

Abstract
Acetylcholinesterase, with an active site located at the bottom of a narrow and deep gorge, provides a striking example of enzymes with buried active sites. Recent molecular dynamics simulations showed that reorientation of five aromatic rings leads to rapid opening and closing of the gate to the active site. In the present study the molecular dynamics trajectory is used to quantitatively analyze the effect of the gate on the substrate binding rate constant. For a 2.4-Å probe modeling acetylcholine, the gate is open only 2.4% of the time, but the quantitative analysis reveals that the substrate binding rate is slowed by merely a factor of 2. We rationalize this result by noting that the substrate, by virtue of Brownian motion, will make repeated attempts to enter the gate each time it is near the gate. If the gate is rapidly switching between the open and closed states, one of these attempts will coincide with an open state, and then the substrate succeeds in entering the gate. However, there is a limit on the extent to which rapid gating dynamics can compensate for the small equilibrium probability of the open state. Thus the gate is effective in reducing the binding rate for a ligand 0.4 Å bulkier by three orders of magnitude. This relationship suggests a mechanism for achieving enzyme specificity without sacrificing efficiency.
Rapid hydrolysis of acetylcholine by acetylcholinesterase (AChE; EC 3.1.1.7) is essential for cholinergic neurotransmission. This importance is underlined by the large value of kcat/Km, ≈109 M−1⋅s−1 (1–3), which ranks as one of the highest catalytic efficiencies known (4). This magnitude of kcat/Km is striking for AChE, considering the fact that the substrate has to find the active site at the bottom of a narrow but 20-Å deep gorge (5) by diffusion. What is even more remarkable is the fact that, in the x-ray structure of the enzyme (5), the aromatic rings of Tyr-121 and -334 and Phe-290, -330 and -331 completely block the entrance of a 2.4-Å sphere modeling acetylcholine at ≈12 Å from the bottom of the gorge. These rings must reorient to temporarily make way for the incoming substrate (see Fig. 1). Molecular dynamics (MD) simulations (6) demonstrated that, by such reorientation, passage of molecules having the size of acetylcholine is allowed only a small fraction of the time (see Fig. 2). How can the enzyme retain such a high catalytic efficiency?
Figure 1.

Opening and closing of the gate to the active site of AChE. The five aromatic residues serving as the gate are shown such that the viewer is looking down the active-site gorge and into the active site. The surface accessible to a 1.4-Å probe is shown as dots. (Upper) Configuration at 12 ps along a 750-ps MD trajectory. The gate is closed to the 1.4-Å probe (but is open to a 1.2-Å probe). (Lower) Configuration at 68 ps along the MD trajectory. The gate is open even to a 2.6-Å probe (but is closed for a 2.8-Å probe).
Figure 2.

Equilibrium probability po that the gate opens wide enough to admit a substrate with size s. Each circle represents the fraction of time the half-width x of the gate is larger than a particular value of s, calculated from the 750-ps MD trajectory. The curve is a fit assuming that x has a Gaussian distribution.
It was recognized long ago that substrate binding could be slowed down by dynamic modulation of the active site accessibility (7), which may be called conformation gating. In one extreme, conformation gating simply turns the active site from reactive to inert toward the substrate and vice versa, but the enzyme is otherwise identical in the two states. If the switching between the reactive and inert states is stochastic (with rates wr and wi, respectively), Szabo et al. (8, 9) found that the substrate binding rate constant under conformation gating is given by
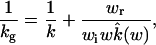 |
1 |
where k is the rate constant when the active site is reactive all the time and wk̂(w) is a related quantity for the same situation, and w = wr + wi. Eq. 1 predicts that, when the switching between the gating states is extremely slow, kg is reduced from k by the equilibrium probability of the reactive state. In the opposite—i.e., fast gating—limit, kg will approach k if the substrate binding is controlled by diffusion.
