

Abstract
Pointing has long been considered to be a uniquely human, universal, and biologically based gesture. However, pointing emerges spontaneously, without explicit training, in captive chimpanzees. Because pointing is commonplace in captive chimpanzees and virtually absent in wild chimpanzees, and because both captive and wild chimpanzees are sampled from the same gene pool, pointing by captive apes is attributable to environmental influences on communicative development. If pointing by captive chimpanzees is so variably expressed in different rearing environments, this suggests that pointing by humans may also be attributable to situational factors that make pointing effective in certain developmental contexts.
Keywords: pointing, chimpanzees, communication, epigenesis
A defining characteristic of the human species is our capacity to rapidly establish topics for mutual contemplation. For example, we can discuss the health of the cat that lies on the rug in front of the fireplace in the room with us. The identification and location of entities for mutual contemplation is also possible without language. By one year of life, children begin to point to events and objects in their environments. Points do not stand for or represent the objects indicated in the way that words do. The ‘‘standing for’’ relation is termed by linguists reference; in verbal reference, words that represent particular entities are not iconic—e.g., the relationship between the physical features of the word dog and actual dogs is arbitrary. In contrast, with pointing, the relationship between the signal and the thing indicated is not arbitrary; a point’s specific meaning is determined in large part by the spatial locations of the pointer, the thing indicated, and the communicative partner. An act of pointing, thus, creates a referential triangle that incorporates distant objects into the relationship between a signaler and the recipient of the gesture (e.g., Butterworth, 2003).
The capacity to create this referential triangle is widely believed to be foundational for humans’ acquisition of speech during infancy. Butterworth (2003) referred to pointing as the ‘‘royal road to language’’ (p. 9) because when a child looks in the direction of a point at the same time that a mature speaker utters the name of the entity indicated, the link between the word and the entity for which it stands is facilitated through an auditory–visual learned association. For this and other reasons, developmental psychologists have long claimed that pointing, like speech, is a human species-specific adaptation for reference.
One line of argument in support of this hypothesis has been the widespread but incorrect claim that apes do not point (Povinelli, Bering, & Giambrone, 2003). Experimental work in our laboratory (Leavens & Hopkins, 1998; Leavens, Hopkins, & Bard, 1996; Leavens, Hopkins, & Thomas, 2004; reviewed by Leavens, Russell, & Hopkins, 2005) demonstrates that chimpanzees in captivity commonly point to unreachable food. Between 41% and 71% of chimpanzees in our studies point to unreachable food, with sample sizes ranging from 29 to 115 subjects. Sometimes they point with their index fingers, though more usually chimpanzees in this population point with all fingers extended (pointing with the whole hand). Some researchers refer to this latter kind of pointing as ‘‘reaching,’’ but we know that these are communicative signals because chimpanzees will not reach towards obviously unreachable food if there is nobody around to see them do it (Fig. 1; see Leavens et al., 1996, 2004). Like human infants, chimpanzees in captivity point with no explicit training whatsoever; pointing emerges ‘‘spontaneously.’’ The behavior is not limited to chimpanzees; it has also been observed in the other great apes, including orangutans, gorillas, and bonobos (Leavens & Hopkins, 1999).
Fig. 1.
