

Abstract
The first 10 residues within the Src homology domain (SH)–4 domain of the Src family kinase Fyn are required for binding to the immune receptor tyrosine-based activation motif (ITAM) of T cell receptor (TCR) subunits. Recently, mutation of glycine 2, cysteine 3, and lysines 7 and 9 was shown to block binding of Fyn to TCR ζ chain ITAMs, prompting the designation of these residues as an ITAM recognition motif (Gauen, L.K.T., M.E. Linder, and A.S. Shaw. 1996. J. Cell Biol. 133:1007–1015). Here we show that these residues do not mediate direct interactions with TCR ITAMs, but rather are required for efficient myristoylation and palmitoylation of Fyn. Specifically, coexpression of a K7,9A-Fyn mutant with N-myristoyltransferase restored myristoylation, membrane binding, and association with the cytoplasmic tail of TCR ζ fused to CD8. Conversely, treatment of cells with 2-hydroxymyristate, a myristoylation inhibitor, blocked association of wild-type Fyn with ζ. The Fyn NH2 terminus was necessary but not sufficient for interaction with ζ and both Fyn kinase and SH2 domains were required, directing phosphorylation of ζ ITAM tyrosines and binding to ζ ITAM phosphotyrosines. Fyn/ζ interaction was sensitive to octylglucoside and filipin, agents that disrupt membrane rafts. Moreover, a plasma membrane bound, farnesylated Fyn construct, G2A,C3S-FynKRas, was not enriched in the detergent insoluble fraction and did not associate with ζ. We conclude that the Fyn SH4 domain provides the signals for fatty acylation and specific plasma membrane localization, stabilizing the interactions between the Fyn SH2 domain and phosphotyrosines in TCR ζ chain ITAMs.
Keywords: Src homology domains, acylation, cell membrane, protein-tyrosine kinase, receptor/ antigen
Proteins belonging to the Src family of nonreceptor tyrosine kinases critically rely on binding to the cytoplasmic leaflet of the plasma membrane lipid bilayer to fulfill their biological functions (32, 40). Membrane targeting is specifically guided by the Src homology domain (SH)1–4 motif, a short sequence in the NH2-terminal region of Src family kinases (30, 31). SH4 motifs contain two independent signals (myristate and either palmitate or a polybasic amino acid sequence) that control stable membrane attachment when acting together, but not when operating individually (31, 32). For example, specific and rapid plasma membrane targeting of the Src family kinase Fyn is encoded by dual fatty acylation with myristate at glycine 2 and palmitate at cysteine 3 (1, 35, 41, 43).
Protein myristoylation involves the cotranslational attachment of the 14 carbon fatty acid myristate to the NH2-terminal glycine (glycine 2) on target proteins (8, 17, 42). A chemically stable amide bond is formed between myristate and glycine and is maintained for the lifetime of the protein. The enzymatic reaction is catalyzed by N-myristoyl transferase (NMT), a cytosolic protein that is ubiquitously expressed in eukaryotic cells. The minimum consensus sequence for N-myristoylation is GXXXS/T; glycine 2 is a critical component and a serine or threonine at position 6 is preferred (17). Additional amino acids have been shown to be important for recognition by NMT in the context of specific protein sequences. N-myristoylation of Fyn is a prerequisite for posttranslational and dynamic palmitoylation to occur. Mutations that ablate myristoylation result in loss of palmitoylation (1, 35, 41, 43). Efficient myristoylation of Fyn, therefore, is essential for correct processing and intracellular targeting of Fyn (41, 43).
One of the functional consequences of dual fatty acylation and plasma membrane targeting is to allow Src family kinases to interact with transmembrane receptor complexes. For example, Fyn coimmunoprecipitates with the activated T cell receptor (TCR) and has been reported to interact with multiple TCR subunits (10, 33). Fyn tyrosine kinase activity is stimulated when the TCR is cross-linked, but the molecular nature of the Fyn/TCR interaction is not entirely clear, as the complex is only detected by the use of mild detergents to lyse cells (10, 33, 40). The presence of immune receptor tyrosine-based activation motifs (ITAM) on the ζ chain of the TCR was shown to be essential for interaction with Fyn, although no specific amino acid within the ITAM seemed to be responsible for this interaction (12). Recently, mutation of glycine 2, cysteine 3, and lysines 7 and 9 within the SH4 domain of Fyn was reported to block binding to the ζ chain of the TCR, prompting the designation of these four resides as an ITAM recognition motif (11). However, glycine 2 is required for myristoylation and cysteine 3 is the main site of palmitoylation in Fyn (1, 35, 43). In addition, lysine 7 has been implicated in directing efficient myristoylation of Src (6, 19). Thus, it remained possible that alteration in fatty acylation levels was responsible for the inability of the mutant Fyn to interact with ζ chain.
We therefore reevaluated the role of the SH4 motif of Fyn, focusing on the significance of lysines 7 and 9 in fatty acylation and membrane targeting of Fyn, and its interactions with the TCR ζ chain. Here we show that dual fatty acylation of a K7,9A-Fyn mutant was drastically impaired. A gain of function experiment revealed that overexpression of NMT restored fatty acylation levels of K7,9A-Fyn and allowed the mutant protein to interact with the cytoplasmic tail of the TCR ζ chain fused to CD8. Using the myristoylation inhibitor 2-hydroxymyristate, we observed that association of wild-type Fyn with the ζ chain can be disrupted by blocking fatty acylation. The Fyn NH2 terminus was necessary but not sufficient for interaction with ζ. Both functional Fyn kinase and SH2 domains were required, directing phosphorylation of ζ ITAM tyrosines and subsequent binding of the Fyn SH2 domain to ζ ITAM phosphotyrosines. The interactions between Fyn and ζ were sensitive to conditions that disrupt membrane rafts and a farnesylated Fyn construct that was not enriched in the detergent insoluble fraction did not stably associate with ζ.
These results provide additional explanations for the sequence requirements and mild detergent conditions needed for coimmunoprecipitation of Fyn with TCR subunits. We conclude that dually fatty acylation of Fyn is required for correct targeting to specific membrane subdomains, that is a prerequisite for stable association with the cytoplasmic tail of the TCR ζ chain. Fyn kinase activity is required for phosphorylation of tyrosine residues in the ITAMs of the ζ chain, to which the Fyn SH2 domain subsequently binds. The localization of Fyn in plasma membrane rafts therefore serves an essential role in stabilizing SH2–phosphotyrosine interactions between Fyn and TCR ζ.
Materials and Methods
Materials
Cell culture reagents were purchased from Gibco Laboratories. cDNAs for human wild-type Fyn, G2A-Fyn, and C3,6S-Fyn (1) and NMT (28) and rabbit polyclonal antiserum raised against Fyn SH43 protein (3) were from lab stocks. pEFBOS-CD8-ζ, CD8-ζT76, and CD8-ζ4F (16, 22) were kindly donated by Dr. Arthur Weiss (Department of Immunology, University of California, San Francisco, San Francisco, CA). pSVΔSH2-Fyn, containing mutant Fyn lacking amino acids 144–248, was kindly provided by Drs. Nicolas Dunant and Kurt Balmer-Hofer (Friedrich Miescher Institute, Basel, Switzerland). pEGFP-N1 and rabbit polyclonal antibody against green fluorescent protein (GFP) were obtained from Clontech. Anti–CD8 monoclonal antibody solution and high purity digitonin were from Calbiochem. Protein A–agarose and protein A/G+ agarose beads and antiphosphotyrosine (PY99) monoclonal antibody solution were purchased from Santa Cruz Biotechnology. dl-α-Hydroxymyristic acid (2-hydroxytetradecanoic acid), defatted BSA, and filipin were from Sigma Chemical Co. and n-octylglucoside was from Boehringer Mannheim. Tran35S-label and 125I-NaI were from DuPont-NEN. Synthesis and radioiodination of IC16 (16-iodohexadecanoic acid) and IC13 (13-iodotridecanoic acid) fatty acid analogues were performed as previously described (2).
Plasmid Construction
Construction of plasmids containing wild-type Fyn, G2A-Fyn, C3,6S-Fyn, and Gαo-Fyn was described before (1, 41). For generation of K7A-, K9A-, K7,9A-Fyn, and Lck(10)Fyn constructs, sense oligonucleotides were synthesized to encode the upstream region of pGEM3Z, an NcoI site, and either the first 11–14 amino acids of human Fyn, containing lysines to alanine substitutions at positions 7 and/or 9, or the first 10 amino acids of human Lck, followed by amino acids 11–16 of human Fyn. An antisense oligonucleotide was used corresponding to a region of the SH3 domain of Fyn (1). Sense and antisense primers were used with pGEM3Z-Fyn as a template to generate mutant cDNAs, which were digested with NcoI and BstXI to produce 76-bp fragments that were used to replace the corresponding fragments of Fyn in pSP65. A G2A-FynKRas construct was generated by PCR using a sense oligonucleotide encoding nucleotides 1110–1130 of human Fyn containing a unique BglII site, and an antisense oligonucleotide corresponding to the last 18 nucleotides of Fyn, followed by the last 54 nucleotides of KRas4B, a stop codon, and a SalI site. The primers were used with pGEM3Z-Fyn as a template. The PCR product was digested with BglII and SalI to produce a 500-bp fragment that was used to replace a corresponding carboxy-terminal fragment of G2A-Fyn in pSP65. All Fyn constructs in pSP65 were subsequently digested with EcoRI and SalI, and ligated into pCMV5. FynKRas was subsequently generated by digestion of pCMV5-G2A-FynKRas with BglII and SalI, followed by ligation into BglII and SalI cut pCMV5-Fyn. G2A,C3S-FynKRas was generated by PCR, with pGEM3Z-Fyn as a template, using a sense oligonucleotide encoding an NcoI site followed by the first eight amino acids of human Fyn, containing a glycine-to-alanine substitution at position 2 and a cysteine-to-serine substitution at position 3, and an antisense oligonucleotide corresponding to the SH3 domain of Fyn (1). The PCR product was digested with NcoI and BstXI to generate a 76-bp fragment that was used to replace the corresponding fragment of pSP65-FynKRas. G2A,C3S-FynKRas subcloned into pCMV5 as described above. ΔSH2-Fyn was subcloned into pSP65 by digestion of pSVΔSH2-Fyn with BstXI and BstE2 and ligation into BstXI- and BstE2-digested pSP65, and subcloned into pCMV5, as described above. K299M-Fyn and R176K-Fyn constructs were generated using the Quickchange site-directed mutagenesis kit (Stratagene Inc.). Fyn(16)-GFP was generated by PCR, using a sense oligonucleotide, encoding an EcoRI site followed by the first 16 amino acids of human Fyn fused in frame to amino acid 2 of eGFP, and an antisense oligonucleotide corresponding to the carboxy terminus of eGFP and an XbaI site. These primers were used with pEGFP-N1 as a template to generate a 1.6-kb DNA fragment that was digested with EcoRI and XbaI, followed by ligation into pCMV5.
