

Abstract
ZO-1, a 220-kD peripheral membrane protein consisting of an amino-terminal half discs large (dlg)-like domain and a carboxyl-terminal half domain, is concentrated at the cadherin-based cell adhesion sites in non-epithelial cells. We introduced cDNAs encoding the full-length ZO-1, its amino-terminal half (N-ZO-1), and carboxyl-terminal half (C-ZO-1) into mouse L fibroblasts expressing exogenous E-cadherin (EL cells). The full-length ZO-1 as well as N-ZO-1 were concentrated at cadherin-based cell–cell adhesion sites. In good agreement with these observations, N-ZO-1 was specifically coimmunoprecipitated from EL transfectants expressing N-ZO-1 (NZ-EL cells) with the E-cadherin/α, β catenin complex. In contrast, C-ZO-1 was localized along actin stress fibers. To examine the molecular basis of the behavior of these truncated ZO-1 molecules, N-ZO-1 and C-ZO-1 were produced in insect Sf9 cells by recombinant baculovirus infection, and their direct binding ability to the cadherin/catenin complex and the actin-based cytoskeleton, respectively, were examined in vitro. Recombinant N-ZO-1 bound directly to the glutathione-S-transferase fusion protein with α catenin, but not to that with β catenin or the cytoplasmic domain of E-cadherin. The dissociation constant between N-ZO-1 and α catenin was ∼0.5 nM. On the other hand, recombinant C-ZO-1 was specifically cosedimented with actin filaments in vitro with a dissociation constant of ∼10 nM. Finally, we compared the cadherin-based cell adhesion activity of NZ-EL cells with that of parent EL cells. Cell aggregation assay revealed no significant differences among these cells, but the cadherin-dependent intercellular motility, i.e., the cell movement in a confluent monolayer, was significantly suppressed in NZ-EL cells. We conclude that in nonepithelial cells, ZO-1 works as a cross-linker between cadherin/catenin complex and the actin-based cytoskeleton through direct interaction with α catenin and actin filaments at its amino- and carboxyl-terminal halves, respectively, and that ZO-1 is a functional component in the cadherin-based cell adhesion system.
ZO-1 is a peripheral membrane protein with a molecular mass of 220 kD that was first identified as a component of tight junctions (TJ) of epithelial and endothelial cells (Stevenson et al., 1986; Anderson et al., 1988). As a ZO-1–binding protein, another peripheral membrane protein called ZO-2 with a molecular mass of 160 kD has been identified (Gumbiner et al., 1991). Sequence analysis of the cDNAs encoding mammalian ZO-1 and ZO-2 revealed that both show similarity to the product of lethal (1) discs large-1 (dlg),1 one of the tumor suppressor molecules in Drosophila (Itoh et al., 1993; Tsukita et al., 1993; Willott et al., 1993; Jesaitis and Goodenough, 1994), and that there are at least two isotypes of ZO-1 generated by alternative splicing (α+ and α−) (Willot et al., 1992). Recently, in addition to these proteins, many dlg-like proteins have been identified, indicating the existence of a novel gene family named membrane-associated guanylate kinase homologues (MAGUKs) (Woods and Bryant, 1993; Kim, 1995; Anderson, 1995).
We developed an isolation procedure for cell-to-cell adherens junctions (AJ) from rat liver (Tsukita and Tsukita, 1989). Using this fraction, we identified a 220-kD peripheral membrane protein that was highly concentrated at AJ of cardiac muscle cells (intercalated discs) and cultured fibroblasts (Itoh et al., 1991), suggesting that this 220-kD protein is involved in some function of AJ. Cloning of its cDNA, however, revealed that this protein is identical to ZO-1, indicating that ZO-1 is concentrated not only at TJ in epithelial and endothelial cells but also at AJ in cardiac muscle and fibroblastic cells (Itoh et al., 1993). Similar observations were also reported by other laboratories (Jesaitis and Goodenough, 1994).
TJ is an element of epithelial and endothelial junctional complexes and functions as a primary barrier to the diffusion of solutes through the paracellular pathway (Schneeberger and Lynch, 1992; Gumbiner, 1987, 1993) as well as a fence between the apical and basolateral plasma membrane domains to create and maintain their polarity (Rodriguez-Boulan and Nelson, 1989). ZO-1 and ZO-2 are thought to constitute the undercoat structure of TJ together with other peripheral membrane proteins such as cingulin, 7H6 antigen, and symplekin (Citi et al., 1988; Zhong et al., 1993; Keon et al., 1996). An integral membrane protein localized at TJ was recently identified and named occludin (Furuse et al., 1993; Ando-Akatsuka et al., 1996). Occludin has four transmembrane domains in its amino-terminal half and a long carboxyl-terminal cytoplasmic domain. ZO-1 is directly associated with the carboxyl-terminal 150 amino acids (aa) of occludin in TJ (Furuse et al., 1994).
AJ is a specialized region of the plasma membrane, where cadherin molecules (uvomorulin, L-CAM, A-CAM, etc.) function as adhesion molecules and actin filaments are densely associated with the plasma membrane through a well-developed plasmalemmal undercoat (Farquhar and Palade, 1963; Volk and Geiger, 1984; Boller et al., 1985; Takeichi, 1988, 1991). This type of junction is important for the formation and maintenance of tissues. AJ appears beltlike in simple epithelial cells and endothelial cells, but appears spotlike in the intercalated discs of cardiac muscle cells. Small spot-like AJ also occur in cultured fibroblasts (Heaysman and Pegrum, 1973; Yonemura et al., 1995). At least two cytoplasmic proteins, α and β catenins, are tightly associated with the cytoplasmic domain of cadherins (Ozawa et al., 1989, 1990; Nagafuchi and Takeichi, 1989). Those catenins show a similarity to vinculin and the Drosophila armadillo gene product, respectively (Herrenknecht et al., 1991; Nagafuchi et al., 1991; McCrea et al., 1991), and constitute the undercoat of AJ together with other cytoplasmic proteins such as vinculin and α-actinin. As described above, only in nonepithelial cells, ZO-1 is also concentrated in the undercoat of AJ.
Our knowledge is still limited regarding the molecular basis of the association of ZO-1 with the cadherin adhesion system in nonepithelial cells and why this association is not observed in epithelial cells (Itoh et al., 1993). To examine this issue, mouse L cell transfectants expressing exogenous E-cadherin (EL cells; Nagafuchi et al., 1987) provide an advantageous experimental system. They express neither endogenous cadherins nor occludin and show strong cell adhesion activity only through the exogenous E-cadherin. They express endogenous α and β catenin, which form a stable complex with E-cadherin (Nagafuchi et al., 1991). Furthermore, they express endogenous ZO-1 but not ZO-2. In this study, we analyzed the behavior of introduced exogenous full-length, amino-terminal half and carboxyl-terminal half ZO-1 molecules in EL cells and also examined the binding abilities of these molecules to E-cadherin, α catenin, β catenin, and actin filaments in vitro. These analyses led us to conclude that ZO-1 is involved in the cadherin-based cell adhesion system through its direct binding to α catenin and actin filaments.
Materials and Methods
Antibodies and Cells
Mouse anti–ZO-1 mAbs, T8-754 and T7-713, and rat anti–α catenin mAb, α18, have been described (Itoh et al., 1991, 1993; Nagafuchi et al., 1991). T8-754 recognizes the amino-terminal half of ZO-1, while T7-713 is specific for its carboxyl-terminal half. Rabbit anti–E-cadherin pAb and rat anti–E-cadherin mAb, ECCD-2, were provided by Dr. M. Takeichi (Kyoto University, Kyoto, Japan). Mouse anti–β catenin mAb and mouse anti–c-myc tag mAb, 9E10, were purchased from Transduction Labs (Lexington, KY), and Sigma Chemical Co. (St. Louis, MO), respectively.
Sf9 cells were obtained from InVitrogen (San Diego, CA) and cultured in TC-100 medium (GIBCO BRL, Gaithersburg, MD) supplemented with tryptose phosphate broth and 10% FCS at 27°C. Mouse L cells and E-cadherin–transfected L cells (EL cells; clone ELβ1) were provided by Dr. M. Takeichi and cultured in DME supplemented with 10% FCS (Nose et al., 1988).
