

Abstract
Islets of Langerhans are microorgans scattered throughout the pancreas, and are responsible for synthesizing and secreting pancreatic hormones. While progress has recently been made concerning cell differentiation of the islets of Langerhans, the mechanism controlling islet morphogenesis is not known. It is thought that these islets are formed by mature cell association, first differentiating in the primitive pancreatic epithelium, then migrating in the extracellular matrix, and finally associating into islets of Langerhans. This mechanism suggests that the extracellular matrix has to be degraded for proper islet morphogenesis. We demonstrated in the present study that during rat pancreatic development, matrix metalloproteinase 2 (MMP-2) is activated in vivo between E17 and E19 when islet morphogenesis occurs. We next demonstrated that when E12.5 pancreatic epithelia develop in vitro, MMP-2 is activated in an in vitro model that recapitulates endocrine pancreas development (Miralles, F., P. Czernichow, and R. Scharfmann. 1998. Development. 125: 1017–1024). On the other hand, islet morphogenesis was impaired when MMP-2 activity was inhibited. We next demonstrated that exogenous TGF-β1 positively controls both islet morphogenesis and MMP-2 activity. Finally, we demonstrated that both islet morphogenesis and MMP-2 activation were abolished in the presence of a pan-specific TGF-β neutralizing antibody. Taken together, these observations demonstrate that in vitro, TGF-β is a key activator of pancreatic MMP-2, and that MMP-2 activity is necessary for islet morphogenesis.
Keywords: pancreatic development, islets of Langerhans, morphogenesis, MMP-2, TGF-β1
In the adult rat, the endocrine pancreatic tissue represents 2% of the pancreatic mass. This endocrine tissue is composed of cells associated into spherical microorgans: the islets of Langerhans. These islets (around 5,000 per pancreas) contain a few thousand cells that express insulin (beta cells), glucagon, somatostatin, or pancreatic polypeptides. Beta cells are localized in the center of the islets, while the other endocrine cells are localized in the periphery.
During development, it is thought that precursor cells that will give rise to pancreatic endocrine cells are present in the primitive pancreatic duct epithelium where they differentiate into endocrine cells (30). Once differentiated, these cells will migrate into the interstitial matrix, and finally aggregate to form the islets of Langerhans (9). While progress has recently been made concerning the early steps of endocrine cell differentiation (1, 19, 40, 43), the mechanism leading to islet morphogenesis remains poorly understood. The focus of the present work is islet morphogenesis.
Cell migration of pancreatic endocrine cells through the extracellular matrix (ECM)1 is thought to constitute an essential step in islet morphogenesis (8, 9). In the pancreas as in other organs, the major components of the ECM are essentially collagens, laminins, and fibronectins. Enzymes known to be able to degrade the different components of the ECM are thus possibly implicated in endocrine cell migration.
Matrix metalloproteinases (MMPs) are a large family of related proteolytic enzymes that include collagenases, gelatinases, and stromelysins. These enzymes have been implicated in cancer cell invasion (45). MMPs have also been implicated in the morphogenesis of a large number of organs. Indeed, an appropriate balance between metalloproteinases and their inhibitors (the tissue inhibitors of metalloproteinases [TIMPs]), is necessary for proper morphogenesis of the mammary gland (44), as well as for the branching of salivary glands and ureter buds (21, 27).
Recently we have described an in vitro model that recapitulates the development of the endocrine pancreas (24). In this system, mature endocrine cells differentiate from rat embryonic pancreatic epithelium cultured into tridimensional collagen gels. Interestingly, once differentiated, the endocrine cells organize with the characteristic architecture of islets of Langerhans, with beta cells in the center and endocrine nonbeta cells in the periphery. Thus, in that model, both morphogenesis and cytodifferentiation occur. We have used this in vitro model to investigate the mechanisms that lead to islet morphogenesis during early fetal life. Our study indicates that (a) in vivo, MMP-2 is expressed in the pancreas, and its activity is maximum between E17 and E19 when islet morphogenesis is important; (b) in vitro, MMP-2 activity is necessary for proper islet morphogenesis, and is tightly controlled by TGF-β. Thus TGF-β plays an important role in islet morphogenesis, one of its targets being MMP-2.
Materials and Methods
Animals and Pancreatic Rudiments Dissection
Pregnant Wistar rats were purchased from Janvier breeding center (Le Genet, France). The morning the vaginal plug was discovered was designated as embryonic day 0.5 (E0.5). Pregnant rats at 12.5 d of gestation were killed by cervical dislocation, and the embryos were removed. Their whole digestive tract was isolated and used to dissect the dorsal pancreatic primordium (11). The pancreatic epithelium was prepared as previously described (24). In brief, the dorsal pancreatic rudiment was incubated in a 0.03% collagenase A (Boehringer Mannheim Corp., Meylan, France) solution in RPMI 1640 synthetic medium (GIBCO BRL, Eragny, France) for 1 h at 37°C (18). The enzymatic digestion was stopped by a 30-min incubation in a 1:1 mixture of RPMI 1640 medium and FCS. Thereafter, the dorsal pancreatic epithelial bulge was freed of its surrounding mesenchyme using tungsten needles.
Culture of the Dorsal Pancreatic Epithelia in Three-dimensional Collagen Gels
Dorsal pancreatic epithelia were grown in three-dimensional collagen gels. The collagen gels were prepared according to Montesano et al. and as previously described (24, 26). 200 μl of collagen were dispensed into 15-mm wells of four-well plates and allowed to gel at 37°C. The pancreatic epithelia were placed on the surface of the collagen gel and covered with 300 μl of collagen solution. After the second layer had gelled, 500 μl of complete culture medium was added. The complete medium consisted of RPMI 1640 supplemented with 10% heat-inactivated FCS, 2 mM glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin. In some experiments, recombinant human TGF-β1 (R&D Systems Europe, Oxon, United Kingdom) was added to the culture medium at a final concentration of 1 ng/ml. In other experiments, a pan-specific TGF-β neutralizing antibody (AB-100-NA; R&D Systems) was used at a final concentration of 2 μg/ml. Rabbit preimmune sera was used as a control for the experiences using the pan TGF-β neutralizing antibody. The antibody was added every day either from the first or third day of the culture. For perturbing experiments, a stock solution (10 mM) of the metalloproteinases inhibitor BB-3103 (British Biotechnology Pharmaceuticals, Ltd., Oxford, UK) was prepared in DMSO. Working dilution of the inhibitor and comparable DMSO controls were made in culture medium. Cultures were maintained at 37°C in a humidified atmosphere of 95% air and 5% CO2. Medium was replaced every 48 h. At the indicated times, the pancreatic epithelia were photographed and fixed for immunohistology as described below.
Immunohistology
The pancreatic epithelia were fixed at 4°C in 4% paraformaldehyde in PBS for 2 h, briefly rinsed with PBS, cryoprotected overnight at 4°C in 30% sucrose, and frozen. Consecutive sections (6 μm thick) were cut and collected on gelatinized glass slides. For immunostaining, the sections were first incubated for 30 min in PBS containing 3% BSA. Subsequently, the sections were incubated for 2 h at room temperature (or overnight at 4°C) with the primary antibodies. After washing in PBST (PBS containing 0.5% Tween-20), the sections were incubated with the appropriate fluorescent secondary antibodies. Finally, the sections were extensively washed in PBST and mounted with a fluorescence protecting medium (Vectashield, Vector Laboratories, Inc., Burlingame, CA). The sections were examined and photographed with a Leitz DMRD microscope. Colocalization was determined by double-labeling immunofluorescence. For this purpose, antibodies prepared in two different species were used and revealed using anti-species antibodies labeled with two different fluorochromes. Single-labeled sections incubated with mismatched secondary antibodies showed no immunostaining, confirming the specificity of the secondary antisera.
