rSgII of noninfected cells are indicated by open double arrowheads and open arrows, respectively. Bottom: quantitation of virally expressed recombinant proteins across the equilibrium gradients shown in top and middle panels. The position of CV and ISG is indicated.
Discussion
The Disulfide-bonded Loop Is Essential for Sorting of CgB during Vaccinia Expression
In our study we provide biochemical and morphological evidence that the 22 amino acids of hCgB comprising the disulfide-bonded loop are essential for sorting of CgB to secretory granules and that they affect sorting at the level of the TGN. This result was accomplished using a vaccinia virus–based expression system which inhibits host cell protein synthesis. We have shown by different methods that under these conditions wt-hCgB but not Δcys-hCgB is sorted to secretory granules. First, sucrose gradient analysis showed a differential exit of wt-hCgB and Δcys-hCgB from the TGN. Whereas wt-hCgB was sorted mainly into ISG, which were shown to undergo maturation, Δcys-hCgB was found exclusively in CV (Fig. 10). The reduction in sorting efficiency of wt-hCgB in the vaccinia system (∼70% versus ∼100% after transfection) is most likely due to the shut-off of host cell protein synthesis, which could make factors involved in sorting rate-limiting. Consistent with this explanation, we observed that the sorting efficiency of virally expressed wt-hCgB decreased with increasing infection times (data not shown). Second, after pulse-chase labeling, Δcys-hCgB was secreted like the constitutive marker protein AT whereas wt-hCgB was stored ∼4.5-fold more (Fig. 11). Third, immunofluorescence analysis after 90 min of cycloheximide treatment revealed a pattern for wt-hCgB, which was reminiscent of granule staining. For the deletion mutant, no signal was detected after 90 min of cycloheximide treatment reflecting its efficient secretion (Fig. 12). Furthermore, we have demonstrated that sorting of Δcys-hCgB to ISG can be rescued by coexpression with full-length CgB (Fig. 13), resembling the presence of endogenous granins, as in the case of transfection.
From the finding that the disulfide-bonded loop is necessary for the sorting of hCgB to secretory granules, we conclude that the disulfide-bonded loop comprises either a part or the entire sorting signal. This explanation is consistent with the data obtained by Chanat et al. (1993), who showed that reductive cleavage of the disulfide bond of CgB by DTT caused its missorting to the constitutive secretory pathway. However, the previous DTT data did not strictly exclude an effect of DTT on a component of the secretory machinery for CgB, rather than CgB itself. Our results obtained by expression of Δcys-hCgB clearly indicate that missorting is due to the mutation in CgB.
Sorting of Transfected Δcys-hCgB by Coaggregation
In contrast to the data obtained with the vaccinia expression system, Δcys-hCgB was efficiently sorted to the regulated pathway after transfection. This was determined by pulse-chase labeling (Fig. 2), sucrose gradient centrifugation (Fig. 3), depolarization-induced calcium-dependent release (Fig. 4), and confocal double immunofluorescence analysis (Fig. 5). How can one reconcile the opposing results for Δcys-hCgB obtained by the two different expression systems? The major difference between the two systems is the ongoing synthesis of endogenous proteins after transfection, in contrast to a shut-off of host cell protein synthesis during viral infection, resulting in the presence or absence of endogenous granins in the TGN (Fig. 14). Ongoing protein synthesis during transfection would allow the formation of aggregates between transfected and endogenous regulated secretory proteins enabling the former to enter the regulated secretory pathway by coaggregation. The fact that approximately one-third of Δcys-hCgB was missorted to CV after transfection may be explained by the formation of homotypic aggregates of the mutant. Because, under transfection, Δcys-hCgB is expressed approximately two to three times higher than endogenous granins (not shown), the probability of homotypic Δcys-hCgB aggregates is increased. Indeed the absence and presence of rescuing molecules (full-length CgB) as the principal difference between the two expression systems is clearly shown by coexpression in the vaccinia system of Δcys-hCgB with full-length CgB, which rescues sorting of the mutant to secretory granules (Fig. 13). This implies that sorting of the deletion mutant is accomplished by coaggregation with loop-bearing CgB, i.e., endogenous rCgB in the case of transfection and recombinant wt-hCgB in the vaccinia system (Fig. 14).
Figure 14.

