

Abstract
Mouse and human macrophages express a plasma membrane receptor for extracellular ATP named P2Z/P2X7. This molecule, recently cloned, is endowed with the intriguing property of forming an aqueous pore that allows transmembrane fluxes of hydrophylic molecules of molecular weight below 900. The physiological function of this receptor is unknown. In a previous study we reported experiments suggesting that the P2Z/P2X7 receptor is involved in the formation of macrophage-derived multinucleated giant cells (MGCs; Falzoni, S., M. Munerati, D. Ferrari, S. Spisani, S. Moretti, and F. Di Virgilio. 1995. J. Clin. Invest. 95:1207– 1216). We have selected several clones of mouse J774 macrophages that are characterized by either high or low expression of the P2Z/P2X7 receptor and named these clones P2Zhyper or P2Zhypo, respectively. P2Zhyper, but not P2Zhypo, cells grown to confluence in culture spontaneously fuse to form MGCs. As previously shown for human macrophages, fusion is inhibited by the P2Z/P2X7 blocker oxidized ATP. MGCs die shortly after fusion through a dramatic process of cytoplasmic sepimentation followed by fragmentation. These observations support our previous hypothesis that the P2Z/P2X7 receptor is involved in macrophage fusion.
Purinergic P2X receptors are emerging as one of the most interesting new families of plasma membrane receptors recently described. Molecular cloning has shown that they are formed by subunits possessing only two probable transmembrane domains, with both the carboxy and amino termini on the cytoplasmic side of the plasma membrane and a central (280 amino acids) extracellular domain, rich in cysteine residues (Brake et al., 1994; Valera et al., 1994; Surprenant et al., 1996). Other plasma membrane receptor families sharing in part this structural motif are the inward rectifying K+ channel (Kir) of insulin-secreting cells, the amiloride-sensitive Na+ channel of epithelial cells, the mechano-sensitive channel of Escherichia coli, and the mechano-sensitive channels of Caenorhabditis elegans (deg-1, mec-4, and mec-10; Corey and Garcia-Anoveros, 1996; North, 1996). Among these, P2X and Kir are the only members that are known to be controlled by a soluble ligand, ATP in the case of P2X and ADP in that of Kir (Surprenant et al., 1995; Nichols et al., 1996).
P2Z/P2X7, the largest receptor/channel of the P2X subfamily (595 amino acids), differs from other members of the P2X subfamily by the presence of a long cytoplasmic carboxy tail that is essential for the pore-forming activity, as elegantly shown by Surprenant et al. (1996). Although P2Z/P2X7 is the plasma membrane molecule that is responsible for the long known but little understood permeabilization of the plasma membrane consequent to stimulation of many cell types with extracellular ATP (Rozengurt et al., 1977; Cockcroft and Gomperts, 1979; Steinberg et al., 1987; Di Virgilio and Steinberg, 1993), the physiological function of this process has remained unknown.
It has been previously suggested that a possible role of P2Z/P2X7 receptor could be in cellular communication, in a gap junction-like fashion (Steinberg et al., 1990; Di Virgilio et al., 1995). Two years ago we provided preliminary evidence in support of this hypothesis by observing that specific blockade of this receptor with oxidized ATP (oATP) almost completely inhibits formation of multinucleated giant cells (MGCs),1 triggered by incubation of human macrophages with Con A and interferon-γ (IFN-γ; Falzoni et al., 1995). Treatment with oATP on the other hand did not affect chemotaxis, cell aggregation, or expression of the adhesion molecules CD11a, CD18, and CD54.
To further investigate the role of P2Z/P2X7 receptor, we have selected J774 macrophage cell clones that express this receptor at different levels, from virtually none (P2Zhypo cells) to very high levels (P2Zhyper cells). Experiments reported in this paper show that P2Zhyper macrophages become exceedingly fragile and susceptible to ATP-mediated cell death and spontaneously fuse during in vitro culture.
Materials and Methods
Cells
J774 mouse macrophages and P2Zhyper and P2Zhypo clones were grown in DME supplemented with 10% heat-inactivated horse serum, penicillin (100 U/ml), and streptomycin (100 μg/ml) (complete DME medium). P2Zhypo variants were selected by repeated rounds of incubation in the presence of 5 mM ATP, followed by cloning by limiting dilution. P2Zhyper variants were obtained by cloning by limiting dilution and selection of the clones that showed a higher ATP-dependent uptake of lucifer yellow. Stable transfectants of HEK293 cells expressing the rat P2X2 or P2X7 receptors were described previously (Evans et al., 1995; Surprenant et al., 1996) and were grown in DME F12 medium supplemented with 10% FCS and 300 μg/ml of G418 (Inalco, Milan, Italy).
Phase Contrast and Fluorescence Microscopy
Phase contrast and fluorescence photographs were taken with an inverted fluorescence microscope (Olympus IMT-2; Olympus Optical Co., Ltd., Tokyo, Japan) equipped with 40 and 100× (oil immersion) objectives and fluorescein and rhodamine filters.
Transmission Electron Microscopy
Cell monolayers were fixed in 2.5% glutaraldehyde in 0.1 M cacodylate buffer (pH 7.2–7.4) and post fixed in 1% OsO4 in the same buffer. Samples were then dehydrated and embedded in Araldite Durcupan (Fluka Chemie AG, Buchs, Switzerland). Blocks were cut with a microtome (Ultracut S; Reichert, Vienna, Austria), and ultra-thin sections were stained with uranyl acetate and lead citrate with an Ultrostainer (Reichert). Examination was performed in a transmission electron microscope (H-800; Hitachi Instr., San Jose, CA).
In Situ Hybridization
J774 mouse macrophages, P2Zhyper, and P2Zhypo clones were allowed to adhere to polylysine-coated coverslides for 2 h. Preparations were fixed with 3% paraformaldehyde and incubated with either sense or anti-sense digoxigenin-UTP–labeled cRNA probes at a probe concentration of 50 ng/ml (Schaeren-Wiemers and Gerfin-Moser, 1993). After 16 h at 72°C, hybridization was detected with an anti-digoxigenin antibody conjugated to alkaline phosphatase (Boehringer Mannheim, Indianapolis IN) using 4-nitroblue tetrazolium chloride and 5-bromo–4-chloro-3-indolyphosphate (Boehringer Mannheim). Results are shown for an 18-h phosphatase reaction. The P2X7 riboprobe corresponds to nucleotides 1,150– 1,900 of the corresponding cDNA.
Western Blotting
Cells were detached from the tissue culture using a scraper and were resuspended in PBS in the presence of protease inhibitors (phenylmethylsulfonyl fluoride, 4 mM; pepstatin, 2 mg/ml; leupeptin, 2 mg/ml; trypsin inhibitor, 2 mg/ml; aprotinin, 2 mg/ml). Cells were freeze thawed three times, alternating dry ice and 37°C water bath, and membranes were pelleted. Proteins were extracted in lysis buffer (10 mM Tris-HCl, pH 8.5, 150 mM NaCl, 1 mM CaCl2, 2% Triton X-100) in the presence of protease inhibitors. The extracts were centrifuged at 40,000 rpm to remove insoluble material. Equal concentrations of protein extracts were separated on an 8% polyacrylamide gel in the presence of SDS and transferred to nitrocellulose membrane.
Nitrocellulose membranes were incubated with primary antibody followed by peroxidase-conjugated swine anti–rabbit immunoglobulins (DAKO Spa, Milan, Italy) and developed using the chemiluminescent substrate ECL (Amersham Intl., Little Chalfont, UK).