Cell Culture and Transfection
COS-1 cells (American Type Culture Collection) were cultured and transfected as previously described (41). For cotransfection experiments with Fyn and NMT, 2 μg Fyn cDNA was used with 4 or 10 μg NMT cDNA (ratios 1:2 and 1:5). For CD8-ζ coimmunoprecipitation experiments, 5 μg pEFBOS-CD8-ζ was used with 5 μg pCMV5-Fyn construct and 10 μg pCMV5-NMT or pCMV5.
Metabolic Radiolabeling, Cell Fractionation and Pretreatment with 2-Hydroxymyristate and Filipin
Transfected COS-1 cells were starved for 1 h at 37°C in DMEM minus methionine/cysteine, supplemented with 2% dialyzed FBS, and labeled for 4 h at 37°C with 25 μCi/ml Tran35S-label. For radiolabeling with fatty acid analogues, cells were starved for 1 h at 37°C in DMEM containing 2% dialyzed FBS, followed by radiolabeling for 4 h at 37°C with either 25 μCi/ml 125I-IC13 or 125I-IC16 (1, 41). Fractionation into cytosolic (S100) and total membrane (P100) fractions and extraction with the nonionic detergent Triton X-100 was performed as described previously (41). 2-Hydroxymyristate was stored as a 100-mM stock solution in DMSO and used for experiments at 1 mM in DMEM containing 1% defatted BSA. Before addition to cells, 1 mM 2-hydroxymyristate solution was sonicated briefly and filtered to remove any undissolved myristate analogue. After preincubation for 2 h at 37°C, DMEM containing 5% FBS was added, followed by overnight incubation at 500 μM 2-hydroxymyristate. Where indicated, cells were pretreated for 1 h at 37°C with 10 μg/ml filipin in DMEM containing 1% defatted BSA.
Immunoprecipitation, Gel Electrophoresis, and Quantitation of Fatty Acylation
For immunoprecipitation, each clarified lysate in RIPA buffer (10 mM Tris/HCl, pH 8.0, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, 0.1% SDS, 1% deoxycholate) was mixed with 3 μl anti–Fyn or 3 μl anti–GFP and 10 μl protein A–agarose solution, and incubated for 2–12 h at 4°C. Immunoprecipitates were washed twice with RIPA buffer, once with STE (50 mM Tris/HCl, pH 7.4, 150 mM NaCl, 1 mM EDTA), resuspended in SDS sample buffer containing 0.1 M DTT, and applied to SDS-PAGE. Radiolabeled proteins in dried gels were visualized by exposure for 8–24 h to X-Omat or BioMax film (Eastman Kodak Co.) or to PhosphorImager screens. Quantitation of radiolabeled proteins was performed using the ImageQuant software provided with the PhosphorImager system (Molecular Dynamics, Inc.). In each labeling experiment, constructs were labeled in duplicate with 125I-IC13, 125I-IC16, or Tran35S-label, and the mean of the duplicates was calculated and used as the value for each individual experiment. Numbers were corrected for the background in each individual lane of the gel. Efficiency of myristoylation was calculated as 125I-IC13 incorporation per unit of 35S-label, which is accurate since all Fyn constructs used in this report (see Table I) contain identical amounts of methionines and cysteines. Numbers for myristoylation of mutant constructs were expressed relative to wild-type Fyn within each experiment. Each construct was analyzed in two to four experiments.
Table I.
NH2-terminal Sequences of Wild-Type and Mutant Fyn Constructs
| Amino acid position | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||||||||||
| Fyn | G | C | V | Q | C | K | D | K | E | |||||||||
| G2A-Fyn | A | C | V | Q | C | K | D | K | E | |||||||||
| C3,6S-Fyn | G | S | V | Q | S | K | D | K | E | |||||||||
| K7A-Fyn | G | C | V | Q | C | A | D | K | E | |||||||||
| K9A-Fyn | G | C | V | Q | C | K | D | A | E | |||||||||
| K7,9A-Fyn | G | C | V | Q | C | A | D | A | E | |||||||||
| Lck | G | C | G | C | S | S | H | P | E | |||||||||
| Gαo | G | C | T | L | S | A | E | E | R | |||||||||
Sequences of the first ten amino acids of each Fyn construct are shown with mutant amino acids underlined.
CD8-ζ Coimmunoprecipitation Experiments
10% wt/vol stock solutions of high purity digitonin (Calbiochem Corp.) were prepared according to the manufacturer's instructions. After prolonged storage at 4°C, stocks produce precipitates that were removed by reheating and filtration. For experiments, digitonin lysis buffer (1% digitonin, 25 mM Tris, pH 8.0, 150 mM NaCl, 300 mM KCl, 25 mM NaF, 0.1 mM Na3VO4) was prepared freshly using a clear 10% digitonin stock. Transfected COS-1 cells were lysed for 15 min at 4°C in digitonin buffer supplemented with protease inhibitors (1.5 μg/ml of each leupeptin, aprotinin, and pepstatin A [Boehringer Mannheim], 20 μg/ml each of AEBSF, TLCK, TPCK, and benzamidine [Calbiochem Corp.]). Where indicated, the digitonin lysis buffer was supplemented with 60 mM n-octylglucoside. Lysates were centrifuged for 15 min at top speed in an Eppendorf microfuge at 4°C and incubated for 4 h at 4°C with 2 μl anti–CD8 antibody solution and 10 μl protein A/G+ agarose, or with 10 μl protein A/G+ agarose alone. The anti–CD8 immunoprecipitates were centrifuged for 2 min at top speed in an Eppendorf centrifuge, washed twice with digitonin buffer, and once with ST buffer (10 mM Tris, pH 7.4, 150 mM NaCl). Western blotting analysis for Fyn was performed as described (43), using a rabbit anti–Fyn polyclonal in the primary antibody step. To evaluate the levels of CD8-ζ expression, Fyn blots were stripped and reprobed with monoclonal anti–CD8 solution. As controls for total Fyn expression, supernatants of the anti–CD8 immunoprecipitation step were supplemented with 5× RIPA buffer, 3 μl rabbit anti–Fyn antibody, 10 μl protein A–agarose and incubated for >4 h at 4°C. Immunoprecipitates were washed three times in RIPA buffer and subjected to Western blotting using monoclonal antibody against Fyn (Transduction Laboratories) (43).
Results
Replacement of Lysines 7 and 9 in Fyn Results in Reduced Fatty Acylation and Membrane Anchoring
The first set of experiments was designed to quantitatively assess the contributions of lysine residues within the Fyn SH4 motif towards fatty acylation. COS-1 cells were transfected with cDNAs encoding either wild-type Fyn, G2A-Fyn, or the K7,9A-Fyn construct, in which lysines at positions 7 and 9 are replaced by alanine residues (Table I). Radiolabeling with Tran35S-label, followed by immunoprecipitation of Fyn, SDS-PAGE and autoradiography, showed that all constructs were expressed at similar levels (Fig. 1). The K7,9A-Fyn construct displayed moderately elevated levels of expression and migrated as a doublet (Fig. 1). Wild-type Fyn was labeled with both 125I-IC13 (a myristate analogue) and 125I-IC16 (a palmitate analogue), while G2A-Fyn was not (Fig. 1, and Table II), as described previously (1, 41, 43). Although the K7,9A-Fyn construct contains both the glycine at position 2 and the cysteine at position 3 required for efficient myristoylation and palmitoylation of Fyn (1, 41, 43), levels of 125I-IC13 and 125I-IC16 incorporation were drastically reduced (Fig. 1). In subsequent experiments, K7A-Fyn and K9A-Fyn mutants, containing alanine replacements of either lysine 7 or 9, were included to analyze the relevance of the individual lysines for fatty acylation. Radiolabeling with Tran35S-label and 125I-IC13, followed by calculation of expression levels and relative myristoylation (Table II), showed that mutation of lysine 7 or 9 individually reduced myristoylation of Fyn by 51 and 40%, respectively (Table II). Myristoylation of the double mutant, K7,9A-Fyn, was inhibited by 83% (Table II), implying that the effect of each lysine mutation on myristoylation is additive. In agreement with the dependence of palmitoylation on myristoylation, the level of 125I-IC16-palmitate labeling of K7,9A-Fyn was also found to be reduced to 22 ± 3% (n = 4) of wild-type levels.