Proposed trafficking of wt-hCgB and Δcys-hCgB after transfection and vaccinia expression. Transfection (left): Both recombinant (filled squares) and endogenous (open squares) granins are present in the TGN. wt-hCgB and Δcys-hCgB are sorted upon exit from the TGN into ISG. Sorting of wt-hCgB (top left, squares with semicircle) is accomplished by the formation of aggregates and by the disulfide-bonded loop. Sorting to ISG of Δcys-hCgB, which lacks the disulfide-bonded loop (middle left, plain squares) is accomplished by its coaggregation with endogenous granins bearing the disulfide-bonded loop. A minor amount of Δcys-hCgB exits from the TGN into CV, probably due to formation of homotypic aggregates lacking endogenous granins. Vaccinia system (right): Recombinant hCgB is present in the TGN whereas endogenous granins are absent due to the shut-off of host cell protein synthesis. After single infection, wt-hCgB (top right, filled squares with semicircle) is mostly sorted to ISG. The small amount exiting from the TGN into CV is due to factors involved in sorting, which become rate-limiting under shut-off. In contrast, Δcys-hCgB lacking the disulfide-bonded loop (middle right, plain filled squares) exits from the TGN completely into CV after single infection. Thus, due to the absence of loop-bearing endogenous granins, sorting of Δcys-hCgB to ISG cannot be rescued by coaggregation. However, after double infection, i.e., expression of Δcys-hCgB together with wt-hCgB-EGFP (filled squares with semicircle and triangle), sorting of Δcys-hCgB to ISG is rescued by coaggregation with loop-bearing wt-hCgB-EGFP, and only a minor portion exits into CV. Thickness of arrows reflects various amounts of trafficking proteins; dashed arrow, absence of traffic.
This explanation is supported by the demonstration of heterophilic interactions between regulated secretory proteins in vitro (Gorr et al., 1989; Palmer and Christie, 1992; Colomer et al., 1996). In particular, it was shown that CgB interacts with CgA at low pH (Yoo, 1996). Because the aggregative properties of CgB are maintained after reduction of the disulfide bond (Chanat et al., 1994) and deletion of the disulfide-bonded loop (A. Krömer and H.-H. Gerdes, unpublished observations), Δcys-hCgB, like wt-hCgB, can be expected to interact with other regulated secretory proteins. Taken together, our data suggest that Δcys-hCgB lacking the disulfide-bonded loop, when expressed by transfection, is sorted to the regulated pathway of secretion by coaggregation with endogenous regulated secretory protein, the latter bearing the sorting signal.
Constitutive-like Secretion in PC12 Cells
Gradient analysis of transfected PC12 cells showed that wt-hCgB and Δcys-hCgB pulse labeled for 5 min with [35S]sulfate exited completely within 12 min from the TGN into post-TGN vesicles. At the end of a 12-min chase, transfected wt-hCgB was detected exclusively in ISG, whereas Δcys-hCgB was found in ISG and to a minor extent in CV (Fig. 3), indicating that ∼30% of the mutant had entered the constitutive pathway of secretion upon exit from the TGN. These data are consistent with a role of the disulfide-bonded loop in sorting to ISG. We assume that the proportion of Δcys-hCgB entering CV did not coaggregate with endogenous granins but formed homotypic aggregates (Fig. 14). After 90 min of chase, a time point at which ISG have largely been converted to mature secretory granules (t 1/2 ∼45 min; Tooze et al., 1991), 70.9 and 40.8% of transfected wt-hCgB and Δcys-hCgB, respectively, were stored in secretory granules (Fig. 2 C). Comparison of the storage efficiencies determined after 90 min of chase with the proportion sorted to ISG after 12 min suggests that a similar proportion of both proteins was secreted in an unstimulated manner after entering the ISG. Two mechanisms, basal release of ISG (Matsuuchi and Kelly, 1991) and constitutive-like secretion (Kuliawat and Arvan, 1992), have been described which could account for this effect. For PC12 cells it has been shown, following rSgII as a marker protein, that basal release occurred at a very low level if at all (Gerdes et al., 1989; Tooze and Huttner, 1990). Similar results were obtained in this study for transfected PC12 cells (Seff of rSgII: 93.4% ± 2.7 SD). Therefore, it is unlikely to be basal release but rather constitutive-like secretion that accounts for unstimulated secretion of wt-hCgB and Δcys-hCgB between 12 and 90 min of chase. This explanation is consistent with data obtained from pancreatic islet cells, in which constitutive-like secretion was restricted to the maturation period of ISG (Kuliawat and Arvan, 1992). During vaccinia expression, unstimulated secretion of wt-hCgB (sorting into ISG ∼65% versus Seff ∼32%) from ISG was observed to a similar extent as in the case of transfection (sorting into ISG ∼100% versus Seff ∼71%). This suggests that in both systems, transfection and infection, the difference between sorting and storage of wt-hCgB is caused by constitutive-like secretion.
