

Abstract
KLP64D and KLP68D are members of the kinesin-II family of proteins in Drosophila. Immunostaining for KLP68D and ribonucleic acid in situ hybridization for KLP64D demonstrated their preferential expression in cholinergic neurons. KLP68D was also found to accumulate in cholinergic neurons in axonal obstructions caused by the loss of kinesin light chain. Mutations in the KLP64D gene cause uncoordinated sluggish movement and death, and reduce transport of choline acetyltransferase from cell bodies to the synapse. The inviability of KLP64D mutations can be rescued by expression of mammalian KIF3A. Together, these data suggest that kinesin-II is required for the axonal transport of a soluble enzyme, choline acetyltransferase, in a specific subset of neurons in Drosophila. Furthermore, the data lead to the conclusion that the cargo transport requirements of different classes of neurons may lead to upregulation of specific pathways of axonal transport.
Keywords: Drosophila, kinesin-like proteins, axonal transport, microtubule motors, neurobiology
Axonal transport is crucial for the movement of cellular components from their sites of synthesis in neuronal cell bodies to their sites of utilization at nerve termini. Fast axonal transport of organelles and vesicles requires a variety of molecular motors of the kinesin and dynein superfamilies (for reviews see Goldstein and Philp 1999; Goldstein and Yang 2000). These motor proteins generate active ATP-dependent transport along microtubule tracks. How soluble proteins in neurons are moved remains unknown, although there have been suggestions that motors such as kinesin or dynein might play important roles (Nixon 1998; Goldstein and Philp 1999; Goldstein and Yang 2000).
At present, most axonal kinesins are thought to play roles in anterograde axonal transport. The best evidence for such functions comes from studies of motors of the unc104/KIF1 family, kinesin heavy chain (KHC)1 family, and the kinesin-II/KIF3 family (for reviews see Hirokawa 1998; Goldstein and Philp 1999; Goldstein and Yang 2000). It has been proposed that these motors all have important functions in axonal transport and have radiated during evolution so that a common motor domain has acquired divergent tails to interact with different specific cargos. Recent examples of this mechanism in axonal transport include unc104/KIF1A and KIF1B, which appear to interact with vesicles carrying synaptic vesicle components and mitochondria, respectively (Hall and Hedgecock 1991; Otsuka et al. 1991; Nangaku et al. 1994; Okada et al. 1995). Of particular interest in this context are motors of the kinesin-II family, which are generally heterotrimeric complexes composed of two different motor subunits and a third nonmotor subunit. The nonmotor subunits of kinesin-II are generally referred to as kinesin-associated proteins or KAPs (Scholey 1996). The motor subunits of kinesin-II include KRP85 and KRP95 in sea urchin (Cole et al. 1993); KIF3A, KIF3B, and KIF3C in mouse (Kondo et al. 1994; Yamazaki et al. 1995; Yang and Goldstein 1998); FLA10 in Chlamydomonas (Walther et al. 1994); OSM3 in Caenorhabditis (Shakir et al. 1993); and KLP64D and KLP68D (Stewart et al. 1991; Pesavento et al. 1994) in Drosophila. Several of these motor subunits were tested for in vitro motility, and thus far, they invariably demonstrated plus end–directed microtubule dependent motility (Cole et al. 1993; Pesavento et al. 1994; Yamazaki et al. 1995). These observations, in concert with axonal ligation experiments, suggest a role for these motors in anterograde axonal transport (Kondo et al. 1994; Yang and Goldstein 1998).
Investigation of the cargos transported by kinesin-II has provided some intriguing clues. In nonneural cells, kinesin-II has been suggested to transport melanosomes in melanophores (Rogers et al. 1997), ER or Golgi components in the secretory pathway (Le Bot et al. 1998), and flagellar components in ciliated cells (Kozminski et al. 1993, Kozminski et al. 1995; Walther et al. 1994; Piperno et al. 1996; Vashishtha et al. 1996; Morris and Scholey 1997; Piperno and Mead 1997; Nonaka et al. 1998; Marszalek et al. 1999; Takeda et al. 1999). Interestingly, in cilia and flagella, the cargo transported may be in the form of protein complexes rather than membrane-bound organelles (Rosenbaum et al. 1999). Similarly, there is intriguing evidence that these motors may also be used in the evolutionarily divergent immotile sensory cilia in Caenorhabditis elegans (Tabish et al. 1995; Orozco et al. 1999; Signor et al. 1999). In neuronal axons, the kinesin-II cargo is unknown, but there is some evidence for a membrane-bound form based on fractionation and localization experiments (Henson et al. 1995; Yamazaki et al. 1995; Muresan et al. 1998; Yang and Goldstein 1998). However, at present, definitive evidence about the cargo of kinesin-II in axons is absent, as is proper understanding of its in vivo functions in nonciliated cells. Here we report our investigations of kinesin-II in Drosophila and present evidence that at least one function is the axonal transport of choline acetyltransferase (ChAT) in a subset of neurons.
Materials and Methods
Unless noted, all molecular biology experiments were done by standard methods (Sambrook et al. 1989). Fly husbandry and other related procedures were as described (Ashburner 1989; Goldstein and Fyrberg 1994). All fly stocks, unless otherwise mentioned, are described in Lindsley and Zimm 1992.
Cloning and Sequencing
A 150-bp PCR amplified KLP64D (KLP4 in Stewart et al. 1991) sequence was used as a probe to isolate several truncated cDNA clones from a Drosophila 0–4-h embryonic library cloned in pNB40 (Brown and Kafatos 1988). A composite 2.5-kb KLP64D coding sequence was created from several cDNA clones. Nested deletions were created in the cloned cDNA fragments in either direction using an ExoIII kit (Promega Corp.) according to the protocol provided by the manufacturers. Both strands were sequenced using either the T7 or the T3 primers, and later the whole parental clones were sequenced using the internal primers generated from the primary sequences. Several overlapping genomic fragments were isolated from a Drosophila genomic library in Lambda DASH® II using the KLP64D cDNA probe, and the coding sequences were further mapped to a 3.6-kb EcoRI/HindIII fragment, which was then cloned into pBluescript SK+ (Stratagene Inc.) and called pB11-4. Sequencing this genomic fragment revealed a single continuous open reading frame without any intron, and matched the KLP64D cDNA sequence.
Screen for KLP64D Mutants
Several deletions in the KLP64D gene were isolated previously by imprecise excision of P-transposable elements inserted at the 3′ end of the gene (Perez and Steller 1996). All mutant chromosomes were maintained over a TM3, Ser <y+> balancer in a yw background. We generated point mutations in the KLP64D gene by scoring failure of ethyl methanesulphonate (EMS)-induced lethal mutations to complement a deletion that removes the KLP64D gene, Df(3L)Klp64DA8 .n123. In brief, mutagenesis followed standard methods (Grigliatti 1986) in which 2-d-old adult isogenic w males were fed with 27 mM EMS in 2% sucrose solution for 16 h, put on fresh food for 1 d, and then groups of 10 males were mated to 40 w; TM3, Sb Ser/TM6B, Tb Hu (Lindsley and Zimm 1992) virgin females (60 matings). Individual male progeny that were TM3, Sb Ser/* or TM6, Tb Hu/* (where * indicates a mutagenized third chromosome) were then mated to four yw Df(3L)Klp64DA8 .n123/TM3, Ser <y+> virgins (9,000 matings). The progeny of these matings were screened for the absence of */Df(3L)Klp64DA8 .n123flies. TM3,Ser /* males from appropriate lines were then mated to yw, TM3, Ser <y+>/MKRS Sb virgins to create a stock. The mutagenized third chromosome was cleaned up and the lethal mutations mapped by recombination with a chromosome containing ru h th st cu sr e ca. We found eight recessive lethal mutants that failed to complement Df(3L)Klp64DA8 .n123 and that mapped to the KLP64D region. These lethal mutations fell into three complementation groups, one of which was found to identify the KLP64D gene.
Transgenic Rescue of KLP64D Mutant Phenotypes
A 3.6-kb KLP64D genomic fragment in pB11-4 was cleaved out by NotI and XhoI endonuclease digest and ligated to the same sites of pUAST (Brand and Perrimon 1993) and named pUAS64D. This maintained proper orientation of the KLP64D coding sequences with respect to the UAS sites in pUAST. We recovered several stable transformant lines (marked by w+ eye) in the X and in the second chromosome by injecting pUAS64D with a helper plasmid, containing the P-transposase gene, in yw embryos. These lines were then crossed to the Klp64D mutants. Rescue of lethality was scored both at the larval stages as well as in the adult, where the homozygous mutants are marked by yellow body color.
For rescue experiments using the mouse KIF3A gene, an inducible transgene was created by inserting a full length KIF3A cDNA (Kondo et al. 1994) obtained by standard methods between the EcoRI and the NotI restriction sites of the pUAST vector (Brand and Perimon, 1993). This arrangement placed the 5′ end of the KIF3A coding sequence near the GAL-UAS transcription activation sites. Stable transformant lines were then obtained by P-element–mediated transformation. Expression of the KIF3A transgene was induced in all neurons by Gal4 driven from the promoter of the elav gene inserted in the third chromosome (Luo et al. 1994).