Constructs and Transfections
To express a protein carrying seven c-myc epitopes (EQKLISEEDL) in tandem at its carboxyl end, a tandem array of seven copies of an oligonucleotide encoding the c-myc epitope was ligated into the expression plasmid pME18S producing pME18S-7myc. The tandem c-myc tag and pME18S were provided by Drs. K. Green (Northwestern University, Chicago, IL) and K. Maruyama (Tokyo Medical and Dental University, Tokyo, Japan), respectively. The cDNA fragment containing the entire open reading frame of mouse ZO-1 (α+ isotype) was produced by PCR using clone 220-1 as a template (Itoh et al., 1993). The first half of this fragment was amplified using the sense primer from position 404 containing an EcoRI site (5′-CAGAATTCATGTCCGCCAGGGCCGCG-3′) and the antisense primer from position 2686 (5′-AAGCTTATGAGAGCGTTC-3′), which contained an internal HindIII site. The second half was amplified using the sense primer from position 2681 (5′-AAGCTTCTGAAGAACAAT-3′) and the antisense primer from position 5638 containing a SalI site (5–CCGTCGACAAAGTGGTCAATCAGGAC-3′). The first half was digested with EcoRI and HindIII, the second half was digested with HindIII and SalI, and then both fragments were ligated. This single recombinant fragment was subcloned into the EcoRI/SalI sites of pBluescript SK(−) (designated as pBFZO-1), and then the EcoRI/SalI-digested fragment was inserted into pME18S-7myc (Fig. 1). The cDNA fragments encoding the ZO-1 amino-terminal half dlg-like (N-ZO-1;1–862 aa) and carboxyl-terminal half (C-ZO-1;863–1745 aa) domains were also produced by PCR using clone 220-1 as a template (Itoh et al., 1993). As primers, 5′-CAGAATTCATGTCCGCCAGGGCCGCG-3′ (sense for amino-terminal half), 5′-CCGTCGACAAGAGTTTCATCTAGTTC-3′ (antisense for amino-terminal half), 5′-CCGAATTCAATGATGAGGTGGGGACT-3′ (sense for carboxyl-terminal half), and 5′-CCGTCGACAAAGTGGTCAATCAGGAC-3′ (antisense for carboxyl-terminal half) were used. Each amplified fragment was subcloned into EcoRI/SalI sites of pBluescript SK(−) (designated as pBNZO1 and pBCZO1, respectively) and then into pME18S-7myc (Fig. 1).
Figure 1.

Structure of c-myc– tagged full-length ZO-1 (F-ZO-1), dlg-like amino-terminal half ZO-1 (N-ZO-1), and carboxyl-terminal half ZO-1 (C-ZO-1). A tandem array of seven c-myc epitopes (myc; EQKLISEEDL) was tagged to the carboxyl terminus of each molecule. F-ZO-1 contains three PDZ domains (PDZ1-3), one SH3 domain (SH3), one guanylate kinase domain (GUK), and a proline-rich domain (proline-rich). The α+ isotype with an insert (α) was used in this study.
EL cells (1 × 107 cells) were cotransfected with 2 μg of each expression vector and 0.1 μg of pSV2bsr (Funakoshi Pharmaceutical Co., Tokyo, Japan) using Lipofectamine reagent (GIBCO BRL) for 5 h in Opti-MEM (GIBCO BRL), followed by addition of 3 ml DME supplemented with 10% FCS. After incubation for 43 h, cells were replated onto four 9-cm dishes in the presence of 2 μg/ml of blasticidin-S to select stable transfectants. Colonies of blasticidin-resistant cells were isolated and screened by immunoblotting with anti–c-myc tag mAb. The positive clones were recloned. Clones expressing full-length ZO-1, N-ZO-1, or C-ZO-1 were designated as FZ-EL, NZ-EL, or CZ-EL cells, respectively. Three independent clones were isolated in each transfection (e.g., FZ-EL-1, FZ-EL-2, and FZ-EL-3) and examined. Since these three independent clones gave the same results, we will mainly discuss FZ-EL-1, NZ-EL-1, and CZ-EL-1 cells in this paper. Furthermore, we performed analyses using the α− isotype of ZO-1 and obtained the same results (data not shown).
Immunofluorescence Microscopy
Cells plated on glass coverslips were rinsed in PBS and fixed with 1% formaldehyde in PBS for 15 min at room temperature. The fixed cells were treated with 0.3% Triton X-100 in PBS for 15 min and washed three times with PBS. After soaking in PBS containing 1% BSA, the samples were treated with primary antibodies for 1 h in a moist chamber. They were then washed three times with PBS, followed by incubation for 30 min with FITC-conjugated sheep anti–mouse IgG and/or rhodamine-conjugated donkey anti–rabbit IgG (Chemicon, Temecula, CA). In some experiments, rhodamine phalloidin (Molecular Probes, Inc., Eugene OR) was added to detect F-actin. The samples were washed with PBS three times, embedded in 95% glycerol-PBS containing 0.1% para-phenylendiamine and 1% n-propylgalate, and examined with a photomicroscope (model Axiophot; Carl Zeiss, Jena, Germany).
Immunoprecipitation
Confluent monolayers of cultured EL or NZ-EL cells on 9-cm dishes were washed three times with ice-cold HMF buffer (HCMF [Ca2+/Mg2+-free Hepes-buffered saline, pH 7.4] containing 2 mM CaCl2) and then lysed in 2 ml of extraction buffer (0.5% NP-40, 2 mM PMSF, 2 μg/ml leupeptin in HMF). The cell lysate clarified by centrifugation at 100,000 g for 30 min was incubated with 100 μl of protein G–Sepharose bead slurry (Zymed Laboratories, Inc., San Francisco, CA) coupled with rat anti–E-cadherin mAb, ECCD-2, or mouse anti–myc epitope tag mAb, 9E10, for 3 h at 4°C. For control, rat or mouse IgG was used. After five washes with the extraction buffer, immunoprecipitates were eluted with 500 μl of 1 M acetic acid twice. These acid eluates were combined, lyophilized, and resolved in SDS sample buffer.
Production of the Amino- and Carboxyl-terminal Halves of ZO-1 by Recombinant Baculovirus Infection
The EcoRI/SalI-digested fragments from pBFZO-1, pBNZO-1, and pBCZO-1 were integrated into the baculovirus genome (Hirao et al., 1996). The recombinant virus containing each cDNA was isolated and condensed using a MAXBAC kit (InVitrogen). Insect Sf9 cells were infected with recombinant viruses and cultured in 100 ml spinner flasks at 27°C for 72 h. Cells were collected and washed with PBS. After suspension in PBS containing 0.1% Triton X-100, 2 mM PMSF, and 4 μg/ml of leupeptin, the cells were homogenized in a tight-fitting Dounce homogenizer, sonicated, and clarified by centrifugation at 100,000 g for 1 h at 4°C. Immunoblotting revealed that the level of expression of the full-length ZO-1 in Sf9 cells was too low to be used for in vitro binding assay with glutathione-S-transferase (GST) fusion proteins or actin filaments. Therefore, we used only recombinant N-ZO-1 and C-ZO-1, which were abundantly expressed in Sf9 cells, for the in vitro binding analyses.
In Vitro Binding Assay Using GST Fusion Proteins
The cDNA encoding the full length of the cytoplasmic domain of mouse E-cadherin (2,268–2,720 bp) with BamHI and EcoRI sites at its ends was produced by PCR using pBATEM2 (Nose et al., 1988) as the template and subcloned into pBluescript SK(−). The insert was then subcloned into the pGEX2T expression vector (Pharmacia LKB Biotechnology, Uppsala, Sweden), which was designed to express a GST fusion protein in Escherichia coli. The cDNAs containing the entire coding regions of mouse α or β catenin with BamHI and EcoRI sites at the ends were produced by PCR using PSK102B (Nagafuchi and Tsukita, 1994) or pMβcat (Nagafuchi et al., 1994), respectively, as the template and subcloned into pBluescript SK(−). The inserts were then subcloned into the pGEX2T expression vector. The GST fusion protein with the cytoplasmic domain of occludin (358–504 aa; GST-Oc358) was previously described (Furuse et al., 1994).
GST fusion proteins were expressed in E. coli and purified using glutathione-Sepharose 4B beads (Pharmacia LKB Biotechnology) as previously described (Furuse et al., 1994). After the 200 μl of glutathione-Sepharose bead slurry containing GST fusion proteins was washed with 100 vol of PBS containing 0.1% Triton X-100, 2 mM PMSF, and 4 μg/ml of leupeptin by brief centrifugation, 2 ml of the cell lysate of Sf9 cells expressing N-ZO-1 or C-ZO-1 was added, followed by incubation for 3 h at 4°C. The beads were washed with 40 vol of the same solution, and then bound proteins were eluted with 1 ml of 50 mM Tris-HCl buffer, pH 8.0, containing 10 mM glutathione. The amounts of GST fusion proteins in each eluate were determined by SDS-PAGE. An appropriate amount of each eluate was again applied to SDS-PAGE to contain the same amount of GST fusion proteins, and the amount of N-ZO-1 or C-ZO-1 in each eluate was evaluated by CBB staining and/or immunoblotting.