Sorting during Host Cell Synthesis Shut-off
The observation that virally expressed wt-hCgB is sorted and packaged into secretory granules under host cell protein synthesis shut-off allows the following conclusions: First, sorting of wt-hCgB to secretory granules is independent of other regulated secretory proteins, i.e., it carries all signals necessary for sorting to the regulated pathway of secretion. Second, involvement of chaperones with a short half-life in the secretory pathway assisting transport or aggregation of wt-hCgB is unlikely. Third, recruitment of other components necessary for granule formation, e.g., membrane proteins and cytosolic factors is, at least for the infection time analyzed, mostly independent of ongoing protein synthesis, and thus can be obtained from preexisting pools.
In the past, the vaccinia virus has been used frequently as a mammalian expression system because of its ability to infect a wide spectrum of cell types yielding high expression levels of the recombinant proteins (Hruby et al., 1986). In particular, processing and sorting of prohormones and proneuropeptides have been studied by infecting fibroblast-like cells, which have only a constitutive pathway of protein secretion, and AtT20 cells, with recombinant vaccinia viruses usually at very low viral multiplicity (Thomas et al., 1986; Edwards et al., 1988; Thomas et al., 1988; Seethaler et al., 1991). However, inhibition of host cell protein synthesis was not analyzed under the conditions used. Because the degree of inhibition varies significantly with viral multiplicity (Moss, 1968) and with cell type (Bablanian et al., 1978), it remains unclear whether processing and sorting was determined in the presence or absence of host cell protein synthesis.
Secretory granule biogenesis under protein synthesis arrest was subject of a study by Moore and co-workers (Brion et al., 1992). Using cycloheximide to block protein synthesis and analyze glycosaminoglycan chain trafficking, Brion et al. found that formation of secretory vesicles was severely impaired. In contrast to a total block of protein synthesis after cycloheximide application, vaccinia infection inhibits only host cell protein synthesis. Under the latter condition, we have shown that expression of a single regulated secretory protein is sufficient for the formation of secretory granules.
Analysis of Sorting Signals in Other Regulated Secretory Proteins
Prosomatostatin is a regulated secretory protein whose sorting to secretory granules has been studied intensively. Its sorting information was shown to reside in the first 82 (Stoller and Shields, 1989) or 78 amino acids (Sevarino et al., 1989) of prosomatostatin. A more detailed mutational analysis of prosomatostatin did not reveal a unique amino acid sequence sufficient for sorting (Sevarino and Stork, 1991). Taking into account that the proregion of somatostatin encodes for more than half of the prohormone, the defined region might exhibit aggregative properties rather than functioning as a sorting signal. Furthermore, by using stimulated secretion to monitor sorting to the regulated pathway of protein secretion, it is not clear to which extent sorting in the TGN or sorting by retention in mature secretory granules had occurred.
Proopiomelanocorticotrophic hormone (POMC) is another model protein for studying sorting to the regulated pathway. When mutants of this hormone with a 25– or 26– amino acid deletion at the NH2 terminus of the proregion were analyzed in Neuro 2A cells, opposing results for the existence of an NH2-terminal–sorting motif were reported (Roy et al., 1991; Chevrier et al., 1993; Cool and Peng Loh, 1994; Cool et al., 1995). Using a retroviral expression system (which does not shut off host cell protein synthesis) Roy et al. showed that the NH2-terminal 26 amino acids are not necessary for sorting. In addition, they provided evidence that sorting information is present in multiple domains of POMC and speculated that cooperation between these domains is needed for sorting of the precursor to secretory granules (Chevrier et al., 1993). In contrast, Cool et al. reported that transfection of a deletion mutant of POMC lacking amino acids 2–26 or 8–20 resulted in its constitutive secretion (Cool and Peng Loh, 1994; Cool et al., 1995). Furthermore, the authors present data that in AtT20 cells the NH2-terminal 26 amino acids of POMC are sufficient to sort the constitutive reporter protein chloramphenicol acetyltransferase to secretory granules (Tam et al., 1993). However, it remains unclear to what degree homo- or heterophilic interactions of the transfected proteins with endogenously regulated secretory proteins were involved in the sorting process. It is possible that such interactions vary considerably with the expression level of the transfected proteins and might account for the opposing results. In addition, as for prosomatostatin, sorting was studied at the level of mature secretory granules, and not in the TGN. Nevertheless, it is interesting that the proposed sorting motif of POMC like the loop of CgB is stabilized by a disulfide bond. This raises the question of whether an exposed loop of amino acids is a general feature of the signal for granular sorting. From theoretical calculations based on the primary sequence analysis of regulated proteins it was suggested that the sorting signal may contain a degenerated amphipathic helix (Kizer and Tropsha, 1991). Consistent with the latter suggestion, the disulfide-bonded loop in hCgB forms an amphipathic helix, as determined by secondary structure prediction.