RNA In Situ Hybridization
In situ hybridization was done according to Tautz and Pfeifle 1989, with modification as described in Pesavento et al. 1994 and Perez and Steller 1996. An antisense strand-specific ribo-probe was made from the 1.6-kb Spe I/Hind III fragment subcloned in pBluescript SK.
Antibody Purification by Antigen Affinity Column
An affinity column was prepared by cross-linking about 10 mg of truncated, bacterially expressed and purified glutathione S-transferase–KLP68D (Pesavento et al. 1994) to 1 ml Affigel-10 (Bio-Rad Laboratories) beads according to the protocol supplied by the manufacturer. These were packed in a 15-ml plastic column (Bio-Rad Laboratories) and blocked thoroughly with 100 ml of 10% BSA (Sigma Chemical Co.) in 10 mM Tris-Cl, pH 7.0, 100 mM NaCl, and 5 mM KCl (TBS-BSA), then washed in succession with 15 ml of 100 mM glycine, pH 2.5, TBS-BSA, 100 mM triethanolamine, pH 11.5, and finally twice with TBS-BSA. It was then loaded with the antiserum, washed twice with 15 ml of TBS-BSA, and the bound antibody was extracted with 1 ml of 100 mM glycine, pH 2.5 (Harlow and Lane 1988). We tried all three methods of 1 M MgCl2, 100 mM glycine, and 100 mM triethylamine extraction and found glycine to be the most suitable. The purified antibody was dialyzed against TBS overnight and stored at 4°C with 0.2% NaN2.
Protein Extraction, Western Blotting, and Immunoprecipitation
For Western blotting, ∼50 heads, thoraces, and abdomens from male and female adult flies were dissected and collected separately in plastic microcentrifuge tubes on ice. They were squashed in 500 μl sample loading buffer containing 0.1 M β-mercaptoethanol, 10% glycerol, and 0.1% bromophenol blue, and centrifuged at 14,000 g for 5 min at room temperature. The supernatant was boiled for 3 min and then separated by 10% SDS-PAGE and transferred to a polyvinylidene difluoride membrane (Harlow and Lane 1988). The Western blots thus prepared were stained with several primary antisera and corresponding peroxidase-coupled secondary antisera (Jackson Laboratories, Inc.), and developed using the ECL kit (Amersham Pharmacia Biotech) according to the protocol supplied by the manufacturer. For coimmunoprecipitation experiments, methods were essentially as described in Yang and Goldstein 1998. In brief, extracts of isolated Drosophila heads in RIPA buffer (50 mM Tris-Cl, pH 8.0, 1% NP-40, 0.1% SDS, 0.5% sodium deoxycholate, 150 mM NaCl) were subjected to immunoprecipitation, run on SDS polyacrylamide gels, and then probed in Western blots by standard methods.
Immunohistochemistry
Embryos were dechlorinated in 50% commercial bleach for 3 min and washed thoroughly in running distilled water, then fixed in heptane–fixative (2:1) interface as described in protocol 90 (Ashburner 1989) for 20 min at room temperature, and then devitelinized using methanol. The fixative contained 4% formaldehyde (EM grade) in 100 mM Pipes, pH 6.9, 1 mM EGTA, and 2 mM MgSO4. Larval tissues were dissected in Drosophila Ringers containing 130 mM NaCl, 4.7 mM KCl, 1.8 mM MgCl2, 0.7 mM KH2PO4, and 0.35 mM Na2HPO4 and then fixed in the same fixative as mentioned above for 20 min at room temperature.
Fixed tissue samples and embryos were washed with PBS and 0.1% Triton X-100 (PTX) and incubated with the primary antibody solution in PTX for 1 h. They were then washed with five changes of PTX and incubated with fluorescent secondary antibody (1:250 dilution in PTX; Jackson Laboratories, Inc.) for 1 h. Finally, after another wash with five changes of PTX, they were mounted with a drop of Vectashield™ mounting media (Vector Laboratories, Inc.). We used goat anti-HRP (1:1,000 in PTX; Cappel Inc.), purified rabbit anti-KLP68D (1:5 in PTX; Pesavento et al. 1994), mAb anti-ChAT (1:1,500 in PTX; Kitamoto et al. 1995), rabbit anti-synaptotagmin (SYT) (1:200 in PTX; Littleton et al. 1993), and mAb anti–cysteine string protein (CSP) (1:50 in PTX; Zinsmaier et al. 1994). The secondary antibodies alone do not stain anything in the control tissue.
The stained tissue samples were observed using a Bio-Rad MRC1024 laser scanning confocal device which was fitted to a Nikon Diaphot 200, and the images were captured by the Laser-Sharp 3.0 software. Images were further processed in Adobe Photoshop 4.0 for presentation. Most of the pictures contain data from 3–10-μm thick regions in the tissue.
Results
KLP64D Is the Drosophila Homologue of KIF3A and KRP85 and Belongs to the Kinesin-II Family
KLP64D (previously named KLP4) was identified during a PCR screen for new Drosophila genes with homology to the highly conserved regions of the KHC motor domain (Stewart et al. 1991). The gene was mapped to salivary gland chromosome band 64D. Northern analysis revealed a 2.6-kb mRNA enriched in the head (Stewart et al. 1991). Sequence of the full-length KLP64D coding region was obtained from several cDNA clones as well as a genomic clone. Analysis of the sequence indicated a 2032-bp open reading frame (ORF), which encodes a 674–amino acids (aa) long polypeptide with a predicted molecular mass of 76.4 kD. Secondary structure prediction analysis suggested three distinct domains in the polypeptide, including a globular NH2-terminal domain of 385 aa, an alpha-helical coiled-coil central domain of 200 aa, and a globular COOH-terminal tail domain of ∼100 aa. The KLP64D sequence is most similar to the kinesin-II subunit KIF3A (94% similarity) of mouse and KRP85 (93% similarity) of sea urchin (Fig. 1). Considerable similarity (66%) was found with another Drosophila kinesin-II polypeptide encoded by the KLP68D gene (Pesavento et al. 1994). Therefore, we placed KLP64D in the kinesin-II family with KIF3A and KRP85 as the likely homologues. This assignment is also supported by the finding that the lethality of KLP64D mutants can be rescued by expression of mouse KIF3A (see below).
Figure 1.

The protein sequence of KLP64D is homologous to KIF3A of mouse and KRP85 of sea urchin. The identical sequences are shown as white letters in black background, whereas functionally similar ones are marked with gray boxes in the background. Multiple sequence alignments were done using the pileup program of the GCG analysis package with default options set. Sequence identities were highlighted using Box shade 3.21 shareware provided by the ISREC Bioinformatics Group (http://www-isrec.unil.ch/isrec.htm). The location of Klp64D mutant changes are noted.
KLP64D and KLP68D Are Expressed in a Subset of Peripheral and Central Nervous System Neurons
To obtain clues about the potential functions of kinesin-II in neurons, we examined the expression patterns of KLP64D and KLP68D in the developing Drosophila nervous system. In overview, we found that KLP64D and KLP68D are both expressed in a restricted set of neurons, which appear to correlate with the cholinergic subset.
In situ hybridization to RNA revealed that KLP64D is expressed in neurons of both the central (CNS) and peripheral (PNS) nervous system. KLP64D RNA was first observed in the CNS at stage 12 (Fig. 2 A, arrows) when many CNS neurons begin extending their processes (Broadie and Bate 1993; Goodman and Doe 1993; Campos-Ortega and Hartenstein 1997). Later in embryogenesis, at stage 16, KLP64D expression continues in the CNS, including the optic lobes (Fig. 2 B, large arrow). Prominent expression was also found in the PNS, where KLP64D RNA was observed in the cells of the dorsal (Fig. 2 B, arrowheads) and lateral chordotonal organs (Fig. 2 B, small arrows). Careful observation revealed that the staining in the abdominal lateral chordotonal (lch5) organs (Fig. 2 C, arrows) was restricted to the neuronal cells. In addition, we also found staining in the ventral and dorsal chordotonal neurons, the anterior sense organ and in the antenno-maxillary complex (data not shown). Expression in the CNS and PNS continues in the first instar larva and was still detectable in the third instar larval brain, although at a lower level (Fig. 2 D). Expression in the third instar brain includes cells of the ventral ganglion (Fig. 2 D, arrowheads) and a few cells in the optic lobe (Fig. 2 D, arrow).
Figure 2.