Figure 1.

Fatty acylation of Fyn, G2A-Fyn, and K7,9A-Fyn in COS-1 cells. COS-1 cells were transfected with cDNAs encoding wild-type Fyn, G2A-Fyn, and K7,9A-Fyn, and radiolabeled with either Tran35S-label, 125I-IC13, or 125I-IC16. Cell lysates were immunoprecipitated with anti–Fyn antibody and analyzed by SDS-PAGE and autoradiography. Gels were exposed to film for 8–24 h, and strips corresponding to the region of the gel containing p59Fyn are shown. No other radiolabeled proteins were apparent on the gel.
Table II.
Relative Myristoylation Levels of Wild-Type and Mutant Fyn Constructs in COS-1 Cells
| Construct | 125I-IC13 incorporation per 35S-labeled unit of Fyn protein | |||||||
|---|---|---|---|---|---|---|---|---|
| 35S-Met | 125I-IC13 | 125I-IC13/35S-Met | n | |||||
| Fyn | 100% | 100% | 100% | 8 | ||||
| G2A-Fyn | 87 ± 5 | 1 ± 1 | 0 | 2 | ||||
| K7A-Fyn | 145 ± 8 | 73 ± 6 | 49 ± 2 | 2 | ||||
| K9A-Fyn | 195 ± 6 | 116 ± 8 | 60 ± 4 | 2 | ||||
| K7,9A-Fyn | 160 ± 12 | 28 ± 6 | 17 ± 3 | 5 | ||||
| Lck(10)Fyn | 125 ± 3 | 104 ± 3 | 91 ± 3 | 2 | ||||
COS-1 cells were transfected with Fyn cDNA as indicated and radiolabeled as described in Fig. 1. Gels were exposed to PhosphorImager screens for 24 or 48 h, and incorporation of Tran35S-label or 125I-IC13 into wild-type and mutant Fyn was measured using the ImageQuant software provided with the PhosphorImager system. Values for 35S-label in Fyn ranged from 50,000 to 200,000 PhosphorImager units, and for 125I-labeled Fyn from 10,000 to 50,000 units. Background numbers comprised 5–10% of total values for both labeling procedures. Incorporation of radiolabel into the Fyn constructs is expressed relative to wild type. n, number of experiments performed for each construct.
Lck is the only Src family member that lacks lysines at positions 7 and 9 (36, 37). Quantitative analysis of fatty acylation of full length Lck was hampered by inconsistent immunoprecipitation results using different batches of anti–Lck antibody. Therefore, to test whether the absence of lysines 7 and 9 impairs myristoylation of the Lck amino terminus, we used an Lck(10)Fyn fusion construct (Table I). As shown in Table II, myristoylation of Lck(10)Fyn was very similar (∼90%) to that of Fyn, and palmitoylation of Lck(10)Fyn was ∼1.4× more efficient than that of Fyn (data not shown). These results show that despite the lack of lysines 7 and 9 in Lck, the overall fatty acylation profile at its amino terminus is very similar to that of Fyn. It is relevant to note that Lck contains a serine at position 6 (Table II), a preferred residue at this site for mammalian NMT recognition (17).
Since fatty acylation of Fyn is essential for membrane binding (1, 41, 43), the reduced levels of fatty acid modification of K7,9A-Fyn would be expected to alter the subcellular distribution of this construct. Transfected cells expressing the various Fyn constructs were radiolabeled with Tran35S-label, 125I-IC13 or 125I-IC16, followed by homogenization and fractionation into high-speed supernatant (S100) and membrane (P100) fractions, and the distribution of Fyn was analyzed. Fig. 2 shows that more than 95% of wild-type Fyn was recovered from the membrane fraction as dually acylated protein, whereas ∼75% of G2A-Fyn was recovered from the soluble fraction, in agreement with previous results (1, 41, 43). In addition, slightly more than 50% of K7,9A-Fyn was found in the soluble fraction (Fig. 2 B), specifically consisting of the protein band with the slower migration, that is neither myristoylated nor palmitoylated (A). The reason for this difference in migration is not clear, and it cannot be explained simply by the absence of fatty acylation since the G2A-Fyn mutant does not show this effect. The remainder of K7,9A-Fyn was recovered from the membrane fraction as dually acylated protein (Fig. 2 A). The calculated ratio of 125I-IC13 label to Tran35S-labeled in the P100 fraction was 25% lower for K7,9A-Fyn compared with wild-type Fyn (data not shown), implying that some of the protein in the membrane fraction is not acylated. This is confirmed by the results with G2A-Fyn, for which ∼25% of the protein is found in the membrane pellet, even though none of the protein is fatty acylated (Fig. 2 and Table II).
Figure 2.

Fatty acylation and subcellular distribution of Fyn, G2A-Fyn, and K7,9A-Fyn in COS-1 cells. (A) Transfected cells were radiolabeled with either Tran35S-label, 125I-IC13, or 125I-IC16. Cells were homogenized and fractionated into S100 (S) and P100 (P) fractions, followed by immunoprecipitation, SDS-PAGE, and autoradiography. Gels were exposed to film for 8–24 h. (B) Quantitation of the subcellular distribution of 35S-labeled wild-type and mutant Fyn by PhosphorImaging. Gels were exposed to PhosphorImager screens for 8–24 h. Values represent the mean of four independent experiments.
Overexpression of Enzymatically Active NMT Rescues the Impaired Fatty Acylation and Membrane Binding of K7,9A-Fyn
Cotransfection experiments were performed to investigate whether the reduced myristoylation of the K7,9A-Fyn mutant was related to levels of NMT activity inside cells. COS-1 cells were transfected with Fyn or K7,9A-Fyn cDNA alone (Fig. 3, lanes 1 and 2), or cotransfected with cDNA encoding either wild-type NMT (lanes 3 and 4) or H218N-NMT (lanes 5 and 6). H218N-NMT contains a point mutation in the active site that abolishes transfer of myristoyl-CoA to protein (28), and serves as a negative control for the requirement of NMT activity. The ratio of NMT cDNA to Fyn cDNA used for transfection was 2:1. Fig. 3 shows that Fyn (lanes 1, 3, and 5) and K7,9A-Fyn protein (lanes 2, 4, and 6) were expressed at similar levels, with K7,9A-Fyn exhibiting an extra band with a slower migration and reduced myristoylation relative to wild-type Fyn (compare lanes 1 and 2). Cotransfection of K7,9A-Fyn with wild-type NMT increased 125I-IC13 labeling two- to threefold (compare Fig. 3, lanes 1 and 2 with lanes 3 and 4, and see Fig. 4 B), whereas cotransfection with enzymatically inactive H218N-NMT had no effect (compare Fig. 3, lanes 1 and 2 with lanes 5 and 6). Cotransfection of K7,9A-Fyn with wild-type NMT also increased 125I-IC16 incorporation of K7,9A-Fyn twofold (data not shown). In addition, the relative intensity of the lower band in the K7,9A-Fyn doublet increased (Fig. 3, lane 4), consistent with an increase in fatty acylation levels.
Figure 3.

Myristoylation of wild-type Fyn and K7,9A-Fyn coexpressed in COS-1 cells with wild-type or mutant NMT. Cotransfection of Fyn and NMT constructs in COS-1 cells was performed using 2 μg Fyn cDNA and 4 μg NMT cDNA. Radiolabeling of transfected cells, followed by immunoprecipitation, SDS-PAGE, and autoradiography was performed as described in Materials and Methods.
Figure 4.
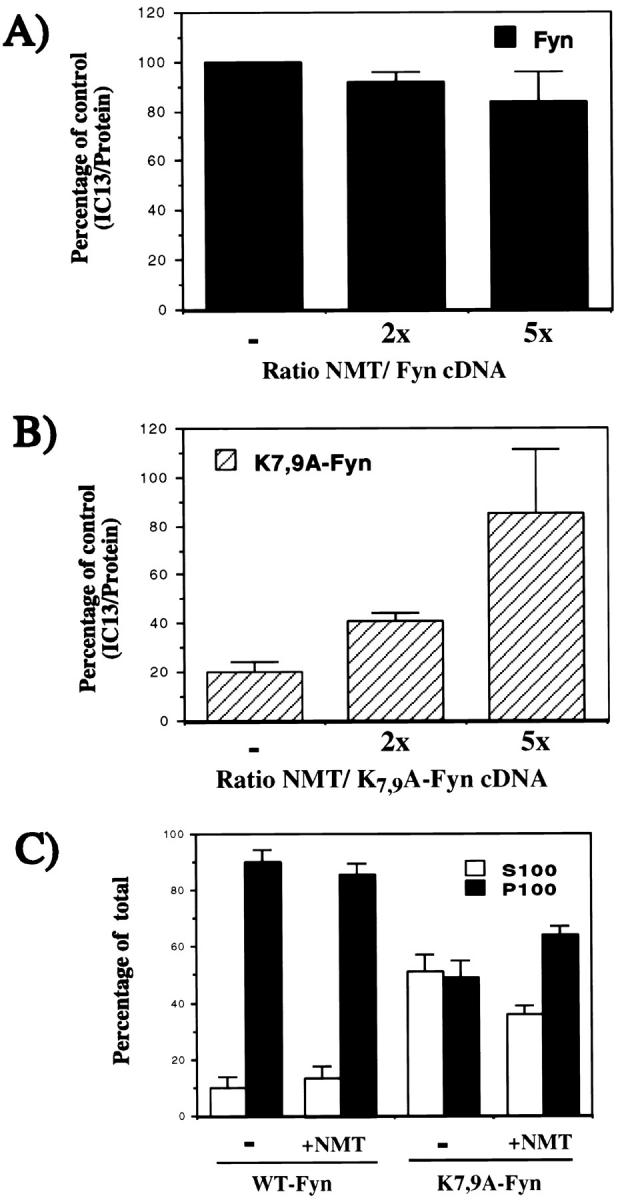
Myristoylation efficiency and subcellular distribution of wild-type Fyn and K7,9A-Fyn after cotransfection in COS-1 cells with varying amounts of NMT cDNA. Cotransfection of Fyn and NMT constructs in COS-1 cells was performed as described in Materials and Methods, using 2 μg Fyn cDNA and 0, 4, or 10 μg NMT cDNA. Radiolabeling of cells transfected with (A) wild-type Fyn or (B) K7,9A-Fyn, followed by immunoprecipitation, SDS-PAGE, and quantitation of 125I-IC13 incorporation per unit 35S-labeled protein by PhosphorImaging was performed as described in Table II. Values are expressed relative to wild-type Fyn, expressed in the absence of exogenous NMT, and numbers represent the mean of four independent experiments. (C) Radiolabeling of cells transfected with 2 μg Fyn or K7,9A-Fyn cDNA and 4 μg NMT cDNA, followed by homogenization and subcellular fractionation. Analysis of protein distribution was performed as described in Fig. 2. Values represent the mean of four independent experiments.