Antibody
The rabbit polyclonal anti-P2X7 antiserum was raised against the synthetic peptide corresponding to the last 20 amino acids of the rat P2X7 protein (Collo, G., S. Neidhart, E. Kawashima, M. Kosco, R.A. North, and G. Buell, manuscript submitted for publication).
Results
Phenotypic Characterization of P2Zhyper and P2Zhypo Cells
P2Z/P2X7 receptor function can be demonstrated by the ATP-mediated uptake of extracellular hydrophilic fluorescent markers such as lucifer yellow or YO-PRO. Fig. 1 shows that incubation in the presence of 3 mM ATP triggered a massive uptake of lucifer yellow by P2Zhyper cells (Fig. 1, A and D), while on the contrary almost no uptake by P2Zhypo cells was detectable (Fig. 1, C and F). P2Zwt showed an intermediate behavior (Fig. 1, B and E). Nearly all P2Zhyper cells were positive for lucifer yellow uptake to varying levels. Because the sustained activation of the P2Z/P2X7 receptor leads to cell death, we tested for the release of the cytoplasmic enzyme lactic dehydrogenase in response to ATP. Whereas P2Zhypo were unaffected, P2Zhyper rapidly released lactic dehydrogenase, and P2Zwt again showed an intermediate behavior (Fig. 2).
Figure 1.
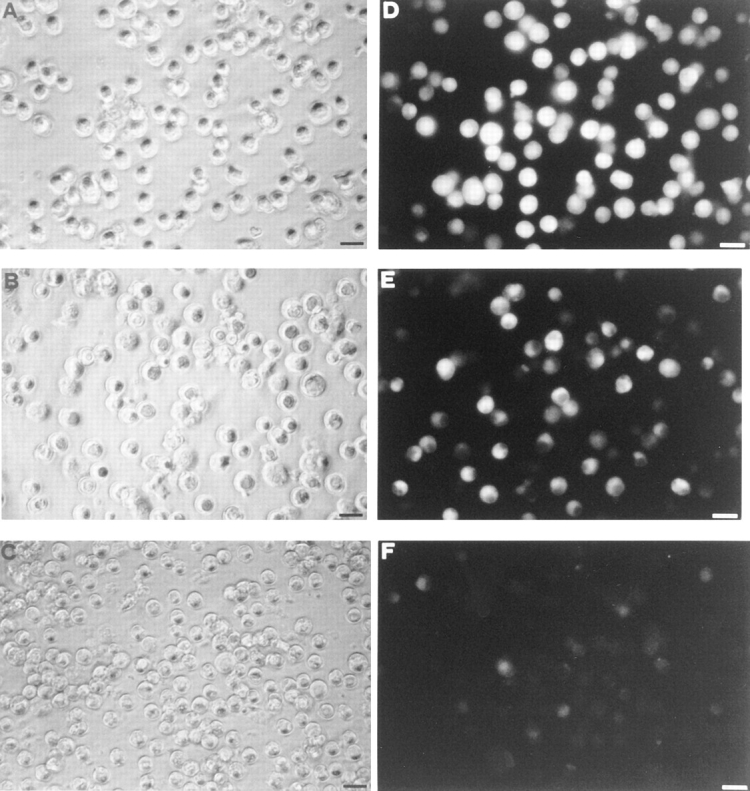
Enhancement of ATP-dependent lucifer yellow uptake by P2Zhyper cells. Macrophages were incubated in complete RPMI medium at 37°C and stimulated with 3 mM ATP in the presence of 1 mg/ml of lucifer yellow for 15 min. After this incubation time, they were rinsed several times with complete RPMI medium and photographed with a fluorescence microscope (40× objective). (A and D) P2Zhyper; (B and E) P2Zwt; (C and F) P2Zhypo. Bars, 25 μm.
Figure 2.

Differential sensitivity to ATP-mediated cytotoxicity of P2Zhyper, P2Zwt, and P2Zhypo macrophages. Macrophages were incubated in serum-free RPMI medium at 37°C at a concentration of 5 × 105/ml and stimulated with 3 mM ATP for 6 h. Releases of lactic dehydrogenase (LDH) from two clones of P2Zhyper cells (□ □, ♦ ♦), two clones of P2Zhypo cells (⋄ ⋄, ▴ ▴), and P2Zwt (▪ ▪) are shown.
P2Zhyper cells overexpress the P2Z/P2X7 mRNA and protein as seen by in situ hybridization (Fig. 3) and western (Fig. 4) blot analysis. P2Zhyper macrophages (Fig. 3, A) show a very strong reactivity to the specific P2X7 anti-sense riboprobe, while P2Zhypo cells (Fig. 3 B) have little, if any, reactivity with the notable exception of one highly reactive cell. J774 wild-type cells (Fig. 3 C) show an intermediate reactivity. A stable transfectant (HEK293 cells) for rat P2X7 is shown for comparison (Fig. 3 D). On average, we found that in the P2Zhyper population ∼83 ± 5% of the cells were strongly positive, as compared to ∼65 ± 8% of positive cells in the J774 wild-type population and only 2–3% of positive cells in the P2Zhypo macrophages. No hybridization was detected with the sense riboprobe (not shown). Fig. 4 shows that the specific polyclonal Ab raised against the carboxy-terminal tail of the receptor recognized in J774 wild-type cells a protein of the expected molecular weight of ∼72. This band was stronger in P2Zhyper cells and absent in P2Zhypo and control cells.
Figure 3.
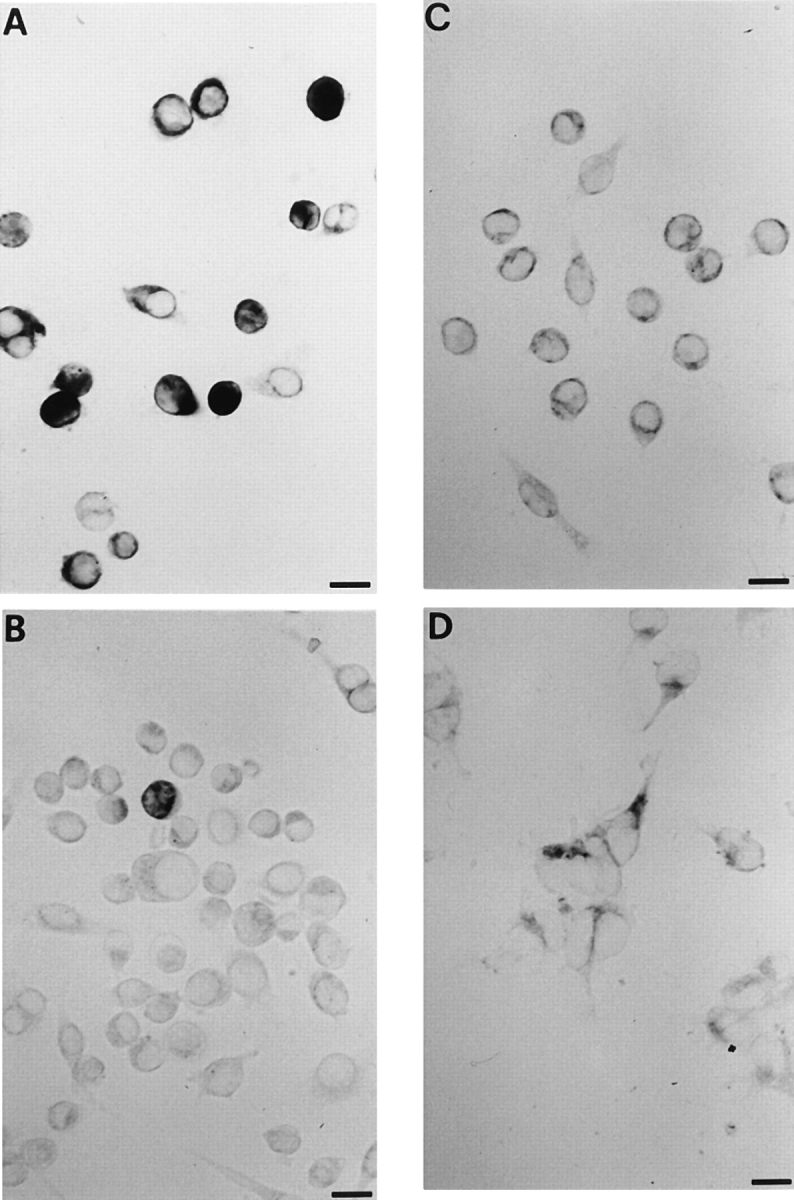
P2X7 RNA expression in J774 mouse macrophages, P2Zhyper, and P2Zhypo clones. P2Zhyper and P2Zhypo clones, wild-type J774 macrophages, and HEK293 cells stably transfected with P2X7 cDNA were incubated with digoxigenin antisense riboprobe (A–D, respectively). Bars, 10 μm.