In summary, our data show that the vaccinia-based expression is a powerful tool for the identification of sorting signals of regulated secretory proteins. Most importantly, it allows us to distinguish sorting signals from aggregative properties. Using this system it should be possible to identify two classes of regulated secretory proteins: (a) proteins containing all the structural information necessary for sorting to secretory granules and which, therefore, are properly sorted when they are present in the TGN as the only cargo protein and (b) proteins lacking a specific sorting signal, but which are competent for aggregation, and whose sorting therefore depends on the former class of regulated secretory proteins. A paradigm for the former class is CgB analyzed in this study. Recently, Natori and Huttner (1996) provided in vivo evidence for a helper function of CgB in sorting of proteins to secretory granules. In AtT20 cells, CgB was shown to promote sorting of the 23-K fragment of POMC, which was further processed in ISG to ACTH. Other candidates that may rely on helper proteins for sorting may be thyroid hormone and luteinizing hormone whose aggregative properties have been analyzed in vitro by Cohn and co-workers (Gorr et al., 1989), and Rindler and co-workers (Colomer et al., 1996), respectively.
We have demonstrated here using the vaccinia expression system that the NH2-terminal disulfide-bonded loop of CgB is necessary for its sorting to secretory granules. Likewise, this system can be used in future studies to analyze sorting of regulated secretory proteins for which transfection studies failed in the identification of a sorting signal. In particular, it will be interesting to identify the nature of sorting signals in regulated secretory proteins that do not contain cysteins, e.g., SgII. It may turn out that exposed loops of amino acids stabilized by other means than disulfide-bonds serve as signal in these proteins.
Acknowledgments
We thank Henk Stunnenberg for advice in vaccinia work, Andrea Betz (MPI für experimetelle Medizin, Göttingen) for preparation of plasmid transfer vector pAB and the recombinant virus vv:Δcys-hCgB, Martin Spieß (Biozeutrum, Basel, Switzerland) for providing the COOH-terminally modified cDNA of α1-antitrypsin, Petra Samenfeld for excellent technical assistance, and Denis Corbeil, Jeremy Garwood, Matthew Hannah, and Christoph Kaether for critical comments on the manuscript. The antiserum against α1-antitrypsin was a gift from Jerry L. Brown.
Abbreviations used in this paper
- AT
α1-antitrypsin containing a tyrosine sulfation site
- CgA
chromogranin A
- CgB
chromogranin B
- CV
constitutive secretory vesicles
- EGFP
enhanced green fluorescent protein
- gpt
guanine phosphoribosyltransferase
- hCgB
human CgB
- hsPG
heparansulfate proteoglycan
- ISG
immature secretory granules
- pfu
plaque-forming unit
- PI
post infection
- POMC
proopiomelanocorticotrophic hormone
- rCgB
rat CgB
- rSgII
rat SgII
- SgII
secretogranin II
- Seff
storage efficiency
- vv
vaccinia virus
Footnotes
W.B. Huttner and H.-H. Gerdes are recipients of grants from the Deutsche Forschungsgemeinschaft (SFB 317/C2 and C7) and the European Commission (Biotechnology ERBBIO4CT960058, TMR ERB-FMRX-CT96-0023).
Address all correspondence to Hans-Hermann Gerdes, Department of Neurobiology, University of Heidelberg, Im Neuenheimer Feld 364, 69120 Heidelberg, Germany. Tel.: 49 6221 548317. Fax: 49 6221 548301. E-mail: hhgerdes@sun0.urz.uni-heidelberg.de
Andreas Krömer's present address is Institut für Klinische Pharmakologie Bobenheim, Prof. Dr. Lücker GmbH, Richard-Wagner-Straße 20, D-67269 Grünstadt, Germany.