For AChE, the effect of the conformation gating by the five aromatic rings at ≈12 Å from the bottom of the active-site gorge is better described as modulating the access of the substrate to the active site rather than as directly switching the reactivity of the active site toward the substrate. If this modulation is simplified as a stochastic switch between a fully open and a fully closed state (with transition rates wo and wc, respectively), then the substrate binding rate constant under conformation gating is given by (10)
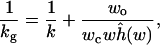 |
2 |
where now w = wo + wc and wĥ(w) is a quantity defined for the situation in which the gate to the active site is open all the time. When gating is fast, it was found (10)
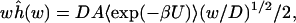 |
3 |
where D is the diffusion constant of the substrate, A is the cross section of the gate to the active site, U is the interaction potential, and 〈exp(−βU)〉 is the average Boltzmann factor on the gate cross section. Previous results (10) indicate that Eq. 3 is useful when 1/w is less than 10−3 of the characteristic diffusion time tD = R2/D, where R is the size of the enzyme. For the present system, R is ≈32 Å and D is 78.3 Å2/ns (11), so Eq. 3 applies when 1/w < 10 ps. Eqs. 2 and 3 are used below to quantitatively demonstrate that, despite conformation gating, AChE can bind acetylcholine with a rate constant as large as 109 M−1⋅s−1. Analysis is specifically done on Torpedo californica AChE because of previous MD and Brownian dynamics work on this species (6, 12), but the results should be relevant for other AChEs because the implicated structural and functional features are also seen in other species (13, 14).
METHODS AND RESULTS
Equilibrium Probability of Open State.
According to the MD simulations (6), the half-width x of the gate to the active site fluctuates and has a Gaussian distribution (Fig. 2 and Fig. 3 Inset). The mean xm and standard deviation σ were estimated to be 1.95 and 0.22 Å, respectively. For a substrate with size s, the open and closed states are equivalent to the regions x > s and x < s, respectively. The equilibrium probability of the open state po = wc/(wo + wc) is thus erfc(η)/2 with η = (s − xm)/√2σ. For s = 2.4 Å, one finds po = 2.4%.
Figure 3.

Number correlation function for the transition of the gate half-width x between x < s = 2.4 Å and x > s. Circles are calculated from the 750-ps MD trajectory as 〈{H[x(t) − s] − po]}{H[x(0) − s] − po]}〉/〈{H[x(0) − s] − po]}2〉 (ref. 15), where H(x) is the step function. The curve is a fit to Eq. 4, which is calculated for diffusion in a harmonic potential. (Inset) Fluctuations of the gate half-width along the 750-ps MD trajectory.
Transition Rates Between Open and Closed States.
We modeled the width fluctuation of the gate as a diffusive motion in a harmonic potential, consistent with the Gaussian distribution of the gate width. The transition between the open and closed states is equivalent to a transition between x > s and x < s. The latter is described by the number correlation function
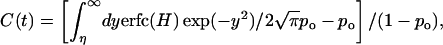 |
4 |
where H = (η − ξy)/(1 − ξ2)1/2, ξ = exp(−Dgt/σ2), and Dg is the effective diffusion constant for the gate width fluctuation. This is calculated by using the Green function for diffusion in a harmonic potential (15, 16). By fitting the number correlation function obtained from a 750-ps trajectory of AChE to Eq. 4, the value of Dg was estimated to be 15 Å2/ns (see Fig. 3). The value of 1/w was identified as the relaxation time of C(t). From Eq. 4 one has
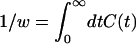 |
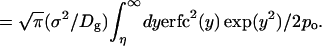 |
5 |
For substrate size s = 2.0, 2.4, and 2.8 Å, Eq. 5 gives 1/w = 1.9, 0.47, and 0.19 ps respectively. This means that Eq. 3 is adequate for calculating wĥ(w).
Average Boltzmann Factor and Cross Section of Gate.
The interaction potential U was calculated by treating the substrate as a test charge in the presence of the electrostatic potential of the enzyme. The latter was calculated by using the uhbd program (17) on the x-ray structure of AChE (5), as described previously (12). From this, the average Boltzmann factor 〈exp(−βU)〉 on the gate cross section was found to be 349 at an ionic strength of 150 mM. The total area A of the entrance to the active site when the gate is fully open was taken to be the substrate-accessible part of a cylindrical cross section modeling the active-site gorge. Let the radius of this cylinder be ras, then
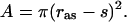 |
6 |
Close to the constriction around the five aromatic rings, the active-site gorge has a more or less uniform cross section, with a radius of roughly 3 Å. This was taken to be the value of ras. For s = 2.4 Å, Eq. 3 gives wĥ(w) = 48 × 109 M−1⋅s−1, and the inverse of the second term on the right-hand side of Eq. 2 is 1.2 × 109 M−1⋅s−1.
Binding rates in the presence and absence of gate.