Figure 4.

Western blot analysis with a specific anti-P2X7 antiserum. Proteins (30 μg/lane) extracted from J774 mouse macrophages, P2Zhyper, and P2Zhypo clones were separated by electrophoresis and transferred to nitrocellulose. P2X7 was visualized by incubation with the P2X7 specific antibody, followed by peroxidase-conjugated secondary antibody and chemiluminescent substratum. Stable P2X7 refers to proteins extracted from HEK293 cells stably transfected with P2X7 (positive control). Negative control refers to proteins extracted from HEK293 cells stably transfected with P2X2.
Spontaneous Cell Fusion in P2Zhyper Monolayers
P2Zhyper cells were very fragile and easily died upon reaching confluence. Nonetheless, in many cultures we observed that shortly after reaching confluence (usually 3 d after plating), macrophages formed dense aggregates in which MGCs were easily detected (Fig. 5). During the last two years we have selected >20 P2Zhyper cell clones, and invariably, although to a different extent, we have observed formation of MGCs in all these cell populations. MGCs had different shapes: round, with short pseudopods (Fig. 5 A); star-like, with dendritic-like elongations (Fig. 5 B); polygonal, with a regular cellular contour (Fig. 5 C). MGCs contained from 2 to >10 large nuclei usually located in the center and were also readily observed in monolayers of HEK293 cells stably transfected with the P2X7 receptor cDNA (Fig. 5 D); on the contrary, fused cells were never observed in monolayers of P2Zhypo or P2Zwt macrophages (Fig. 6). In principle, MGCs could originate either from a real membrane fusion event or from kariokinesis without cell division. To solve this issue, we pooled two batches of P2Zhyper cells that had been previously labeled with different endosome–lysosome markers, lucifer yellow, and Texas red. Real fusion events were detected by observing both yellow/green and red vesicles within the same MGC. Two examples of such fusion are shown by arrowheads, which identify MGCs of different shapes in Fig. 7. On the contrary, individual macrophages are labeled by either the yellow/green or red tracer (Fig. 7, closed triangles). Obviously, cell fusion can also occur between macrophages labeled with the same tracer, such as the spindle-shaped MGC visible in the left-hand side of Fig. 7 A (closed diamond), that is stained only by Texas red.
Figure 5.
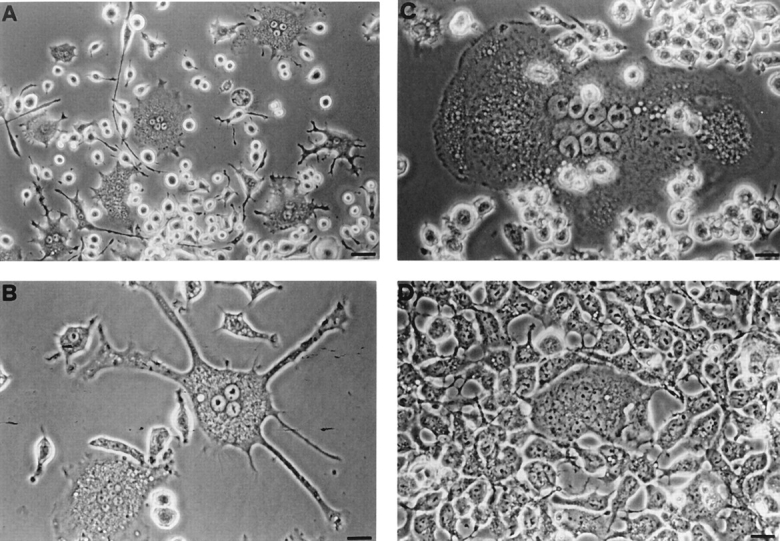
Spontaneous formation of multinucleated giant cells in monolayers of P2Zhyper cells and HEK293 cells transfected with the P2X7 receptor. Cells were plated at the concentration of 5 × 105 cells/ml and grown for 3 d. (A–C) P2Zhyper macrophages; (D) HEK293 cells stably transfected with the P2X7 receptor. Photographs were taken with a 20× objective in A and a 40× objective in B–D. Bars: (A) 50 μm; (B–D) 25 μm.
Figure 6.
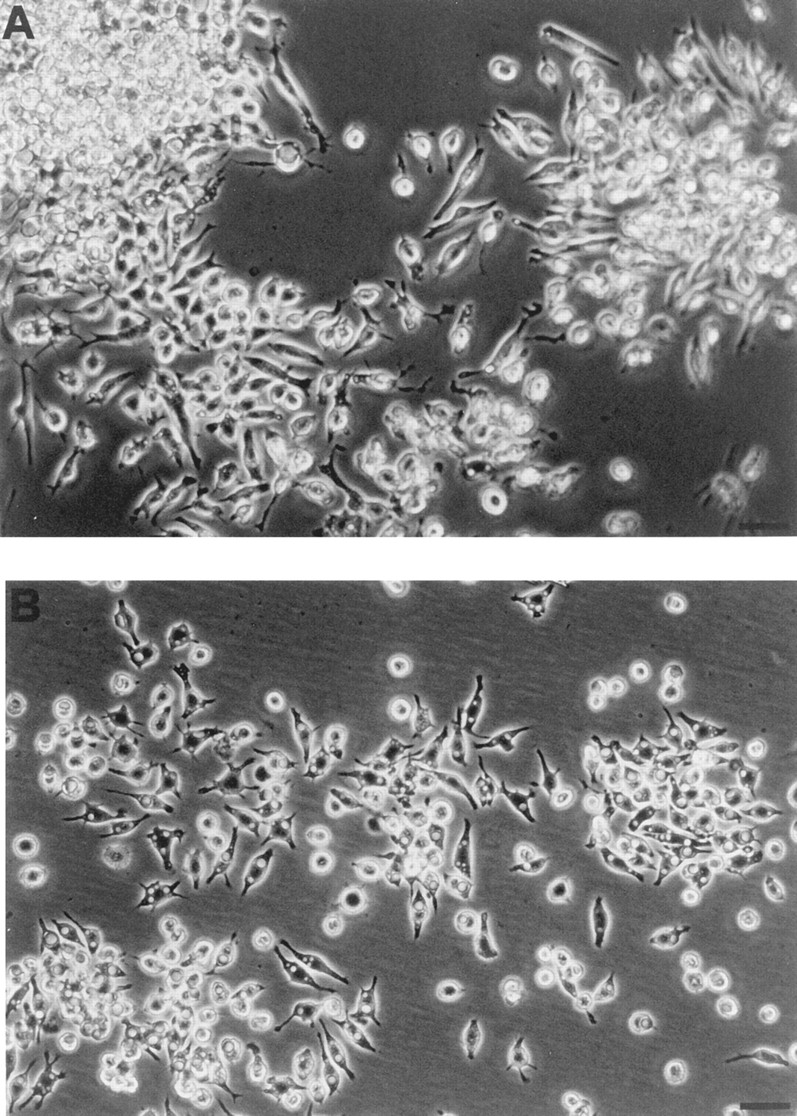
Lack of formation of multinucleated giant cells in monolayers of P2Zwt and P2Zhypo macrophages. P2Zhypo (A) and P2Zwt (B) macrophages were plated as described in Fig. 5 and photographed with a 20× objective. Bar, 50 μm.