References
- Arvan P, Castle D. Protein sorting and secretion granule formation in regulated secretory cells. Trends Cell Biol. 1992;2:327–331. doi: 10.1016/0962-8924(92)90181-l. [DOI] [PubMed] [Google Scholar]
- Bablanian R, Esteban M, Baxt B, Sonnabend JA. Studies on the mechanisms of vaccinia virus cytopathic effects. I. Inhibition of protein synthesis in infected cells is associated with virus-induced RNA synthesis. J Gen Virol. 1978;39:391–402. doi: 10.1099/0022-1317-39-3-391. [DOI] [PubMed] [Google Scholar]
- Baeuerle PA, Huttner WB. Tyrosine sulfation is a trans-Golgi– specific protein modification. J Cell Biol. 1987;105:2655–2664. doi: 10.1083/jcb.105.6.2655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bauerfeind R, Jelinek R, Huttner WB. SynaptotagminI- and II- deficient PC12 cells exhibit calcium-independent, depolarization-induced neurotransmitter release from synaptic-like microvesicles. FEBS Lett. 1995;364:328–334. doi: 10.1016/0014-5793(95)00419-a. [DOI] [PubMed] [Google Scholar]
- Benedum UM, Lamouroux A, Konecki DS, Rosa P, Hille A, Baeuerle PA, Frank R, Lottspeich F, Mallet J, Huttner WB. The primary structure of human secretogranin I (chromogranin B): comparison with chromogranin A reveals homologous terminal domains and a large intervening variable region. EMBO (Eur Mol Biol Organ) J. 1987;6:1203–1211. doi: 10.1002/j.1460-2075.1987.tb02355.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brion C, Miller SG, Moore H-PH. Regulated and constitutive secretion. Differential effects of protein synthesis arrest on transport of glycosaminoglycan chains to the two secretory pathways. J Biol Chem. 1992;267:1477–1483. [PubMed] [Google Scholar]
- Burgess TL, Kelly RB. Constitutive and regulated secretion of proteins. Ann Rev Cell Biol. 1987;3:243–293. doi: 10.1146/annurev.cb.03.110187.001331. [DOI] [PubMed] [Google Scholar]
- Burgess TL, Craik CS, Matsuuchi L, Kelly RB. In vitro mutagenesis of trypsinogen: role of the amino terminus in intracellular protein targeting to secretory granules. J Cell Biol. 1987;105:659–668. doi: 10.1083/jcb.105.2.659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castle AM, Stahl LE, Castle JD. A 13-amino acid N-terminal domain of a basic proline-rich protein is necessary for storage in secretory granules and facilitates exit from the endoplasmic reticulum. J Biol Chem. 1992;267:13093–13100. [PubMed] [Google Scholar]
- Chanat E, Huttner WB. Milieu-induced, selective aggregation of regulated secretory proteins in the trans-Golgi network. J Cell Biol. 1991;115:1505–1519. doi: 10.1083/jcb.115.6.1505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chanat E, Weiβ U, Huttner WB, Tooze SA. Reduction of the disulfide bond of chromogranin B (secretogranin I) in the trans-Golgi network causes its missorting to the constitutive secretory pathway. EMBO (Eur Mol Biol Organ) J. 1993;12:2159–2168. doi: 10.1002/j.1460-2075.1993.tb05864.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chanat E, Weiβ U, Huttner WB. The disulfide bond in chromogranin B, which is essential for its sorting to secretory granules, is not required for its aggregation in the trans-Golgi network. FEBS Lett. 1994;351:225–230. doi: 10.1016/0014-5793(94)00865-5. [DOI] [PubMed] [Google Scholar]
- Chevrier D, Fournier H, Nault C, Zollinger M, Crine P, Boileau G. Targeting of pro-opiomelanocortin to the regulated secretory pathway may involve cooperation between different protein domains. Mol Cell Endocrinol. 1993;94:213–221. doi: 10.1016/0303-7207(93)90170-o. [DOI] [PubMed] [Google Scholar]
- Chocran MA, Puckett C, Moss B. In vitro mutagenesis of the promotor region for a vaccinia virus gene: evidence for tandem early and late regulatory signals. J Virol. 1985;54:30–37. doi: 10.1128/jvi.54.1.30-37.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu WN, Baxter JD, Reudelhuber TL. A targeting sequence for dense secretory granules resides in the active renin protein moiety of human preprorenin. Mol Endocrinol. 1990;90:1905–1913. doi: 10.1210/mend-4-12-1905. [DOI] [PubMed] [Google Scholar]