The inverse of the remaining term in Eq. 2 is the substrate binding rate constant k for a fully open gate. We simply approximated this by the rate constant obtained for a substrate with a reduced size, i.e., 2.0 Å, binding to the enzyme with the x-ray structure, which was previously found to be 1.7 × 109 M−1⋅s−1 by Brownian dynamics simulations (12). Combining this result with the earlier result for the second term, one finds that the substrate rate constant under conformation gating is kg = 0.7 × 109 M−1⋅s−1. Hence conformation gating merely slows down the substrate binding by a factor of 2. This is despite the fact that the gate is open only 2.4% of the time.
Rationalization of the Insignificant Effect of Gate on Substrate Binding Rate.
To gain better insight to the above result, we now write Eq. 3 in a physically meaningful form. Let the linear dimension of the gate cross section be l so A ≈ l2 (see Eq. 6). Then Dl〈exp(−βU)〉 has roughly the same order of magnitude as the “ungated” rate constant k (18). Hence the rate decrease by conformation gating is determined by G = (wc/wo)(wl2/D)1/2, which involves both the equilibrium and the dynamic properties of the gate width fluctuation. A small equilibrium probability of the open state can be compensated by a fast transition between the open and closed states to give a moderate value of G, resulting in an insignificant decrease of the rate constant. Physically, what happens is that, during the period in which the substrate is near the gate (≈l2/D), it will make repeated attempts to enter the gate. If the gating transition is fast so wl2/D ≫ 1, then the gate would have been in the open state numerous times. This fact ensures that at least one of the attempts coincides with an open state of the gate so the substrate can successfully enter the gate.
Binding Rate for a Bulkier Ligand.
The result for a 2.4-Å substrate is to be contrasted with that for a ligand that is bulkier by only 0.4 Å. Now one finds wc/wo = 8.2 × 10−5 and wĥ(w) = 8.6 × 109 M−1⋅s−1. If the ungated rate constant is still taken to be 1.7 × 109 M−1⋅s−1, Eq. 2 gives kg = 0.7 × 106 M−1⋅s−1. This rate is smaller than the earlier result for the 2.4-Å substrate by three orders of magnitude. It is tempting to suggest that this difference in substrate binding rate contributes in part to the difference of three orders of magnitude in kcat/Km for the hydrolysis of acetylcholine and the bulkier butyrylcholine by AChE (4, 19, 20). This difference illustrates that conformation gating may impose substrate specificity without sacrificing efficiency. In comparing these model results to those for real systems, it should be noted that the exact contribution of the gate dynamics to the binding rate may be influenced by the nonharmonic behavior expected for larger fluctuations, and that changes in the size of the substrate may influence steps in the binding process subsequent to passage through the gate.
DISCUSSION
Aromatic Residues Serving as Gate.
The abundance of aromatic residues lining the walls of the active-site gorge has attracted considerable attention (5, 21, 22). In the present context, one may ask why the gate is composed of five aromatic rings. The aromatic rings are distinguished by their oblateness, so a small rotation leads to a large change in the width of the gate to the active site. MD simulations have shown that tyrosine rings may undergo torsional fluctuations on the order of 30° on the picosecond time scale (23). Hence with the aromatic rings serving as the gate, it can very rapidly switch between the open and closed states. As discussed earlier, such rapid switching is required for the proper functioning of conformation gating as a mechanism for enzyme specificity.
Implications for Other Enzymes with Buried Active Sites.
The results obtained in the present study are expected to provide guidance for understanding the specificity and efficiency of other enzymes with buried active sites. In particular, the methodology presented here appears to be directly applicable to cytochrome P450cam, in which the purported entrance to the buried active site is also lined by three aromatic rings (Phe-87 and -98 and Tyr-96) (24, 25). This observation opens the possibility that protein dynamics, just like protein structures, can now be quantitatively analyzed to examine functional implications.
Acknowledgments
This work is supported in part by grants to H.X.Z. from the Research Grants Council of Hong Kong and to J.A.M. from the National Institutes of Health, the National Science Foundation, and the San Diego Supercomputer Center.
ABBREVIATIONS
- AChE
acetylcholinesterase
- MD
molecular dynamics
Footnotes
This paper was submitted directly (Track II) to the Proceedings Office.