Figure 7.
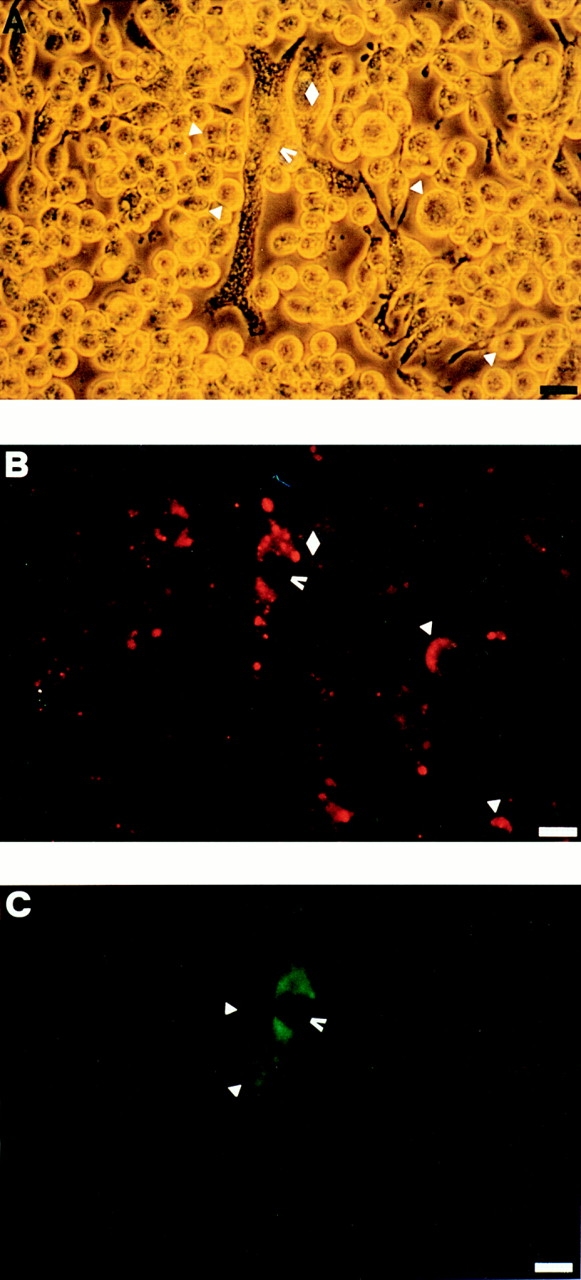
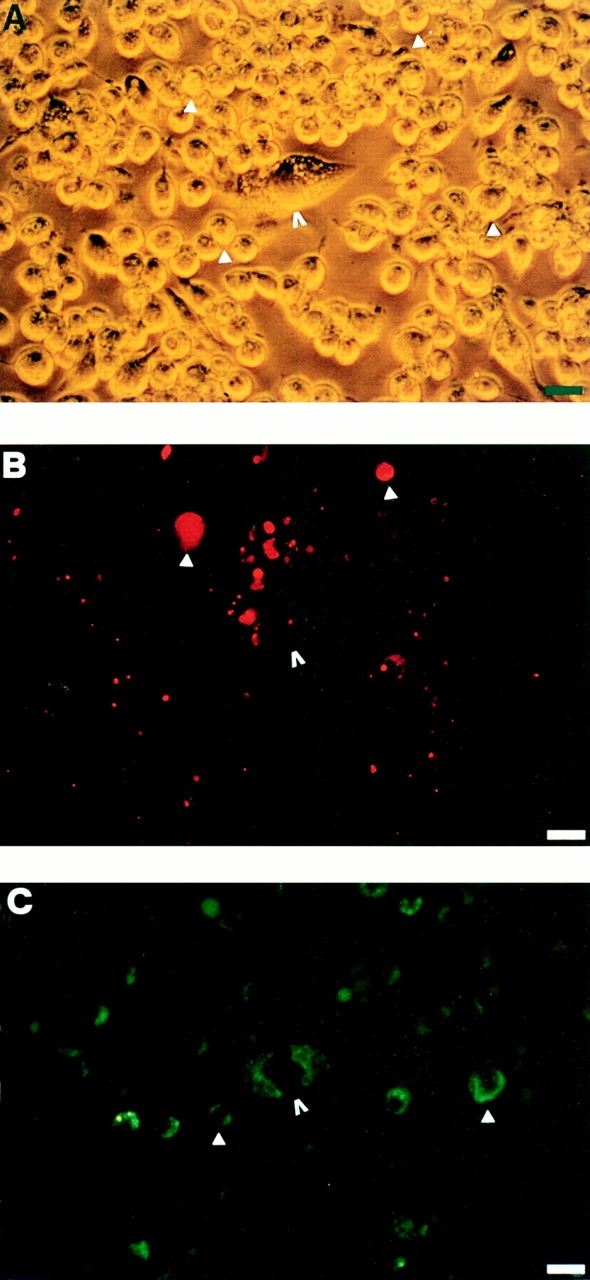
Double staining of multinucleated giant cells with lucifer yellow and Texas red. Two batches of P2Zhyper cells were separately allowed to pinocytose lucifer yellow (10 mg/ml) or Texas red (1 mg/ml) for 1 h at 37°C and then rinsed several times with complete RPMI medium, mixed together, and layered in 24-well dishes at the concentration of 5 × 105/well. After 3 d, the cultures were examined for formation of MGCs and photographed with a fluorescence microscope. (A) Phase contrast; (B) rhodamine filter; (C) fluorescein filter. Bars, 25 μm.
P2Zhyper macrophages formed cell aggregates that preceeded fusion. It was very common to find within such aggregates, images suggestive of initial cytoplasmic communication between adjacent cells (Fig. 8, A and B, arrows). Most often, an already-formed MGC was itself a center of aggregation to which other cells were attracted up to the eventual fusion (Fig. 8 C). We examined some of these aggregates by transmission electron microscopy. As shown in Fig. 9, there were sites of close apposition between membranes of adjacent cells, and sometimes the juxtaposed plasma membranes were about to fuse (Fig. 9, arrows).
Figure 8.