To estimate the dissociation constant between N-ZO-1 and α catenin, 200 μl of the glutathione-Sepharose bead slurry containing 40 μg of GST–α catenin was incubated with 2 ml of the Sf9 cell lysate containing 0.012–0.6 μg of N-ZO-1. The beads were washed, and bound proteins were eluted with 1 ml of 50 mM Tris-HCl buffer, pH 8.0, containing 10 mM glutathione. The amounts of N-ZO-1 in the cell lysate or in each eluate were estimated as follows: N-ZO-1 was precipitated out from the Sf9 cell lysate using GST–occludin (GST-Oc358) as shown in Fig. 5 a. This was tentatively called ‘purified' N-ZO-1, and the amount of this purified N-ZO-1 was determined densitrometrically by comparing the intensity of its band after Coomassie brilliant blue staining with those of various amounts of BSA using Adobe PhotoshopTM 3.0J histogram (San Jose, CA). We then compared the intensity of immunoblotted N-ZO-1 bands in cell lysates or in eluates with those of immunoblotted bands of various amounts of purified N-ZO-1 resolved and immunoblotted in the same gel. The intensity of the immunoblotted band was also measured by densitrometry using Adobe PhotoshopTM 3.0J histogram, and a Scatchard plot of the data was generated. Experiments were repeated three times for each estimation of K d.
Figure 5.



Association of N-ZO-1 with α catenin in vitro. (a) GST fusion proteins with the cytoplasmic domain of occludin, the cytoplasmic domain of E-cadherin, α catenin, and β catenin, which were bound to glutathione-Sepharose beads, were incubated with the lysate of Sf9 cells expressing N-ZO-1. After washing, the proteins associated with GST fusion proteins were eluted from the beads with a buffer containing glutathione. The Sf9 cell lysate (Sf9 lysate), the eluate from GST– occludin beads (GST/Occ-eluate), the eluate from GST–E-cadherin beads (GST/Cad-eluate), the eluate from GST–α catenin beads (GST/ α cat-eluate), the eluate from GST–β catenin beads (GST/β cat-eluate), and purified GST–α catenin (GST/α cat) were separated by SDS-PAGE followed by immunoblotting with anti–ZO-1 mAb, T8-754. Although the N-ZO-1 band was not resolved from the GST–α catenin band by CBB staining, immunoblotting identified N-ZO-1 (arrowhead) in the eluate from GST–α catenin beads at the same amount as in that from GST–occludin beads (arrow), in which the N-ZO-1 band was clearly visualized by CBB staining (asterisk). N-ZO-1 was not detected in the eluates from GST–E-cadherin or GST–β catenin beads. Comparison of the CBB staining patterns between the GST–α catenin beads eluate (GST/α cat-eluate) and the purified GST–α catenin (GST/α cat) revealed that only N-ZO-1 was precipitated out from the crude Sf9 lysate by GST–α catenin, indicating the direct α catenin/N-ZO-1 interaction. 1, 2, 3, and 4 indicate the bands of GST–occludin, GST–E-cadherin, GST–α catenin, and GST–β catenin, respectively, and the other lower molecular mass bands are their degradation products. (b) GST fusion protein with α catenin, which was bound by glutathione-Sepharose beads, was incubated with the lysate of Sf9 cells expressing C-ZO-1. After washing, the proteins associated with the GST fusion protein were eluted from the beads with a buffer containing glutathione. The Sf9 cell lysate (Sf9-lysate) and the eluate from GST–α-catenin beads (GST/α cat-eluate) were separated by SDS-PAGE followed by immunoblotting with anti–C-ZO-1 mAb, T7-713. 5, the band of GST–α catenin. No specific binding was detected between GST–α-catenin and C-ZO-1 (arrow). (c) Quantitative analysis of the binding between N-ZO-1 and α catenin. Glutathione-Sepharose bead slurry containing 40 μg of GST–α catenin was incubated with the Sf9 cell lysate containing 0.012–0.6 μg of N-ZO-1. The amounts of N-ZO-1 in the Sf9 cell lysate and in each eluate (inset) were estimated as described in Materials and Methods. Each point represents the mean ± SD of triplicate determinations. The binding was saturable, and Scatchard analysis (inset) indicated that the K d was ∼0.5 nM.
Cosedimentation Experiments with Actin Filaments
Actin was purified from rabbit skeletal muscle according to the method of Pardee and Spudich (1982) and further purified by gel filtration on a Sephacryl S-200 column (Pharmacia LKB Biotechnology). Gel-filtered G-actin was stored in buffer A (2 mM Tris-HCl, pH 8.0, 0.2 mM ATP, 0.5 mM DTT, 0.2% NaN3) and diluted at room temperature to 2–3 μM with buffer F (20 mM Tris-HCl, pH 7.4, 75 mM KCl, 10 mM NaCl, 2 mM DTT, 2.5 mM MgCl2) to initiate polymerization. The cell lysate of Sf9 cells expressing C-ZO-1 was dialyzed against buffer F at 4°C overnight and centrifuged at 100,000 g for 30 min. After actin filaments were polymerized for 30 min, 100 μl of the supernatant was added to 100 μl of the actin filament solution and incubated for 30 min at room temperature. After centrifugation at 100,000 g for 30 min at 20°C, the supernatant and pellet were resolved by SDS-PAGE, followed by immunoblotting with anti–ZO-1 mAb.
To estimate the dissociation constant between C-ZO-1 and actin filaments, 100 μl of the actin filament solution was incubated with 100 μl of Sf9 cell lysates containing 0.2–2 μg of C-ZO-1 for 30 min at room temperature. After centrifugation, the pellet was resolved by SDS-PAGE (see Fig. 6). The amount of cosedimented C-ZO-1 was estimated densitrometrically by comparing the intensity of its Coomassie brilliant blue-stained band with those of various amounts of BSA using Adobe PhotoshopTM 3.0J histogram. A Scatchard plot of the data was then generated. Experiments were repeated three times for each estimation of K d.
Figure 6.


Association of C-ZO-1 with actin filaments in vitro. (a) Sf9 cell lysate containing C-ZO-1 (Sf9-lysate) was incubated with actin filaments (+Actin) or BSA (+BSA). After centrifugation, the supernatant (S) and pellet (P) were resolved by SDS-PAGE, followed by immunoblotting with anti–ZO-1 mAb, T7-713. From crude Sf9 cell lysate, only the C-ZO-1 band was precipitated out by actin filaments (arrows), but not by BSA. Arrowheads indicate actin or BSA. (b) Quantitative analysis of the binding between C-ZO-1 and actin filaments. Actin filaments were incubated with the Sf9 cell lysate containing 0.2–2 μg of C-ZO-1. The amounts of C-ZO-1 in the Sf9 cell lysate and in each pellet (inset) were estimated as described in Materials and Methods. Each point represents the mean ± SD of triplicate determinations. The binding was saturable, and Scatchard analysis (inset) indicated that the K d was ∼10 nM.
Cell Aggregation and Intercellular Motility Assays
Cell aggregation assay was performed basically according to the method developed by Takeichi (1977). Cells were treated with 0.1% trypsin in the presence of 2 mM CaCl2 at 37°C for 20 min and then washed with HCMF to obtain single-cell suspensions. Cells suspended in DME medium were placed in 1% agar-coated four-well plates and rotated at 37°C on a gyratory shaker at 80 rpm for 30 min, 2 h, or 6 h.
For the intercellular motility assay, cells were labeled with DiI according to the method developed by Honig and Hume (1986) and treated with trypsin. Aliquots of 1 × 103 labeled cells were seeded on 6-cm dishes with a monolayer of 2 × 106 cells. After 48 h of culture, four sister cells that seemed to be derived from one seeded cell were examined by fluorescence microscopy. When cell line A was seeded on confluent cultures of cell line B, we designate the experiment as A/B analysis. For quantification of intercellular motility, intercellular distances of all combinations between four cells were measured and summed as Dc. As a control experiment, labeled cells were seeded on the dishes in the absence of a cell sheet. In this case, the intercellular distances were summed as Dd. The degree of intercellular motility was represented as Dc/Dd. At least 24 independent samples were picked up to determine Dc or Dd for each transfectant cell line.
Gel Electrophoresis and Immunoblotting
One-dimensional SDS-PAGE (10–12.5% gel) was based on the method of Laemmli (1970). Gels were stained with Coomassie brilliant blue R-250 or with a silver staining kit (Wako Pure Chemical Industries, Osaka, Japan). For immunoblotting, proteins separated by SDS-PAGE were electrophoretically transferred onto nitrocellulose sheets, which were then incubated with primary antibodies. The antibodies were detected with a blotting detection kit (Amersham Corp., Arlington Heights, IL).