- Colomer V, Kieska GA, Rindler MJ. Secretory granule content proteins and the luminal domains of granule membrane proteins aggregate in vitro at mildly acidic pH. J Biol Chem. 1996;271:48–55. doi: 10.1074/jbc.271.1.48. [DOI] [PubMed] [Google Scholar]
- Cool, D.R., and Y. Peng Loh. 1994. Identification of a sorting signal for the regulated secretory pathway at the N-terminus of pro-opiomelanocortin. Biochimie (Paris). 76:265–270. [DOI] [PMC free article] [PubMed]
- Cool DR, Fenger M, Snell CR, Loh YP. Identification of the sorting signal motif within pro-opiomelanocortin for the regulated secretory pathway. J Biol Chem. 1995;270:8723–8729. doi: 10.1074/jbc.270.15.8723. [DOI] [PubMed] [Google Scholar]
- Cool DR, Normant E, Shen F-S, Chen H-C, Pannell L, Zhang Y, Loh YP. Carboxypeptidase E is a regulated secretory pathway sorting receptor: genetic obliteration leads to endocrine disorders in Cpefatmice. Cell. 1997;88:73–83. doi: 10.1016/s0092-8674(00)81860-7. [DOI] [PubMed] [Google Scholar]
- Edwards RH, Selby MJ, Mobley WC, Weinrich SL, Hruby DE, Rutter WJ. Processing and secretion of nerve growth factor: expression in mammalian cells with a vaccinia virus vector. Mol Cell Biol. 1988;8:2456–2464. doi: 10.1128/mcb.8.6.2456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falkner FG, Moss B. Escherichia coligpt gene provides dominant selection for vaccinia virus open reading frame expression vectors. J Virol. 1988;62:1849–1854. doi: 10.1128/jvi.62.6.1849-1854.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerdes, H.-H., and W.B. Huttner. 1995. Chromogranin B. A. Haeberli, editor, VCH Verlagsges. mbH, Weinheim 3rd Installment.
- Gerdes H-H, Rosa P, Phillips E, Baeuerle PA, Frank R, Argos P, Huttner WB. The primary structure of human secretogranin II, a widespread tyrosine-sulfated secretory granule protein that exhibits low pH- and calcium-induced aggregation. J Biol Chem. 1989;264:12009–12015. [PubMed] [Google Scholar]
- Gorr S-U, Shioi J, Cohn DV. Interaction of calcium with porcine adrenal chromogranin A (secretory protein-I) and chromogranin B (secretogranin I) Am J Physiol. 1989;257:247–254. doi: 10.1152/ajpendo.1989.257.2.E247. [DOI] [PubMed] [Google Scholar]
- Hänggi M, Bannwarth W, Stunnenberg HG. Conserved TAAAT motif in vaccinia virus late promotors: overlapping TATA box and site of transcription initiation. EMBO (Eur Mol Biol Organ) J. 1986;5:1071–1076. doi: 10.1002/j.1460-2075.1986.tb04324.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heumann R, Kachel V, Thoenen H. Relationship between NGF-mediated volume increase and “priming effect” in fast and slow reacting clones of PC12 pheochromocytoma cells. Exp Cell Res. 1983;145:179–190. doi: 10.1016/s0014-4827(83)80019-6. [DOI] [PubMed] [Google Scholar]
- Hruby DE, Thomas G, Herbert E, Franke CA. Use of vaccinia virus as a neuropeptide expression vector. Methods Enzymol. 1986;124:295–309. doi: 10.1016/0076-6879(86)24022-7. [DOI] [PubMed] [Google Scholar]
- Huttner WB, Gerdes H-H, Rosa P. The granin (chromogranin/ secretogranin) family. Trends Biochem Sci. 1991;16:27–30. doi: 10.1016/0968-0004(91)90012-k. [DOI] [PubMed] [Google Scholar]
- Iacangelo AL, Grimes M, Eiden LE. The bovine chromogranin A gene: structural basis for hormone regulation and generation of biologically active peptides. Mol Endocrinol. 1991;5:1651–1660. doi: 10.1210/mend-5-11-1651. [DOI] [PubMed] [Google Scholar]
- Kaether C, Salm T, Glombik M, Almers W, Gerdes H-H. Targeting of green fluorescent protein to neuroendocrine secretory granules: a new tool for real time studies of regulated protein secretion. Eur J Cell Biol. 1997;74:133–142. [PubMed] [Google Scholar]
- Kizer JS, Tropsha A. A motif found in propeptides and prohormones that may target them to secretory vesicles. Biochem Biophys Res Commun. 1991;174:586–592. doi: 10.1016/0006-291x(91)91457-n. [DOI] [PubMed] [Google Scholar]