The data in Figs. 1–3 imply that levels of endogenous NMT in COS-1 cells are limiting for myristoylation of K7,9A-Fyn. To verify this supposition, the effect of varying the ratio of NMT cDNA to Fyn cDNA was tested. Myristoylation levels of K7,9A-Fyn increased twofold, using twice the amount of NMT cDNA and three- to fivefold using five times more NMT cDNA (Fig. 4 B). No significant effect on myristoylation of wild-type Fyn was observed (Fig. 4 A), implying that the amount of endogenous NMT was sufficient to achieve maximal myristoylation levels of this protein. Analysis of NMT expression by Western blotting confirmed that increasing levels of NMT protein were expressed after cotransfection with NMT cDNA (data not shown). Radiolabeling with Tran35S-label, followed by subcellular fractionation, showed that the distribution of wild-type Fyn between the soluble and the membrane fraction was not affected during coexpression of NMT (Fig. 4 C). In contrast, K7,9A-Fyn expressed in the presence of NMT displayed a significant shift from the soluble to the membrane fraction (Fig. 4 C). Taken together, these experiments demonstrate that the fatty acylation defect of the K7,9A-Fyn mutant can be restored by expressing excess amounts of exogenous NMT.
Coimmunoprecipitation of Fyn with the ζ Chain of the TCR Depends on the Fatty Acylation Status of Fyn
The ability to manipulate fatty acylation of Fyn provided us with a unique opportunity to quantitate the individual contributions of the fatty acids and the surrounding amino acids towards interactions of Fyn with other membrane bound proteins. We chose to evaluate the interactions of Fyn with the ζ chain of the TCR complex as a model system. COS-1 cells were cotransfected with cDNAs for Fyn and CD8-ζ, a chimeric construct containing the extracellular and transmembrane regions of CD8, fused to the cytoplasmic tail of the TCR ζ chain (16). Cells were lysed in a 1% digitonin lysis buffer (10, 33) and subjected to immunoprecipitation with anti–CD8 monoclonal antibody. Incubation with protein A/G+ agarose beads alone was used as a negative control. When anti–CD8 immunoprecipitates were analyzed by Western blotting using polyclonal anti– Fyn antibody, a signal for Fyn was obtained using lysates from cells coexpressing Fyn and CD8-ζ (Fig. 5 A, lane 3). Relative to the amount of wild-type Fyn, only marginal amounts (<5%) of G2A-Fyn and C3,6S-Fyn were detected after immunoprecipitation with anti–CD8 antiserum (Fig. 5, A, lanes 5 and 7, and B). Since G2A-Fyn is neither myristoylated nor palmitoylated and C3,6S-Fyn is myristoylated but not palmitoylated (1, 43), these data indicate that fatty acylation of Fyn with both myristate and palmitate is required for association of Fyn with CD8-ζ. K7,9A-Fyn was also impaired (∼70%) in its ability to coimmunoprecipitate with CD8-ζ (Fig. 5), a result similar to that obtained by Gauen et al. (11) for association of K7,9A-Fyn with a VSV G-ζ chimera. However, the level of coimmunoprecipitation of K7,9A-Fyn with CD8-ζ could be increased twofold by overexpression of exogenous NMT (Fig. 5, A, lane 11, and B) and this correlates well with the twofold increase in myristoylation levels (Fig. 4 B). This gain of function experiment indicates that the presence of lysine residues at positions 7 and 9 in Fyn is not required per se for an interaction with the ζ chain, but rather for efficient fatty acylation of Fyn. In addition, Gαo-Fyn (41) and Lck(10)Fyn, fusion proteins in which the first 10 amino acids of Fyn are replaced with those of human Gαo or human Lck (Table I) were analyzed. Both constructs are efficiently dually fatty acylated (see reference 41 and Table II), yet their NH2-terminal sequences are distinct from that of Fyn outside the met-gly-cys motif (Table I). As depicted in Fig. 5 B, the levels of Gαo-Fyn and Lck(10)Fyn coimmunoprecipitating with CD8-ζ were similar (∼90%) to that of wild-type Fyn (Fig. 5 B), providing two additional examples of the need for dual fatty acylation rather than specific lysine residues for interaction of Fyn with CD8-ζ. Controls indicated that expression levels of CD8-ζ and Fyn constructs were constant throughout the experiments (see Fig. 5 A).
Figure 5.

Coimmunoprecipita-tion of wild-type or mutant Fyn constructs with CD8-ζ. (A) COS-1 cells were cotransfected with cDNAs encoding wild-type or mutant Fyn (see Table I), CD8-ζ, and NMT where indicated. Cells were lysed in digitonin buffer and subjected to immunoprecipitation with anti– CD8 monoclonal antibody (+) or with protein A/G+ agarose beads alone (−). Immunoprecipitates were analyzed by SDS-PAGE and Western blotting using polyclonal anti–Fyn antibody (top). Subsequently, the blot was stripped and reprobed using anti–CD8 monoclonal antibody; the distribution of the 41-kD form of CD8-ζ is shown (middle). After immunoprecipitation with anti–CD8 antibody, lysates were subjected to a second immunoprecipitation step, using polyclonal anti–Fyn antibody, followed by SDS-PAGE and Western blotting using monoclonal anti–Fyn antibody (bottom). (B) Levels of Fyn protein coimmunoprecipitating with CD8-ζ were quantified by scanning of films. Films were exposed for different times to the blot and measurements were made in the linear range of the film. The measured values for coimmunoprecipitation with CD8-ζ were corrected for total expression levels and values are expressed relative to the amount of wild-type Fyn that coimmunoprecipitates with CD8-ζ. Given values represent the mean of two experiments.
Inhibition of Myristoylation, Membrane Binding, and CD8-ζ Association of Wild-Type Fyn by Treatment with 2-Hydroxymyristate
In an attempt to manipulate fatty acylation levels of wild-type Fyn, the effect of 2-hydroxymyristate, a potent inhibitor of NMT (25, 27) was tested. Transfected COS-1 cells expressing wild-type Fyn were treated with 2-hydroxymyristate and radiolabeled with either Tran35S-label or 125I-IC13. Analysis of immunoprecipitated Fyn after SDS-PAGE showed that the level of 35S-labeled protein was unaffected but that the level of 125I-IC13 labeling was significantly decreased (83%) after treatment with 2-hydroxymyristate (Fig. 6 A). Subsequently, analysis of the subcellular distribution of Fyn after treatment with 2-hydroxymyristate showed a dramatic increase in the amount of soluble Fyn. More than 95% of Fyn was recovered from the membrane P100 fraction of control cells, whereas >40% was observed in the soluble fraction after 2-hydroxymyristate treatment (Fig. 6 A). Next, the effect on coimmunoprecipitation of wild-type Fyn with CD8-ζ was analyzed. 2-Hydroxymyristate treatment caused a significant decrease in the amount of wild-type Fyn coimmunoprecipitating with CD8-ζ (Fig. 6 B), to <40% of control levels. After 2-hydroxymyristate treatment, wild-type Fyn still retains glycine 2, cysteine 3, and lysines 7 and 9, all of the residues previously implicated as part of an ITAM binding motif on Fyn, yet it cannot associate with ζ.
Figure 6.

Myristoylation, subcellular distribution, and coimmunoprecipitation of Fyn with CD8-ζ after 2-hydroxymyristate treatment. Transfected cells expressing Fyn were preincubated overnight with 500 μM 2-hydroxymyristate, as described in Materials and Methods. (A) Cells were radiolabeled in the absence (−) or presence (+) of 500 μM 2-hydroxymyristate with Tran35S-label, or 125I-IC13, and analyzed directly (left) or subjected to homogenization and subcellular fractionation, followed immunoprecipitation (right). (B) COS-1 cells were cotransfected with cDNAs for Fyn and CD8-ζ, and incubated in the absence (−) or the presence (+) of 2-hydroxymyristate. Cells were lysed in digitonin buffer and coimmunoprecipitation of Fyn with CD8-ζ was analyzed using anti–CD8 antibody (+) or protein A/G+ agarose beads alone (−).