KLP64D gene expression pattern revealed by RNA in situ hybridization with a specific probe encoding the COOH terminus made from a 1.6 kb Spe I/HindIII fragment, labeled with digoxygenin-dUTP, and visualized using alkaline phosphatase activity linked to an antidigoxygenin antiserum. Bars: A, 100 μm (A, B, and D); C, 25 μm. (A) Arrows indicate staining in the developing CNS of a stage 12/13 embryo when germ band retraction is partly complete. The anterior side of the embryo is towards the left and the ventral side is down. (B) The expression in the PNS is first detected at late stage 17 of embryogenesis, after the completion of germ band retraction and dorsal closure. Most of the neuronal development is complete by this time. The small arrows indicate staining in the lateral chordotonal (lch) neurons. All eight lch5 cells of the abdominal segments and one lch3 in the third thoracic segment are visible. The arrowheads indicate dorsal chordotonal organs in each of the abdominal segments. The large arrow indicates the optic lobe in the CNS. (C) An enlarged view of lateral pentascolopedial chordotonal organs (lch5) of a late stage 17 embryo stained by the KLP64D probe revealed the cytoplasmic outline of the cells. There are five lch5 neurons clustered together at the basal side of the epidermis. (D) Staining in the brain of a third instar larva. A low level of expression is seen in the cells of the ventral ganglion (arrowheads) and in the optic lobe (arrow). Arrows indicate the staining in the optic lobe region where inputs from the bolwig nerve as well as the photoreceptor neurons of the larval eye disc is received. A faint staining is also observed in the crescent shaped developing lamina region of the optic lobe.
Antibody staining revealed preferential expression of KLP68D in a subset of neurons similar to those shown to express KLP64D, and shown previously to express KLP68D RNA (Pesavento et al. 1994). The antibody used was an affinity-purified antiserum raised against the 432-aa COOH-terminal domain of KLP68D (Pesavento et al. 1994). In Western blots of adult head lysate, the purified serum stained a single band of 90 kD, which is close to the expected molecular mass of full-length KLP68D. In stage 17 embryos, the purified KLP68D antibody strongly stained a subset of PNS neurons, including chordotonal neurons, cells in the antenno-maxillary complex (Fig. 3A, Fig. C, and Fig. E), bolwig organ cells (Fig. 3 A, asterisk), and other sensory neuron cells in the epidermis. To visualize all neurons, the same embryos were also stained with anti-HRP antibody (Fig. 3B, Fig. D, and Fig. F), which, as expected, labeled the cytoplasm and membranes of all neuronal cell bodies and processes (Jan and Jan 1982). This double staining experiment helped establish the identity of the KLP68D-positive sensory neurons in the epidermis (Fig. 3), and revealed that KLP68D may be specific to a subset of PNS sensory neurons, whose common thread may be that they are all cholinergic (see below).
Figure 3.
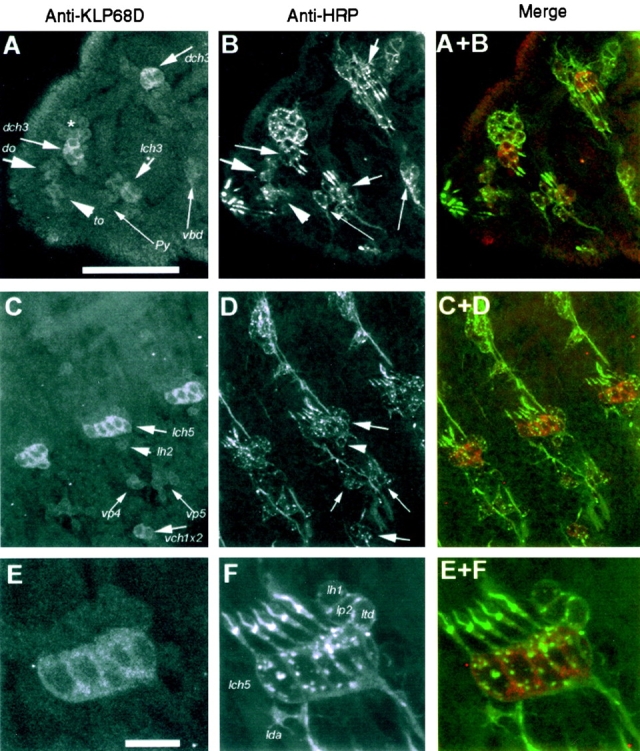
Immunostaining pattern of rabbit anti-KLP68D (A, C, and E) and anti-HRP (B, D, and F) in the epidermis of a late stage 16 embryo. The anterior side of the embryo in all the figures is placed towards the left and the dorsal side is upwards. The KLP68D antiserum stained neuronal soma of a subset of sensilla while anti-HRP marked all the sensory neurons in the epidermis (Jan and Jan 1982). Sense organ nomenclature is followed as in Campos-Ortega and Hartenstein 1997. Bars: A, 50 μm (A–D); E, 10 μm (E and F). (A) The anterior part of the embryo is shown, including the first two thoracic segments. Relatively strong staining of the dorsal (dch3) and lateral (lch3) chordotonal organ neurons in the first and second thoracic segments is evident. The polyinnervated external sense organ (vbd) and external papilla (Py) in the second and first thoracic segments, respectively, are only weakly stained. Similarly, the dorsal (do) or antennalles organ and the terminal organ (to) or maxillarorgan are weakly stained. Most of the sensory cells of these two organs are just beyond the plane of focus. The faint staining observed in a group of cells (indicated by an asterisk) in the dorsal region of the first thoracic segment are identified as bolwig organ. (B) The same region as in A labeled with goat anti-HRP, which stains all neuronal cell bodies and processes and clearly shows that only a subset of neurons are stained for KLP68D. (C) Staining in the abdominal segments is shown. The cell bodies of the lateral (lch5) and the ventral (vch1x2) chordotonal neurons are relatively strongly stained. The neuronal cell bodies of ventral papillae (vp4 and vp5) and the lateral trichoid sensillum (lh2) are less intensely stained. In addition, there is weak staining in some unrecognized cells in between vp4 and vp5. (D) Same field as in C stained with anti-HRP. (E) A high magnification image of a region of the lateral epidermis containing the pentascholopedial organ (lch5) and other sensilla. Staining is restricted to the neuronal cytoplasm of the lch5. (F) The same field as in E stained with anti-HRP, which highlights all the neurons. Identified neurons are marked in the figure. The neurons of the monoinnervated external sensilla, lh1 and lp2, and the multi-dendritic neurons, ltd and lda, are not stained significantly relative to lch5.
KLP68D was also detectable in the CNS starting at stage 16 in the embryo, where it was most pronounced in the first instar larva stage in the ventral ganglion as well as in the optic lobe. Unlike the sensory neurons, the expression in the CNS was not distinguishable in specific cell types, owing to the small size and high density of cell bodies in this region. Some cells in the optic lobe were strongly stained early in the first instar stage, but the staining was not seen at a later stage. In the ventral ganglion, uniform staining appeared in the neuropil and cell cortex. No specific staining was detectable in CNS at the third instar stage.
A comparison of the KLP68D and KLP64D expression patterns (Fig. 2) suggests that both KLP64D and KLP68D are expressed in similar tissue subsets and most likely in the same set of cells. However, the overlap could be determined with certainty only in the neurons of the chordotonal organs (Fig. 2 C and Fig. 3 E), bolwig organ cells, and in the antenno-maxillary complex. We noted that the KLP64D and KLP68D distribution was similar to that of acetyl cholinesterase RNA expression (Zador 1995) and ChAT RNA and lacZ reporter expression in the PNS (Yasuyama et al. 1995), which are good markers for cholinergic presynaptic neurons in Drosophila and other insects. The neurons innervating the chemosensory, chordotonal, and larval bolwig organs (Kitamoto et al. 1995; Yasuyama et al. 1995) are known to be cholinergic. In addition, many interneurons of the CNS also express ChAT and acetyl cholinesterase (Yasuyama et al. 1995; Zador 1995). Unlike vertebrates, motor neurons in Drosophila generally use glutamate instead of acetylcholine as their neurotransmitter (Johansen et al. 1989). Similar to what others have found, our efforts to localize ChAT antigen itself in the neuronal cell bodies of the PNS and CNS using ChAT antiserum were not successful, and so we could not determine the overlap of ChAT and KLP68D in the cell bodies with precision. However, since all of the cholinergic synapses are located in the CNS, ChAT staining was detected in the synaptic bulbs of the CNS neuropil (see Fig. 5 A), which provided an assay for ChAT transport in KLP64D mutants (see below).
Figure 5.

Coimmunoprecipitation analysis. Immunoprecipitation from wild-type (WT) and mouse KIF3A expressing (3A+) Drosophila using K2.4 mAb raised against sea urchin KRP85, affinity-purified anti-KLP68D, or nonspecific Ig. Western blots of immunoprecipitates were probed with K2.4 or affinity-purified anti-KLP68D.
KLP68D Coaccumulates with ChAT in Axonal Obstructions
To test further whether KLP68D and ChAT are coexpressed in cholinergic neurons, we used kinesin light chain (Klc−) mutant larvae. Homozygous Klc mutant larvae die at the late second instar and early third instar stages with progressive distal paralysis. The axons of these larvae develop focal swellings along their length and contain accumulated cargos of various types that appear to block single axons in axon bundles (Hurd and Saxton 1996; Gindhart et al. 1998). We observed numerous KLP68D and ChAT immunoreactive clogs in the axons of these animals (Fig. 4A and Fig. B, arrows), which probably result from the blockage of ChAT and KLP68D transport in cholinergic neurons. Quantitation in several nerve bundles revealed strong coincidence between ChAT immunoreactivity and that of KLP68D in the axons (Table ). This observation confirms the suggestion that both ChAT and KLP68D are expressed in the same cells and transported in the same axons. These findings are significantly different than observed for ChAT and a ubiquitous axonal cargo, SYT (Fig. 4, C–F; Table ). Although there are a large number of SYT-positive clogs in the axon, only a small percentage (37.4%) are also ChAT-positive (Table ; Fig. 4E and Fig. F, arrows). These data suggest that not all axons and axonal clogs contain ChAT and therefore, that the observed ChAT and KLP68D association is not fortuitous.