The antisera used in this study were used at the following dilutions: guinea pig anti-porcine insulin (1:500; DAKO A/S, Glostrup, Denmark); mouse monoclonal anti-porcine glucagon (1:2,000; Sigma Chemical Co., St. Louis, MO); rabbit anti-human amylase (1:2,000; Sigma Chemical Co.); mouse monoclonal anti-human insulin (1:2,000; Sigma Chemical Co.); and sheep anti-human MMP-2 (1:100; reference 15).
The fluorescent secondary antibodies were fluorescein anti-guinea pig antibodies (1:500; DAKO A/S) and the following secondary antibodies from Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA): fluorescein anti-rabbit antibodies (1:200); fluorescein anti-mouse antibodies (1:200); Texas red anti-rabbit antibodies (1:500); and Texas red anti-mouse antibodies (1:200).
Quantitative Analysis
To determine the number of insulin-positive cells per pancreatic epithelium, serial 6-μm sections were cut and collected on multiwell glass slides. One out of two consecutive sections (that is, sections separated by 12 μm) was analyzed by immunocytochemistry to avoid counting the same cell twice. A mean of 50 ± 5 sections was analyzed per rudiment. Only the immunoreactive cells for which a nucleus was clearly visible were counted. The results of each experimental point were obtained by quantifying the absolute number of each pancreatic cell type in at least five pancreatic rudiments. Data are presented as mean ± SEM. Statistical analysis was performed using a Student's t test.
Reverse Transcriptase (RT) PCR Analyses
Total RNA was extracted from pancreatic rudiments (6) and reverse-transcribed as previously described (3). In brief, first-strand cDNA was prepared from DNase-treated RNA using random hexamer primers: 200 μM each of dNTP, and 100 U of reverse transcriptase (SuperScript; Life Technologies, Inc., Grand Island, NY). The reaction was also carried out in the absence of reverse transcriptase to ensure that the amplified material derive from RNA. The oligonucleotides used for amplification were as follows:
MMP-2 (sense) 5′-TTACACACCTGACCTGGACCCT-3′;
MMP-2 (antisense) 5′-CGTTCCCATACTTACTCGGACC-3′;
MMP-9 (sense) 5′-GAGGGACGCTCCTATTTGTCCT-3′;
MMP-9 (antisense) 5′-GGGAAGACGCACATCTCTCCT-3′;
MT-MMP (sense) 5′-GGAATGACATCTTCTTGGTGGC-3′;
MT-MMP (antisense) 5′-TAGGTGGGGTTTCTGGGCTTAT-3′;
TIMP-1 (sense) 5′-GATATGTCCACAAGTCCCAGAACC-3′;
TIMP-1 (antisense) 5′-GCAGGCAAAGTGATCGCTCT-3′;
TIMP-2 (sense) 5′-CGGAAGGAGATGGCAAGATG-3′;
TIMP-2 (antisense) 5′-GCGCAAGAACCATCACTTCTCT-3′;
TIMP-3 (sense) 5′-AAAGAAGCTGGTGAAGGAAGGG-3′;
TIMP-3 (antisense) 5′-CTGGGACAGTGTGAGGTGGTC-3′;
TGF-β1 (sense) 5′-GACTCTCCACCTGCAAGACCAT-3′;
TGF-β1 (antisense) 5′-TCCCGAATGTCTGACGTATTGA-3′;
TGF-β2 (sense) 5′-CGCCCACTTTCTACAGACCCTA-3′;
TGF-β2 (antisense) 5′-GAGGTGCCATCAATACCTGCA-3′;
TGF-β3 (sense) 5′-GAAGAAGAGGGTGGAAGCCATT-3′;
TGF-β3 (antisense) 5′-GGTTCCATTTTTCTCCACTGAGG-3′;
TGF-βR1 (sense) 5′-TTTCAGAGGGCACCACCTTAAA-3′;
TGF-βR1 (antisense) 5′-CCATGCTCATGATAATCCGACA-3′;
TGF-βR2 (sense) 5′-AGAAGTGAAAGATTATGAGCCCCC-3′;
TGF-βR2 (antisense) 5′-GTGTTCAGGGAGCCGTCTTCT-3′;
Cyclophilin (sense) 5′-ATGGTCAACCCCACCGTGTT-3′;
Cyclophilin (antisense) 5′-CGTGTGAAGTCACCACCCT-3′.
30 cycles of amplification were performed. Amplification parameters included a 1-min denaturation step at 94°C, a 1-min annealing step at 57°C, and a 30-s extension step at 72°C. The products of amplification were separated on a 2% agarose gel, and were photographed.
Zymography
Tissue-associated and secreted metalloproteinases were detected and characterized by zymography (14). This technique was applied to pools of 10 solubilized epithelia, and to concentrated media conditioned from pools of 10 epithelia cultured for 24 h. The pancreatic rudiments were directly lysed in SDS-PAGE sample buffer (50 mM Tris, 1% SDS, 5% glycerol, 0.002% bromophenol blue). Concentrated conditioned media was diluted in the same buffer. Then, the solubilized rudiments and concentrated media were loaded on 8% SDS-polyacrylamide gels copolymerized with 1 mg/ml of gelatin (Sigma Chemical Co.). Electrophoresis was performed under nonreducing conditions at 20 mA for 2 h at room temperature. Gels were washed twice for 30 min in 2.5% Triton X-100 to remove SDS, incubated in substrate buffer (50 mM Tris-HCl, 5 mM CaCl2, 1 mM ZnCl2, pH 7.5) for 24 h at 37°C, stained in 0.5% Coomassie blue (prepared in 30% ethanol, 10% acid acetic, 1% formaldehyde) for 30 min at room temperature, and destained three times for 15 min in the same buffer devoid of Coomassie blue. The presence of metalloproteinase was indicated by an unstained proteolytic zone of the substrate.
Results
Metalloproteinase Expression and Activity In Vivo during Pancreatic Development
Extracts of rat pancreas were examined by gelatin zymography in order to identify metalloproteinase activity in the pancreas at specific stages of development. A metalloproteinase of 72 kD, the precursor form of MMP-2 (15), was detected at all the pancreatic stages analyzed. The levels of this enzyme did not appear to change significantly over time (Fig. 1 A). In contrast, a proteinase of ∼66 kD, which represents the activated form of the MMP-2, was detected in extracts from E17 to E19 pancreases. By E19, the 66-kD form of the enzyme represented roughly 50% of the total enzyme present in the pancreas (Fig. 1 A). In the newborn and adult pancreas, the enzyme was found only in its inactive form (72 kD). No activity associated with the 92-kD forms of MMP-9 was detected (Fig. 1 A). As shown in Fig. 1 B, the MMP activity present at E19 was inhibited by the synthetic compound BB-3103, a hydroxamic acid–based inhibitor of proteases (2, 23). Inhibition of the gelatinase zymograms was effective in the range of 50–1,000 μM (Fig. 1 B).
Figure 1.

Metalloproteinases activity in the developing pancreas: (A) Equal amounts of protein extracts from each of the stages indicated were analyzed by zymography. A proteinase of 70–72 kD, representing the proform of the enzyme MMP-2, was detected in samples of all the stages of pancreatic development. The 62–64 kD activated form of the enzyme is essentially detectable between E17 and E19. (B) Protein extracts from E19 rat pancreas were run on a 8% SDS-polyacrylamide gel copolymerized with 1 mg/ml of gelatin. The gel was next divided into sections for subsequent overnight digestion at 37°C in incubation buffer (50 mM Tris, 10 mM CaCl2, pH 7.6) containing (a) no additive; (b) 10 μM BB-3103; (c) 50 μM BB-3103; (d) 1 mM compound BB-3103; or (e) 0.1 μl/ml DMSO diluent.