Association of Fyn with ζ Chain: The NH2 Terminus of Fyn Alone Is Not Sufficient and Functional SH2- and SH1-kinase Domains Are Essential
To further define the nature of the observed association between Fyn and TCR ζ, the following series of experiments was performed. Fig. 7 A shows that between 10 and 15% of the total population of Fyn coimmunoprecipitated with CD8-ζ (compare lanes 7 and 11). Fyn protein was not detected in the absence of anti–CD8 antibody during immunoprecipitation (Figs. 5–7), showing that nonspecific binding of Fyn to the protein A/G+ agarose beads did not occur. Fyn was only detected in anti–CD8 immunoprecipitates from cells expressing both CD8-ζ and Fyn (Figs. 5 A, lane 3, 6 B, and 7 A, lane 7), but not from mock transfected cells (Fig. 7 A, lane 1), cells expressing CD8-ζ alone (Figs. 5 A, lane 1, and 7 A, lane 3), or Fyn alone (Fig. 7 A, lane 5). In addition, the mixing of digitonin lysates of cells that were transfected separately with either Fyn or CD8-ζ, followed by immunoprecipitation with anti–CD8-ζ antibodies, did not result in coimmunoprecipitation of Fyn (Fig. 7 A, lane 9), highlighting that Fyn and CD8-ζ need to be present within the same cell.
Figure 7.

Analysis of the molecular determinants in Fyn required for coimmunoprecipitation with CD8-ζ. (A) COS-1 cells were transfected with empty pCMV5 vector (lanes 1 and 2), cDNA encoding CD8-ζ alone (lanes 3 and 4), Fyn alone (lanes 5 and 6), or cotransfected with both CD8-ζ and Fyn (lanes 7, 8, 11, and 12). Cells were lysed in digitonin buffer and subjected to immunoprecipitation with (+) or without (−) monoclonal antibody against CD8 (lanes 1–10) or Fyn (lanes 11 and 12). In addition, immunoprecipitation was performed using a mixed digitonin lysate, derived from cells transfected separately with CD8-ζ or Fyn (lanes 9 and 10). Immunoprecipitates were analyzed by SDS-PAGE and Western blotting using polyclonal anti–Fyn antibody. (B) COS-1 cells were transfected with Fyn(16)GFP cDNA and either lysed in RIPA buffer and subjected to immunoprecipitation using anti–GFP antibody (Total) or fractionated into 1% Triton X-100 soluble (S) or resistant (R) fractions before immunoprecipitation (left). For coimmunoprecipitation analysis, COS-1 cells were transfected with Fyn(16)GFP and CD8-ζ cDNA, lysed in digitonin buffer, and subjected to immunoprecipitation with (+) or without (−) anti–GFP or anti–CD8 antibody (right). Immunoprecipitates were analyzed by SDS-PAGE and Western blotting using polyclonal anti–GFP. (C) COS-1 cells were transfected with cDNAs encoding CD8-ζ and either wild-type Fyn, K299M-Fyn, ΔSH2-Fyn, or R176K-Fyn. Digitonin lysates were subjected to immunoprecipitation with anti–CD8 antibody (left) or anti–Fyn monoclonal antibody (right) and immunoprecipitates were analyzed by SDS-PAGE and Western blotting with anti–Fyn antibody. The locations of full length Fyn and ΔSH2-Fyn are indicated by the arrows. The weak signal in the K299M-Fyn lane (left) may be accounted for by phosphorylation of CD8-ζ by endogenous Fyn (see Fig. 8 A, legend).
Next, the functional requirements within the Fyn protein structure for association with CD8-ζ were evaluated. We first tested whether the Fyn NH2 terminus alone is sufficient for interaction with the ζ chain in cells, using Fyn(16)GFP, a chimeric construct containing the first 16 amino acids of Fyn fused to GFP. Fyn(16)GFP expressed in COS-1 cells was dually acylated with myristate and palmitate (data not shown), and after extraction with nonionic detergent buffer, ∼60% of total Fyn(16)GFP was recovered from the detergent insoluble fraction (Fig. 7 B). Thus, Fyn(16)GFP is processed and targeted like wild-type Fyn and other chimeras containing the Fyn SH4 motif (41, 43). After coexpression of Fyn(16)GFP and CD8-ζ in COS cells, followed by digitonin lysis, no Fyn(16)GFP was detected in anti–CD8 immunoprecipitates, although there was a strong signal after immunoprecipitation with anti– GFP antibody (Fig. 7 B). Similar results were obtained with constructs containing the NH2 terminus of Fyn fused to GST or β galactosidase (data not shown). These results indicate that the NH2-terminal SH4 motif of Fyn by itself is not sufficient for association with CD8-ζ and that additional domains of the Fyn protein are required. To identify these requirements, CD8-ζ was coexpressed with three different Fyn mutants: K299M-Fyn carries a point mutation in the SH1 domain that renders Fyn kinase-inactive, ΔSH2-Fyn lacks amino acids 144–248 encoding the Fyn SH2 domain, and R176K-Fyn contains a point mutation of a critical arginine residue in the SH2 domains of Src family kinases that reduces phosphotyrosine binding (26). Less than 15% of ΔSH2-Fyn and only ∼35% of K299M-Fyn and R176K-Fyn was detected in anti–CD8 immunoprecipitates (Fig. 7 C, left, and Table III). The expression levels were similar for all constructs (Fig. 7 C, right). These findings establish that functional SH1 and SH2 domains in Fyn are important for optimal interaction with CD8-ζ.
Table III.
Quantitative Analysis of Coimmunoprecipitation of Fyn with CD8-ζ
| Construct | Relative coimmunoprecipitation | ||||
|---|---|---|---|---|---|
| CD8-ζ | + Fyn | 100% | |||
| + K299M-Fyn | 35% | ||||
| + ΔSH2-Fyn | 14% | ||||
| + R176K-Fyn | 38% | ||||
| CD8-ζT76 | + Fyn | 91% | |||
| CD8-ζ4F | + Fyn | 19% | |||
| CD8-ζ + Fyn | (Octylglucoside) | 14% | |||
| (Filipin) | 54% | ||||
| CD8-ζ | + FynKRas | 93% | |||
| + G2A,C3S-FynKRas | 26% | ||||
COS-1 cells were transfected with cDNAs encoding the indicated constructs, and coimmunoprecipitation experiments were performed as described in Fig. 5. In each experiment, coimmunoprecipitation of Fyn protein constract with anti–CD8 antibody was measured in duplicate and multiple films representing different exposure times to the blot were used to ensure linarity of the signal. These values were first corrected for the total level of Fyn protein in each immunoprecipitation, as measured by the second Fyn immunoprecipitation step (see Materials and Methods). Given numbers for each test are expressed relative to the amount of wild-type Fyn that coimmunoprecipitates with full length CD8-ζ and represent the mean of two experiments, with errors ranging from 5 to 15%. The given 100% value of wild-type Fyn coimmunoprecipitating with full length CD8-ζ represents 10–15% of total Fyn expressed (see Fig. 7).
Association of Fyn with ζ Chain Requires Phosphorylation of ζ ITAMs by Fyn
Based on the findings described above that kinase activity and the SH2 domain in Fyn are critical, we addressed the relevance of the ITAM motifs in the ζ chain for association with Fyn. First, the tyrosine phosphorylation status of CD8-ζ was tested by immunoprecipitation with anti–CD8 antibody followed by antiphosphotyrosine Western blotting analysis. As shown in Fig. 8 A, CD8-ζ was strongly tyrosine phosphorylated when coexpressed with Fyn, ΔSH2-Fyn, or R176K-Fyn, but not when expressed alone or when coexpressed with kinase dead K299M-Fyn. Levels of CD8-ζ protein were constant (Fig. 8 A, bottom). As observed elsewhere (18), CD8-ζ is present in three forms in COS-1 cells, migrating at 36, 38, and 41 kD, respectively (Fig. 8 A, arrows). Comparison of the migration profiles indicated that the 41-kD band and the smear above it represent the tyrosine phosphorylated forms of CD8-ζ (Fig. 8 A, top arrow). Next, the specificity of ζ chain ITAM phosphorylation by Fyn was analyzed by use of a CD8-ζ construct in which the cytoplasmic tail was truncated after the second ITAM motif (CD8-ζT76), and a CD8-ζT76 construct in which all four remaining ITAM tyrosines were mutated to phenylalanine (CD8-ζ4F) (22). As measured by radiolabeling and immunoprecipitation, the total levels of protein expression in COS-1 cells were similar for each construct (Fig. 8 B, bottom). Truncated CD8-ζT76 was observed as one main form that migrated at ∼40 kD and the major radiolabeled form of CD8-ζ4F was observed to migrate at ∼35 kD, as indicated by the arrows. The 40-kD band of CD8-ζT76 was tyrosine phosphorylated when coexpressed with Fyn, with moderately reduced levels compared with full length CD8-ζ, whereas no tyrosine phosphorylation was observed for the CD8-ζ4F mutant (Fig. 8 B). Analysis of the association of Fyn with the different CD8-ζ constructs showed that similar amounts (∼90%) of Fyn coimmunoprecipitated with CD8-ζT76, whereas very little (<20%) Fyn was observed in the CD8-ζ4F immunoprecipitate (Fig. 8 C, and Table III). These results show that phosphorylation of ITAM tyrosines in the ζ chain by Fyn is critical for association between Fyn and ζ. A similar finding was reported by Pleiman et al. (29), who showed that the SH2 domains of Fyn and Lyn bind to the tyrosine phosphorylated ITAM in the Igα receptor.
Figure 8.