References
- 1.Nolte H-J, Rosenberry T L, Neumann E. Biochemistry. 1980;19:3705–3711. doi: 10.1021/bi00557a011. [DOI] [PubMed] [Google Scholar]
- 2.Pryor A N, Selwood T, Leu L-S, Andracki M A, Lee B H, Rao M, Rosenberry R, Doctor B P, Silman I, Quinn D M. J Am Chem Soc. 1992;114:3896–3900. [Google Scholar]
- 3.Radic Z, Gibney G, Kawamoto S, MacPhee-Quigley K, Bongiorno C, Taylor P. Biochemistry. 1992;31:9760–9767. doi: 10.1021/bi00155a032. [DOI] [PubMed] [Google Scholar]
- 4.Fersht A. Enzyme Structure and Mechanism. 2nd Ed. New York: Freeman; 1985. pp. 150–154. [Google Scholar]
- 5.Sussman J L, Harel M, Frolow F, Oefner C, Goldman A, Toker L, Silman I. Science. 1991;253:872–879. doi: 10.1126/science.1678899. [DOI] [PubMed] [Google Scholar]
- 6.Wlodek S T, Clark T W, Scott L R, McCammon J A. J Am Chem Soc. 1997;119:9513–9522. [Google Scholar]
- 7.McCammon J A, Northrup S H. Nature (London) 1981;293:316–317. doi: 10.1038/293316a0. [DOI] [PubMed] [Google Scholar]
- 8.Szabo A, Shoup D, Northrup S H, McCammon J A. J Chem Phys. 1982;77:4484–4493. [Google Scholar]
- 9.Zhou H-X, Szabo A. Biophys J. 1996;71:2440–2457. doi: 10.1016/S0006-3495(96)79437-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhou H-X. J Chem Phys. 1998;108:8146–8154. [Google Scholar]
- 11.Antosiewicz J, Gilson M K, Lee I H, McCammon J A. Biophys J. 1995;68:62–68. doi: 10.1016/S0006-3495(95)80159-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zhou H-X, Briggs J M, McCammon J A. J Am Chem Soc. 1996;118:13069–13070. [Google Scholar]
- 13.Bourne Y, Taylor P, Marchot P. Cell. 1995;83:502–512. doi: 10.1016/0092-8674(95)90128-0. [DOI] [PubMed] [Google Scholar]
- 14.Radic Z, Kirchhoff P D, Quinn D M, McCammon J A, Taylor P. J Biol Chem. 1997;272:23265–23277. doi: 10.1074/jbc.272.37.23265. [DOI] [PubMed] [Google Scholar]
- 15.Zhou H-X. Chem Phys Lett. 1989;164:285–290. [Google Scholar]
- 16.Risken H. The Fokker–Planck Equation. 2nd Ed. Berlin: Springer; 1989. pp. 422–424. [Google Scholar]
- 17.Madura J D, Briggs J M, Wade R C, Davis M E, Luty B A, Ilin A, Antosiewicz J, Gilson M K, Bagheri B, Scott L R, McCammon J A. Comp Phys Comm. 1995;91:57–95. [Google Scholar]
- 18.Zhou H-X. J Chem Phys. 1996;105:7235–7237. [Google Scholar]
- 19.Vellom D C, Radic Z, Li Y, Pickering N A, Camp S, Taylor P. Biochemistry. 1993;32:12–17. doi: 10.1021/bi00052a003. [DOI] [PubMed] [Google Scholar]
- 20.Ordentlich A, Barak D, Kronman C, Flashner Y, Leitner M, Segall Y, Ariel N, Cohen S, Velan B, Shafferman A. J Biol Chem. 1993;268:17083–17095. [PubMed] [Google Scholar]
- 21.Ripoll D R, Faerman C H, Axelsen P H, Silman I, Sussman J L. Proc Natl Acad Sci USA. 1993;90:5128–5132. doi: 10.1073/pnas.90.11.5128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Nair H K, Seravalli J, Arbuckle T, Quinn D M. Biochemistry. 1994;33:8566–8576. doi: 10.1021/bi00194a023. [DOI] [PubMed] [Google Scholar]
- 23.McCammon J A, Wolynes P G, Karplus M. Biochemistry. 1979;18:927–942. doi: 10.1021/bi00573a001. [DOI] [PubMed] [Google Scholar]
- 24.Poulos T L, Finzel B C, Howard A. J Mol Biol. 1987;195:687–700. doi: 10.1016/0022-2836(87)90190-2. [DOI] [PubMed] [Google Scholar]
- 25.Poulos T L. Curr Opin Struct Biol. 1995;5:767–774. doi: 10.1016/0959-440x(95)80009-3. [DOI] [PubMed] [Google Scholar]