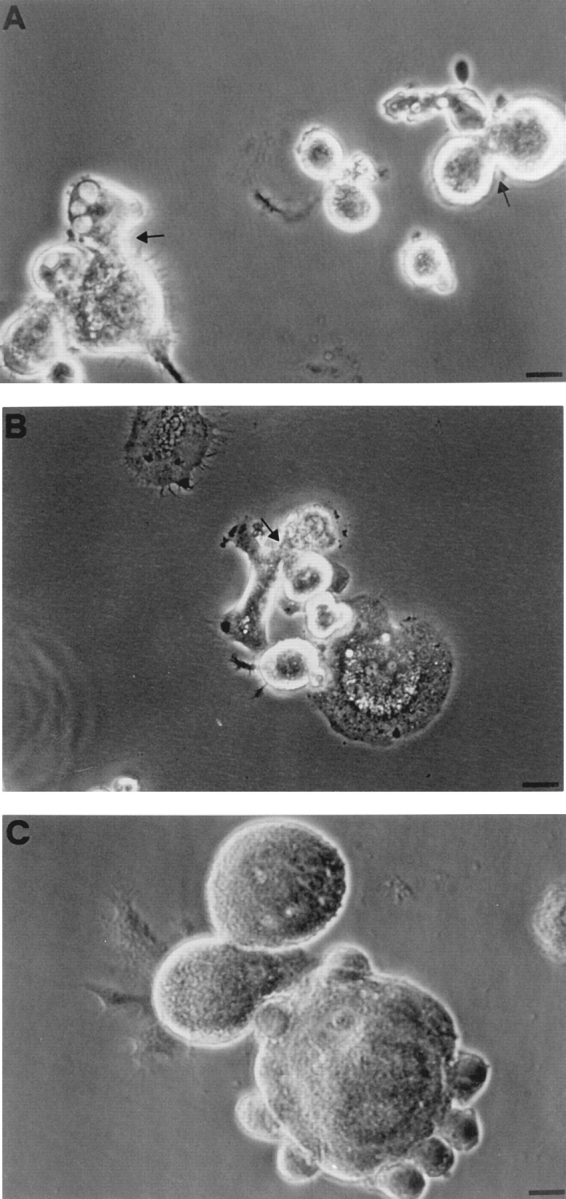
Early cellular interactions in cultures of P2Zhyper macrophages. Experimental conditions as in Fig. 5.
Figure 9.
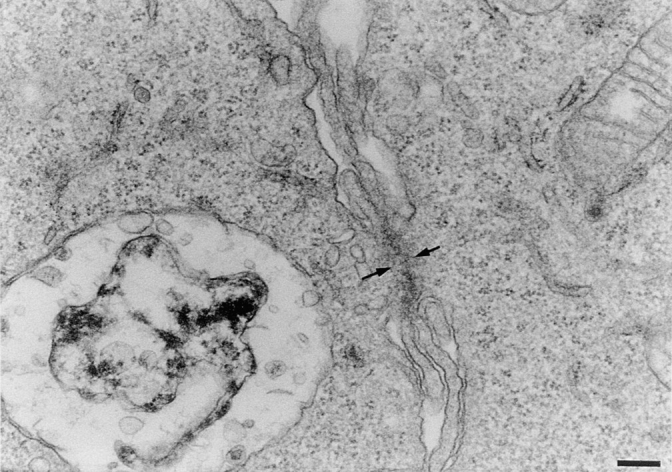
Detail of sites of close cellular interaction in cultures of P2Zhyper macrophages. Macrophage monolayers were incubated as described in Fig. 5, fixed, and processed for electron microscopy, as described in Materials and Methods. Bar, 0.17 μm.
Our group introduced a few years ago the only available inhibitor of P2Z/P2X7 receptor, the dialdehyde compound oATP (Murgia et al., 1993). This reagent covalently binds to the receptor and causes an irreversible inactivation. Pretreatment of P2Zhyper cells with oATP for 2 h completely blocked spontaneous MGC formation without inhibiting aggregation (Fig. 10). Similar inhibition has been reported for monocyte-derived human macrophages (Falzoni et al., 1995). It has been recently suggested (Baricordi et al., 1996; Buell et al., 1996) that during in vitro culture, immune cell P2X receptors may be in a state of chronic desensitization due to a continuous leakage of ATP into the exracellular milieu. This hypothesis has received strong support from the recent demonstration that macrophages and microglial cells can release ATP via a nonlytic mechanism (Ferrari et al., 1997). Therefore, we tested the effect of the ATP-consuming enzyme hexokinase on MGC formation in P2Zhyper monolayers. Treatment with 100 μg/ ml of hexokinase almost doubled the number of MGCs, from 6 ± 3 to 12 ± 2/field, as quantitated by observing 4 randomly selected microscopic fields from 2 different wells for each experimental condition, i.e., control and hexokinase-treated monolayers.
Figure 10.
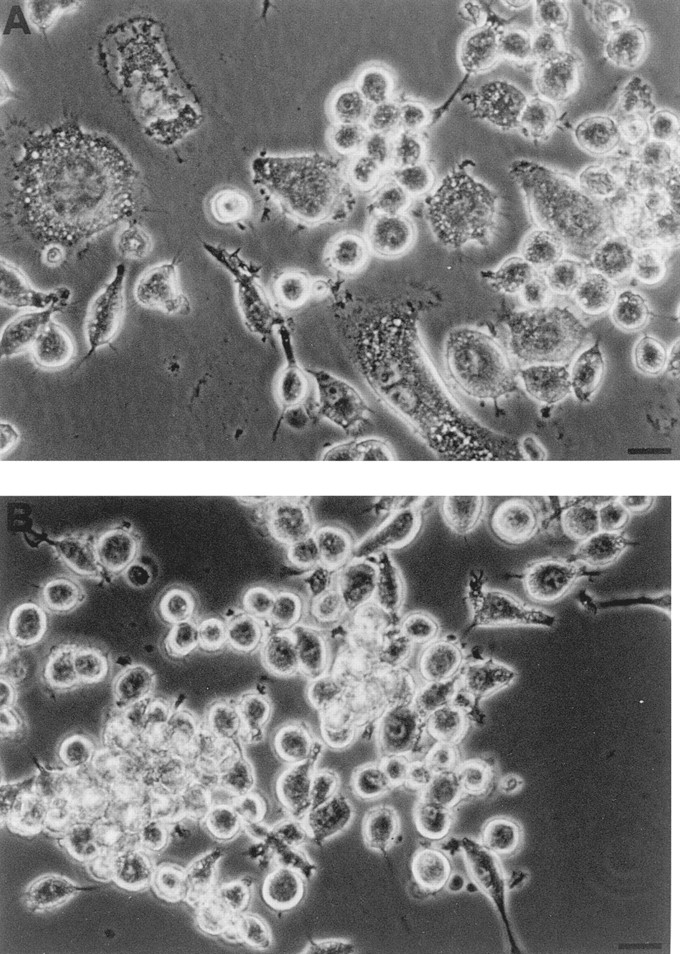
Inhibition of multinucleated giant cell formation by oxidized ATP. Macrophages from the same batch of P2Zhyper cells were plated in 24-well dishes at a concentration of 5 × 105/ ml. 6 h after the plating, cells shown in B were treated with 300 μM oATP for 2 h and then rinsed thoroughly and supplemented with fresh, complete RPMI medium.
An interesting question is the fate of MGCs. Within 24–36 h after formation, MGCs started to deteriorate: cytoplasmic vacuoles appeared, nuclei condensed, and finally the cell body fragmented (Fig. 11, A-D). An MGC's fate did not depend on the age of the culture but rather on that of the individual MGC; no MGC survived longer than 72 h after fusion.
Figure 11.

Multinucleated giant cells die after fusion. P2Zhyper macrophages were plated as described in Fig. 5. Sequential pictures were taken from the same field at ∼12-h interval.