Analysis of the molecular interactions between Fyn and CD8-ζ. (A) COS-1 cells were transfected with cDNAs encoding CD8-ζ and either pCMV5, wild-type Fyn, K299M-Fyn, ΔSH2-Fyn, or R176K-Fyn. Transfected cells were lysed in RIPA buffer and were subjected to immunoprecipitation with anti– CD8 antibody, followed by Western blotting analysis using antiphosphotyrosine (α-PTyr) monoclonal antibody (top). As controls for expression levels, transfected cells were radiolabeled with Tran35S-label (bottom), lysed in RIPA buffer, and subjected to immunoprecipitation with anti–CD8 antibody, followed by SDS-PAGE and autoradiography. CD8-ζ expressed in COS-1 cells is present in three forms that migrate at 36, 38, and 41 kD, respectively, as indicated by the arrows. The 41-kD form represents the main tyrosine phosphorylated band. Overexposure of the blot in the top row revealed a low level of phosphorylation of CD8-ζ in the pCMV5 and K299M lanes, presumably reflecting the activity of endogenous Fyn or other kinases. (B) COS-1 cells were transfected with cDNAs encoding wild-type Fyn and either CD8-ζ, CD8-ζT76, or CD8-ζ4F. Antiphosphotyrosine Western blotting and radiolabeling were performed as described in A. Truncated CD8-ζT76 was present in one main form that migrated at ∼40 kD while the main radiolabeled form of CD8-ζ4F migrated at ∼35 kD, as indicated by the arrows. (C) COS-1 cells were transfected as described in B and coimmunoprecipitation analysis of Fyn with CD8-ζ constructs was performed as described in Fig. 7 C, legend.
Association of Fyn with CD8-ζ Is Stabilized in Membrane Rafts
Dual fatty acylation has previously been shown to be required for localization of Fyn to plasma membrane microdomains known as rafts (5, 35, 38, 41, 43). To test for the involvement of rafts in directing the interactions between Fyn and ζ chain, two different approaches were used. First, we analyzed the sensitivity of coimmunoprecipitation of Fyn with CD8-ζ to treatment with octylglucoside and filipin, agents that disrupt raft stability (5, 38). The association of Fyn with CD8-ζ was almost entirely abolished by addition of octylglucoside to the digitonin lysis buffer (Table III). The interaction was also greatly reduced (∼50%) by pretreatment of cells with filipin (Table III), an agent that binds cellular cholesterol, an essential component for raft integrity (5, 38). An alternate approach was based on the recent observation that, in contrast to fatty acylated proteins, farnesylated proteins are largely excluded from detergent-resistant membrane rafts (23). Fyn was tagged at the carboxy terminus with the KRas4B tail (FynKRas), which promotes farnesylation and plasma membrane targeting (15). This construct is both fatty acylated and prenylated (data not shown). A second construct containing only a functional farnesylation signal was generated by mutating the amino-terminal fatty acylation sites (G2A,C3S-FynKRas). When expressed in COS-1 cells, wild-type Fyn and the two KRas-tagged constructs were completely membrane bound (Fig. 9 A). Wild-type Fyn and FynKRas were also enriched in the Triton X-100 insoluble fraction (Fig. 9 B). In contrast, the amount of G2A,C3S-FynKRas in the Triton X-100 insoluble fraction was significantly lower (Fig. 9 B), suggesting that farnesylated Fyn exhibits reduced association with plasma membrane rafts. Fig. 9 C and Table III show that coimmunoprecipitation of G2A,C3S-FynKRas with CD8-ζ was significantly reduced, whereas association of FynKRas with CD8-ζ was very similar to that of wild-type Fyn. FynKRas and G2A,C3S-FynKRas kinase activity and ability to phosphorylate ζ ITAM tyrosines were similar to wild-type Fyn (data not shown). These observations imply that fatty acylation not only provides plasma membrane binding of Fyn, but also guides specific localization to membrane subdomains, which is essential for stabilization of the protein–protein interactions between Fyn and the ζ chain.
Figure 9.

Membrane binding, Triton X-100 sensitivity, and coimmunoprecipitation of fatty acylated and farnesylated Fyn constructs. (A) COS-1 cells were transfected with cDNAs encoding wild-type Fyn, FynKRas, or G2A,C3S-FynKRas. Transfected cells were radiolabeled with Tran35S-label, homogenized, and fractionated as described in Materials and Methods. Distribution into the P100 fraction was quantitated by phosphorimaging. Values represent the mean of two experiments. (B) COS-1 cells were transfected as described in A, followed by fractionation into 1% Triton X-100 soluble (S) or resistant (R) fractions. Distribution of Fyn constructs into the TX100 resistant fraction was quantitated after immunoprecipitation by Western blotting analysis, as described in Materials and Methods. Values represent the mean of two experiments. (C) COS-1 cells were transfected as described in A and coimmunoprecipitation analysis of Fyn protein with CD8-ζ was performed as described in Fig. 7 C, legend.
Discussion
Lysines 7 and 9 Are Required for Efficient N-Myristoylation of Fyn
Lysine residues at positions 7 and 9 are conserved in nearly all Src family members (36, 37), and, in the context of the NH2 terminus of v-Src, the lysine at position 7 is critical for efficient myristoylation (6, 19). We were therefore interested in testing the fatty acylation status of K7,9A-Fyn, a mutant containing alanine substitutions for lysines 7 and 9. To obtain sensitive measurements of myristoylation levels, different Fyn constructs (Table I) were expressed at high levels in COS cells, and radiolabeling with the iodinated myristate analogue 125I-IC13 was performed. After immunoprecipitation and SDS-PAGE, quantitation of 125I-IC13 incorporation into Fyn was accomplished by PhosphorImaging (2), which is not feasible after radiolabeling with conventional 3H-labeled myristate analogues used in other studies (11). Since all Fyn constructs used in this report contain the same number of methionines and cysteines, labeling with Tran35S-label in parallel gave precise numbers for expression levels of the protein backbone. Subsequent calculation of 125I-IC13 per unit of 35S-labeled protein provided a reliable measure of the efficiency of myristoylation for wild-type and mutant Fyn constructs.
Our results show that, as previously established for v-Src (6, 19), lysine 7 is required for efficient myristoylation of Fyn (Table II). In addition, we show that lysine 9 plays an important role in guiding effective myristoylation of Fyn (Table II). Interestingly, the effects of the individual lysine replacements add up to the total effect observed with the double K7,9A-Fyn mutant (Fig. 1, and Table II), suggesting that each lysine residue plays an independent role. The conclusion that these lysines are directly involved in a myristoylation event is further substantiated by our finding that the K7,9A-Fyn myristoylation defect is specifically reversed by coexpression with wild-type human NMT, but not with enzymatically impaired H218N-NMT (28) (Figs. 3 and 4).
Reversal of the K7,9A-Fyn myristoylation defect by additional exogenous NMT in our experiments was almost complete (Fig. 4 B). In the experiments described here, we have used cDNA encoding a 49-kD form of human NMT (28), which has full enzymatic activity in vitro and is capable of functionally complementing myristoylation deficiency in yeast (9). Additional cellular forms of NMT, with higher molecular weights (∼60 kD) and different intracellular distribution patterns, have been observed (14), and recently a second mammalian NMT was characterized (13). Most notably, missing from the 49-kD form of NMT (9, 28) is an NH2-terminal sequence containing a polybasic motif that is proposed to function in ribosomal targeting of NMT (14), as well as other cotranslationally active enzymes, such as N-methionylaminopeptidases (21). We have observed that neither the 60-kD form nor the NMT 1 or 2 isoforms (13) reverse the myristoylation defect in K7,9A-Fyn more efficiently than the 49-kD form (data not shown).
Lck is the only Src family member that lacks lysines at positions 7 and 9 (36, 37) and one might question whether myristoylation of Lck is impaired relative to the other Src-related kinases. Since quantitation of fatty acylation of full length Lck proved technically difficult, we used an Lck(10)Fyn fusion construct (Table I). Our results show that the fatty acylation profile of the Lck amino terminus is very similar to that of Fyn (Table II), despite the absence of lysines 7 and 9. We infer that in Lck the serine at position 6 (Table I), a preferred residue at this site for recognition by mammalian NMT (17) is sufficient to direct efficient fatty acylation of this sequence. If myristoylation levels of full length Lck were substoichiometric, one would expect that either its membrane association would not be complete or that additional factors (e.g., CD4) would be required to guide membrane binding of Lck. However, nearly all of the Lck in cells appears to be membrane bound (18, 46). Moreover, two recent studies have shown that plasma membrane targeting of Lck can be achieved in the absence of CD4 and that the first 10 amino acids of Lck are sufficient for plasma membrane binding (4, 46). These experiments imply that myristoylation and palmitoylation of full length Lck occur efficiently.
The NH2 Terminus of Fyn Directs Fatty Acylation and Membrane Localization, but Does Not Specify an ITAM Binding Motif
Several earlier reports have described coimmunoprecipitation of the thymic isoform of Fyn with the TCR/CD3 complex (10, 12, 33). The association was only observed under mild detergent conditions and the exact nature of the interaction remained unclear. Recently, the first 10 amino acids of Fyn were shown to be essential for specific binding of Fyn to the ζ subunit, as well as other chains of the TCR complex in heterologous systems (10). On the basis of mutagenesis experiments, it was concluded that the amino acids glycine 2, cysteine 3, and lysines 7 and 9 define a motif for binding to ITAMs and for plasma membrane localization (11).
Our results described here (Figs. 1–4), as well as earlier work from our group and others (1, 35, 41, 43), indicate that these amino acids within the Fyn SH4 domain are essential for optimal fatty acylation and membrane binding of Fyn. Using the coimmunoprecipitation of various Fyn constructs with a CD8-ζ fusion protein (16) as a model, we present three lines of evidence to demonstrate that the Fyn SH4 domain does not represent a specific ITAM binding motif.