Figure 4.
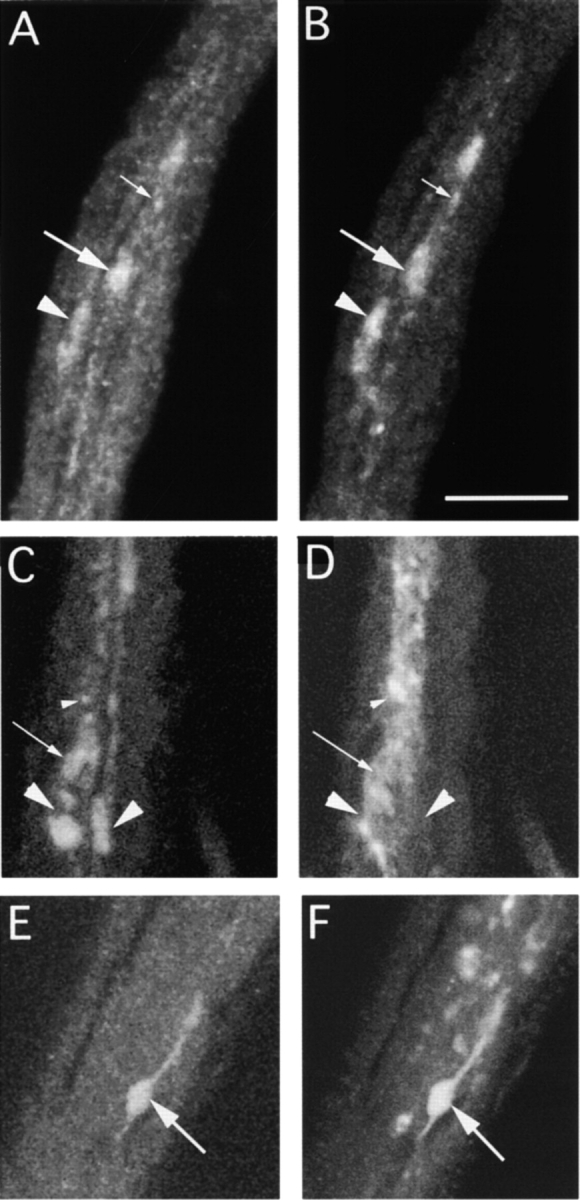
Segmental nerve bundles from a Klc1/Df(3L)8ex94 third instar larva, simultaneously stained with either mouse anti-ChAT (A) and rabbit anti-KLP68D (B) or with anti-ChAT (C and E) and rabbit anti-SYT (D and F). They are visualized using FITC anti–mouse and Cy5 anti–rabbit. Both frames are simultaneously excited and captured by separate photomultipliers in a single optical scan of 1-μm thickness. Accumulations of the respective antigens in the axons in individual clogs are marked by different sized arrows and arrowheads. Both KLP68D and ChAT antisera usually stain the same foci (clogs) in the nerve roots (see Table for details). SYT generally does not associate (arrowheads in C and D) with the ChAT in these foci although occasional coincident staining is seen (arrows in E and F). Bar, 10 μm.
Table 1.
| No. SYT+ ChAT− clogs | No. SYT− ChAT+ clogs | No. SYT+ ChAT+ clogs | Percentage of colocalization among ChAT clogs | No. Nerves |
|---|---|---|---|---|
| >800 | 36 | 22 | 38% (22/58) | 11 |
| No. KLP68D+ ChAT− clogs | No. KLP68D− ChAT+ clogs | No. KLP68D+ ChAT+ clogs | Percentage of colocalization among ChAT clogs | No. Nerves |
| 4 | 3 | 47 | 94% (47/50) | 14 |
Relative distribution of KLP68D and SYT antigen with respect to ChAT in Klc1/Df(3L)8ex94 axons where these antigens accumulate in focal swellings (clogs). The numbers of clogs that stained positively only for SYT (SYT+ ChAT−), only for ChAT (SYT− ChAT+), or both (SYT+ ChAT+) are shown. The relative overlap (percentage of colocalization among ChAT clogs) of either SYT or KLP68D staining with that of ChAT is expressed as the percentage of doubly stained clogs among the total number of ChAT positive clogs. Three individual larvae were examined for each set.
Since other members of the kinesin-II family are often associated in heterodimeric complexes, we conducted coimmunoprecipitation experiments to probe the association of KLP68D and KLP64D. These experiments were complicated by the absence of an antibody specific for KLP64D and the lack of cross-reactivity in Western blot experiments of antibodies reactive with the KIF3A and KRP85 homologous proteins (data not shown). However, it was possible to express functional mouse KIF3A in Drosophila using the GAL4-UAS system (Brand and Perrimon 1993), and to demonstrate that KIF3A rescues KLP64D mutants (see below). Flies expressing KIF3A could then be used in coimmunoprecipitation experiments. Immunoprecipitations using the K2.4 mAb raised against sea urchin KRP85 (Cole et al. 1993) precipitated both KIF3A and KLP68D in flies expressing KIF3A, but not in wild-type (Fig. 5). Complementary immunoprecipitations using anti-KLP68D also precipitated both KIF3A and KLP68D in flies expressing KIF3A, but not in wild-type (Fig. 5). Control immunoprecipitations precipitated no immunoreactive material in any genotype. Thus, we propose that KLP64D, which is the sequence and functional homologue of KIF3A, also associates with KLP68D in vivo.
Mutations in the KLP64D Gene Cause Uncoordinated Movements and Death
Previous work led to the generation and characterization of a set of deletions in and around the KLP64D gene (Perez and Steller 1996). These deletions were obtained by imprecise excision of several P-transposable elements inserted near the 3′ end of the KLP64D gene (Perez and Steller 1996). One such deletion, Df(3L)Klp64DA8 .n123, has a left breakpoint that extends to the extreme 5′ end of the KLP64D protein coding sequence. To generate KLP64D mutations, we used EMS mutagenesis and screened for lethal mutants that failed to complement Df(3L)Klp64DA8 .n123. We isolated eight mutants that fell into three different complementation groups. Meiotic recombination mapping with respect to roughoid and hairy placed the lethality in all of these chromosomes near the KLP64D gene. One of the complementation groups containing three of the EMS alleles, Klp64Dk1, Klp64Dk5, and Klp64Dk33, identifies the KLP64D gene, since these mutants show changes in the KLP64D coding sequence. Specifically, Klp64Dk1has a change from Gln (CAA) to a stop (UAA) at codon 13, Klp64Dk5has a change from a highly conserved Glu (GAG) to Lys (AAG) at codon 551, and Klp64Dk33 has a change from T to G in the 3′ untranslated region 4 bp after the predicted normal stop codon. We presume that Klp64Dk33 causes mRNA instability, since its phenotype is similar to that of Klp64Dk1, which is likely to be a null-mutant since it has a stop codon early in the gene. Heteroallelic combinations among these mutants give some complementation and a few very uncoordinated and near-paralyzed adults.
All three KLP64D mutants are homozygous lethal. In a typical crowded culture, these mutants die at or before the early third instar larval stage. If the mutants are raised in conditions of low density and without competition from wild-type larvae, a few mutant animals will survive to be pharate pupae and occasionally eclose as fully formed adults. All of these mutant adults exhibited an acutely uncoordinated walk, are unable to stand, and died in a few hours. Mutant third instar larvae exhibit slight to severe sluggishness, and roll abnormally from side to side during crawling. These observations suggest a neuronal function of the KLP64D gene.
To confirm that mutations in the KLP64D gene were responsible for the lethality and other behavioral defects, we used genetic rescue experiments. Although large KLP64D genomic constructs were unstable in bacteria, we were able to construct pUAS64D, which contained an ∼3.6 kb EcoR1/HindIII genomic fragment with a full-length KLP64D coding sequence, 447 bp of 5′ upstream sequences, including the TATA box, and about 0.8 kb of 3′ sequences in pUAST (Brand and Perrimon 1993). We reasoned that expression of KLP64D from this construct would be initiated from its own promoter if the necessary enhancer element is present within the limited 5′ upstream region. Alternatively, the GAL4-UAS at the 5′ end can be used to drive KLP64D expression through the tissue-specific expression of specific promoter–Gal4 fusion genes (Brand and Perrimon 1993). We obtained several independent stable transformants and crossed them to various Klp64D alleles to test whether they can rescue the lethality and other abnormalities. The pUAS64D construct by itself rescued the lethality and uncoordinated behavior of Klp64Dk1 and Klp64Dk5, either in homozygous condition or hemizygous with the Df(3L)Klp64DA8 .n123 chromosome. These results confirm that the lesions in the KLP64D gene caused the lethality and the uncoordinated behavior.