Discussion
It is increasingly appreciated that ATP is an important mediator of intercellular communication (Burnstock, 1972, 1996; Zimmermann, 1994; Di Virgilio et al., 1996). Biological effects of ATP as an extracellular messenger are mediated through P2 purinergic receptors. Macrophages express both G protein–coupled P2Y and intrinsic ion channel P2X receptors (Greenberg et al., 1988). An intriguing P2X receptor expressed by macrophages is P2Z/ P2X7, whose most characteristic property is to form an aqueous pore that, depending on the ATP dose and frequency of stimulation, forms an ion channel or a non– selective pore that admits hydrophylic molecules of molecular mass up to 900 D (Steinberg et al., 1987; Di Virgilio et al., 1995). This receptor is specifically activated by ATP, probably in its ATP4− form, like the ATP-gated pore of mast cells (Cockcroft and Gomperts, 1979), and inhibited by the dialdehyde derivative oATP (Murgia et al., 1993). The only selective agonist, the ATP derivative benzoylbenzoic ATP, is about one order of magnitude more potent than ATP (Nuttle and Dubyak, 1994).
The only well characterized and generally accepted effect consequent to P2Z/P2X7 stimulation is cell death (Murgia et al., 1992; Molloy et al., 1994; Blanchard et al., 1995). Opening of the P2Z/P2X7 pore causes large transmembrane ion fluxes leading to Na+ and Ca2+ overflooding and K+ depletion accompanied by loss of cytoplasmic low molecular weight metabolites (Steinberg and Silverstein, 1987). Short permeabilizations (up to 10–15 min) are in general harmless, in that once extracellular ATP is removed, the pore reseals, and intracellular ion homeostasis is re-established. However, a sustained activation is invariably followed by cell death by either necrosis or apoptosis, depending on the length of the stimulation and the ATP concentration (Murgia et al., 1992; Molloy et al., 1994).
Why macrophages and a few other cell types should express such a dangerous receptor is not clear. It might be assumed that the receptor is never active, because in the body the extracellular ATP concentrations necessary for its opening (hundreds of micromolar or millimolar) are never reached. However, this is clearly not the case (Hoyle and Burnstock, 1996). More and more cell types are shown to be capable of releasing ATP via a number of different mechanisms. The most straightforward is exocytotic release of granular ATP by platelets, adrenal chromaffine cells, or neurons (Born and Kratzer, 1984; Zimmermann, 1994), but nonexocytotic mechanisms have also been implicated (Pearson and Gordon, 1979; Gordon, 1990). For example, some membrane transporters such as P-glycoprotein and the CFTR protein have been proposed to carry ATP as well as other smaller ions (Abraham et al., 1993; Reisin et al., 1994; Schwiebert et al., 1995; but see Reddy et al., 1996, for contrasting results). In addition, because cytoplasmic ATP concentration is in the 5–10 mM range, any damage of the plasma membrane is likely to cause substantial release of ATP in the extracellular milieu. Events able to release ATP can range from mild injuries such as shear stress applied to the vessel wall to outright necrotic lesions. Under normal physiological conditions these events could be uncommon, but at sites of acute or chronic inflammatory reactions they might be frequent and thus contribute significantly to the accumulation of high concentrations of extracellular ATP.
Our lab and others originally proposed that the P2Z/ P2X7 receptor could have a role as a suicide receptor exploited by the immune system to eliminate unwanted cells and more generally down regulate the immune response (Di Virgilio et al., 1989, 1990; Filippini et al., 1990). This hypothesis is clearly very speculative, but a number of groups have already provided data in its support (Blanchard et al., 1991; Modderman et al., 1994; Molloy et al., 1994). However, it cannot be excluded that this receptor is also endowed with other functions not related to cytotoxicity. The P2Z/P2X7 receptor is expressed during macrophage differentiation and is upregulated by IFN-γ (Blanchard et al., 1991; Hickman et al., 1994; Falzoni et al., 1995), a cytokine that is massively released during the immune and inflammatory responses. Under these conditions, macrophages are known to establish close contact with each other and with different cell types, such as helper or cytotoxic lymphocytes, and to release several cytokines. Thus, it can be hypothesized that the P2Z/P2X7 receptor could be involved in intercellular communication, maybe to help establish bridges between the cytoplasm of adjacent macrophages, in a gap junction-like fashion. This process might contribute to formation of MGCs, a typical histologic feature of granulomatous inflammation. MGCs are known to arise from the fusion of adjacent macrophages and not from endomitosis (i.e., nuclear division without cytoplasmic division; McInnes and Rennick, 1988; Takashima et al., 1993), but the mechanism and the membrane molecules involved are still unknown. Several cytokines (e.g., IFN-γ, TNF-α, IL-3, and IL-4) have been implicated in different stages of MGC formation, but no clear conclusion has been reached about the identity of the fusogenic cytokine (Weinberg et al., 1984; Most et al., 1990). Molecules of the LFA-1 family might be involved, but again no firm evidence has been provided (Most et al., 1990).
J774 macrophage cells have been instrumental in the identification and characterization of the P2Z/P2X7 receptor. Steinberg and Silverstein performed a thorough investigation of the pharmacological and physiological properties of this molecule and introduced the ATP-resistant clones that became an invaluable tool at a time in which the cDNA was not available (Steinberg and Silverstein, 1987; Steinberg et al., 1987; Greenberg et al., 1988). We have in our laboratory ATP-resistant clones that were selected >10 yr ago. The cellular basis for the ability to select stable ATP-resistant cell clones rests on the marked heterogeneity of the wild-type J774 population with respect to P2Z/P2X7 expression. In general, susceptibility to ATP-mediated permeabilization of J774 cells assessed by uptake of hydrophylic fluorescent markers ranges from zero (dark cells) to very high (bright cells), with the full range of intermediate conditions. Very bright J774 cells can be selected, by limiting dilution, and clonally expanded. As previously shown by our group (Chiozzi et al., 1996), J774 hyper clones thus selected not only undergo a massive ATP-dependent lucifer yellow uptake but also cell death, confirming that high expression of the P2Z/ P2X7 receptor confers an unusual sensitivity to ATP cytotoxic effects. P2Zhyper macrophages turned out to be very delicate cells to grow in culture, very probably as a direct consequence of high expression of the P2Z/P2X7 receptor, but very interestingly they also underwent a high rate of spontaneous fusion after 2 or 3 d of in vitro culture. Although fusion was at its peak 3 d after plating, scattered MGCs could be observed even 1 d after seeding. Furthermore, HEK293 cells transfected with P2X7 cDNA, but not the wild-type control population, also showed several MGCs. These observations suggest that the mere hyperexpression of P2Z/P2X7 receptor may be sufficient to drive fusion in the absence of secreted or diffusible factors. High rate of spontaneous fusion correlated with high expression of P2Z/P2X7 mRNA and reactivity with specific Abs; furthermore, fusion was abolished by oATP, the only selective inhihitor so far available. These observations, together with earlier convergent reports from our laboratory (Falzoni et al., 1995), point to a crucial role of this receptor in macrophage fusion, although they do not allow us to pinpoint exactly at what stage of the fusion process the receptor is involved.
Once formed, MGCs have a very short life. Within a few hours the cytoplasm becomes filled with phase-lucent vacuoles, fuzzy, and fragmented. At the end of this process, only a shadow of the original MGCs is left. Adjacent, nonfused macrophages on the contrary continue to grow normally, thus suggesting that it is the fusion itself that drives the cells along a terminal pathway.