First, we show that the reduced coimmunoprecipitation of the K7,9A-Fyn mutant with CD8-ζ (Fig. 5, and reference 11) is specifically reversed by overexpression of NMT (Fig. 5). The effect of NMT overexpression is to increase fatty acylation and membrane binding of K7,9A-Fyn (Figs. 3 and 4). This gain of function experiment clearly establishes that coimmunoprecipitation of Fyn with CD8-ζ does not require lysines 7 and 9 per se, but their presence is crucial for directing efficient fatty acylation. This conclusion is further substantiated by the observations made with the Gαo-Fyn and Lck(10)Fyn constructs, which coimmunoprecipitate with CD8-ζ as efficiently as wild-type Fyn (Fig. 5 B). The NH2-terminal sequences of Gαo-Fyn and Lck(10)Fyn do not contain lysines at position 7 and 9 (Table I), but they are efficiently fatty acylated (reference 41, and Table I). Second, the loss of function experiment with 2-hydroxymyristate (Fig. 6) shows that, even in the presence of all the amino acids proposed to constitute the ITAM binding motif (glycine 2, cysteine 3, and lysines 7 and 9), coimmunoprecipitation of wild-type Fyn with CD8-ζ can be ablated. Thus, loss of association of wild-type Fyn to CD8-ζ coincides with a reduction of fatty acylation and membrane binding (Fig. 6). Third, Fyn(16)GFP did not coimmunoprecipitate with CD8-ζ (Fig. 7 B), showing that the first 16 amino acids of Fyn are not sufficient for interaction with the ITAMs on ζ, in agreement with the results of cross-linking studies (34). This implies that other protein determinants in Fyn are involved in mediating the interactions with ζ chain. Indeed, coimmunoprecipitation of Fyn was observed to require both a functional kinase as well as an SH2 domain (Fig. 7 C).
Colocalization of Fyn and ζ Chain in Cell Surface Rafts
Coimmunoprecipitation of Fyn with TCR subunits occurs only under mild detergent conditions (10, 12, 33), which has been taken as evidence for low affinity protein–protein interactions. Our findings with Fyn, and a recently growing interest in membrane microdomains (5, 38), suggests the involvement of additional mechanisms for stabilization of the interactions between Src kinases and TCR subunits, which is supported by several recent independent reports (24, 44, 45). In biological membranes, especially plasma membranes, phase-separated “rafts” exist, representing lipid domains enriched in cholesterol and sphingolipids that provide scaffolds for clustering of transmembrane and peripherally associated proteins (5, 38). These rafts or microdomains can be recovered from cells using mild detergent conditions (5). There is growing evidence that key components of TCR-mediated signaling events are localized to rafts. For example, palmitoylation of the T cell transmembrane protein LAT is essential for localization in rafts and T cell tyrosine phosphorylation events (45). Src family kinases, especially Fyn and Lck, are constitutively localized in rafts via dual fatty acylation motifs (35, 41, 43). Activation of the TCR results in specific accumulation of ζ chains in plasma membrane rafts (24). In COS cells, ∼20% of CD8-ζ is in the detergent-insoluble fraction (data not shown), similar to the distribution of the ζ chain in resting T cells (7), and this may explain in part why only a limited amount (<20%) of total Fyn coimmunoprecipitates with CD8-ζ in COS cells. The functional significance of raft localization is evidenced by the finding that disruption of cell surface rafts interferes with early T cell signaling events (44). These observations establish a connection between local accumulation of signaling molecules in plasma membrane rafts and regulated T cell signaling events.
Based on the results reported here and by others (35, 39, 41, 43), the following model emerges. Myristoylation and palmitoylation within the NH2-terminal SH4 domain directs Fyn into detergent-resistant plasma membrane rafts and positions the downstream SH2 and kinase domains in close proximity to the TCR ζ chain. Phosphorylation of the ζ ITAMs by Fyn (Fig. 8) allows subsequent interaction between the phosphorylated ITAMs and the Fyn SH2 domain (Figs. 7 and 8). Accumulation of Fyn and the ζ chain in rafts increases the local concentration of both proteins and drives their association. The stability and the extent of Fyn SH2/ζ phosphotyrosine interaction is apparently dependent on the integrity of the rafts since treatment with octylglucoside and filipin, agents that disrupt rafts (5, 38), results in dissociation of the Fyn/ζ complex (Table III), presumably due to a decrease of the effective local protein concentrations. This implies that the Fyn SH2/ζ phosphotyrosine interaction is relatively weak, as found for several other SH2/phosphotyrosine interactions (20). In T cells, dissociation of the Fyn SH2 domain from the ζ chain would make ITAM phosphotyrosine residues on ζ accessible for binding to the SH2 domains of other signaling molecules, such as ZAP-70. This model explains why undermyristoylated and underpalmitoylated forms of Fyn, that are not efficiently membrane bound, as well as a farnesylated Fyn construct that is not enriched in membrane subdomains (Fig. 9), do not interact well with the ζ chain. These studies illustrate that the role of the NH2-terminal SH4 motif is to properly position Src kinases within specific locations of cellular membranes for optimal interaction of downstream Src homology domains with other membrane-bound signaling molecules.
Acknowledgments
We thank Dr. Art Weiss for his generous gifts of CD8-ζ plasmids. We are grateful to Raya Louft-Nisenbaum for technical assistance and Lori Klausner for secretarial support.
This work was supported by grant GM 57966 from the National Institutes of Health and grant BE235 from the American Cancer Society to M.D. Resh. M.D. Resh is an Established Scientist of the American Heart Association. W. van't Hof is a Miriam and Benedict Wolf Cancer Research fellow.
Abbreviations used in this paper
- GFP
green fluorescent protein
- ITAM
immune-receptor tyrosine-based activation motif
- NMT
N-myristoyl transferase
- SH
Src homology domain
- TCR
T cell receptor
Footnotes
Dr. van't Hof's present address is Pulmonary Research Laboratories, Division of Pulmonary and Critical Care Medicine, Weill Medical College of Cornell University, 1300 York Ave., F-401, New York, NY 10021.
References
- 1.Alland L, Peseckis SM, Atherton RE, Berthiaume L, Resh MD. Dual myristylation and palmitylation of Src family member p59fynaffects subcellular localization. J Biol Chem. 1994;269:16701–16705. [PubMed] [Google Scholar]
- 2.Berthiaume L, Peseckis SM, Resh MD. Synthesis and use of Iodo-fatty acid analogs. Methods Enzymol. 1995;250:454–466. doi: 10.1016/0076-6879(95)50090-1. [DOI] [PubMed] [Google Scholar]
- 3.Berthiaume L, Resh MD. Biochemical characterization of a palmitoyl acyltransferase activity that palmitoylates myristylated proteins. J Biol Chem. 1995;270:22399–22405. doi: 10.1074/jbc.270.38.22399. [DOI] [PubMed] [Google Scholar]
- 4.Bijlmakers MJ, Isobe-Nakamura M, Ruddock LJ, Marsh M. Intrinsic signals in the unique domain target p56(lck) to the plasma membrane independently of CD4. J Cell Biol. 1997;137:1029–1040. doi: 10.1083/jcb.137.5.1029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Brown DA, London E. Functions of lipid rafts in biological membranes. Annu Rev Cell Dev Biol. 1998;14:111–136. doi: 10.1146/annurev.cellbio.14.1.111. [DOI] [PubMed] [Google Scholar]
- 6.Buss JE, Der CJ, Solski PA. The six amino-terminal amino acids of p60src are sufficient to cause myristylation of p21v-ras . Mol Cell Biol. 1988;8:3960–3963. doi: 10.1128/mcb.8.9.3960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Caplan S, Zeliger S, Wang L, Baniyash M. Cell-surface– expressed T-cell antigen-receptor zeta chain is associated with the cytoskeleton. Proc Natl Acad Sci USA. 1995;92:4768–4772. doi: 10.1073/pnas.92.11.4768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Deichaite I, Casson LP, Ling H-P, Resh MD. In vitro synthesis of pp60v-src: myristylation in a cell-free system. Mol Cell Biol. 1988;8:4295–4301. doi: 10.1128/mcb.8.10.4295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Duriono RJ, Reed SI, Gordon JI. Mutations of human myristoyl-CoA: protein N-myristoyltransferase cause temperature-sensitive myristic acid auxotrophy in Saccharomyces cerevisiae. . Proc Natl Acad Sci USA. 1992;89:4129–4133. doi: 10.1073/pnas.89.9.4129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gauen LKT, Kong A-NT, Samelson LE, Shaw AS. p59fyn tyrosine kinase associates with multiple T-cell receptor subunits through its unique amino-terminal domain. Mol Cell Biol. 1992;12:5438–5446. doi: 10.1128/mcb.12.12.5438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gauen LKT, Linder ME, Shaw AS. Multiple features of the p59fyn src homology 4 domain define a motif for immune-receptor tyrosine-based activation motif (ITAM) binding and for plasma membrane localization. J Cell Biol. 1996;133:1007–1015. doi: 10.1083/jcb.133.5.1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gauen LKT, Zhu Y, Letourneur F, Hu Q, Bolen JB, Matis LA, Klausner RD, Shaw AS. Interactions of p59fynwith T-cell receptor activation motifs: defining the nature of a signalling motif. Mol Cell Biol. 1994;14:3729–3741. doi: 10.1128/mcb.14.6.3729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Giang DK, Cravatt BF. A second mammalian N-myristoyltransferase. J Biol Chem. 1998;273:6595–6598. doi: 10.1074/jbc.273.12.6595. [DOI] [PubMed] [Google Scholar]