Results
Behavior of Amino-terminal and Carboxyl-terminal Halves of ZO-1 in EL Cells
As shown previously (Itoh et al., 1993), endogenous ZO-1 is concentrated at the cadherin-based cell adhesion sites in EL cells (L cells expressing exogenous E-cadherin) (Fig. 2, a and b). When the myc-tagged full-length mouse ZO-1 was introduced into EL cells, it was also concentrated at the cadherin-based cell adhesion sites in a serrated pattern (Figs. 1 and 2, c and d). Then, to narrow down the region responsible for this characteristic distribution, we divided the ZO-1 molecule into two parts: the amino-terminal half dlg-like domain (N-ZO-1) and the carboxyl-terminal half domain (C-ZO-1) (see Fig. 1). After the myc tag was fused with their carboxyl-end, they were transfected into EL cells. As shown in Fig. 3, a, b, and b′, the introduced N-ZO-1 was concentrated at cell–cell contact sites together with E-cadherin. In these transfectants (NZ-EL), N-ZO-1 and E-cadherin showed a tendency to concentrate in a linear fashion, suggesting a dominant-negative effect of N-ZO-1 in the cadherin-based junction distribution. However, since depending on the culture conditions they showed a serrated concentration pattern (Fig. 3 b, inset), it was difficult to conclusively evaluate the dominant-negative effect in this transfection system. In contrast, C-ZO-1 was distributed along the stress fibers (Fig. 3, c, d, and d′). Of course, the possibility was not excluded that C-ZO-1 was also present at the serrated points of cell contact in these transfectants (CZ-EL). N-ZO-1 and C-ZO-1 showed the same behavior when transfected into the rat fibroblast cell line, 3Y1 cells (data not shown).
Figure 2.
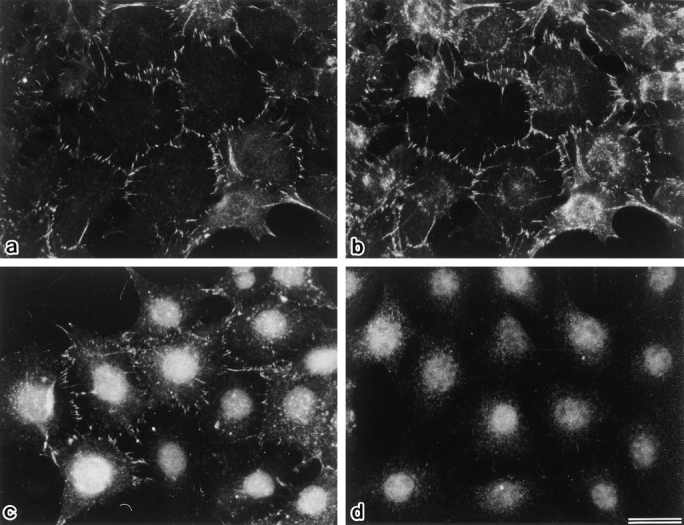
Concentration of full-length ZO-1 at cadherin-based cell adhesion sites. When parent EL cells were doubly stained with anti– ZO-1 mAb, T8-754 (a), and anti–E-cadherin pAb (b), both signals were colocalized at the cell–cell border. Introduced myc-tagged full-length ZO-1 was similarly distributed in EL transfectants (FZ-EL cells) as revealed by anti–c-myc tag mAb, 9E10 (c). This mAb gave a signal in nuclei but not in cell–cell borders of parent EL cells (d). Bar, 10 μm.
Figure 3.
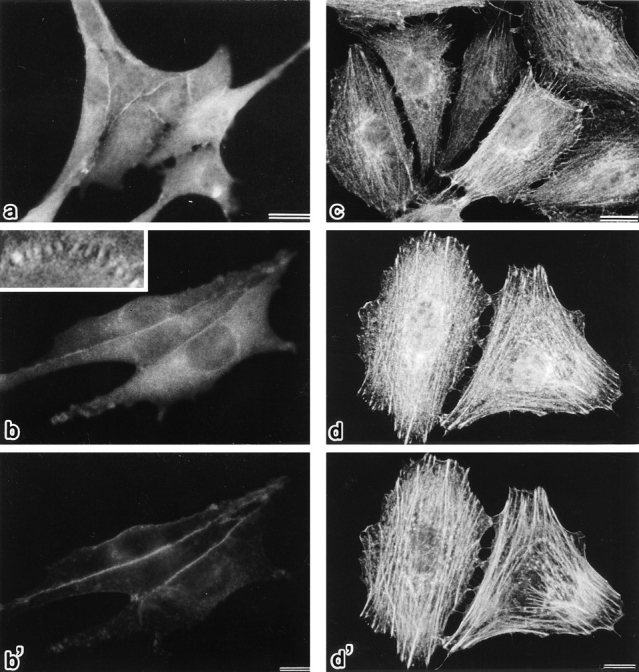
Subcellular distribution of N-ZO-1 and C-ZO-1 in EL transfectants (NZ-EL and CZ-EL, respectively). NZ-EL cells (a, b, and b′) were singly stained with anti–c-myc tag mAb, 9E10 (a), or doubly stained with 9E10 (b) and anti–E-cadherin pAb (b′). In addition to the diffuse cytoplasmic distribution, introduced N-ZO-1 was significantly concentrated at the cell–cell border, where E-cadherin was colocalized. N-ZO-1 and E-cadherin showed a tendency to distribute in a linear fashion, but depending on the culture conditions they showed a serrated concentration pattern (inset, 3,000×). CZ-EL cells (c, d, and d′) were singly stained with anti–c-myc tag mAb, 9E10 (c), or doubly stained with 9E10 (d) and rhodamine phalloidin (d′). Introduced C-ZO-1 was distributed along actin stress fibers visualized with rhodamine phalloidin. Bars: (a–c) 10 μm; (d and d′) 20 μm.
Association of the Amino-terminal Half of ZO-1 with the Cadherin/Catenin Complex In Vivo
The transfection results suggested that endogenous ZO-1 is concentrated at cell–cell contact sites through the direct or indirect association of its amino-terminal dlg-like domain with the cadherin/catenin complex. The interaction of ZO-1 with the cadherin/catenin complex in vivo was then examined by immunoprecipitation. The E-cadherin immunoprecipitates from the 0.5% NP-40 lysate of EL cells contained α and β catenins as major components, whereas ZO-1 was hardly detected even by immunoblotting (Fig. 4 a). Considering that most of endogenous ZO-1 is resistant against detergent extraction (Itoh et al., 1991), it is expected that the cadherin/catenin complex–associated ZO-1 was not extracted by 0.5% NP-40. Then, E-cadherin was immunoprecipitated from NZ-EL cells. As shown in Fig. 4 a, in the E-cadherin immunoprecipitate containing α and β catenins, N-ZO-1 was detected by immunoblotting. Furthermore, when the introduced N-ZO-1 was immunoprecipitated with anti-myc mAb, E-cadherin, α catenin, and β catenin were coimmunoprecipitated (Fig. 4 b). By contrast, when E-cadherin was immunoprecipitated from CZ-EL cells, C-ZO-1 was not detected in the precipitated cadherin/catenin complex (data not shown). We then concluded that N-ZO-1 (and probably endogenous ZO-1) was concentrated at cadherin-based cell adhesion sites in EL cells through its interaction with the cadherin/catenin complex in vivo.
Figure 4.

Association of N-ZO-1 with the E-cadherin/ catenin complex in NZ-EL cells. (a) Silver staining pattern of E-cadherin immunoprecipitates from EL or NZ-EL cells and accompanying immunoblots with anti–ZO-1 mAb, T8-754, specific for the amino-terminal half of ZO-1. When E-cadherin (Cad) was immunoprecipitated with rat anti–E-cadherin mAb, ECCD-2 (anti-cad), from the NP-40 extract of EL or NZ-EL cells, α and β catenins were coimmunoprecipitated. (α and β, respectively). In the E-cadherin immunoprecipitates from NZ-EL cells, β catenin appeared to be degraded to some extent. Rat IgG (r-IgG) was used for control immunoprecipitation. Asterisks indicate IgG. Immunoblotting with anti–ZO-1 mAb detected N-ZO-1 (arrowhead) in the E-cadherin immunoprecipitate from NZ-EL cells but did not detect endogenous full-length ZO-1 in the E-cadherin immunoprecipitate from either NZ-EL or EL cells. (b) Immunoprecipitation with anti–c-myc tag mAb, 9E10 (anti-myc), from the NP-40 extract of NZ-EL cells. Silver staining detected the N-ZO-1 band (arrow) in addition to IgG bands (asterisks), and immunoblotting identified E-cadherin, α catenin, and β catenin in the immunoprecipitate. Mouse IgG (m-IgG) was used for control immunoprecipitation.