This report provides further evidence in support of the hypothesis that the pore-forming P2Z/P2X7 receptor participates in a crucial step of macrophage fusion leading to MGCs formation, a role of potential interest in the evolution of chronic granulomatous inflammation.
Acknowledgments
This work was supported by the Italian Ministry for Scientific Research (40% and 60%), the National Research Council of Italy (Target Project Clinical Applications of Cancer Research, ACRO), the Italian Association for Cancer Research (AIRC), the IX AIDS Project, the I Tuberculosis Project, and Telethon of Italy. J.M. Sanz is supported by a fellowship awarded by the Spanish Ministry for Education and Science.
Abbreviations used in this paper
- IFN-γ
interferon-γ
- MGC
multinucleated giant cells
Footnotes
The help of Dr. Sybille Neidhart is gratefully acknowledged. We also thank Dr. Annmarie Surprenant for the kind gift of HEK293 cells stably transfected with P2X2 cDNA.
Please address all correspondence to Dr. Francesco Di Virgilio, Institute of General Pathology, University of Ferrara, Via Borsari, 4, I-44100 Ferrara, Italy. Tel.:(39)532-291-353; Fax:(39)532-247-278; E-mail: fdv@ifeuniv.unife.it
References
- Abraham EH, Prat AG, Gerweck L, Seneveratne T, Arceci RJ, Kramer R, Guidotti G, Cantiello HF. The multidrug resistance (mdr1)gene product functions as an ATP channel. Proc Natl Acad Sci USA. 1993;90:312–316. doi: 10.1073/pnas.90.1.312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baricordi, O., D. Ferrari, L. Melchiorri, P. Chiozzi, S. Hanau, E. Chiari, M. Rubini, and F. Di Virgilio. 1996. An ATP-activated channel is involved in mitogenic stimulation of human T lymphocytes. Blood, 87:682–690. [PubMed]
- Blanchard DK, McMillen S, Djeu JY. IFN-γ enhances sensitivity of human macrophages to extracellular ATP-mediated lysis. J Immunol. 1991;147:2579–2585. [PubMed] [Google Scholar]
- Blanchard DK, Wei S, Duan C, Pericle F, Diaz JI, Djeu JY. Role of extracellular adenosine triphosphate in the cytotoxic T-lymphocyte-mediated lysis of antigen presenting cells. Blood. 1995;11:3173–3182. [PubMed] [Google Scholar]
- Born GVR, Kratzer MAA. Source and concentration of extracellular adenosine triphosphate during haemostasis in rats, rabbits, and man. J Physiol. 1984;354:419–429. doi: 10.1113/jphysiol.1984.sp015385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brake AJ, Wagenbach MJ, Julius D. New structural motif for ligand-gated ion channels defined by an ionotropic ATP receptor. Nature (Lond) 1994;371:519–523. doi: 10.1038/371519a0. [DOI] [PubMed] [Google Scholar]
- Buell, G., A.D. Michel, C. Lewis, G. Collo, P.P.A. Humphrey, and A. Surprenant. 1996. P2X1 receptor activation in HL60 cells. Blood, 87:2659–2664. [PubMed]
- Burnstock G. Purinergic nerves. Pharmacol Rev. 1972;24:509–560. [PubMed] [Google Scholar]
- Burnstock G. P2 purinoceptors: historical perspective and classification. Ciba Found Symp. 1996;198:1–34. doi: 10.1002/9780470514900.ch1. [DOI] [PubMed] [Google Scholar]
- Chiozzi P, Murgia M, Falzoni S, Ferrari D, Di Virgilio F. Role of the purinergic P2Z receptor in spontaneous cell death in J774 macrophage cultures. Biochem Biophys Res Commun. 1996;218:176–181. doi: 10.1006/bbrc.1996.0031. [DOI] [PubMed] [Google Scholar]
- Cockcroft S, Gomperts BD. ATP induces nucleotide permeability in rat mast cells. Nature (Lond) 1979;279:541–542. doi: 10.1038/279541a0. [DOI] [PubMed] [Google Scholar]
- Cockcroft S, Gomperts BD. The ATP4−receptor of rat mast cells. Biochem J. 1980;188:789–798. doi: 10.1042/bj1880789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corey DP, Garcia-Anoveros J. Mechanosensation and the DEG/ ENaC ion channels. Science (Wash DC) 1996;273:323–324. doi: 10.1126/science.273.5273.323. [DOI] [PubMed] [Google Scholar]
- Di Virgilio F. The P2Z purinoceptor: an intriguing role in immunity, inflammation and cell death. Immunol Today. 1995;16:524–528. doi: 10.1016/0167-5699(95)80045-X. [DOI] [PubMed] [Google Scholar]
- Di Virgilio, F., and T.H. Steinberg. 1993. ATP permeabilization. In Cell and Tissue Culture: Laboratory Procedures. Wiley & Sons Inc., New York. 5B: 4.1–4.5.
- Di Virgilio F, Bronte V, Collavo D, Zanovello P. Responses of mouse lymphocytes to extracellular adenosine 5′-triphosphate (ATP). Lymphocytes with cytotoxic activity are resistant to the permeabilizing effects of ATP. J Immunol. 1989;143:1955–1960. [PubMed] [Google Scholar]
- Di Virgilio F, Pizzo P, Zanovello P, Bronte V, Collavo D. Extracellular ATP as a possible mediator of cell-mediated cytotoxicity. Immunol Today. 1990;11:274–277. doi: 10.1016/0167-5699(90)90111-l. [DOI] [PubMed] [Google Scholar]
- Di Virgilio, F., D. Ferrari, M. Munerati, S. Falzoni, M. Villalba, V. Bronte, A. Zambon, P. Zanovello, and T.H. Steinberg. 1995. The P2Z receptor and its regulation of macrophage function. In Adenosine and Adenine Nucleotides: From Molecular Biology to Integrative Physiology. L. Belardinelli and A. Pelleg, editors Kluver Academic Publisher, Boston, MA. 329–335.
- Di Virgilio F, Ferrari D, Falzoni S, Chiozzi P, Munerati M, Steinberg TH, Baricordi OR. P2 purinoceptors in the immune system. Ciba Found Symp. 1996;198:290–305. doi: 10.1002/9780470514900.ch17. [DOI] [PubMed] [Google Scholar]
- Evans RJ, Lewis C, Buell G, North RA, Surprenant A. Pharmacological characterization of heterologously expressed ATP-gated cation channels (P2X-purinoceptors) Mol Pharmacol. 1995;48:178–183. [PubMed] [Google Scholar]
- Falzoni S, Munerati M, Ferrari D, Spisani S, Moretti S, Di Virgilio F. The purinergic P2Z receptor of human macrophage cells. Characterization and possible physiological role. J Clin Invest. 1995;95:1207–1216. doi: 10.1172/JCI117770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrari D, Chiozzi P, Falzoni S, Hanau S, Di Virgilio F. Purinergic modulation of interleukin-1β release from microglial cells stimulated with bacterial endotoxin. J Exp Med. 1997;185:579–582. doi: 10.1084/jem.185.3.579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Filippini A, Taffs RE, Agui T, Sitkovsky MV. Ecto-ATPase activity in cytolytic lymphocytes. Protection from the cytolytic effects of extracellular ATP. J Biol Chem. 1990;265:334–340. [PubMed] [Google Scholar]
- Gordon JL. The effects of ATP on endothelium. Ann NY Acad Sci. 1990;603:46–52. doi: 10.1111/j.1749-6632.1990.tb37660.x. [DOI] [PubMed] [Google Scholar]
- Greenberg S, Di Virgilio F, Steinberg TH, Silverstein SC. Extracellular nucleotides mediate Ca2+fluxes in J774 macrophages by two distinct mechanisms. J Biol Chem. 1988;263:10337–10343. [PubMed] [Google Scholar]
- Hickman SE, El J, Khoury, Greenberg S, Schieren I, Silverstein SC. P2Z adenosine triphosphate receptor activity in cultured human monocyte-derived macrophages. Blood. 1994;84:2452–2456. [PubMed] [Google Scholar]
- Hoyle, C.H.V., and G. Burnstock. 1996. Purines. In Principles of Medical Biology. E.E. Bittar and N. Bittar, editors JAI Press Inc., Greenwich, CT. 49–75.