RT-PCR was used in a qualitative manner to determine expression in the developing pancreas of the transcripts encoding the different components of the metalloproteinase system. MMP-2, MT1-MMP, TIMP-1, and TIMP-2 transcripts in the pancreatic rudiments were found to remain expressed throughout the entire pancreatic development as well as in the adult pancreas (Fig. 2 A). This observation correlates well with the levels of expression of the 72-kD form of the enzyme detected by zymography. Similar results were obtained for expression of the transcripts encoding MT1-MMP, the membrane activator of MMP-2, and the metalloproteinase inhibitors TIMP-1 and TIMP-2. To determine the cell types expressing MMP-2, sections of fetal pancreas in the late stages of development (E20) were labeled with sheep anti-human MMP-2 antibodies, which recognize mice and rat MMP-2 (21). At that stage, MMP-2 immunoreactivity was detected in insulin-positive cells (Fig. 2 B). Indeed, a perfect colocalization was found between insulin-immunoreactivity and MMP- 2-immunoreactivity.
Figure 2.
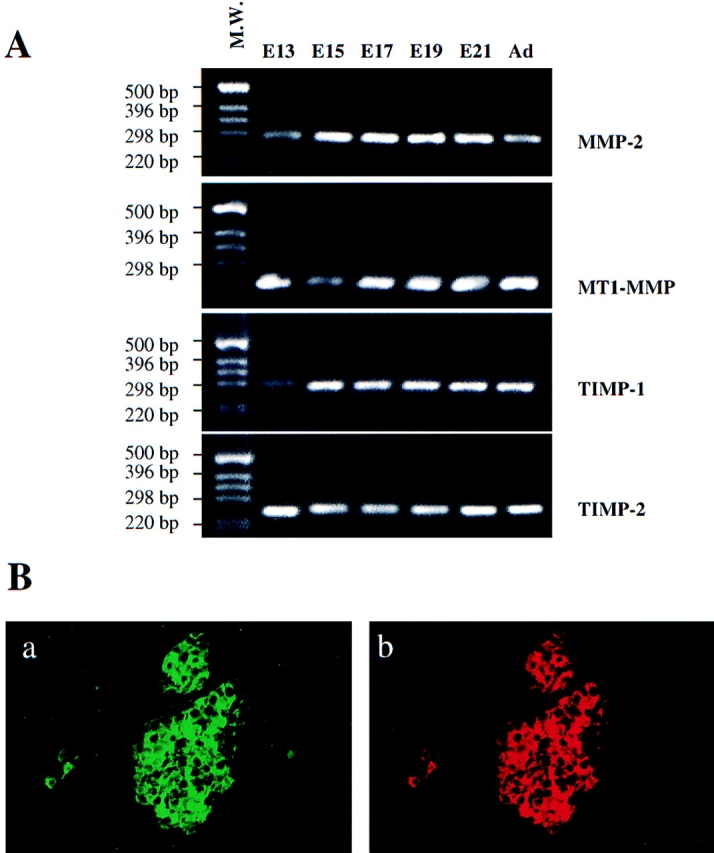
Expression of matrix metalloproteinase components in the pancreas in development. (A) RT-PCR analysis of MMP-2, MT1-MMP, TIMP-1, and TIMP-2 expression. First-strand cDNA was synthesized and used as template in PCR reactions. 30 cycles of amplification were performed for each reaction. (B) Sections of newborn rat pancreas were costained with (a) anti-insulin and (b) anti-MMP-2 antibodies.
Development of E12.5 Pancreatic Epithelia In Vitro
When E12.5 rat pancreatic epithelia were cultured into a three-dimensional collagen gel, they rapidly acquired a spherical shape. This spherical shape remained unchanged during the first 4 d of culture (Fig. 3 A). During these first 4 d, differentiation into endocrine cells occurred. Indeed, while very few endocrine cells were present before culture (24), a considerable number of glucagon- and insulin-positive cells were detected by immunohistochemistry after 4 d in culture. At that stage of culture, endocrine cells were found into the central epithelial mass of the pancreatic rudiment, and did not associate to form typical islet structures (Fig. 3 B).
Figure 3.

Morphological and immunohistological analysis of pancreatic rudiments developed in vitro. Pancreatic rudiments devoid of their surrounding mesenchyme were grown into tridimensional collagen gels. Pancreatic rudiments were photographed (A and C) and analyzed by immunohistochemistry (B and D) for insulin (green) or glucagon (red) expression after 4 (A and B) and 7 d (C and D) of culture.
On the other hand, after 7 d in vitro as previously described (24), a variable number of translucent structures have emerged from the central epithelial sphere (Fig. 3 C). The immunohistological analysis revealed that these translucent structures were constituted exclusively by endocrine cells with glucagon-positive cells present at the periphery of large clusters of insulin-positive cells (Fig. 3 D). Thus, in the present in vitro system, pancreatic epithelial cells first differentiate into endocrine cells. Once differentiated, these endocrine cells migrate away from the central epithelial mass and associate into islets of Langerhans. The distribution of the specific endocrine cells into the round compact structures resembles the islets found in vivo.
Implication of MMP Activity in Islet Morphogenesis In Vitro
As we have described that MMP-2 is activated in vivo in the developing pancreas between E17 and E19, a period where islets develop, we asked whether MMP-2 activity was implicated in islet formation found in the in vitro model described above. We first tested whether the different components of the metalloproteinase system were expressed in vitro. The RT-PCR analysis showed that transcripts encoding the MMP-2 metalloproteinase, its membrane-associated receptor MT1-MMP, and the inhibitor/activators TIMP-1, TIMP-2, and TIMP-3, were expressed in the epithelia developed in culture for 4 d (Fig. 4 A). To define the cell types expressing MMP-2 in vitro, immunohistological analysis was performed using a sheep anti-human MMP-2 antibody. As shown in Fig. 4 B, the antibody reacted positively with the majority of the insulin-positive cells present in pancreatic epithelia cultured for 4 d. The remaining tissue stained negatively.
Figure 4.


Expression of the matrix metalloproteinases components in pancreatic rudiments in culture. (A) RT-PCR analysis of MMP-2, MT1-MMP, and TIMPs 1–3 expression in pancreatic rudiments cultured for 4 d. First-strand cDNA was synthesized and used as template in PCR reactions. 30 cycles of amplification where performed for each reaction. (B) Immunohistochemical analysis of insulin and MMP-2 expression in pancreatic rudiments. Sections were costained with (a) anti-insulin and (b) anti-MMP-2 antibodies.
To further examine the role of MMPs in islet morphogenesis, perturbing experiments were performed. For that purpose, the effect of the protease inhibitor BB-3103, which as shown above inhibited MMP-2 activity present in E19 pancreases, was tested in tissue culture. As shown in Fig. 5 when pancreatic epithelia were cultured in the presence of 1 or 10 μM BB-3103, islet formation was impaired. Indeed, endocrine structures fail to bud from the central mass in the presence of the inhibitor (compare A to B and C). On the other hand, cell differentiation was not impaired in the presence of the inhibitor. Indeed, immunohistological analysis showed that after 7 d in culture, differentiation into insulin- or glucagon-expressing cells did occur in the presence of the compound BB-3103 (Fig. 5, D–F). Differentiation into amylase-expressing cells did also occur properly in the presence of inhibitor (Fig. 5, G–I). Moreover, quantification of the absolute number of endocrine and exocrine cells in the explants treated with 1–10 μM indicated that the absolute number of differentiated cells was not significantly different from the one found in control rudiments (Table I, A).
Figure 5.