Direct Binding of the Amino-terminal Half of ZO-1 with α Catenin In Vitro
Immunoprecipitation experiments suggested that the cytoplasmic domain of E-cadherin, α catenin, or β catenin directly binds to the amino-terminal half of ZO-1. To determine the binding partner for ZO-1 in the cadherin/catenin complex, we produced GST fusion proteins with the cytoplasmic domain of E-cadherin, α catenin, and β catenin and performed in vitro binding assays with the recombinant N-ZO-1 produced in Sf9 cells by baculovirus infection (Fig. 5 a). In this assay, the electrophoretic mobility of recombinant N-ZO-1 was almost the same as those of GST–α catenin and GST–β catenin fusion proteins, making it difficult to estimate the bound N-ZO-1 by CBB staining. Thus, to estimate the amount of N-ZO-1 by immunoblotting, N-ZO-1 was first purified from the crude lysate of Sf9 cells using the GST/cytoplasmic domain of occludin, as previously reported (Furuse et al., 1994). By comparison with the amount of this purified N-ZO-1 determined by CBB staining and immunoblotting, we estimated the amounts of α catenin– or β catenin–bound N-ZO-1, if any. As shown in Fig. 5 a, N-ZO-1 was directly and specifically bound to GST–α catenin with similar affinity to the cytoplasmic domain of occludin, but not to the cytoplasmic domain of E-cadherin or to β catenin. Using the same system, we checked the possible interaction of recombinant C-ZO-1 with GST fusion proteins with the cytoplasmic domain of E-cadherin, α catenin, and β catenin, but no specific binding was detected (Fig. 5 b).
Next, using this in vitro binding assay, we estimated the dissociation constant between N-ZO-1 and α catenin. For this purpose, the interaction of N-ZO-1 with α catenin was examined by incubating various concentrations of N-ZO-1 with beads bearing GST–α catenin fusion protein or GST (for control). The dissociation constant was estimated by subtracting the binding to the GST-conjugated Sepharose beads. The binding was saturable, and Scatchard analysis revealed a single class of affinity binding sites with a K d of ∼0.5 nM (Fig. 5 c).
Direct Binding of the Carboxyl-terminal Half of ZO-1 with Actin Filaments
Immunofluorescence microscopy revealed that C-ZO-1 was concentrated along stress fibers. We then examined the binding affinity of C-ZO-1 with actin filaments in vitro. When actin filaments were incubated with the cell lysate of Sf9 cells expressing C-ZO-1, only the recombinant C-ZO-1 in the lysate was selectively cosedimented with actin filaments (Fig. 6 a). In control experiments, BSA was used instead of actin, and no significant amount of C-ZO-1 was sedimented. Then, using this cosedimentation system, we determined the dissociation constant between C-ZO-1 and actin filaments. For this purpose, the interaction of C-ZO-1 with actin filaments was examined by incubating various concentrations of C-ZO-1 with actin filaments or BSA (for control). The dissociation constant was estimated by subtracting the amount of pelleted C-ZO-1 in the presence of BSA. The binding was saturable, and Scatchard analysis revealed a single class of affinity binding sites with a K d of ∼10 nM (Fig. 6 b).
Dominant-negative Effects of the Amino-terminal Half of ZO-1 on the Cadherin-based Cell Adhesion System
If in EL cells the endogenous ZO-1 was functionally involved in the cadherin-based cell adhesion, some dominant-negative effect of N-ZO-1 in cell adhesion was expected in NZ-EL cells. Conventional cell aggregation assay (Takeichi, 1977) was then performed to compare the cadherin-dependent cell adhesion activity between EL and NZ-EL cells. However, no significant difference was detected (Fig. 7).
Figure 7.
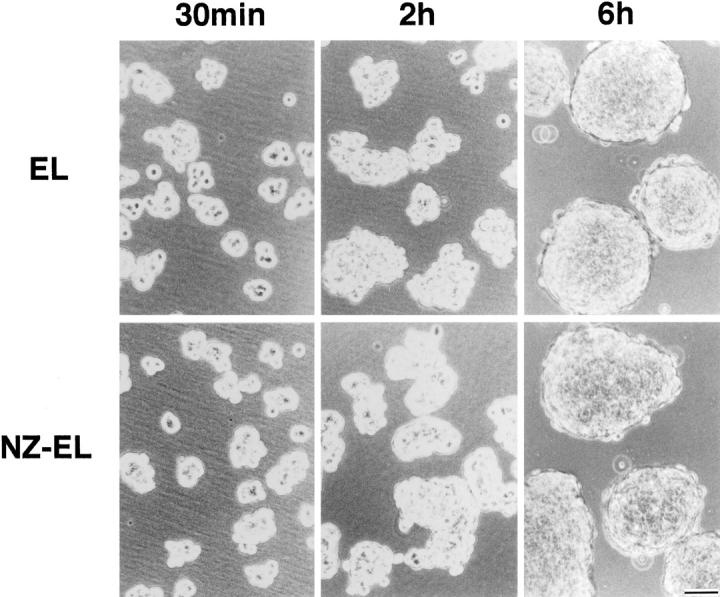
Cell aggregation assay of EL and NZ-EL cells. Single cells were rotated for 30 min, 2 h, or 6 h on a gyratory shaker at 80 rpm. No significant differences were detected in cell aggregation activity between EL and NZ-EL cells. Bar, 50 μm.
To detect the qualitative changes in cadherin-based cell adhesion, the intercellular motility assay was used (Nagafuchi et al., 1994) (Fig. 8). In this assay, a single L, EL, or NZ-EL cell labeled with a fluorescent dye (DiI) was seeded onto a confluent culture of nonlabeled L, EL, or NZ-EL cells, respectively, and after 48 h in culture (twice the doubling time), the cell scattering of four labeled sister cells was analyzed by measuring the mass distance among these four cells. As shown previously (Nagafuchi et al., 1994), when the labeled EL cells were seeded on the EL cell sheet, the four sister cells were more scattered as compared to labeled L cells on the L cell sheet. In contrast, when the labeled NZ-EL cells were seeded on the NZ-EL cell sheet, the scattering of the labeled cells was significantly decreased. The intercellular motility of NZ-EL cells was comparable to that of L cells on the L cell sheet. Fig. 8 b summarizes the quantitative results of the intercellular motility assay, where the index was calculated as described in Materials and Methods.
Figure 8.


Intercellular motility of L, EL, and NZ-EL cells. (a) In the intercellular motility assay (on cell sheet), the fluorescence microscopic images of four DiI-stained sister cells (white) were superimposed on the phase contrast image of a confluent cell sheet. As compared to EL/EL analysis (see Materials and Methods for details), the intercellular motility in the NZ-EL/NZ-EL analysis was suppressed down to the level of that in L/ L analysis. These cell lines showed similar scattering properties on plastic dishes in the absence of a cell sheet (on dish). (b) Quantitative analysis of the intercellular motility. Higher values on the abscissa (intercellular motility index [Dc/Dd]; see Materials and Methods for details) represent the higher degrees of intercellular motility. At least 24 independent samples were picked up to determine the intercellular motility index for each analysis. Bar, 10 μm.
Discussion
Cadherins are responsible for Ca2+-dependent cell–cell adhesion (Gumbiner et al., 1988; Takeichi, 1988, 1991; Kemler, 1992; Birchmeier, 1995). One of the major characteristics of cadherins is that they interact intimately with the actin-based cytoskeleton to exert their physiological function, i.e., strong cell adhesion (Hirano et al., 1987; Nagafuchi and Takeichi, 1988). The identification of cytoplasmic proteins responsible for cadherin/actin interaction is thus required to better understand how the functions of cadherin are regulated. E-cadherin has been immunoprecipitated from the detergent extracts of various cell types, and in the immunoprecipitate at least two proteins, named α and β catenins, have been identified (Ozawa et al., 1989; Nagafuchi and Takeichi, 1989). It is now accepted that the cytoplasmic domain of cadherins directly binds to β catenin, which then binds to α catenin (Ozawa and Kemler, 1992; Aberle et al., 1994; Hinck et al., 1994; Näthke et al., 1994; Oyama et al., 1994; Jou et al., 1995). However, immunoprecipitation analysis along this line is expected to face technical difficulties, since the functioning cadherins that show strong cell adhesion activity are associated with actin-based cytoskeletons and resist detergent extraction (Nagafuchi and Takeichi, 1988; Tsukita et al., 1992).