We also used genetic rescue experiments to establish further that KLP64D is a true functional homologue of KIF3A and KRP85. Thus, a complete KIF3A cDNA was placed under the control of GAL4-UAS sequences and introduced into the Drosophila genome by P-element transformation. Then we combined this transgene with a transgene encoding GAL4 driven by the neural elav promoter (Luo et al. 1994). This combination of elements gave complete rescue of Klp64Dk1 heterozygous with a Df(3L)Klp64DA8 .n123 chromosome. Thus, when yw/Y; UAS-Kif3A<w+>/+; Df(3L)Klp64DA8 .n123 elav-Gal4 <w+>/TM3, Ser e <y+> was crossed to yw/yw; +/+; Klp64Dk1/TM3 Ser e <y+>, 92 progeny carrying UAS-Kif3A<w+>/+; Df(3L)Klp64DA8 .n123 elav-Gal4 <w+>/Klp64Dk1were recovered, whereas no progeny carrying +/+; Df(3L)Klp64DA8 .n123 elav-Gal4 <w+>/Klp64Dk1were recovered out of a total of 414 progeny, which is well within expected proportions.
KLP64D Is Needed for Axonal Transport of ChAT
The expression pattern and the behavioral defect in Klp64D mutant animals led to the hypothesis that the KLP64D motor complex might be involved in transporting the components of the acetylcholine-producing machinery. We tested this view by staining Klp64D mutant animals with ChAT antiserum. In wild-type third instar larvae, we found ChAT in abundance at many synapses in the neuropil of the ventral nerve chord and in the optic lobes of the larval brain (Fig. 6 A), whereas very little was observed in the cell bodies of the brain cortex. Counter staining with rhodamine-labeled α-bungarotoxin, which binds to the nicotinic acetylcholine receptors (nAChR) (Schloss et al. 1992), showed that the ChAT antigen is distributed in the neuropil similar to nAChR (Fig. 6 E). No ChAT immunoreactivity was observed in the axons (Fig. 6 C) of wild-type larvae. This staining pattern is consistent with previous reports (Yasuyama et al. 1995). In Klp64Dk1 and Klp64Dk5homozygous third instar larvae, we observed accumulation of ChAT immunoreactivity in the cell cortex (Fig. 6 B, arrowheads) of the ventral ganglion. Closer examination of these mutant samples revealed considerable ChAT immunogen in the nerve roots (Fig. 6B and Fig. D, arrows) and in the cell bodies (Fig. 6 B, arrowheads). In some samples, the ChAT immunoreactivity in the neuropil was reduced strikingly (data not shown) relative to the wild-type control. The abnormal staining pattern suggests that loss of KLP64D affects ChAT transport to the synapse, since no such ChAT staining is observed in the wild-type axons (Fig. 6 A, arrows). The α-bungarotoxin (Schloss et al. 1992) staining of nAChR on the postsynaptic membrane in the Klp64Dk1 homozygous mutants (Fig. 5 F) showed no detectable difference from wild-type (Fig. 6 E) control.
Figure 6.
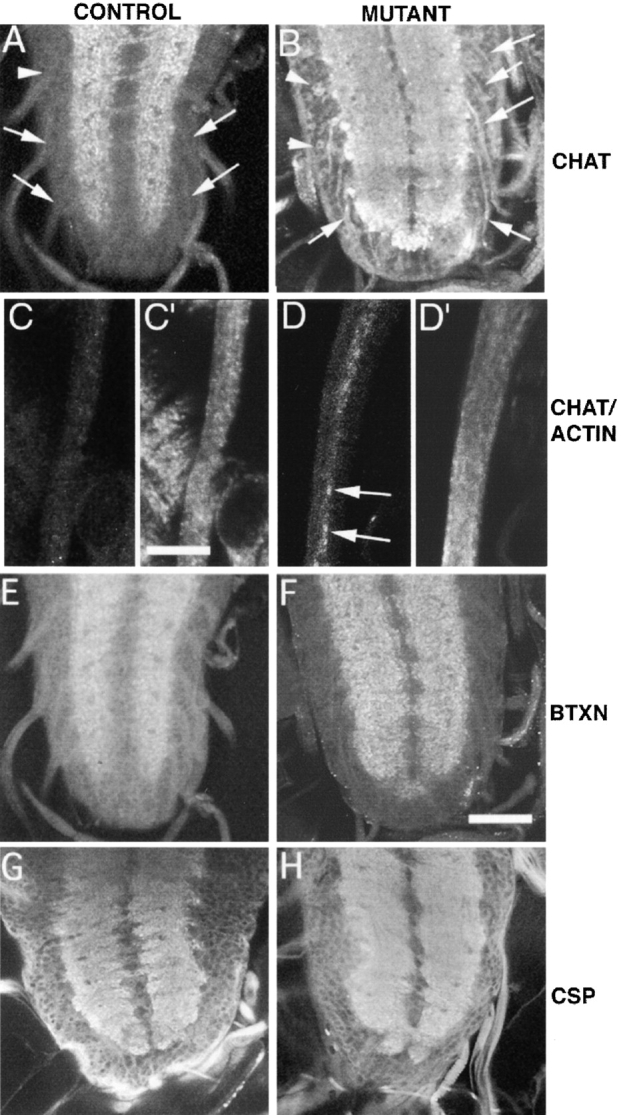
Staining of various markers of axonal transport in the cell bodies and axons of heterozygous (A and C) and homozygous (B and D) klp64D mutant larval brains and segmental nerves. Samples are stained with mouse anti-ChAT (A–D), XTRITC-coupled α-bungarotoxin (E and F) and with mouse anti-CSP antibody (G and H), respectively. A few were also stained with rabbit antiactin antibody (C′ and D′). Bars: C′, 10 μm (C, C′, D, and D′); F, 100 μm (A, B, E–H). (A) Klp64Dk1/TM3 third instar larva ventral ganglion has anti-ChAT staining confined to the neuropil. The punctate nature of the staining indicates localization of ChAT in synaptic bulbs. There was no staining in the cell cortex (arrowhead) or in the nerve roots (arrows). (B) The homozygous Klp64Dk1third instar larval brain exhibited significant ChAT staining in the axons (arrows) of the nerve roots and in the cell cortex (arrowheads). Abnormally strong staining can also be seen at the boundary of the neuropil. (C and C′) A part of a segmental nerve bundle is shown from a Klp64Dk1/TM3 larva stained with mouse anti-ChAT and rabbit antiactin antibody, respectively. Both cases showed no specific accumulation of these two antigens in any of the axons. (D and D′) The equivalent part of a similar nerve bundle from a Klp64Dk1 homozygous larva showed strong accumulations of ChAT antigen in some of the axons (arrows). The antiactin staining in the axons is slightly higher in intensity than the control, but it is not selectively higher in the axons that showed accumulated ChAT as shown in F. E–H show equivalent regions of ventral ganglia from Klp64Dk1/TM3 (E and G), and the Klp64Dk1 homozygous third instar larvae, respectively. They are stained with α-bungarotoxin (E and F) and anti-CSP (G and H) and the pattern is similar between the control and mutants.
We tested whether the loss of ChAT immunoreactivity at the synapse and the abnormal accumulation in the axons are a secondary effect of the lethality or other neural defects in the mutant animals lacking KLP64D. We examined Khc − and Klc − larvae, which die at the second instar and third instar larva stage, respectively, with significant axonal transport defects (Hurd and Saxton 1996; Gindhart et al. 1998). We found that the intensity of ChAT staining in late second instar Khc − as well as in late third instar Klc − brains was not significantly reduced or altered as compared with the wild-type. Hence, it is unlikely that reduction of ChAT immunoreactivity is a secondary effect of lethality or general axonal transport defects.
To test whether ChAT accumulation in Klp64Dk1axons is a specific effect of KLP64D malfunction or a result of general axonal clogging, we stained Klp64Dk1 homozygous larvae at the third instar stage with antiactin (Fig. 6C′ and D′), anti-SYT (data not shown) (Littleton et al. 1993), and anti-CSP (Fig. 6G and Fig. H, and Fig. 5 H; Zinsmaier et al. 1994). We used Klp64Dk1/TM3 larvae as the wild-type control (Fig. 6 G). Since both SYT and CSP are integral parts of synaptic vesicles, they are transported to all chemical synapses and are likely to accumulate in general axonal clogs as seen in Khc and Klc mutants (Gindhart et al. 1998; Hurd and Saxton 1996). We found no accumulation of SYT and CSP in the axons of the Klp64D mutant larvae (Fig. 6 H). In addition, we also stained the larvae with anti-HRP, antitubulin, and antiactin (Fig. 6 D′) and found no detectable accumulation in the axon. Finally, we also noted that ChAT accumulation in Klp64D mutant axons is distinct from the clogs found in Klc or Khc mutants (see Fig. 4), since these accumulations do not cause obvious swelling of the axons. In fact, EM observation suggests that general CNS neuronal and axonal morphology is normal in Klp64D mutants (data not shown). Therefore, mutations in the Klp64D gene appear to cause selective accumulation of ChAT as opposed to general clogging. In fact, these results suggest that there are probably no clogs in these axons, which is dissimilar to Klc and Khc mutants.