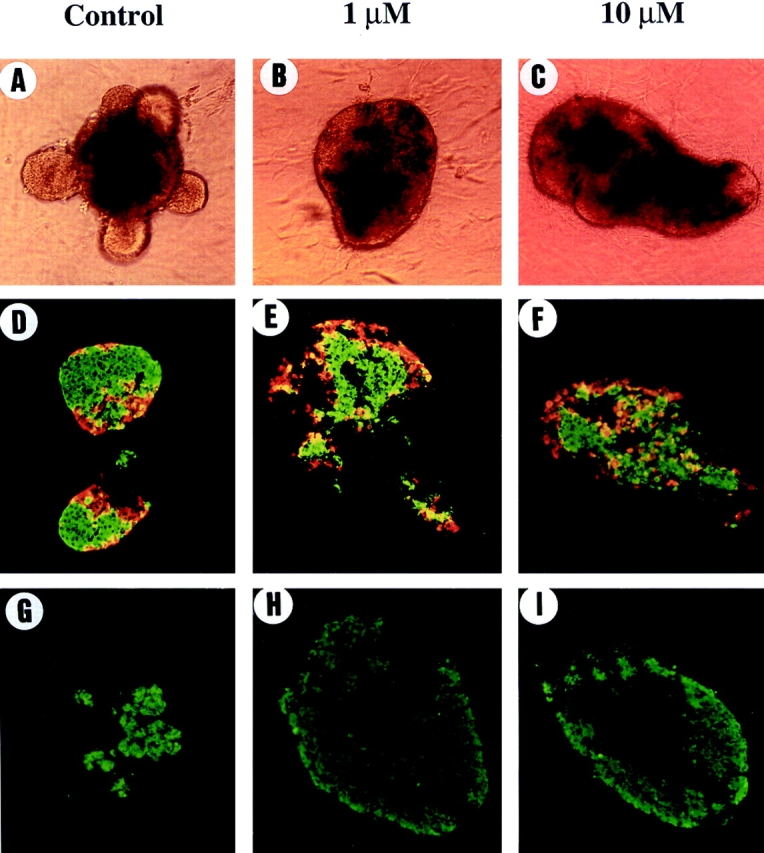
Compound BB-3130 inhibits islet morphogenesis without affecting cell differentiation. The rudiments were grown for 7 d in the absence (control) or in the presence of BB-3103 (1 μM or 10 μM). Adding 1 μM or 10 μM of compound BB-3103 inhibits islet morphogenesis (compare A to B and C). On the other hand, immunohistochemical analysis revealed that differentiation into insulin-positive cells in green or into glucagon-positive cells in red occured both in the absence (D) or in the presence (E and F) of inhibitor. The differentiation into amylase-positive cells did also occur in the absence (G) or in the presence (H and I) of BB-3103.
Table I.
Quantitative Analysis of Pancreatic Development In Vitro
| Experience | Average cell number/pancreatic rudiment |
n
|
||||||
|---|---|---|---|---|---|---|---|---|
| Endocrine | Exocrine | |||||||
| A | ||||||||
| Control | 1,900 ± 100 | NS | 900 ± 80 | NS | 5 | |||
| BB-3103 | 1,800 ± 90 | 836 ± 100 | 8 | |||||
| B | ||||||||
| Control | 1,900 ± 100 | NS | 940 ± 110 | NS | 10 | |||
| TGF-β1 | 1,700 ± 80 | 1,150 ± 100 | 10 | |||||
| C | ||||||||
| Control | 1,800 ± 100 | NS | 1,000 ± 120 | NS | 11 | |||
| Anti-TGF-β (d0–d7) | 1,500 ± 200 | 1,400 ± 100 | 16 | |||||
| D | ||||||||
| Control | 1,600 ± 200 | NS | 1,100 ± 250 | NS | 5 | |||
| Anti-TGF-β (d3–d7) | 1,700 ± 100 | 2,000 ± 300 | 5 | |||||
Quantification of the number of endocrine and exocrine cells developed in the pancreatic rudiments grown in vitro. The absolute number of endocrine and exocrine cells developed in the pancreatic rudiments was determined by counting respectively the number of endocrine cells (insulin- and glucagon-immunoreactive cells) and the number of exocrine cells (amylase-immunoreactive cells). The data represent the mean ± SEM. n = number of pancreatic rudiments analyzed per experience.
The effect of BB-3103 was fully reversible. Indeed, 2 d after removing the compound BB-3103, endocrine structures started to bud from the core of the explants, and 3 d later the explants showed the same appearance than the control explants (data not shown).
As controls, pancreatic explants were cultured in the presence of zinc metalloendopeptidases (thiorphan, 0.1 mM), serine proteases (aprotinin, 100 μg/ml), or cystein proteinases (leupeptin, 10 mM). In these conditions, the explants developed large endocrine buds identical to those found in controls (data not shown).
TGF-β1 Positively Controls Both Islet Morphogenesis and MMP-2 Activity
TGF-β has been shown to play an important role in endocrine pancreatic development (37). Since it has also been shown that TGF-β controls MMP activities in different models (22, 29, 31, 32, 34, 35, 48), we analyzed in vitro the effect of TGF-β1 on both islet morphogenesis and MMP activity. After 4 d in culture in the presence of TGF-β1, translucent buds have already emerged, which was not the case after 4 d in the absence of exogenous TGF-β1 (Fig. 6, compare A and B). In the TGF-β1–treated rudiments, many of the endocrine cells showed a spindled shape characteristic of cells that are in migration (Fig. 6, compare C and D). Islet morphogenesis thus occurs faster in the presence of TGF-β1 than in the absence of TGF-β1. During the next 3 d the buds continue to develop (Fig. 6, E and H). Note that in the presence of exogenous TGF-β1 after 7 d in culture, the segregation between beta and nonbeta cells was perturbed when compared with controls. Indeed, insulin-positive cells were not surrounded by glucagon-positive cells (Fig. 6 H). Finally, when the total number of endocrine cells developed in the absence or in the presence of TGF-β1 were counted after 4 or 7 d of culture, no significant difference was found between rudiments grown in the presence or absence of TGF-β1, suggesting that exogenus TGF-β1 treatment did not perturb cell differentiation (Table I, B).
Figure 6.
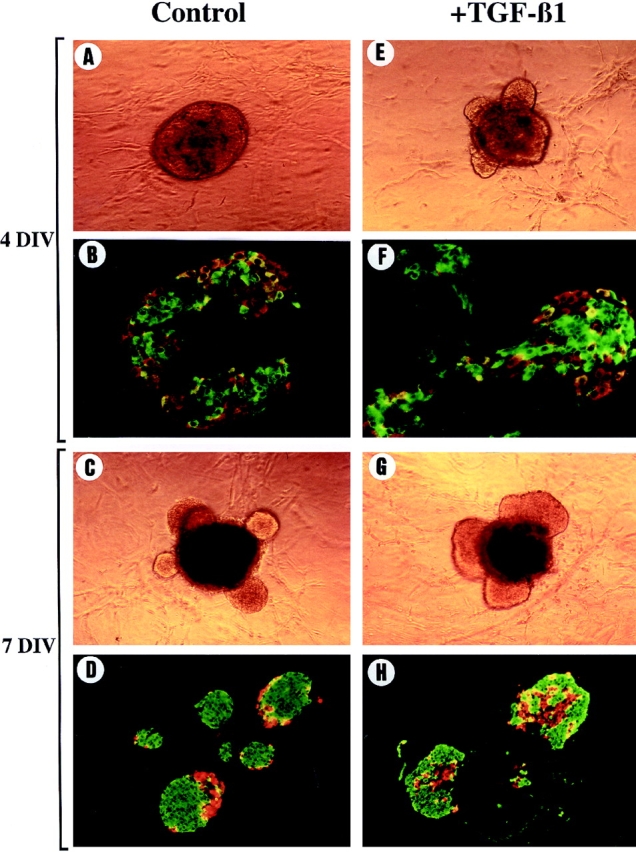
Morphological and immunohistological analysis of pancreatic rudiments developed in vitro in the presence of exogenous TGF-β1. Pancreatic rudiments devoid of their surrounding mesenchyme were grown into tridimensional collagen gels in the absence (A, C, E, and G) or in the presence (B, D, F, and H) of TGF-β1. Pancreatic rudiments were photographed and analyzed by immunohistochemistry for insulin (green) or glucagon (red) expression after 4 (A–D) and 7 d (E–H) of culture. Note that after 4 d in culture in the presence of TGF-β1, translucent buds were already emerging (B), and many endocrine cells showed a spindled shape characteristic of cells that are in migration (D), which was not the case after 4 d in the absence of exogenous TGF-β1.