In nonepithelial cells lacking TJ, ZO-1 is exclusively concentrated at cell–cell adhesion sites, where the functioning cadherins are intimately associated with the actin-based cytoskeletons (Itoh et al., 1991, 1993). Furthermore, ZO-1 in these cells is highly resistant to detergent extraction, suggesting that ZO-1 is a good candidate as a cross-linker between the functioning cadherin/catenin complex and the actin-based cytoskeleton. In this study, we divided the ZO-1 molecule into the amino-terminal half (N-ZO-1) and the carboxyl-terminal half (C-ZO-1). In EL cells expressing these truncated ZO-1 constructs (NZ-EL and CZ-EL cells, respectively), N-ZO-1 was concentrated at the cadherin-based cell–cell adhesion sites, whereas C-ZO-1 was localized along the actin stress fibers. From NZ-EL cells, N-ZO-1, but not the endogenous full-length ZO-1, was coimmunoprecipitated with the cadherin/catenin complex. In this case, we circumvented the above technical difficulty in cadherin immunoprecipitation, probably because N-ZO-1 lost the cross-linking activity while retaining its binding activity to the cadherin/catenin complex. Actually, recombinant N-ZO-1 and C-ZO-1 directly bound in vitro to α catenin and actin filaments, respectively. The dissociation constants of N-ZO-1/α catenin and C-ZO-1/actin filaments in vitro were ∼0.5 and ∼10 nM, respectively. Both of these values are small enough to postulate that binding occurs in vivo. Taken together, we conclude that ZO-1 functions as a cross-linker between α catenin and actin filaments to insolubilize the cadherin/catenin complex in nonepithelial cells. On the other hand, α catenin has been reported to bear at least two cytoskeleton-interacting domains (Nagafuchi et al., 1994) and directly binds to actin filaments and to α-actinin, an actin-binding protein (Knudsen et al., 1995; Rimm et al., 1995). The physiological significance of these possible multiple linkage pathways from α catenin to actin filaments remains elusive.
In fully polarized epithelial cells bearing TJ, ZO-1 is not localized at AJ but at TJ. At the initial phase of polarization of these epithelial cells, however, small spotlike AJ are formed, where ZO-1 is coconcentrated with E-cadherin (Yonemura et al., 1995). It is then possible that also in epithelial cells ZO-1 functions as a cross-linker between α catenin and the actin-based cytoskeleton at the initial stage of the junction formation. In good agreement with this, α and β catenins were reported to be coimmunoprecipitated with ZO-1 only at the initial stage of junction formation in Madin-Darby canine kidney (MDCK) cells, although cadherins were not identified in the immunoprecipitate (Rajasekaran et al., 1996). During the maturation process of beltlike AJ of fully polarized epithelial cells, ZO-1 may be transferred from AJ to TJ through its direct interaction with the cytoplasmic domain of occludin. Taking it into consideration that the dissociation constant between occludin and ZO-1 is ∼1 nM (Itoh, M., and Sh. Tsukita, manuscript in preparation), the molecular mechanism for this peculiar behavior of ZO-1 is one of the most interesting issues from the viewpoint of epithelial polarization.
ZO-1 belongs to the MAGUK family, members of which are characterized by PDZ (DHR) domains. Although the physiological relevance of this domain remains unclear, PSD95, one of the MAGUK family members, was reported to facilitate the lateral aggregation of its membrane binding partners, such as the NMDA receptor and the K+ channel (Kim et al., 1995; Kornau et al., 1995; Niethammer et al., 1996). Thus, it is possible that ZO-1 is also involved in the lateral aggregation of cadherin/catenin complexes in nonepithelial cells as well as in epithelial cells at the initial stage of junction formation. Furthermore, ZO-1 is possibly involved in the lateral aggregation of occludin at TJ in fully polarized epithelial cells.
Recently, ZO-1 was reported to be localized in the nucleus, suggesting its possible involvement in intracellular signaling (Gottardi et al., 1996). The sequence similarity of ZO-1 to the dlg gene product, one of the tumor suppressors in Drosophila, also suggests this possibility. On the other hand, evidence has accumulated that β catenin is directly involved in intracellular signaling to the nucleus (Funayama et al., 1995; Gumbiner, 1995; Behrens et al., 1996; Huber et al., 1996; Miller and Moon, 1996). Furthermore, as discussed above, β catenin as well as α catenin were reported to be coimmunoprecipitated with ZO-1 in MDCK cells at the initial phase of junction formation (Rajasekaran et al., 1996). It is thus tempting to speculate that β catenin directly interacts with ZO-1. However, at least in vitro, this direct interaction was not detected. Recently, we introduced a fusion protein between nonfunctional E-cadherin and α catenin into L fibroblasts (Nagafuchi et al., 1994). This fusion molecule lacked the β catenin–binding site but functioned as an adhesion molecule with strong adhesion activity at the cell–cell adhesion sites. ZO-1, but not β catenin, was concentrated at these cell–cell adhesion sites (Nagafuchi, A., and Sh. Tsukita, manuscript in preparation). This observation favors the notion that α catenin is responsible for the concentration of ZO-1 at the cadherin-based cell adhesion sites. Of course, considering that α catenin appears to cross-link β catenin to N-ZO-1 in the E-cadherin immunoprecipitate from NZ-EL cells, it is possible that β catenin interacts with ZO-1 indirectly via α catenin.
Considering that ZO-1 works as a cross-linker and that N-ZO-1 is concentrated at cadherin-based cell adhesion sites, some dominant-negative effect of N-ZO-1 in the cadherin-based cell adhesion is expected. Various deletion mutants of ZO-1 have been introduced into various types of cells, but no obvious dominant-negative effects on cadherin-based cell adhesion were observed (Fanning, A.S., B.T. Jameson, and J.M. Anderson. 1996. Mol. Biol. Cell. 7: 607a; Itoh, M., and Sh. Tsukita, unpublished data). This may be partly explained by the occurrence of some functionally redundant ZO-1-like proteins. ZO-2 is one candidate for such proteins, and L cells (and EL cells) do not express ZO-2. Furthermore, the lack of occludin in EL cells is also advantageous for analysis and interpretation of the possible dominant-negative effects of truncated ZO-1 molecules. In terms of the conventional cell aggregation activity, however, in EL cells N-ZO-1 did not show any detectable dominant-negative effects. To evaluate dynamic aspects of cadherin-based cell adhesion, we previously developed an assay system for intercellular motility, i.e., the cell movement in a confluent sheet, and reported that the expression of E-cadherin in L cells facilitates intercellular motility (Nagafuchi et al., 1994). Interestingly, N-ZO-1 suppressed this elevated intercellular motility of EL cells down to the level of parent L cells. This indicates that the cross-linking activity of ZO-1 between the cadherin/catenin complex and the actin-based cytoskeleton is functionally involved in cadherin-based cell adhesion, although detailed molecular mechanisms of this dominant-negative effect of N-ZO-1 in EL cells still remain elusive. Further analyses, including the targeted disruption of ZO-1 gene, will lead us to better understand the functional implications of ZO-1 in the cadherin-based cell adhesion system.
Acknowledgments
We would like to thank all the members of our laboratory (Department of Cell Biology, Faculty of Medicine, Kyoto University) for their helpful discussions throughout this study. Our thanks are also due to Drs. M. Takeichi, K. Green, and K. Maruyama for their generous gift of anti– E-cadherin antibodies, the tandem c-myc tag, and the expression plasmid pME18S, respectively.
This work was supported in part by a Grant-in-Aid for Cancer Research and a Grant-in-Aid for Scientific Research (A) from the Ministry of Education, Science, and Culture of Japan (to Sh. Tsukita).
Footnotes
1. Abbreviations used in this paper: aa, amino acid(s); AJ, adherens junctions; C-ZO-1 and N-ZO-1, carboxyl-terminal half ZO-1 and amino-terminal half dlg-like ZO-1; dlg, discs large; GST, glutathione-S-transferase; MAGUK, membrane-associated guanylate kinase homologues; TJ, tight junctions.