- Kuliawat R, Arvan P. Protein targeting via the “constitutive-like” secretory pathway in isolated pancreatic islets: passive sorting in the immature granule compartment. J Cell Biol. 1992;118:521–529. doi: 10.1083/jcb.118.3.521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4 . Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- Lee RW, Huttner WB. Tyrosine-O-sulfated proteins of PC12 pheochromocytoma cells and their sulfation by a tyrosylprotein sulfotransferase. J Biol Chem. 1983;258:11326–11334. [PubMed] [Google Scholar]
- Leitinger B, Brown JL, Spiess M. Tagging secretory and membrane proteins with a tyrosine sulfation site. J Biol Chem. 1994;269:8115–8121. [PubMed] [Google Scholar]
- Matsuuchi L, Kelly RB. Constitutive and basal secretion from the endocrine cell line, AtT-20. J Cell Biol. 1991;112:843–852. doi: 10.1083/jcb.112.5.843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moss B. Inhibition of HeLa cell protein synthesis by the vaccinia virion. J Virol. 1968;2:1028–1037. doi: 10.1128/jvi.2.10.1028-1037.1968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mouland AJ, Bevan S, White JH, Hendy GN. Human chromogranin A Gene. J Biol Chem. 1994;269:6918–6926. [PubMed] [Google Scholar]
- Natori S, Huttner WB. Chromogranin B (secretorgranin I) promotes sorting to the regulated secretory pathway of processing intermediates derived from a peptide hormone precursor. Proc Natl Acad Sci USA. 1996;93:4431–4436. doi: 10.1073/pnas.93.9.4431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owen, M.C., and R.W. Carrell. 1992. Alpha-1-proteinase inhibitor. A. Haeberli, ed., VCH Verlagsges. mbH, Weinheim 1. Installment.
- Palmer DJ, Christie DL. Identification of molecular aggregates containing glycoproteins III, J, K (carboxypeptidase H), and H (Kex2- related proteases) in the soluble and membrane fractions of adrenal medullary chromaffin granules. J Biol Chem. 1992;267:19806–19812. [PubMed] [Google Scholar]
- Pogo BGT, Dales S. Biogenesis of vaccinia: separation of early stages from maturation by means of hydroxyurea. Virology. 1971;43:144–151. doi: 10.1016/0042-6822(71)90232-7. [DOI] [PubMed] [Google Scholar]
- Pohl TM, Phillips E, Song K, Gerdes H-H, Huttner WB, Rüther U. The organisation of the mouse chromogranin B (secretogranin I) gene. FEBS Lett. 1990;262:219–224. doi: 10.1016/0014-5793(90)80194-n. [DOI] [PubMed] [Google Scholar]
- Powell SK, Orci L, Craik CS, Moore H-PH. Efficient targeting to storage granules of human proinsulins with altered propeptide domain. J Cell Biol. 1988;106:1843–1851. doi: 10.1083/jcb.106.6.1843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosa P, Gerdes H-H. The granin protein family: markers for neuroendocrine cells and tools for the diagnosis of neuroendocrine tumors. J Endocrinol Invest. 1994;17:207–225. doi: 10.1007/BF03347721. [DOI] [PubMed] [Google Scholar]
- Rosa P, Weiß U, Pepperkok R, Ansorge W, Niehrs C, Stelzer E H K, Huttner W B. An antibody against secretogranin I (chromogranin B) is packaged into secretory granules. J Cell Biol. 1989;109:17–34. doi: 10.1083/jcb.109.1.17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosa P, Bassetti M, Weiß U, Huttner WB. Widespread occurrence of chromogranins/secretogranins in the matrix of secretory granules of endocrinologically silent pituitary adenomas. J Histochem Cytochem. 1992;40:523–533. doi: 10.1177/40.4.1552186. [DOI] [PubMed] [Google Scholar]
- Roy P, Chevrier D, Fournier H, Racine C, Zollinger M, Crine P, Boileau G. Investigation of a possible role of the amino-terminal pro- region of proopiomelanocortin in its processing and targeting to secretory granules. Mol Cell Endocrinol. 1991;82:237–250. doi: 10.1016/0303-7207(91)90037-s. [DOI] [PubMed] [Google Scholar]