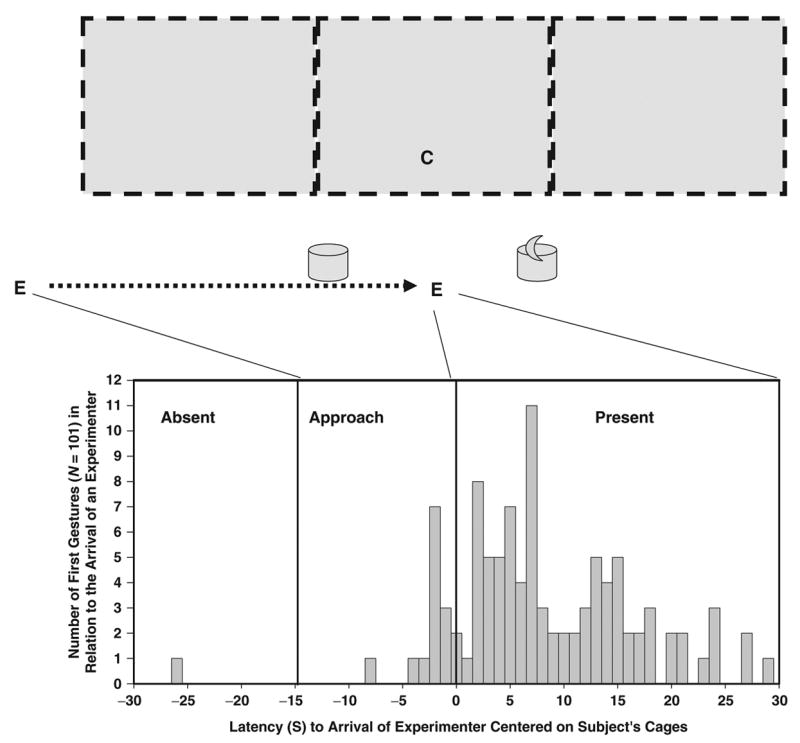
Experimental demonstration of the necessity of an audience (an experimenter, E) for a chimpanzee (C) to display a manual gesture in the presence of unreachable food. Each of 101 chimpanzees was tested twice, once with the food visible (the crescent shape represents a banana), once with it hidden under one of two inverted buckets in front of the cage. In each trial, the timing of the chimpanzee’s first manual gesture, if any, before or after the arrival of a previously absent experimenter in front of its cage (time ‘‘0’’ for reference) was recorded. Number of first gestures during the 1-minute window during which the experimenter was absent, approaching, and present are depicted in the graph at the bottom of the figure; data combine the results of both (food-visible, food-hidden) trials. Between the two trials, 62 chimpanzees exhibited 101 manual gestures, all but one of them when the experimenter was present or nearly so. (Data from Leavens, Hopkins, & Thomas, 2004.)
Strikingly, however, there are few reports of pointing by wild apes. As noted by Povinelli et al. (2003): ‘‘[D]espite intensive observations of the spontaneous interactions of free-ranging chimpanzees, there is no evidence that these animals . . . gesture with the arm and/or index finger toward other objects, animals, or events in space’’ (p. 41). Indeed, the only detailed description of pointing by wild apes is an observation of a bonobo pointing twice to human observers who were hiding in some shrubbery (Veà & Sabater-Pi, 1998). Thus, it seems that apes almost never point in their natural habitats. Because pointing is not part of the communicative repertoires of wild apes, and because wild apes are adapted by natural selection to survive in those wild habitats, there is now a widespread, but incorrect, perception that pointing by captive apes is scientifically uninformative, because findings from captive apes allegedly suffer from a lack of ecological validity.
ECOLOGICAL VALIDITY
Ecological validity is the degree to which laboratory conditions accurately capture relevant features of the natural environment for any given organism. With reference to gesturing, the argument goes like this: Chimpanzees in the wild exhibit their full behavioral repertoires in the environments for which they are adapted through natural selection. In other words, wild apes in their natural habitats manifest the evolutionary interplay between their adaptive milieux and their genetic endowment. Taking chimpanzees out of these environments and raising them in the impoverished circumstances typical of captivity removes the essential feedback from their natural environments, leaving the researcher with a wholly artificial sample of apes whose behavior is unrepresentative of their wild counterparts. Thus, if chimpanzees in captivity point, and wild chimpanzees do not, this fact is taken to be an uninteresting artifact of the bizarre and adaptively irrelevant circumstances of captivity.
The fallacy in this line of reasoning can be clarified with reference to two considerations. First, captive chimpanzees, like all other organisms, exist within an ecological framework. When they point to request food, this constitutes a foraging strategy that emerges frequently in captivity: Chimpanzees in captivity face a foraging problem in which they cannot acquire desirable food except through manipulation of a human. This referential problem, in which a chimpanzee is dependent on another individual to retrieve food that is distant from both of them, is not found in the wild (Leavens, 2004). Captive apes exhibit a suite of communicative behaviors in one ecological context and wild apes exhibit different suites of communicative behaviors in other ecological contexts; that is, there is a behavioral ecology of communication in chimpanzees. (It is worth noting that studies of communication by modern humans in industrialized countries also suffer from the same lack of ecological validity; it cannot be said that people are adapted to contemporary, urban environments by dint of our ancestors’ differential reproduction and survival in such environments, which are historically novel.) Therefore, although studies of captive apes may suffer from a lack of ecological validity, such apes nevertheless constitute ethologically valid populations insofar as their communicative behaviors are influenced by their environments throughout development.