Discussion
In this paper we report two principal findings. First, we found that KLP64D and KLP68D, which appear to be mutually associated motor polypeptides of the kinesin-II family, are expressed preferentially in cholinergic neurons in Drosophila. Second, we found that mutations in the KLP64D gene cause accumulations of ChAT in neuronal cell bodies and axons. Since other apparently generic axonal cargos do not appear to accumulate in these mutants, and since the overall neural morphology of mutant animals is relatively normal, the data suggest that these kinesin motor proteins are required for transport of ChAT, and perhaps a small number of other cargos in axons of cholinergic neurons. There are several important implications of these findings.
A striking conclusion that can be drawn from the preferential expression data and the relative selectivity of the axonal transport defect in KLP64D mutants is that KLP64D and KLP68D in Drosophila are required for axonal transport of a subset of axonal cargos, one of which is ChAT. This conclusion suggests that a common assumption about the general organization of the axonal transport machinery may be incorrect. That is, axonal transport is usually thought of as a neuronal process that uses a general machinery of neuronal motors, adapters, and carriers for transport of both specialized and ubiquitous axonal components (Hirokawa 1998). Consistent with this view, there is good evidence for the existence of a general axonal transport machinery that uses ubiquitous neuronal kinesins such as unc104/KIF1A, KIF1B, and various forms of true kinesin to transport cargos used in all neurons such as mitochondria, synaptic vesicle components, and channels (Hall and Hedgecock 1991; Otsuka et al. 1991; Nangaku et al. 1994; Hurd and Saxton 1996; Yonekawa et al. 1998). However, our data suggest that there also may be specialized transport pathways activated or enhanced in specific classes of neurons. These neuronal classes may be defined by neurotransmitter or other phenotype, and may activate specific motors, adapters, and/or carriers for axonal transport. While kinesin-II in Drosophila (KLP64D and KLP68D) may provide the first recognizable example of such a cell type–specific axonal transport system, there may be others awaiting recognition or discovery.
Although the reported expression patterns of kinesin motor proteins in vertebrate systems do not provide a clear test of the hypothesis that some motors are harnessed in cell type–specific axonal transport pathways, there are a few relevant anecdotal observations in the literature. In particular, KIF3B, KIF3C, and neuronal KHC have been reported to have expression patterns that are not uniform among neurons (Niclas et al. 1994; Muresan et al. 1998; Yang and Goldstein 1998). KIF3B was reported to be expressed in a subset of Purkinje cells (Muresan et al. 1998), whereas neuronal KHC exhibits greatly elevated expression in a subset of neurons (Niclas et al. 1994). However, at present, in the absence of double staining experiments with good markers of neuronal phenotype, it is difficult to gauge whether some kinesins have strict cell type specificity in vertebrate neurons. Nonetheless, it is clear that even ubiquitous neuronal motor proteins, such as neuronal KHC, may not be expressed equivalently in all neuronal types. Although some of this nonuniformity in expression might be a consequence of overall transport requirements in large neurons, some of the heterogeneity might result from the need for cell type–specific axonal transport pathways in phenotypically differentiated neurons.
Kinesin-II Functions
There is considerable information regarding the function of kinesin-II family members in several organisms. In particular, a variety of data suggest that motors of the kinesin-II family are specialized for the transport of cytosolic, nonmembrane enclosed constituents such as dyneins or central pair components of cilia and flagella. In Caenorhabditis, kinesin-II motors are needed for the function of sensory cilia as demonstrated by the phenotype of osm-3 mutants (Shakir et al. 1993; Tabish et al. 1995). In sea urchins, antibody microinjection experiments demonstrate the requirement for these motors in transport of ciliary components, perhaps those of the central pair microtubules (Morris and Scholey 1997). In vertebrates, motors of this family have been found in the immotile dendritic sensory cilium of the rod photoreceptor, where they could move ciliary or sensory components (Beech et al. 1996) and are required to form early embryonic cilia of the node (Nonaka et al. 1998; Marszalek et al. 1999; Takeda et al. 1999). Finally, in Chlamydomonas, there is good evidence to support the view that kinesin-II motors are needed to drive the movement of intraflagellar particles or rafts of axonemal components such as inner arm dyneins (Kozminski et al. 1993, Kozminski et al. 1995; Walther et al. 1994; Piperno et al. 1996; Piperno and Mead 1997). Together, these data have suggested a requirement for kinesin-II in transport in ciliated cells or neurons with degenerate dendritic sensory cilia. What has been unclear is the possible role of this family of motor proteins in axons of sensory and nonsensory neurons.
Our work in Drosophila suggests that a unifying theme for kinesin-II motors may be that they are specialized to move cytosolic proteins in motile cilia and flagella, modified immotile dendritic sensory cilia, and in certain classes of axons. Since ChAT is generally thought to be a cytosolic protein, its dependence upon kinesin-II in Drosophila for transport may reflect this specialization. There is also evidence that kinesin-II motors may be associated with membranous components (Henson et al. 1995, Henson et al. 1997; Yamazaki et al. 1995; Le Bot et al. 1998; Muresan et al. 1998; Tuma et al. 1998; Yang and Goldstein 1998). Thus, kinesin-II motors could, in principle, move both complexes of cytosolic protein components in axons and cilia as well as interact with membranous vesicles. Alternatively, the complexes of cytosolic proteins could interact intermittently with membranes to give the observed associations, similar to the suggestion that has been made for bead movements on the surface of Chlamydomonas flagellar membranes (Kozminski et al. 1995). An intriguing possibility in this regard is that the combinatorial interactions of kinesin-II polypeptides in obligate heterodimers might allow some forms to associate with soluble complexes and some to associate with membranous vesicles (Muresan et al. 1998; Yang et al. 1997). In this context, it remains formally possible that ChAT in Drosophila is not moved as a soluble protein complex, but instead as a “hitch-hiker” on membranous vesicles. Further work will be necessary to fully resolve these issues.
Kinesin-II and Slow Axonal Transport
The transport of soluble proteins such as ChAT is normally attributed to the slow axonal transport machinery, which is driven by an unknown mechanism (Nixon 1998). However, past work is not conclusive on the question of whether ChAT is transported in the fast or slow axonal transport compartment (Malatova et al. 1989; Tandon et al. 1996), and at present there is no evidence as to whether Drosophila has a slow axonal transport pathway. An economical but speculative hypothesis is that although kinesin-II motors are generally thought of as fast axonal transport motors based on their speed of movement in vitro and on the velocity of particle transport in flagella, they could, in principle, also drive some components of the slow transport system. For example, if ChAT-containing particles spent only a small fraction of time interacting with kinesin-II motors, this could lead to slow transport of ChAT. Alternatively, if the overall balance of anterograde versus retrograde transport of ChAT-containing particles is small, then net transport rates of ChAT would be slow. Finally, we note that if either of these parameters can be regulated by the axon, then the actual rate of ChAT transport might also vary in different cell types or circumstances, thus accounting for the diversity in rates reported. In this context, recent work demonstrating that kinesin-I may have a role in movement of intermediate filament subunits in nonneural cells is consistent with this general view (Prahlad et al. 1998). At present the question of how slow transport is driven remains open, but the possibility that kinesin-II motors might play a role warrants further investigation.
Acknowledgments
We thank S. Chen for isolating the cDNA clones and sequencing, and Joe Marszalek for providing the KIF3A cDNA. We are grateful to P. Salvaterra, J.T. Littleton, and K. Zinsmaier for providing various antisera.
S. Perez was supported by a National Science Foundation postdoctoral fellowship. This project was supported by a grant from the National Institutes of Health to L.S.B. Goldstein. L.S.B. Goldstein and H. Steller are investigators of the Howard Hughes Medical Institute.