- McInnes A, Rennick DM. Interleukin 4 induces cultured monocyte/ macrophages to form giant multinucleated cells. J Exp Med. 1988;167:598–605. doi: 10.1084/jem.167.2.598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Modderman WE, Vrijheid-Lammers T, Lowik CWGM, Nijweide PJ. Removal of hematopoietic cells and macrophages from mouse bone marrow cultures: isolation of fibroblastlike stromal cells. Exp Hematol (NY) 1994;22:194–201. [PubMed] [Google Scholar]
- Molloy A, Loachumroonvorapong P, Kaplan G. Apoptosis, but not necrosis, of infected monocytes is coupled with killing of intracellular bacillus Calmette-Guerin. J Exp Med. 1994;180:1499–1509. doi: 10.1084/jem.180.4.1499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Most J, Neumayer HP, Dierich MP. Cytokine-induced generation of multinucleated giant cells in vitro requires interferon-γ and expression of LFA-1. Eur J Immunol. 1990;20:1661–1667. doi: 10.1002/eji.1830200807. [DOI] [PubMed] [Google Scholar]
- Murgia M, Pizzo P, Steinberg TH, Di Virgilio F. Characterization of the cytotoxic effect of extracellular ATP in J774 mouse macrophages. Biochem J. 1992;288:897–901. doi: 10.1042/bj2880897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murgia M, Hanau S, Pizzo P, Rippa M, Di Virgilio F. Oxidized ATP. An irreversible inhibitor of the macrophage purinergic P2Z receptor. J Biol Chem. 1993;268:8199–8203. [PubMed] [Google Scholar]
- Nichols CG, Shyng S-L, Nestorowicz A, Glaser B, Clement JP, IV, Gonzales G, Aguilar-Bryan L, Permutt MA, Bryan J. Adenosine diphosphate as an intracellular regulator of insulin secretion. Science (Wash DC) 1996;272:1785–1787. doi: 10.1126/science.272.5269.1785. [DOI] [PubMed] [Google Scholar]
- North RA. Families of ion channels with two hydrophobic segments. Curr Opin Cell Biol. 1996;8:474–483. doi: 10.1016/s0955-0674(96)80023-8. [DOI] [PubMed] [Google Scholar]
- Nuttle LC, Dubyak GR. Differential activation of cation channels and non-selective pores by macrophages P2Z purinergic receptors expressed in Xenopousoocytes. J Biol Chem. 1994;269:13988–13996. [PubMed] [Google Scholar]
- Pearson JD, Gordon JL. Vascular endothelial and smooth muscle cells in culture selectively release adenine nucleotides. Nature (Lond) 1979;281:384–386. doi: 10.1038/281384a0. [DOI] [PubMed] [Google Scholar]
- Reddy MM, Quinton PM, Haws C, Wine JJ, Grygorczyk R, Tabcharani JA, Hanrahan JW, Gunderson KL, Kopito RR. Failure of the cystic fibrosis transmembrane conductance regulator to conduct ATP. Science (Wash DC) 1996;271:1876–1879. doi: 10.1126/science.271.5257.1876. [DOI] [PubMed] [Google Scholar]
- Reisin IL, Prat AG, Abraham EH, Amara JF, Gregory RJ, Ausiello DA, Cantiello HF. The cystic fibrosis transmembrane conductance regulator is a dual ATP and chloride channel. J Biol Chem. 1994;32:20584–20591. [PubMed] [Google Scholar]
- Rozengurt E, Heppel LA, Friedberg I. Effect of exogenous ATP on the permeability properties of transformed cultures of mouse cell lines. J Biol Chem. 1977;252:4584–4590. [PubMed] [Google Scholar]
- Schaeren-Wiemers, N., and A. Gerfin-Moser. 1993. A single protocol to detect transcripts of various types and expression levels in neural tissue and cultures cells: in situ hybridization using digoxigenin-labeled cRNA probes. Hystochemistry, 100: 431–440. [DOI] [PubMed]
- Schwiebert EM, Egan ME, Hwang T-H, Fulmer SB, Allen SS, Cutting GR, Guggino WB. CFTR regulates outwardly rectifying chloride channels through an autocrine mechanism involving ATP. Cell. 1995;81:1063–1073. doi: 10.1016/s0092-8674(05)80011-x. [DOI] [PubMed] [Google Scholar]
- Steinberg TH, Silverstein SC. Extracellular ATP4−promotes cation fluxes in the J774 mouse macrophage cell line. J Biol Chem. 1987;262:3118–3122. [PubMed] [Google Scholar]
- Steinberg TH, Newman AS, Swanson JA, Silverstein SC. ATP4−permeabilizes the plasma membrane of mouse macrophages to fluorescent dyes. J Biol Chem. 1987;262:8884–8888. [PubMed] [Google Scholar]
- Steinberg TH, Buisman HP, Greenberg S, Di Virgilio F, Silverstein SC. Effects of extracellular ATP on mononuclear phagocytes. Ann NY Acad Sci. 1990;603:120–129. doi: 10.1111/j.1749-6632.1990.tb37666.x. [DOI] [PubMed] [Google Scholar]
- Surprenant A, Buell G, North RA. P2X receptors bring new structure to ligand-gated ion channels. Trends Neurosci. 1995;18:224–229. doi: 10.1016/0166-2236(95)93907-f. [DOI] [PubMed] [Google Scholar]
- Surprenant A, Rassendren F, Kawashima E, North RA, Buell G. The cytolytic P2Z receptor for extracellular ATP identified as a P2X receptor (P2X7) Science (Wash DC) 1996;272:735–738. doi: 10.1126/science.272.5262.735. [DOI] [PubMed] [Google Scholar]
- Takashima T, Ohnishi K, Tsuyuguchi I, Kishimoto S. Differential regulation of formation of multinucleated giant cells from concanavalin A-stimulated human blood monocytes by IFN-γ and IL-4. J Immunol. 1993;150:3002–3007. [PubMed] [Google Scholar]
- Valera S, Hussy N, Evans RJ, Adami N, North RA, Surprenant A, Buell G. A new class of ligand-gated ion channel defined by P2X receptor for extracellular ATP. Nature (Lond) 1994;371:516–519. doi: 10.1038/371516a0. [DOI] [PubMed] [Google Scholar]
- Weinberg JB, Hobbs MM, Misukonis MA. Recombinant human γ-inteferon induces human monocyte polykarion formation. Proc Natl Acad Sci USA. 1984;81:4554–4560. doi: 10.1073/pnas.81.14.4554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmermann H. Signalling via ATP in the nervous system. Trends Neurosci. 1994;17:420–426. doi: 10.1016/0166-2236(94)90016-7. [DOI] [PubMed] [Google Scholar]