- Schmelz M, Sodeik B, Ericcson M, Wolffe EJ, Shida H, Hiller G, Griffiths G. Assembly of vaccinia virus: the second wrapping cisterna is derived from the trans golgi network. J Virol. 1994;68:130–147. doi: 10.1128/jvi.68.1.130-147.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seethaler G, Chaminade M, Vlasak R, Ericsson M, Griffiths G, Toffoletto O, Rossier J, Stunnenberg HG, Kreil G. Targeting of frog prodermorphin to the regulated secretory pathway by fusion to proenkephalin. J Cell Biol. 1991;114:1125–1133. doi: 10.1083/jcb.114.6.1125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sevarino KA, Stork P. Multiple preprosomatostatin sorting signals mediate secretion via discrete cAMP- and tetradecanoylphorbolacetate- responsive pathways. J Biol Chem. 1991;266:18507–18513. [PubMed] [Google Scholar]
- Sevarino KA, Stork P, Ventimiglia R, Mandel G, Goodman RH. Amino-terminal sequences of prosomatostatin direct intracellular targeting but not processing specificity. Cell. 1989;57:11–19. doi: 10.1016/0092-8674(89)90167-0. [DOI] [PubMed] [Google Scholar]
- Sodeik B, Doms RW, Ericsson M, Hiller G, Machamer CE, van't Hof W, van Meer G, Moss B, Griffiths G. Assembly of vaccinia virus: role of the intermediate compartment between the endoplasmic reticulum and the golgi stacks. J Cell Biol. 1993;121:521–541. doi: 10.1083/jcb.121.3.521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoller TJ, Shields D. The propeptide of presomatostatin mediates intracellular transport and secretion of α-globin from mammalian cells. J Cell Biol. 1989;108:1647–1655. doi: 10.1083/jcb.108.5.1647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tam WWH, Andreasson KI, Peng Y, Loh The amino-terminal sequence of pro-opiomelanocortin directs intracellular targeting to the regulated secretory pathway. Eur J Cell Biol. 1993;62:294–306. [PubMed] [Google Scholar]
- Thomas G, Herbert E, Hruby DE. Expression and cell type-specific processing of human preproenkephalin with a vaccinia recombinant. Science. 1986;232:1641–1643. doi: 10.1126/science.3754979. [DOI] [PubMed] [Google Scholar]
- Thomas G, Thorne BA, Thomas L, Allen RG, Hruby DE, Fuller R, Thorner J. Yeast KEX2 endopeptidase correctly cleaves a neuroendocrine prohormone in mammalian cells. Science. 1988;241:226–230. doi: 10.1126/science.3291117. [DOI] [PubMed] [Google Scholar]
- Tooze SA, Huttner WB. Cell-free protein sorting to the regulated and constitutive secretory pathways. Cell. 1990;60:837–847. doi: 10.1016/0092-8674(90)90097-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tooze SA, Huttner WB. Cell-free formation of immature secretory granules and constitutive secretory vesicles from the trans-Golgi network. Methods Enzymol. 1992;219:81–93. doi: 10.1016/0076-6879(92)19012-u. [DOI] [PubMed] [Google Scholar]
- Tooze SA, Flatmark T, Tooze J, Huttner WB. Characterization of the immature secretory granule, an intermediate in granule biogenesis. J Cell Biol. 1991;115:1491–1503. doi: 10.1083/jcb.115.6.1491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tooze, S.A., E. Chanat, J. Tooze, and W.B. Huttner. 1993. Secretory granule formation. In Mechanisms of Intracellular Trafficking and Processing of Proproteins. Y. Peng Loh, editor. CRC Press, Boca Raton. 157–177.
- von Brunn, A., K. Früh, H. Stunnenberg, and H. Bujard. 1988. 22 nm-like particles of the hepatitis B surface antigen containing epitopes of p190 of P. falciparum. In Technologicial Advances in Vaccine Development. Alan R. Liss, Inc. 167–173.
- Wu H-J, Rozansky DJ, Parmer RJ, Gill BM, O'Connor DT. Structure and function of the chromogranin A gene. J Biol Chem. 1991;266:13130–13134. [PubMed] [Google Scholar]
- Yoo SH. pH- and Ca2+-dependent aggregation property of secretory vesicle matrix proteins and the potential role of chromogranins A and B in secretory vesicle biogenesis. J Biol Chem. 1996;271:1558–1565. [PubMed] [Google Scholar]
- Yoo SH, Albanesi JP. Ca2+-induced conformational change and aggregation of chromogranin A. J Biol Chem. 1990;265:14414–14421. [PubMed] [Google Scholar]