Address all correspondence to Shoichiro Tsukita, M.D., Department of Cell Biology, Kyoto University, Faculty of Medicine, Konoe-Yoshida, Sakyo-ku, Kyoto 606, Japan. Tel.: 81-75-753-4372. Fax: 81-75-753-4660. E-mail: htsukita@mfour.med.kyoto-u.ac.jp
References
- Aberle H, Butz S, Stappert J, Weissig H, Kemler R, Hoschuetzky H. Assembly of the cadherin-catenin complex in vitro with recombinant proteins. J Cell Sci. 1994;107:3655–3663. doi: 10.1242/jcs.107.12.3655. [DOI] [PubMed] [Google Scholar]
- Anderson JM. Zonula occludens (ZO)-1 and ZO-2: membrane-associated guanylate kinase homologues (MAGUKs) of the tight junction. Biochem Soc Trans. 1995;23:470–475. doi: 10.1042/bst0230470. [DOI] [PubMed] [Google Scholar]
- Anderson JM, Stevenson BR, Jesaitis LA, Goodenough DA, Mooseker MS. Characterization of ZO-1, a protein component of the tight junction from mouse liver and Madin-Darby canine kidney cells. J Cell Biol. 1988;106:1141–1149. doi: 10.1083/jcb.106.4.1141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ando-Akatsuka Y, Saitou M, Hirase T, Kishi M, Sakakibara A, Itoh M, Yonemura S, Furuse M, Sh, Tsukita Interspecies diversity of the occludin sequence: cDNA cloning of human, mouse, dog, and rat-kangaroo homologues. J Cell Biol. 1996;133:43–47. doi: 10.1083/jcb.133.1.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrens J, von Kries JP, Kuhl M, Bruhn L, Wedlich D, Grosschedl R, Birchmeier W. Functional interaction of β-catenin with the transcription factor LEF-1. Nature (Lond) 1996;382:638–642. doi: 10.1038/382638a0. [DOI] [PubMed] [Google Scholar]
- Birchmeier W. E-cadherin as a tumor (invasion) suppresser gene. Bioessays. 1995;17:97–99. doi: 10.1002/bies.950170203. [DOI] [PubMed] [Google Scholar]
- Boller K, Vestweber D, Kemler R. Cell-adhesion molecule uvomorulin is localized in the intermediate junctions of adult intestinal epithelial cells. J Cell Biol. 1985;100:327–332. doi: 10.1083/jcb.100.1.327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Citi S, Sabanay H, Jakes R, Geiger B, Kendrick-Jones J. Cingulin, a new peripheral component of tight junctions. Nature (Lond) 1988;33:272–276. doi: 10.1038/333272a0. [DOI] [PubMed] [Google Scholar]
- Farquhar MG, Palade GE. Junctional complexes in various epithelia. J Cell Biol. 1963;17:375–409. doi: 10.1083/jcb.17.2.375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Funayama N, Fagotto F, McCrea P, Gumbiner BM. Embryonic axis induction by the armadillo repeat domain of β-catenin: evidence for intracellular signaling. J Cell Biol. 1995;128:959–968. doi: 10.1083/jcb.128.5.959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furuse M, Hirase T, Itoh M, Nagafuchi A, Yonemura S, Tsukita Sa, Tsukita Sh. Occludin: a novel integral membrane protein localizing at tight junctions. J Cell Biol. 1993;123:1777–1788. doi: 10.1083/jcb.123.6.1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furuse M, Itoh M, Hirase T, Nagafuchi A, Yonemura S, Tsukita Sa, Tsukita Sh. Direct association of occludin with ZO-1 and its possible involvement in the localization of occludin at tight junctions. J Cell Biol. 1994;127:1617–1626. doi: 10.1083/jcb.127.6.1617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gottardi CJ, Arpin M, Fanning AS, Louvard D. The junction- associated protein, zonula occludens-1, localizes to the nucleus before the maturation and during the remodeling of cell-cell contacts. Proc Natl Acad Sci USA. 1996;93:10779–10784. doi: 10.1073/pnas.93.20.10779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gumbiner B. Structure, biochemistry, and assembly of epithelial tight junctions. Am J Physiol. 1987;253:C749–C758. doi: 10.1152/ajpcell.1987.253.6.C749. [DOI] [PubMed] [Google Scholar]
- Gumbiner B. Breaking through the tight junction barrier. J Cell Biol. 1993;123:1631–1633. doi: 10.1083/jcb.123.6.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gumbiner B. Signal transduction by β-catenin. Curr Opin Cell Biol. 1995;7:634–640. doi: 10.1016/0955-0674(95)80104-9. [DOI] [PubMed] [Google Scholar]
- Gumbiner B, Stevenson B, Grimaldi A. The role of the cell adhesion molecule uvomorulin in the formation and maintenance of the epithelial junctional complex. J Cell Biol. 1988;107:1575–1587. doi: 10.1083/jcb.107.4.1575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gumbiner B, Lowenkopf T, Apatira D. Identification of a 160-kDa polypeptide that binds to the tight junction protein ZO-1. Proc Natl Acad Sci USA. 1991;88:3460–3464. doi: 10.1073/pnas.88.8.3460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heaysman JE, Pegrum SM. Early contacts between fibroblasts. Exp Cell Res. 1973;78:71–78. doi: 10.1016/0014-4827(73)90039-6. [DOI] [PubMed] [Google Scholar]
- Herrenknecht K, Ozawa M, Eckerskorn C, Lottspeich F, Lenter M, Kemler R. The uvomorulin-anchorage protein α-catenin is a vinculin homologue. Proc Natl Acad Sci USA. 1991;88:9156–9160. doi: 10.1073/pnas.88.20.9156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hinck L, Näthke IS, Papkoff J, Nelson WJ. Dynamics of cadherin/catenin complex formation: novel protein interactions and pathways of complex assembly. J Cell Biol. 1994;125:1327–1340. doi: 10.1083/jcb.125.6.1327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirano S, Nose A, Hatta K, Kawakami A, Takeichi M. Calcium-dependent cell–cell adhesion molecules (cadherins): subclass specificities and possible involvement of actin bundles. J Cell Biol. 1987;105:2501–2510. doi: 10.1083/jcb.105.6.2501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirao M, Sato N, Kondo T, Yonemura S, Monden M, Sasaki T, Takai Y, Tsukita Sh, Tsukita Sa. Regulation mechanism of ERM protein/ plasma membrane association: possible involvement of phosphatidylinositol turnover and rho-dependent signaling pathway. J Cell Biol. 1996;135:37–52. doi: 10.1083/jcb.135.1.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honig MB, Hume RI. Fluorescent carbocyanine dyes allow living neurons of identified origin to be studied in long-term cultures. J Cell Biol. 1986;103:171–187. doi: 10.1083/jcb.103.1.171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huber O, Korn R, McLaughlin J, Ohsugi M, Herrmann BG, Kemler R. Nuclear localization of β-catenin by interaction with transcription factor LEF-1. Mech Dev. 1996;59:3–10. doi: 10.1016/0925-4773(96)00597-7. [DOI] [PubMed] [Google Scholar]
- Itoh M, Yonemura S, Nagafuchi A, Tsukita Sa, Tsukita Sh. A 220-kD undercoat-constitutive protein: its specific localization at cadherin-based cell–cell adhesion sites. J Cell Biol. 1991;115:1449–1462. doi: 10.1083/jcb.115.5.1449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Itoh M, Nagafuchi A, Yonemura S, Kitani-Yasuda T, Tsukita Sa, Tsukita Sh. The 220-kD protein colocalizing with cadherins in non-epithelial cells is identical to ZO-1, a tight junction-associated protein in epithelial cells: cDNA cloning and immunoelectron microscopy. J Cell Biol. 1993;121:491–502. doi: 10.1083/jcb.121.3.491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jesaitis LA, Goodenough DA. Molecular characterization and tissue distribution of ZO-2, a tight junction protein homologous to ZO-1 and the Drosophiladiscs-large tumor suppresser protein. J Cell Biol. 1994;124:949–961. doi: 10.1083/jcb.124.6.949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jou TS, Stewart DB, Stappart J, Nelson WJ, Marrs JA. Genetic and biochemical dissection of protein linkages in the cadherin-catenin complex. Proc Natl Acad Sci USA. 1995;9295:5067–5071. doi: 10.1073/pnas.92.11.5067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kemler R. Classical cadherins. Semin Cell Biol. 1992;3:149–155. doi: 10.1016/s1043-4682(10)80011-x. [DOI] [PubMed] [Google Scholar]
- Keon BH, Schäfer S, Kuhn C, Grund C, Franke WW. Symplekin, a novel type of tight junction plaque protein. J Cell Biol. 1996;134:1003–1018. doi: 10.1083/jcb.134.4.1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim SK. Tight junctions, membrane-associated guanylate kinases and cell signaling. Curr Opin Cell Biol. 1995;7:641–649. doi: 10.1016/0955-0674(95)80105-7. [DOI] [PubMed] [Google Scholar]
- Kim E, Niethammer M, Rothschild A, Jan YN, Sheng M. Clustering of Shaker-type K+ channels by interaction with a family of membrane-associated guanylate kinases. Nature (Lond) 1995;378:85–88. doi: 10.1038/378085a0. [DOI] [PubMed] [Google Scholar]