Footnotes
1.used in this paper: aa, amino acid(s); ChAT, choline acetyltransferase; CNS, central nervous system; CSP, cysteine string protein; EMS, ethyl methanesulphonate; KHC, kinesin heavy chain; nAChR, nicotinic acetylcholine receptor(s); PNS, peripheral nervous system; SYT, synaptotagmin
S. Perez's present address is Division of Biology, California Institute of Technology, Pasadena, CA 91125.
References
- Ashburner M. Drosophila 1989. Cold Spring Harbor Laboratory; Cold Spring Harbor, NY: pp. 1331 [Google Scholar]
- Beech P.L., Pagh-Roehl K., Noda Y., Hirokawa N., Burnside B., Rosenbaum J.L. Localization of kinesin superfamily proteins to the connecting cilium of fish photoreceptors. J. Cell Sci. 1996;109:889–897. doi: 10.1242/jcs.109.4.889. [DOI] [PubMed] [Google Scholar]
- Brand A.H., Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. doi: 10.1242/dev.118.2.401. [DOI] [PubMed] [Google Scholar]
- Broadie K.S., Bate M. Development of the embryonic neuromuscular synapse of Drosophila melanogaster . J. Neurosci. 1993;13:144–166. doi: 10.1523/JNEUROSCI.13-01-00144.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown N.H., Kafatos F.C. Functional cDNA libraries from Drosophila embryos. J. Mol. Biol. 1988;203:425–437. doi: 10.1016/0022-2836(88)90010-1. [DOI] [PubMed] [Google Scholar]
- Campos-Ortega J.A., Hartenstein V. The embryonic development of Drosophila melanogaster 2nd ed 1997. Springer-Verlag New York, Inc; New York: pp. 405 pp [Google Scholar]
- Cole D.G., Chinn S.W., Wedaman K.P., Hall K., Vuong T., Scholey J.M. Novel heterotrimeric kinesin-related protein purified from sea urchin eggs. Nature. 1993;366:268–270. doi: 10.1038/366268a0. [DOI] [PubMed] [Google Scholar]
- Gindhart J.G., Jr., Desai C.J., Beushausen S., Zinn K., Goldstein L.S. Kinesin light chains are essential for axonal transport in Drosophila . J. Cell Biol. 1998;141:443–454. doi: 10.1083/jcb.141.2.443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstein L.S.B., Fyrberg E.A. Drosophila melanogaster: practical uses in cell and molecular biology 1994. Academic Press; San Diego, CA: pp. 755 [Google Scholar]
- Goldstein L.S.B., Philp A.V. The road less travelledemerging principles of kinesin motor utilization. Annu. Rev. Cell Dev. Biol. 1999;In press doi: 10.1146/annurev.cellbio.15.1.141. [DOI] [PubMed] [Google Scholar]
- Goldstein L.S.B., Yang Z. Microtubule-based transport systems in neuronsthe roles of kinesins and dyneins. Annu. Rev. Neurosci. 2000;23:39–72. doi: 10.1146/annurev.neuro.23.1.39. [DOI] [PubMed] [Google Scholar]
- Goodman C.S., Doe C.Q. Embryonic development of the Drosophila central nervous system. In: Bate M., Martinez Arias A., editors. The Development of Drosophila melanogaster. Cold Spring Harbor Laboratory Press; Plainview, NY: 1993. pp. 1131–1206. [Google Scholar]
- Grigliatti T. Mutagenesis. In: Roberts D.B., editor. Drosophila, a Practical Approach. IRL Press; Oxford, UK: 1986. pp. 39–58. [Google Scholar]
- Hall D.H., Hedgecock E.M. Kinesin-related gene unc-104 is required for axonal transport of synaptic vesicles in C. elegans . Cell. 1991;65:837–847. doi: 10.1016/0092-8674(91)90391-b. [DOI] [PubMed] [Google Scholar]
- Harlow E., Lane D. Antibodiesa Laboratory Manual 1988. Cold Spring Harbor Laboratory; Cold Spring Harbor, NY: pp. 726 [Google Scholar]
- Henson J.H., Cole D.G., Terasaki M., Rashid D., Scholey J.M. Immunolocalization of the heterotrimeric kinesin-related protein KRP(85/95) in the mitotic apparatus of sea urchin embryos. Dev. Biol. 1995;171:182–194. doi: 10.1006/dbio.1995.1270. [DOI] [PubMed] [Google Scholar]
- Henson J.H., Cole D.G., Roesener C.D., Capuano S., Mendola R.J., Scholey J.M. The heterotrimeric motor protein kinesin-II localizes to the midpiece and flagellum of sea urchin and sand dollar sperm. Cell Motil. Cytoskelet. 1997;38:29–37. doi: 10.1002/(SICI)1097-0169(1997)38:1<29::AID-CM4>3.0.CO;2-C. [DOI] [PubMed] [Google Scholar]
- Hirokawa N. Kinesin and dynein superfamily proteins and the mechanism of organelle transport. Science. 1998;279:519–526. doi: 10.1126/science.279.5350.519. [DOI] [PubMed] [Google Scholar]
- Hurd D.D., Saxton W.M. Kinesin mutations cause motor neuron disease phenotypes by disrupting fast axonal transport in Drosophila . Genetics. 1996;144:1075–1085. doi: 10.1093/genetics/144.3.1075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jan L.Y., Jan Y.N. Antibodies to horseradish peroxidase as specific neuronal markers in Drosophila and in grasshopper embryos. Proc. Natl. Acad. Sci. USA. 1982;79:2700–2704. doi: 10.1073/pnas.79.8.2700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johansen J., Halpern M.E., Johansen K.M., Keshishian H. Stereotypic morphology of glutamatergic synapses on identified muscle cells of Drosophila larvae. J. Neurosci. 1989;9:710–725. doi: 10.1523/JNEUROSCI.09-02-00710.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kitamoto T., Ikeda K., Salvaterra P.M. Regulation of choline acetyltransferase/lacZ fusion gene expression in putative cholinergic neurons of Drosophila melanogaster . J. Neurobiol. 1995;28:70–81. doi: 10.1002/neu.480280107. [DOI] [PubMed] [Google Scholar]
- Kondo S., Sato-Yoshitake R., Noda Y., Aizawa H., Nakata T., Matsuura Y., Hirokawa N. KIF3A is a new microtubule-based anterograde motor in the nerve axon. J. Cell Biol. 1994;125:1095–1107. doi: 10.1083/jcb.125.5.1095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozminski K.G., Johnson K.A., Forscher P., Rosenbaum J.L. A motility in the eukaryotic flagellum unrelated to flagellar beating. Proc. Natl. Acad. Sci. USA. 1993;90:5519–5523. doi: 10.1073/pnas.90.12.5519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozminski K.G., Beech P.L., Rosenbaum J.L. The Chlamydomonas kinesin-like protein FLA10 is involved in motility associated with the flagellar membrane. J. Cell Biol. 1995;131:1517–1527. doi: 10.1083/jcb.131.6.1517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le Bot N., Antony C., White J., Karsenti E., Vernos I. Role of xklp3, a subunit of the xenopus kinesin II heterotrimeric complex, in membrane transport between the endoplasmic reticulum and the Golgi apparatus. J. Cell Biol. 1998;143:1559–1573. doi: 10.1083/jcb.143.6.1559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindsley D.L., Zimm G.G. The Genome of Drosophila melanogaster 1992. Academic Press; San Diego, CA: pp. 1133 [Google Scholar]
- Littleton J.T., Bellen H.J., Perin M.S. Expression of synaptotagmin in Drosophila reveals transport and localization of synaptic vesicles to the synapse. Development. 1993;118:1077–1088. doi: 10.1242/dev.118.4.1077. [DOI] [PubMed] [Google Scholar]
- Luo L., Liao Y.J., Jan L.Y., Jan Y.N. Distinct morphogenetic functions of similar small GTPasesDrosophila Drac 1 is involved in axonal outgrowth and myoblast fusion. Genes Dev. 1994;15:1787–1802. doi: 10.1101/gad.8.15.1787. [DOI] [PubMed] [Google Scholar]
- Malatova Z., Chavko M., Marsala J. Effect of spinal cord ischemia on axonal transport of cholinergic enzymes in rabbit sciatic nerve. Brain Res. 1989;481:31–38. doi: 10.1016/0006-8993(89)90482-4. [DOI] [PubMed] [Google Scholar]
- Marszalek J.R., Ruiz-Lozano P., Roberts E.A., Chien K.R., Goldstein L.S.B. Situs inversus and embryonic ciliary morphogenesis defects in mouse mutants lacking the KIF3A subunit of kinesin-II. Proc. Natl. Acad. Sci. USA. 1999;96:5043–5048. doi: 10.1073/pnas.96.9.5043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris R.L., Scholey J.M. Heterotrimeric kinesin-II is required for the assembly of motile 9+2 ciliary axonemes on sea urchin embryos. J. Cell Biol. 1997;138:1009–1022. doi: 10.1083/jcb.138.5.1009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muresan V., Abramson T., Lyass A., Winter D., Porro E., Hong F., Chamberlin N.L., Schnapp B.J. KIF3C and KIF3A form a novel neuronal heteromeric kinesin that associates with membrane vesicles. Mol. Biol. Cell. 1998;9:637–652. doi: 10.1091/mbc.9.3.637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nangaku M., Sato-Yoshitake R., Okada Y., Noda Y., Takemura R., Yamazaki H., Hirokawa N. KIF1B, a novel microtubule plus end-directed monomeric motor protein for transport of mitochondria. Cell. 1994;79:1209–1220. doi: 10.1016/0092-8674(94)90012-4. [DOI] [PubMed] [Google Scholar]