The next step was to compare the activity of metalloproteinases in pancreatic rudiments grown for 4 or 7 d in the presence or absence of exogenous TGF-β1. Extracts of pancreatic rudiments were analyzed by zymography. The zymograms showed that a 72-kD band corresponding to the inactive form of MMP-2 was present after both 4 and 7 d of culture in rudiments grown in the presence or absence of TGF-β (Fig. 7). However, while after 4 d in culture in the absence of TGF-β1, no band corresponding to the activated form of MMP-2 was detected (Fig. 7, lane 1), such a band was detected in rudiments developed for 4 d in the presence of TGF-β1 (Fig. 7, lane 2). Finally, after 7 d of culture, a band corresponding to the activated form of MMP-2 was found in rudiments grown in the presence or absence of exogenous TGF-β1. Similar results were obtained when the concentrated conditioned media of pancreatic rudiments were analyzed by zymography (data not shown). Thus, in the presence of TGF-β1, both islet morphogenesis and MMP-2 activation are accelerated.
Figure 7.

Analysis of gelatinase expression and activity in pancreatic rudiments cultured in the presence of exogenous TGF-β1. Pancreatic rudiments grown for 4 or 7 d in the presence or absence of TGF-β were analyzed for MMP-2 activity by gel zymography. Lanes 1 and 2: pancreatic rudiment grown for 4 d without (lane 1) or with (lane 2) 1 ng/ml TGF-β1. Lanes 3 and 4: pancreatic rudiment grown for 7 d without (lane 3) or with (lane 4) 1 ng/ml TGF-β1.
TGF-β Is Necessary for Islet Morphogenesis
Our results described above demonstrate that exogenous TGF-β accelerates islet morphogenesis. However, islet morphogenesis did also occur in the absence of exogenous TGF-β after 7 d of culture. This formation of islets found after 7 d of culture in the absence of exogenous TGF-β1 was also paralleled by an induction of MMP-2 activity (Fig. 7, lanes 1 and 3). This result suggests that if TGF-β is necessary for islet morphogenesis, (a) pancreatic epithelia in culture should produce TGF-β and express TGF-β receptors; and (b) islet morphogenesis should not occur when TGF-β activity is blocked.
RT-PCR analysis was used to define whether TGF-β transcripts as well as transcripts encoding the receptors of TGF-β were expressed in the in vitro–cultured rudiments. As shown in Fig. 8, predicted-size DNA fragments corresponding to TGF-β1, TGF-β2, and TGF-β3 were amplified from pancreatic rudiments cultured for 4 d. More cycles of amplification were necessary to amplify TGF-β1 than TGF-β2 or TGF-β3, suggesting that the level of expression of TGF-β1 was lower than that of TGF-β2 and TGF-β3. TGF-β receptors (TGF-βRI and TGF-βRII) could also be amplified from pancreatic epithelia cultured for 4 d.
Figure 8.

Expression of TGF-β and its receptors in pancreatic rudiments in culture. cDNAs were prepared from pancreatic rudiments devoid of mesenchyme and cultured for 4 d in collagen gels. Then they were used as template in PCR reactions with specific primers for TGF-β1, TGF-β2, TGF-β3, TGF-βRI, and TGF-βRII. (A) 30 cycles of amplification. (B) 35 cycles of amplification.
To further investigate the implication of endogenous TGF-β in islet morphogenesis, pancreatic epithelia were cultured in the presence of a pan-specific TGF-β neutralizing antibody. The antibody was added every day starting either from the first day or from the third day of the culture. Both MMP-2 activation and islet morphogenesis were followed. When the anti-TGF-β is added either from day 0 or from day 3 of the culture and the zymography performed at day 7, MMP-2 is found in its inactive form, while in the absence of antibody, MMP-2 is found activated (Fig. 9). In the absence of neutralizing antibody, islet morphogenesis occurred, the endocrine cells being found outside of the central core of amylase-positive cells (Fig. 10, A and B). When the rudiments were cultured in the presence of the anti-TGF-β neutralizing antibody from day 3 to day 7, endocrine structures fail to bud from the central mass, and remain surrounded by the amylase-positive cells (Fig. 10, C and D) in a way identical to that found when the rudiments were grown in the presence of the metalloproteinase inhibitor BB-3101 (compare Fig. 5, E and H to Fig. 10, C and D). When the rudiments were cultured in the presence of the anti-TGF-β neutralizing antibody from day 0 to day 7, the endocrine cells did not migrate away from the central epithelial sphere either. They were found dispersed in the central core of the rudiments (Fig. 10, E and F), suggesting that during the first days of culture, TGF-β plays a role in endocrine cell adhesion. It is also clear that when the antibody was added from day 0 to day 7, cells negative for insulin, glucagon, and amylase were frequently found in the rudiment, a fact that was found neither in control rudiments nor in rudiments treated with the antibody from day 3 to day 7. Finally, the quantitative analysis showed that the absolute number of either endocrine or exocrine cells was not affected by the panTGF-β neutralizing antibody (Table I, C and D).
Figure 9.

Analysis of gelatinase A expression and activity in pancreatic rudiments grown in the presence of a panTGF-β neutralizing antibody. The pancreatic rudiments were cultured in the presence of a panTGF-β neutralizing antibody, and after a 7-d culture period, the rudiments were recovered and their gelatinase activity was analyzed by gel zymography. The antibody was added either from the third (A) or from the first (B) day of culture. (−) rudiments cultured in the presence of nonimmune serum; (+) rudiments cultured in the presence of the TGF-β neutralizing antibody.
Figure 10.

Analysis of pancreatic rudiments grown in the presence of a panTGF-β neutralizing antibody. To define whether TGF-β is necessary for islet formation, pancreatic rudiments were cultured in the presence of a pan-specific TGF-β neutralizing antibody. The antibody was added either from the first (d0) or the third (d3) day of the culture. The rudiments were analyzed by immunohistochemistry for insulin, glucagon, and amylase expression. In the control rudiments, the insulin- (green) and glucagon- (red) expressing cells are arranged into islet-like structure (A), budding from the core of the rudiment that is mainly composed of amylase-expressing cells (B). When the rudiments are cultured in the presence of the panTGF-β neutralizing antibody from the third day of culture (d3–d7), the endocrine cells differentiate, but do not bud (C), and form small clusters interspersed between the acinar cells (D). When the rudiments are cultured in the presence of the panTGF-β neutralizing antibody from the first day of culture (d0–d7), a more drastic effect is observed. The endocrine cells appear dispersed (E) and surrounded by acinar cells (F).
Discussion
This work presents novel observations with respect to the morphogenesis of the islets of Langerhans of the pancreas. First, we have demonstrated that induction of MMP-2 activity is necessary for proper morphogenesis of the islets of Langerhans. Second, we have shown that TGF-β is a key activator of MMP-2 in the islets, and that inhibition of TGF-β activity represses islet morphogenesis.
During development, the pancreas forms as an epithelial evagination of the foregut endoderm into the surrounding mesenchyme (39). Putative stem cells present in the epithelium will next differentiate into exocrine or endocrine cells. The differentiated endocrine cells will migrate away from the primitive epithelium into the surrounding mesenchyme and aggregate into microorgans: the islets of Langerhans. Mouse aggregation chimera experiments have confirmed this model of development, and have shown that islets are not derived from a few cells that proliferate, but through aggregation of endocrine cells interspersed in the interstitial tissue (9). Thus, cell migration through the ECM represents an essential step for proper morphogenesis of the endocrine islets.