- 14.Glover CJ, Hartman KD, Felsted RL. Human N-myristoyltransferase amino-terminal domain involved in targeting the enzyme to the ribosomal subcellular fraction. J Biol Chem. 1997;272:28680–28689. doi: 10.1074/jbc.272.45.28680. [DOI] [PubMed] [Google Scholar]
- 15.Hancock JF, Paterson H, Marshall CJ. A polybasic domain or palmitoylation is required in addition to the CAAX motif to localize p21ras to the plasma membrane. Cell. 1990;63:133–139. doi: 10.1016/0092-8674(90)90294-o. [DOI] [PubMed] [Google Scholar]
- 16.Irving BA, Weiss A. The cytoplasmic domain of the T cell receptor zeta chain is sufficient to couple to receptor-associated signal transduction pathways. Cell. 1991;64:891–901. doi: 10.1016/0092-8674(91)90314-o. [DOI] [PubMed] [Google Scholar]
- 17.Johnson DR, Bhatnagar RS, Knoll LJ, Gordon JI. Genetic and biochemical studies of protein N-myristoylation. Annu Rev Biochem. 1994;63:869–914. doi: 10.1146/annurev.bi.63.070194.004253. [DOI] [PubMed] [Google Scholar]
- 18.Kabouridis PS, Magee AI, Ley SC. S-acylation of LCK protein tyrosine kinase is essential for its signalling function in T lymphocytes. EMBO (Eur Mol Biol Organ) J. 1997;16:4983–4998. doi: 10.1093/emboj/16.16.4983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kaplan JM, Mardon G, Bishop JM, Varmus HE. The first seven amino acids encoded by the v-src oncogene act as a myristylation signal: lysine 7 is a critical determinant. Mol Cell Biol. 1988;8:2435–2441. doi: 10.1128/mcb.8.6.2435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ladbury JE, Lemmon MA, Zhou M, Green J, Botfield MC, Schlessinger J. Measurement of the binding of tyrosyl phosphopeptides to SH2 domains: a reappraisal. Proc Natl Acad Sci USA. 1995;92:3199–3203. doi: 10.1073/pnas.92.8.3199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Li X, Chang Y-H. Amino-terminal protein processing in Saccharomyces cerevisiaeis an essential function that requires two distinct methionine aminopeptidases. Proc Natl Acad Sci USA. 1995;92:12357–12361. doi: 10.1073/pnas.92.26.12357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lowin-Kropf B, Smith V, Shapiro, Weiss A. Cytoskeletal polarization of T cells is regulated by an immunoreceptor tyrosine-based activation motif–dependent mechanism. J Cell Biol. 1998;140:861–871. doi: 10.1083/jcb.140.4.861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Melkonian KA, Ostermeyer AG, Chen JZ, Roth MG, Brown DA. Role of lipid modifications in targeting proteins to detergent-resistant membrane rafts. J Biol Chem. 1999;274:3910–3917. doi: 10.1074/jbc.274.6.3910. [DOI] [PubMed] [Google Scholar]
- 24.Montixi C, Langlet C, Bernard AM, Thimonier J, Dubois C, Wurbel MA, Chauvin JP, Pierres M, He HT. Engagement of T cell receptor triggers its recruitment to low-density detergent-insoluble membrane domains. EMBO (Eur Mol Biol Organ) J. 1998;17:5334–5348. doi: 10.1093/emboj/17.18.5334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nadler MJS, Harrison ML, Ashendel CL, Cassady JM, Gaehlen RL. Treatment of T cells with 2-hydroxymyristic acid inhibits the myristoylation and alters the stability of p56lck . Biochemistry. 1993;32:9250–9255. doi: 10.1021/bi00086a034. [DOI] [PubMed] [Google Scholar]
- 26.Okamura H, Resh MD. p80/85 Cortactin associates with the Src SH2 domain and colocalizes with v-Src in transformed cells. J Biol Chem. 1995;270:26613–26618. doi: 10.1074/jbc.270.44.26613. [DOI] [PubMed] [Google Scholar]
- 27.Paige LA, Zheng G, DeFrees SA, Cassady JM, Gaehlen RJ. Metabolic activation of 2-substituted derivates of myrisitic acid to form potent inhibitors of myristoyl CoA:protein N-myristoyltransferase. Biochemistry. 1990;29:10566–10573. doi: 10.1021/bi00498a021. [DOI] [PubMed] [Google Scholar]
- 28.Peseckis SM, Resh MD. Fatty acyl transfer by human N-myristyl transferase is dependent upon conserved cysteine and histidine residues. J Biol Chem. 1994;269:30888–30892. [PubMed] [Google Scholar]
- 29.Pleiman CM, Abrams C, Gauen LTK, Bedzyk W, Jongstra J, Shaw AS, Cambier JC. Distinct p53/56lyn and p59fyndomains associate with nonphosphorylated and phosphorylated Ig-α. Proc Natl Acad Sci USA. 1994;91:4268–4272. doi: 10.1073/pnas.91.10.4268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Resh MD. Interaction of tyrosine kinase oncoproteins with cellular membranes. Biochim Biophys Acta. 1993;1155:307–322. doi: 10.1016/0304-419x(93)90012-2. [DOI] [PubMed] [Google Scholar]
- 31.Resh MD. Myristylation and palmitylation of Src family members: the fats of the matter. Cell. 1994;76:411–413. doi: 10.1016/0092-8674(94)90104-x. [DOI] [PubMed] [Google Scholar]
- 32.Resh MD. Regulation of cellular signaling by fatty acid acylation and prenylation of signal transduction proteins. Cell Signalling. 1996;8:403–412. doi: 10.1016/s0898-6568(96)00088-5. [DOI] [PubMed] [Google Scholar]
- 33.Samelson LE, Phillips AF, Luong ET, Klausner RD. Association of the Fyn protein tyrosine kinase with the T cell antigen receptor. Proc Natl Acad Sci USA. 1990;87:4358–4362. doi: 10.1073/pnas.87.11.4358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sarosi GA, Thomas PM, Egerton M, Phillips AF, Kim KW, Bonvini E, Samelson LE. Characterization of the T cell antigen receptor p60fyn protein tyrosine kinase association by chemical cross-linking. Int Immunol. 1992;4:1211–1217. doi: 10.1093/intimm/4.11.1211. [DOI] [PubMed] [Google Scholar]
- 35.Shenoy-Scaria AM, Dietzen DJ, Kwong J, Link DC, Lublin DM. Cysteine 3 of Src family protein tyrosine kinases determines palmitoylation and localization in caveolae. J Cell Biol. 1994;126:353–363. doi: 10.1083/jcb.126.2.353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Silverman L, Resh MD. Lysine residues form an integral component of a novel NH2-terminal membrane targeting motif for myristylated pp60v-src . J Cell Biol. 1992;119:415–422. doi: 10.1083/jcb.119.2.415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Silverman L, Sudol M, Resh MD. Members of the src family of nonreceptor tyrosine kinases share a common mechanism for membrane binding. Cell Growth Differ. 1993;4:475–482. [PubMed] [Google Scholar]
- 38.Simons K, Ikonen E. Functional rafts in cell membranes. Nature. 1997;387:569–572. doi: 10.1038/42408. [DOI] [PubMed] [Google Scholar]
- 39.Thomas PM, Samelson LE. The glycophosphatidylinositol-anchored Thy-1 molecule interacts with the p60fyn protein tyrosine kinase in T cells. J Biol Chem. 1992;267:12317–12322. [PubMed] [Google Scholar]
- 40.Thomas SM, Brugge JS. Cellular functions regulated by Src family kinases. Annu Rev Cell Dev Biol. 1997;13:513–609. doi: 10.1146/annurev.cellbio.13.1.513. [DOI] [PubMed] [Google Scholar]
- 41.van't Hof W, Resh MD. Rapid plasma membrane anchoring of newly synthesized p59Fyn: selective requirement for NH2-terminal myristoylation and palmitoylation at cysteine-3. J Cell Biol. 1997;136:1023–1035. doi: 10.1083/jcb.136.5.1023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wilcox C, Hu J-S, Olson EN. Acylation of proteins with myristic acid occurs cotranslationally. Science. 1987;238:1275–1278. doi: 10.1126/science.3685978. [DOI] [PubMed] [Google Scholar]
- 43.Wolven A, Okamura H, Rosenblatt Y, Resh MD. Palmitoylation of p59fynis reversible and sufficient for plasma membrane association. Mol Biol Cell. 1997;8:1159–1173. doi: 10.1091/mbc.8.6.1159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Xavier R, Brennan T, Li Q, McCormack C, Seed B. Membrane compartmentation is required for efficient T cell activation. Immunity. 1998;8:723–732. doi: 10.1016/s1074-7613(00)80577-4. [DOI] [PubMed] [Google Scholar]
- 45.Zhang W, Trible RP, Samelson LE. LAT palmitoylation: its essential role in membrane microdomain targeting and tyrosine phosphorylation during T cell activation. Immunity. 1998;9:239–246. doi: 10.1016/s1074-7613(00)80606-8. [DOI] [PubMed] [Google Scholar]
- 46.Zlatkine P, Mehul B, Magee AI. Retargeting of cytosolic proteins to the plasma membrane by the Lck protein tyrosine kinase dual acylation motif. J Cell Sci. 1997;110:673–679. doi: 10.1242/jcs.110.5.673. [DOI] [PubMed] [Google Scholar]