- Knudsen KA, Soler AP, Johnson KR, Wheelock MJ. Interaction of α-actinin with the cadherin/catenin cell–cell adhesion complex via α-catenin. J Cell Biol. 1995;130:67–77. doi: 10.1083/jcb.130.1.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kornau HC, Schenker LT, Kennedy MB, Seeburg PH. Domain interaction between NMDA receptor subunits and the postsynaptic density protein PSD-95. Science (Wash DC) 1995;269:1737–1740. doi: 10.1126/science.7569905. [DOI] [PubMed] [Google Scholar]
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (Lond) 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- McCrea PD, Turck CW, Gumbiner B. A homolog of the armadillo protein in Drosophila(plakoglobin) associated with E-cadherin. Science (Wash DC) 1991;254:1359–1361. doi: 10.1126/science.1962194. [DOI] [PubMed] [Google Scholar]
- Miller JR, Moon RT. Signal transduction through β-catenin and specification of cell fate during embryogenesis. Genes Dev. 1996;10:2527–2539. doi: 10.1101/gad.10.20.2527. [DOI] [PubMed] [Google Scholar]
- Nagafuchi A, Takeichi M. Cell binding function of E-cadherin is regulated by the cytoplasmic domain. EMBO (Eur Mol Biol Organ) J. 1988;7:3679–3684. doi: 10.1002/j.1460-2075.1988.tb03249.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagafuchi A, Takeichi M. Transmembrane control of cadherin- mediated cell adhesion: a 94 kDa protein functionally associated with a specific region of the cytoplasmic domain of E-cadherin. Cell Regul. 1989;1:37–44. doi: 10.1091/mbc.1.1.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagafuchi A, Tsukita Sh. The loss of the expression of α catenin, the 102kD cadherin-associated protein, in central nervous tissues during development. Dev Growth Differ. 1994;36:59–71. doi: 10.1111/j.1440-169X.1994.00059.x. [DOI] [PubMed] [Google Scholar]
- Nagafuchi A, Shirayoshi Y, Okazaki K, Yasuda K, Takeichi M. Transformation of cell adhesion properties by exogenously introduced E-cadherin cDNA. Nature (Lond) 1987;329:341–343. doi: 10.1038/329341a0. [DOI] [PubMed] [Google Scholar]
- Nagafuchi A, Takeichi M, Tsukita Sh. The 102 kd cadherin-associated protein: similarity to vinculin and posttranscriptional regulation of expression. Cell. 1991;65:849–857. doi: 10.1016/0092-8674(91)90392-c. [DOI] [PubMed] [Google Scholar]
- Nagafuchi A, Ishihara S, Tsukita Sh. The roles of catenins in the cadherin-mediated cell adhesion: functional analysis of E-cadherin–α catenin fusion molecules. J Cell Biol. 1994;127:235–245. doi: 10.1083/jcb.127.1.235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Näthke IS, Hinck L, Swedlow JR, Papkoff J, Nelson WJ. Defining interactions and distribution of cadherin and catenin complexes in polarized epithelial cells. J Cell Biol. 1994;125:1341–1352. doi: 10.1083/jcb.125.6.1341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niethammer M, Kim E, Sheng M. Interaction between the C terminus of NMDA receptor subunits and multiple members of the PSD-95 family of membrane-associated guanylate kinases. J Neurosci. 1996;16:2157–2163. doi: 10.1523/JNEUROSCI.16-07-02157.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nose A, Nagafuchi A, Takeichi M. Expressed recombinant cadherins mediate cell sorting in model systems. Cell. 1988;54:993–1001. doi: 10.1016/0092-8674(88)90114-6. [DOI] [PubMed] [Google Scholar]
- Oyama T, Kanai Y, Ochiai A, Akimoto S, Oda T, Yanagihara K, Nagafuchi A, Tsukita Sh, Shibamoto S, Ito F, et al. A truncated β-catenin disrupts the interaction between E-cadherin and α-catenin: a cause of loss of intercellular adhesiveness in human cancer cell lines. Cancer Res. 1994;54:6282–6287. [PubMed] [Google Scholar]
- Ozawa M, Kemler R. Molecular organization of the uvomorulin-catenin complex. J Cell Biol. 1992;116:989–996. doi: 10.1083/jcb.116.4.989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ozawa M, Baribault H, Kemler R. The cytoplasmic domain of the cell adhesion molecule uvomorulin associates with three independent proteins structurally related in different species. EMBO (Eur Mol Biol Organ) J. 1989;8:1711–1717. doi: 10.1002/j.1460-2075.1989.tb03563.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ozawa M, Ringwald M, Kemler R. Uvomorulin-catenin complex formation is regulated by a specific domain in the cytoplasmic region of the cell adhesion molecule. Proc Natl Acad Sci USA. 1990;87:4246–4250. doi: 10.1073/pnas.87.11.4246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pardee JD, Spudich JA. Mechanism of K+-induced actin assembly. J Cell Biol. 1982;93:648–654. doi: 10.1083/jcb.93.3.648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajasekaran AK, Hojo M, Huima T, Rodriguez-Boulan E. Catenins and zonula occludens-1 form a complex during early stages in the assembly of tight junctions. J Cell Biol. 1996;132:451–463. doi: 10.1083/jcb.132.3.451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rimm DL, Koslov ER, Kebriaei P, Cianci CD, Morrow JS. α1(E)-catenin is an actin-binding and -bundling protein mediating the attachment of F-actin to the membrane adhesion complex. Proc Natl Acad Sci USA. 1995;92:8813–8817. doi: 10.1073/pnas.92.19.8813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez-Boulan E, Nelson WJ. Morphogenesis of the polarized epithelial cell phenotype. Science (Wash DC) 1989;245:718–725. doi: 10.1126/science.2672330. [DOI] [PubMed] [Google Scholar]
- Schneeberger EE, Lynch RD. Structure, function. and regulation of cellular tight junctions. Am J Physiol. 1992;262:L647–L661. doi: 10.1152/ajplung.1992.262.6.L647. [DOI] [PubMed] [Google Scholar]
- Stevenson BR, Siliciano JD, Mooseker MS, Goodenough DA. Identification of ZO-1: a high molecular weight polypeptide associated with the tight junction (zonula occludens) in a variety of epithelia. J Cell Biol. 1986;103:755–766. doi: 10.1083/jcb.103.3.755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takeichi M. Functional correlation between cell adhesive properties and some cell surface proteins. J Cell Biol. 1977;75:464–474. doi: 10.1083/jcb.75.2.464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takeichi M. The cadherins: cell-cell adhesion molecules controlling animal morphogenesis. Development (Camb) 1988;102:639–655. doi: 10.1242/dev.102.4.639. [DOI] [PubMed] [Google Scholar]
- Takeichi M. Cadherin cell adhesion receptors as a morphogenetic regulator. Science (Wash DC) 1991;251:1451–1455. doi: 10.1126/science.2006419. [DOI] [PubMed] [Google Scholar]
- Tsukita Sh, Tsukita Sa. Isolation of cell-to-cell adherens junctions from rat liver. J Cell Biol. 1989;108:31–41. doi: 10.1083/jcb.108.1.31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsukita Sh, Tsukita Sa, Nagafuchi A, Yonemura S. Molecular linkage between cadherins and actin filaments in cell-cell adherens junctions. Curr Opin Cell Biol. 1992;4:834–839. doi: 10.1016/0955-0674(92)90108-o. [DOI] [PubMed] [Google Scholar]
- Tsukita Sh, Itoh M, Nagafuchi A, Yonemura S, Tsukita Sa. Submembranous junctional plaque proteins include potential tumor suppresser molecules. J Cell Biol. 1993;123:1049–1053. doi: 10.1083/jcb.123.5.1049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volk T, Geiger B. A 135-kd membrane protein of intercellular adherens junctions. EMBO (Eur Mol Biol Organ) J. 1984;3:2249–2260. doi: 10.1002/j.1460-2075.1984.tb02123.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willott E, Balda MS, Heintzelman M, Jameson B, Anderson JM. Localization and differential expression of two isoforms of the tight junction protein ZO-1. Am J Physiol. 1992;262:C1119–1124. doi: 10.1152/ajpcell.1992.262.5.C1119. [DOI] [PubMed] [Google Scholar]
- Willott E, Balda MS, Fanning AS, Jameson B, Van Itallie C, Anderson JM. The tight junction protein ZO-1 is homologous to the Drosophiladiscs-large tumor suppresser protein of septate junctions. Proc Natl Acad Sci USA. 1993;90:7834–7831. doi: 10.1073/pnas.90.16.7834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woods DA, Bryant PJ. ZO-1, DlgA and PSD95/SAP90: Homologous proteins in tight, septate and synaptic cell junctions. Mech Dev. 1993;44:85–89. doi: 10.1016/0925-4773(93)90059-7. [DOI] [PubMed] [Google Scholar]
- Yonemura S, Itoh M, Nagafuchi A, Tsukita Sh. Cell-to-cell adherens junction formation and actin filament organization: similarities and differences between non-polarized fibroblasts and polarized epithelial cells. J Cell Sci. 1995;108:127–142. doi: 10.1242/jcs.108.1.127. [DOI] [PubMed] [Google Scholar]
- Zhong Y, Saitoh T, Minase T, Sawada N, Enomoto K, Mori M. Monoclonal antibody 7H6 reacts with a novel tight junction-associated protein distinct from ZO-1, cingulin and ZO-2. . J Cell Biol. 1993;120:477–483. doi: 10.1083/jcb.120.2.477. [DOI] [PMC free article] [PubMed] [Google Scholar]