In different models, cell migration requires ECM degradation. Among the enzymes implicated in ECM degradation, the collagenase metalloproteases MMP-2 and MMP-9 have focused our attention, since these proteases have been implicated in morphogenetic events associated with organogenesis. For example, MMP-2 and -9 are thought to contribute to morphogenesis of the tongue and Meckel cartilage during mandibule arch development (5). MMP-9 has also been shown to be necessary for branching morphogenesis of the ureter bud (21). MMP-2 also focused our attention because it is implicated not only in the degradation of the ECM, but also in cell migration by providing a signal for cell migration initiation (10). MMP activity is known to depend on a complex balance between MMPs, their cell surface activators (MT-MMPs), and their inhibitors (TIMPs; 45). Our study clearly establishes that MMP-2, MT1-MMP, and various TIMPs are expressed through the entire pancreatic development. However, zymographic studies show that the activated form of MMP-2 is essentially detectable between E17 and E19, a period when endocrine cells leave the primitive epithelium, migrate, and associate into the well-defined architecture of adult islets (13, 25, 47). Between E17 and E19, MMP-2 could thus be implicated in degradation of the ECM, which is at that stage mainly composed of collagen types -I, -III, and -IV (16). This degradation step would be necessary to allow endocrine cells to migrate into the ECM.
To investigate further the eventual implication of MMPs in islet morphogenesis, we have used a recently described in vitro model of pancreatic development (24). Pancreatic epithelium prepared from E12.5 rat pancreatic rudiments was cultured in a tridimensional collagen. In these conditions, the pancreatic epithelium differentiates essentially into endocrine cells (24). As shown in the present study, differentiation occurs in the central epithelial mass. Once differentiated, the cells migrate into the collagen gel and associate into microorgans, which resemble islets of Langerhans found in vivo after birth (28). Thus, this in vitro model mimics the in vivo process of islet morphogenesis, and can thus be used to understand how islet morphogenesis occurs.
Our perturbing experiments performed using the compound BB-3101, a hydroxamic acid–based inhibitor of proteases (2, 23), demonstrate that when MMP-2 activity is blocked, endocrine cell differentiation occurs normally, while islet morphogenesis is abolished. This result demonstrates that morphogenesis is not necessary for endocrine cell differentiation. This last result is not unexpected since it has been shown that pancreatic cytodifferentiation can occur in the absence of morphogenesis. Indeed, when embryonic pancreatic epithelium is cultured in conditions that do not favor pancreatic morphogenesis, cytodifferentiation takes place (41). Moreover, during pancreatic development, endocrine cell differentiation is found before pancreatic morphogenesis (12).
Our results also demonstrate that TGF-β is a strong activator of MMP-2 activity. In other models, TGF-β1 usually increases matrix component production, decreases proteinase activities, and increases proteinase inhibitors synthesis (31, 32, 48). But there are also some examples in which TGF-β displays an stimulating effect on MMP-2 (22, 29) or MMP-9 activities (34, 35). When pancreatic rudiments were cultured in the presence of TGF-β1, the timing of MMP-2 activation and islet morphogenesis were accelerated when compared with controls, suggesting that TGF-β1 acts as an activator of ECM degradation. At that point of the discussion, it is of interest to consider the status of the ECM in transgenic animals overexpressing TGF-β in beta cells. Indeed, in these transgenic animals, increased deposition of ECM was observed (20, 38). These results seem to be in partial contradiction with the data presented here. However, the results obtained in transgenic animals are difficult to compare with the data presented here, because (a) abnormal deposition of ECM could in fact be due to an increase in ECM synthesis and not to a decrease in ECM degradation; and (b) in the transgenic model, progressive accumulation of ECM was observed only after birth, no embryonic/fetal phenotype being found.
In the present study, TGF-β was found to be an activator of islet morphogenesis. Indeed, adding exogenous TGF-β accelerates islet formation. However, islet morphogenesis did also occur in the absence of exogenously added TGF-β, suggesting that endogenously produced TGF-β could be implicated in islet morphogenesis. We demonstrate that: (a) TGF-β1, -β2, and -β3 are produced in the present experimental system; and (b) islet morphogenesis is abolished when a specific panTGF-β neutralizing antibody is added to the culture medium. The demonstration of TGF-β production by pancreatic cells confirms previous data demonstrating that in mice, TGF-β1 mRNA is present in pancreatic buds throughout development (38), and that TGF-β1, -β2, and -β3 are synthesized in the endocrine and exocrine human pancreas (46). However, the fact that a panTGF-β neutralizing antibody abolished islet morphogenesis was not totally expected. Indeed, it does demonstrate that TGF-β is necessary for islet morphogenesis. To our knowledge, no such drastic effect of TGF-β on morphogenesis induction has been described up to now. Islet morphogenesis does thus represent a key model to understand how TGF-β controls morphogenesis.
While a panTGF-β neutralizing antibody perturbed islet morphogenesis, endocrine cell differentiation was not affected. The lack of effect of TGF-β on endocrine cell differentiation is in concordance with data obtained in transgenic mice that overexpress TGF-β1 in beta cells and have a normal number of endocrine cells (20, 38). It seems, however, in contradiction with other studies showing that in vitro TGF-β1 influences the relative proportion of the exocrine and endocrine pancreas (37). In this latter study, the experimental system was quite different from the one used here. Indeed, pancreatic embryonic epithelia were grown in the presence of their surrounding mesenchyme, while we did use mesenchyme-depleted epithelium. However, in the work of Sanvito et al. (37), TGF-β treatment modified the shape and arrangement of stromal cells, and the authors postulated that one possibility would be that some of the effects of TGF-β1 on epithelial cells could in fact be indirect and secondary to changes in the extracellular matrix through effects on stromal cells (37). Such a hypothesis could be right and could represent an explanation of the difference in endocrine cell numbers between our work and the work of Sanvito et al. (37).
In addition to its role in activating MMP-2 and inducing islet morphogenesis, we demonstrate that TGF-β has some additional effects. First, when an excess of TGF-β is added, while islet morphogenesis is accelerated, the final three-dimensional organization of the different endocrine cell types is perturbed. In this case, insulin-positive cells are not surrounded by glucagon-positive cells as in normal islets (28). Second, when the rudiments are grown in the presence of a neutralizing TGF-β antibody, the treatment starting from the beginning of the culture (i.e., before cell differentiation had occurred) islet morphogenesis and endocrine cell adhesion are perturbed. Some effects on cell differentiation could also be seen in those conditions. No such effects are seen when the antibody is added while cell differentiation has already started. The effect of TGF on cell adhesion could be mediated by regulating N-CAM with TGF-β. Indeed, N-CAM has been proposed as being implicated in pancreatic endocrine cell adhesion (7), and TGFs control N-CAM expression in different experimental systems (33, 42). The effect of TGF-β on cell differentiation observed when the neutralizing antibody is added at the beginning of the culture resemble s the one described when a dominant-negative TGF-β type II receptor is expressed in transgenic mice in acinar cells (4). Indeed, we found cells staining negative for insulin, glucagon, and amylase. These cells could represent acinar cells that dedifferentiate into cells with a ductal phenotype as previously proposed (4).
Recently, TGF-β1, TGF-β2, and TGF-β3 knockout mice have been generated (17, 36). To our knowledge, no overt pancreatic phenotype has been described in those animals. This fact can be explained either by the fact that the pancreas was not analyzed in great detail, or by the fact that isoform redundancy among the three TGF-βs isoforms masks the effect. In fact, the latter hypothesis is plausible because all three TGF-β family members are found to be expressed in the pancreas (46, the present work).