- Niclas J., Navone F., Hom-Booher N., Vale R.D. Cloning and localization of a conventional kinesin motor expressed exclusively in neurons. Neuron. 1994;12:1059–1072. doi: 10.1016/0896-6273(94)90314-x. [DOI] [PubMed] [Google Scholar]
- Nixon R.A. The slow axonal transport of cytoskeletal proteins. Curr. Opin. Cell Biol. 1998;10:87–92. doi: 10.1016/s0955-0674(98)80090-2. [DOI] [PubMed] [Google Scholar]
- Nonaka S., Tanaka Y., Okada Y., Takeda S., Harada A., Kanai Y., Kido M., Hirokawa N. Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell. 1998;95:829–837. doi: 10.1016/s0092-8674(00)81705-5. [DOI] [PubMed] [Google Scholar]
- Okada Y., Yamazaki H., Sekine-Aizawa Y., Hirokawa N. The neuron-specific kinesin superfamily protein KIF1A is a unique monomeric motor for anterograde axonal transport of synaptic vesicle precursors. Cell. 1995;81:769–780. doi: 10.1016/0092-8674(95)90538-3. [DOI] [PubMed] [Google Scholar]
- Orozco J.T., Wedaman K.P., Signor D., Brown H., Rose L., Scholey J.M. Movement of motor and cargo along cilia. Nature. 1999;398:674. doi: 10.1038/19448. [DOI] [PubMed] [Google Scholar]
- Otsuka A.J., Jeyaprakash A., Garcia-Anoveros J., Tang L.Z., Fisk G., Hartshorne T., Franco R., Born T. The C. elegans unc-104 gene encodes a putative kinesin heavy chain-like protein. Neuron. 1991;6:113–122. doi: 10.1016/0896-6273(91)90126-k. [DOI] [PubMed] [Google Scholar]
- Perez S.E., Steller H. Molecular and genetic analyses of lama, an evolutionarily conserved gene expressed in the precursors of the Drosophila first optic ganglion. Mech. Dev. 1996;59:11–27. doi: 10.1016/0925-4773(96)00556-4. [DOI] [PubMed] [Google Scholar]
- Pesavento P.A., Stewart R.J., Goldstein L.S. Characterization of the KLP68D kinesin-like protein in Drosophilapossible roles in axonal transport. J. Cell Biol. 1994;127:1041–1048. doi: 10.1083/jcb.127.4.1041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piperno G., Mead K. Transport of a novel complex in the cytoplasmic matrix of Chlamydomonas flagella. Proc. Natl. Acad. Sci. USA. 1997;94:4457–4462. doi: 10.1073/pnas.94.9.4457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piperno G., Mead K., Henderson S. Inner dynein arms but not outer dynein arms require the activity of kinesin homologue protein KHP1(FLA10) to reach the distal part of flagella in Chlamydomonas . J. Cell Biol. 1996;133:371–379. doi: 10.1083/jcb.133.2.371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prahlad V., Yoon M., Moir R.D., Vale R.D., Goldman R.D. Rapid movements of vimentin on microtubule trackskinesin-dependent assembly of intermediate filament networks. J. Cell Biol. 1998;143:159–170. doi: 10.1083/jcb.143.1.159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers S.L., Tint I.S., Fanapour P.C., Gelfand V.I. Regulated bidirectional motility of melanophore pigment granules along microtubules in vitro. Proc. Natl. Acad. Sci. USA. 1997;94:3720–3725. doi: 10.1073/pnas.94.8.3720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenbaum J.L., Cole D.G., Diener D.R. Intraflagellar transportthe eyes have it. J. Cell Biol. 1999;144:385–388. doi: 10.1083/jcb.144.3.385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sambrook J., Fritsch E.F., Maniatis T. Molecular Cloninga Laboratory Manual 2nd ed 1989. Cold Spring Harbor Laboratory; Cold Spring Harbor, NY: pp. 1659 [Google Scholar]
- Schloss P., Mayser W., Gundelfinger E.D., Betz H. Cross-linking of 125I-alpha-bungarotoxin to Drosophila head membranes identifies a 42 kDa toxin binding polypeptide. Neurosci. Lett. 1992;145:63–66. doi: 10.1016/0304-3940(92)90204-k. [DOI] [PubMed] [Google Scholar]
- Scholey J.M. Kinesin-II, a membrane traffic motor in axons, axonemes, and spindles. J. Cell Biol. 1996;133:1–4. doi: 10.1083/jcb.133.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shakir M.A., Fukushige T., Yasuda H., Miwa J., Siddiqui S.S. C. elegans osm-3 gene mediating osmotic avoidance behaviour encodes a kinesin-like protein. Neuroreport. 1993;4:891–894. doi: 10.1097/00001756-199307000-00013. [DOI] [PubMed] [Google Scholar]
- Signor D., Wedaman K.P., Rose L.S., Scholey J.M. Two heteromeric kinesin complexes in chemosensory neurons and sensory cilia of Caenorhabditis elegans . Mol. Biol. Cell. 1999;10:345–360. doi: 10.1091/mbc.10.2.345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stewart R.J., Pesavento P.A., Woerpel D.N., Goldstein L.S. Identification and partial characterization of six members of the kinesin superfamily in Drosophila . Proc. Natl. Acad. Sci. USA. 1991;88:8470–8474. doi: 10.1073/pnas.88.19.8470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tabish M., Siddiqui Z.K., Nishikawa K., Siddiqui S.S. Exclusive expression of C. elegans osm-3 kinesin gene in chemosensory neurons open to the external environment. J. Mol. Biol. 1995;247:377–389. doi: 10.1006/jmbi.1994.0146. [DOI] [PubMed] [Google Scholar]
- Takeda S., Yonekawa Y., Tanaka Y., Okada Y., Nonaka S., Hirokawa N. Left-right asymmetry and kinesin superfamily protein KIF3Anew insights in determination of laterality and mesoderm induction by kif3A−/− mice analysis. J. Cell Biol. 1999;145:825–836. doi: 10.1083/jcb.145.4.825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tandon A., Bachoo M., Weldon P., Polosa C., Collier B. Effects of colchicine application to preganglionic axons on choline acetyltransferase activity and acetylcholine content and release in the superior cervical ganglion. J. Neurochem. 1996;66:1033–1041. doi: 10.1046/j.1471-4159.1996.66031033.x. [DOI] [PubMed] [Google Scholar]
- Tautz D., Pfeifle C. A non-radioactive in situ hybridization method for the localization of specific RNAs in Drosophila embryos reveals translational control of the segmentation gene hunchback. Chromosoma. 1989;98:81–85. doi: 10.1007/BF00291041. [DOI] [PubMed] [Google Scholar]
- Tuma M.C., Zill A., Le Bot N., Vernos I., Gelfand V. Heterotrimeric kinesin II is the microtubule motor protein responsible for pigment dispersion in Xenopus melanophores. J. Cell Biol. 1998;143:1547–1558. doi: 10.1083/jcb.143.6.1547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vashishtha M., Walther Z., Hall J.L. The kinesin-homologous protein encoded by the Chlamydomonas FLA10 gene is associated with basal bodies and centrioles. J. Cell Sci. 1996;109:541–549. doi: 10.1242/jcs.109.3.541. [DOI] [PubMed] [Google Scholar]
- Walther Z., Vashishtha M., Hall J.L. The Chlamydomonas FLA10 gene encodes a novel kinesin-homologous protein. J. Cell Biol. 1994;126:175–188. doi: 10.1083/jcb.126.1.175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamazaki H., Nakata T., Okada Y., Hirokawa N. KIF3A/Ba heterodimeric kinesin superfamily protein that works as a microtubule plus end-directed motor for membrane organelle transport. J. Cell Biol. 1995;130:1387–1399. doi: 10.1083/jcb.130.6.1387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Z., Goldstein L.S. Characterization of the KIF3C neural kinesin-like motor from mouse. Mol. Biol. Cell. 1998;9:249–261. doi: 10.1091/mbc.9.2.249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Z., Hanlon D.W., Marszalek J.R., Goldstein L.S. Identification, partial characterization, and genetic mapping of kinesin-like protein genes in mouse. Genomics. 1997;45:123–131. doi: 10.1006/geno.1997.4901. [DOI] [PubMed] [Google Scholar]
- Yasuyama K., Kitamoto T., Salvaterra P.M. Localization of choline acetyltransferase-expressing neurons in the larval visual system of Drosophila melanogaster . Cell Tissue Res. 1995;282:193–202. doi: 10.1007/BF00319111. [DOI] [PubMed] [Google Scholar]
- Yonekawa Y., Harada A., Okada Y., Funakoshi T., Kanai Y., Takei Y., Terada S., Noda T., Hirokawa N. Defect in synaptic vesicle precursor transport and neuronal cell death in KIF1A motor protein–deficient mice. J. Cell Biol. 1998;141:431–441. doi: 10.1083/jcb.141.2.431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zador E. Expression of the acetylcholinesterase transcript in the chordotonal neurons of Drosophila embryos. Biochem. Genet. 1995;33:41–49. doi: 10.1007/BF00554557. [DOI] [PubMed] [Google Scholar]
- Zinsmaier K.E., Eberle K.K., Buchner E., Walter N., Benzer S. Paralysis and early death in cysteine string protein mutants of Drosophila . Science. 1994;263:977–980. doi: 10.1126/science.8310297. [DOI] [PubMed] [Google Scholar]