Acknowledgments
We wish to acknowledge Dr. Gillian Murphy for the gift of anti–MMP-2 antibodies, and British Biotechnology Pharmaceuticals Ltd. for providing us with the metalloproteinases inhibitor BB-3103. We thank Drs. A. Wandevalle and B. Nguyen for their helpful advice and O. Dubois for technical assistance.
This work was supported by the Juvenile Diabetes Foundation International No. 198207 (to R. Scharfmann) and by the European Community.
Abbreviations used in this paper
- ECM
extracellular matrix
- MMP
matrix metalloproteinase
- RT
reverse transcriptase
- TIMPs
tissue inhibitors of metalloproteinases
Footnotes
Address all correspondence to Francisco Miralles, INSERM U457, Hospital R. Debré, 48 Boulevard Serurier, 75019 Paris, France. Tel.: 33 1 40 03 19 86. Fax: 33 1 40 03 19 03. E-mail: miralles@infobiogen.fr
References
- 1.Ahlgren U, Pfaff S, Jessel T, Edlund T, Edlund H. Independent requirement for ISL1 in formation of pancreatic mesenchyme and islet cells. Nature. 1997;385:257–260. doi: 10.1038/385257a0. [DOI] [PubMed] [Google Scholar]
- 2.Ancuta P, Fahma H, Pons J, LeBlay K, Chaby R. Involvement of the membrane form of tumour necrosis factor-alpha in lipopolysaccharide-induced priming of mouse peritoneal macrophages for enhanced nitric oxide response to lipopolysaccaride. Immunology. 1997;92:259–266. doi: 10.1046/j.1365-2567.1997.00338.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Atouf F, Czernichow P, Scharfmann R. Expression of neuronal traits in pancreatic beta cells: implication of NRSF/REST, a neuron-restrictive silencer. J Biol Chem. 1997;272:1929–1934. doi: 10.1074/jbc.272.3.1929. [DOI] [PubMed] [Google Scholar]
- 4.Bottinger E, Jakubczak J, Roberts I, Mumy M, Hemmati P, Bagnall K, Merlino G, Wakefield L. Expression of a dominant-negative mutant TGF-β type II receptor in transgenic mice reveals essential roles for TGF-β in regulation of growth and differentiation in the exocrine pancreas. EMBO (Eur Mol Biol Organ) J. 1997;16:2621–2633. doi: 10.1093/emboj/16.10.2621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chin J, Werb Z. Matrix metalloproteinases regulate morphogenesis, migration and remodeling of epithelium, tongue skeletal muscle and cartilage in the mandibular arch. Development. 1997;124:1519–1530. doi: 10.1242/dev.124.8.1519. [DOI] [PubMed] [Google Scholar]
- 6.Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987;162:156–159. doi: 10.1006/abio.1987.9999. [DOI] [PubMed] [Google Scholar]
- 7.Cirulli V, Beatens D, Rutishauser U, Halban P, Orci L, Rouiller D. Expression of neural cell adhesion molecule (N-CAM) in rat islets and its role in islet cell type segregation. J Cell Sci. 1994;107:1429–1436. doi: 10.1242/jcs.107.6.1429. [DOI] [PubMed] [Google Scholar]
- 8.Dahl U, Sjodin A, Semb H. Cadherins regulate aggregation of pancreatic β-cells in vivo. Development. 1996;122:2895–2902. doi: 10.1242/dev.122.9.2895. [DOI] [PubMed] [Google Scholar]
- 9.Deltour L, Leduque P, Paldi A, Ripoche M, Dubois P, Jami J. Polyclonal origin of pancreatic islets in aggregation mouse chimaeras. Development. 1991;112:1115–1121. doi: 10.1242/dev.112.4.1115. [DOI] [PubMed] [Google Scholar]
- 10.Giannelli G, Falk-Marzillier J, Schiraldi O, Stetler-Stevenson W, Quaranta V. Induction of cell migration by matrix metalloprotease-2 cleavage of laminin-5. Science. 1997;277:225–228. doi: 10.1126/science.277.5323.225. [DOI] [PubMed] [Google Scholar]
- 11.Gittes G, Galante P. A culture system for the study of pancreatic organogenesis. J Tissue Cult Methods. 1993;15:23–28. [Google Scholar]
- 12.Gittes G, Rutter W. Onset of cell-specific gene expression in the developing mouse pancreas. Proc Natl Acad Sci USA. 1992;89:1128–1132. doi: 10.1073/pnas.89.3.1128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Herrera P, Huarte J, Sanvito F, Meda P, Orci L, Vassali J. Embryogenesis of the murine pancreas; early expression of pancreatic polypeptide gene. Development. 1991;113:1257–1265. doi: 10.1242/dev.113.4.1257. [DOI] [PubMed] [Google Scholar]
- 14.Heussen C, Dowdle E. Electrophoretic analysis of plasminogen activators in polyacrylamide gels containing sodium dodecyl sulfate and copolymerized substrates. Anal Biochem. 1980;102:196–202. doi: 10.1016/0003-2697(80)90338-3. [DOI] [PubMed] [Google Scholar]
- 15.Hipps D, Hembry R, Docherty A, Reynolds J, Murphy G. Purification and characterization of human 72kDa gelatinase (type IV collagenase). Use of immunolocalisation to demonstrate the non-coordinate regulation of the 72-kDa and 95-kDa gelatinasse by human fibroblasts. Biol Chem Hoppe-Seyler. 1991;372:287–296. doi: 10.1515/bchm3.1991.372.1.287. [DOI] [PubMed] [Google Scholar]
- 16.Hisaoka M, Haratake J, Hashimoto H. Pancreatic morphogenesis and extracellular matrix organization during rat development. Differentiation. 1993;53:163–172. doi: 10.1111/j.1432-0436.1993.tb00705.x. [DOI] [PubMed] [Google Scholar]
- 17.Kaartinen V, Voncken J, Shuler W, Warburton C, Bu D, Heisterkamp D, Groffen J. Abnormal lung development and cleft palate in mice lacking TGF-beta 3 indicates defects of epithelial-mesenchymal interaction. Nat Genet. 1995;11:415–421. doi: 10.1038/ng1295-415. [DOI] [PubMed] [Google Scholar]
- 18.Kedinger M, Simon-Assmann P, Lacroix B, Marxer A, Hauri H, Haffen K. Fetal gut mesenchyme induces differentiation of cultured intestinal endodermal and crypt cells. Dev Biol. 1986;113:474–483. doi: 10.1016/0012-1606(86)90183-1. [DOI] [PubMed] [Google Scholar]
- 19.Kim S, Hebrok M, Melton D. Notochord to endoderm signaling is required for pancreas development. Development. 1997;124:4243–4252. doi: 10.1242/dev.124.21.4243. [DOI] [PubMed] [Google Scholar]
- 20.Lee M-S, Gu D, Feng L, Curriden S, Arnush M, Krahl T, Gurushanthaiah D, Wilson C, Loskutoff D, Fox H, Sarvetnick N. Accumulation of extracellular matrix and developmental dysregulation in the pancreas by transgenic production of transforming growth factor-β1. Am J Pathol. 1995;147:42–52. [PMC free article] [PubMed] [Google Scholar]
- 21.Lelongt B, Trugnan G, Murphy G, Ronco P. Matrix metalloproteinases MMP2 and MMP9 are produced in early stages of kidney morphogenesis but only MMP9 is required for renal organogenesis in vitro. J Cell Biol. 1997;136:1363–1373. doi: 10.1083/jcb.136.6.1363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Marti H, Lee L, Kashgarian M, Lovett D. Transforming growth factor-β1 stimulates glomerular mesangial cell synthesis of the 72-kd type IV collagenase. Am J Pathol. 1994;144:82–94. [PMC free article] [PubMed] [Google Scholar]
- 23.Middelhoven P, Ager A, Roos D, Verhoven A. Involvement of a metalloprotease in the shedding of human neutrophil Fc gammaRIIIB. FEBS Lett. 1997;414:14–18. doi: 10.1016/s0014-5793(97)00959-9. [DOI] [PubMed] [Google Scholar]
- 24.Miralles F, Czernichow P, Scharfmann R. Follistatin regulates the relative proportions of endocrine versus exocrine tissue during pancreatic development. Development. 1998;125:1017–1024. doi: 10.1242/dev.125.6.1017. [DOI] [PubMed] [Google Scholar]
- 25.Miralles F, Philippe P, Czernichow P, Scharfmann R. Expression of Nerve Growth Factor and its high affinity receptor Trk-A in the rat pancreas during embryonic and fetal life. J Endocrinol. 1998;156:431–439. doi: 10.1677/joe.0.1560431. [DOI] [PubMed] [Google Scholar]
- 26.Montesano R, Mouron P, Amherdt M, Orci L. Collagen matrix promotes reorganization of pancreatic endocrine cell monolayers into islet-like organoids. J Cell Biol. 1983;97:935–939. doi: 10.1083/jcb.97.3.935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Nakanishi Y, Suguira F, Kishi J, Hayakawa T. Collagenase inhibitor stimulates cleft formation during early morphogenesis of mouse salivary gland. Dev Biol. 1986;113:201–206. doi: 10.1016/0012-1606(86)90122-3. [DOI] [PubMed] [Google Scholar]
- 28.Orci L, Unger R. Functional subdivision of islets of langerhans and possible role of D-cells. Lancet. 1975;2:1243–1244. doi: 10.1016/s0140-6736(75)92078-4. [DOI] [PubMed] [Google Scholar]
- 29.Overall C, Wrana J, Sodek J. Independent regulation of collagenase, 72 kD progelatinase and metalloendoproteinase inhibitor expression in human fibroblasts by transforming growth factor-β. J Biol Chem. 1989;264:1860–1869. [PubMed] [Google Scholar]
- 30.Pictet, R., and W. Rutter. 1972. Development of the embryonic pancreas. In Handbook of Physiology. D.F. Steiner and N. Freinkel, editors. The Williams & Wilkins Co., Baltimore. 25–66.
- 31.Roberts A, McCune B, Sporn M. TGF-β: regulation of extracellular matrix. Kidney Int. 1992;41:557–559. doi: 10.1038/ki.1992.81. [DOI] [PubMed] [Google Scholar]
- 32.Ronco M, Chen Y, Goldfarb S, Ziyadeh F. Elevated glucose stimulates TGFβ gene expression and bioactivity in proximal tubule. Kidney Int. 1992;41:107–114. doi: 10.1038/ki.1992.14. [DOI] [PubMed] [Google Scholar]
- 33.Roubin R, Deagostini-Bazin H, Hirsch M-R, Goridis C. Modulation of NCAM expression by transforming growth factor-beta, serum and autocrine factors. J Cell Biol. 1990;111:673–684. doi: 10.1083/jcb.111.2.673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Rougier J, Moullier P, Piedagnel R, Ronco M. Hyperosmolality suppresses but TGFβ1 increases MMP9 in human peritoneal mesothelial cells. Kidney Int. 1997;51:337–347. doi: 10.1038/ki.1997.42. [DOI] [PubMed] [Google Scholar]
- 35.Salo T, Rahemtulla JLF, Birkedal-Hansen H, Larjava H. Transforming growth factor-B up-regulates type IV collagenase in culture human keratinocytes. J Biol Chem. 1991;266:11436–11441. [PubMed] [Google Scholar]
- 36.Sanford L, Ormsby I, Groot AG-d, Sariola H, Friedman R, Boivin G, Cardell E, Doetschmann T. TGFβ2 knockout mice have multiple developmental defects that are non-overlapping with other TGFβ knockout phenotypes. Development. 1997;124:2659–2670. doi: 10.1242/dev.124.13.2659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Sanvito F, Herrera P, Huarte J, Nichols A, Montesano R, Orci L, Vassali J. TGF-β1 influences the relative development of the exocrine and endocrine pancreas in vitro. Development. 1994;120:3451–3462. doi: 10.1242/dev.120.12.3451. [DOI] [PubMed] [Google Scholar]
- 38.Sanvito F, Nichols A, Herrera P, Huarte J, Wohlvend A, Vassali J, Orci L. TGF-β1 overexpression in murine pancreas induces chronic pancreatitis and, together with TNF-α, triggers insulin-dependent diabetes. Biochem Biophys Res Commun. 1995;217:1279–1286. doi: 10.1006/bbrc.1995.2906. [DOI] [PubMed] [Google Scholar]
- 39.Slack J. Developmental biology of the pancreas. Development. 1995;121:1569–1580. doi: 10.1242/dev.121.6.1569. [DOI] [PubMed] [Google Scholar]
- 40.Sosa-Pineda B, Chowdhury K, Torres M, Oliver G, Gruss P. The Pax4 gene is essential for differentiation of insulin-producing β cells in the mammalian pancreas. Nature. 1997;386:399–402. doi: 10.1038/386399a0. [DOI] [PubMed] [Google Scholar]
- 41.Spooner B, Cohen H, Faubion J. Development of the embryonic mammalian pancreas: the relationship between morphogenesis and cytodifferentiation. Dev Biol. 1977;61:119–130. doi: 10.1016/0012-1606(77)90285-8. [DOI] [PubMed] [Google Scholar]
- 42.Stewart H, Rougon G, Dong Z, Dean C, Jensen K, Mirsky R. TGF-βs upregulate NCAM and L1 expression in cultured Schwann cells, suppress cyclic APMP-induced expression of O4 and galactocerebroside, and are widely expressed in cells of the Schwann cell lineage in vivo. Glia. 1995;15:419–436. doi: 10.1002/glia.440150406. [DOI] [PubMed] [Google Scholar]
- 43.St-Onge L, Sosa-Pineda B, Chowdhury K, Mansouri A, Gruss P. Pax6 is required for differentiation of glucagon-producing α-cells in mouse pancreas. Nature. 1997;387:406–409. doi: 10.1038/387406a0. [DOI] [PubMed] [Google Scholar]
- 44.Sympson C, Talhouk R, Alexander C, Chin J, Clift S, Bissel M, Werb Z. targeted expression of stromelysin-1 in mammary gland provides evidence for a role of proteinases in branching morphogenesis and the requirement for an intact basement membrane for tissue-specific gene expression. J Cell Biol. 1994;125:681–693. doi: 10.1083/jcb.125.3.681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Werb Z. ECM and cell surface proteolysis: regulating cellular ecology. Cell. 1997;91:439–442. doi: 10.1016/s0092-8674(00)80429-8. [DOI] [PubMed] [Google Scholar]
- 46.Yamanaka Y, Friess H, Buchler M, Beger H, Gold L, Korc M. Synthesis and expression of transforming growth factor β-1, β-2, and β-3 in the endocrine and exocrine pancreas. Diabetes. 1993;42:746–756. doi: 10.2337/diab.42.5.746. [DOI] [PubMed] [Google Scholar]
- 47.Yoshinari M, Daikoku S. Ontogenic appearance of immunoreactive endocrine cells in rat pancreatic islets. Anat Embryol. 1982;165:63–70. doi: 10.1007/BF00304583. [DOI] [PubMed] [Google Scholar]
- 48.Ziyadeh F, Sharma K, Ericksen M, Wolf G. Stimulation of collagen gene expressionand protein synthesis in murine mesangial cells by high glucose is mediated by autocrine activation of transforming growth factor-β. J Clin Invest. 1994;93:536–542. doi: 10.1172/JCI117004. [DOI] [PMC free article] [PubMed] [Google Scholar]