A second consideration is that captive apes are sampled from the same gene pool as wild apes. In the last century, hunters often shot ape mothers to capture their infants for the biomedical-research, zoo, exotic-pet, and entertainment industries. It is implausible that these hunters somehow managed to bring into captivity only those rare chimpanzees with the ‘‘pointing gene’’ or ‘‘pointing-gene complex.’’ The communicative behavior of captive chimpanzees (who frequently point) is, therefore, predicated on the same genome as the communicative behavior of wild chimpanzees (who rarely, if ever, point; Leavens, 2004). The difference in the communicative repertoires of wild and captive chimpanzees indicates that these apes will develop pointing when they are faced with the appropriate environmental circumstances; that is, the data permit one to generalize the capacity for pointing to the entire species. Pointing in our nearest living relatives, the chimpanzees, is thus attributable to epigenetic processes: Pointing emerges as a consequence of ecological factors that come into play in some developmental contexts (captivity) but not in others (wild habitats).
EPIGENESIS
Pointing by apes is epigenetic because it does not rely on pre-occurring changes in the genome; novel phenotypes (the observable anatomy, functions, and behaviors of an organism) emerge in certain developmental contexts but not in others, and therefore the most straightforward interpretation of pointing by captive apes is that their communicative repertoires are malleable in the face of environmental variation (Bard, 1998). This flexibility is further underscored by consideration of the apparent group differences within captive ape populations in the specific hand posture used: Chimpanzees raised in close association with humans, such as home-raised or language-trained chimpanzees, point overwhelmingly with their index fingers, whereas chimpanzees housed with more emotionally detached relationships with humans, as in biomedical research centers, point more frequently with all fingers extended (Leavens & Hopkins, 1999). Thus, the extent to which chimpanzees exhibit a preference for pointing with their index fingers depends on the extent to which they experience very close relationships with humans. In chimpanzees, not only is the propensity to point subject to epigenetic processes, so is the specific pointing posture of the hand.
Humans and chimpanzees have a close evolutionary relationship (the two species had a common ancestor as recently as 6 or 7 million years ago). In light of this, if both the propensity to point and the form of pointing is so contingent on environmental factors in chimpanzees, one is authorized to ask whether pointing by humans truly is the ‘‘biologically based and species-specific’’ (Butterworth, 2003, p. 28) gesture that many claim it to be. And since pointing by apes is facilitated in some, but not all, environmental circumstances, it is reasonable to suggest that pointing by humans might also be facilitated in some, but not other, environmental contexts. Recent cross-cultural work suggests that this is, indeed, the case.
Both the propensity to point and the specific form of pointing are, in fact, quite variable across human cultures (Wilkins, 2003). In Western cultures, for example, people generally do not point with their lips, but pointing with the lips coexists with finger pointing in the repertoires of many people (e.g., the Ar-rernte of Australia, the Ewe of Ghana). And although pointing with the index finger is widespread among humans, it is not universal: It is reportedly nonexistent in one culture, the Barai of Papua New Guinea (Olson, personal communication in Wilkins, 2003). People in many cultures also frequently point with their whole hands, rather than their index fingers, in some contexts. That pointing is variable in both incidence and posture between representatives of different cultures, as well as within cultures, suggests that, like chimpanzees, humans exhibit flexibility in their propensity to point. If manual pointing is not a human universal, then claims for its species-specificity or ‘‘innateness’’ are refuted (Wilkins, 2003).
As with humans, there are numerous examples of group differences in the behavior of wild apes (e.g., wild chimpanzees vary the size of a tool and method of extraction of ants, both across different sites and within a single site, in accordance with the type of ant and the specific location of the ant-dipping; Humle & Matsuzawa, 2002). Thus, wild apes adapt their foraging strategies to the local exigencies of their foraging problem spaces (a problem space being the set of related circumstances in which barriers exist to direct attainment of goals, in which the application of indirect means to attain those goals is required). If we are correct, the same is true of captive apes, insofar as pointing constitutes a foraging strategy.
THE REFERENTIAL PROBLEM SPACE
Gottlieb (1992) emphasized that our phenotypic repertoires are underspecified by our genes—that is, there are vastly fewer genes than possible phenotypes. Indeed, in the last 10 years, estimates of the number of human genes have been revised downwards from over 100,000 to fewer than 20,000 genes. Because complex phenotypic traits like communicative behaviors are so underspecified by our genomes, human behavioral repertoires can be seen as the product of environmental feedback on developmental processes. That different groups of chimpanzees sampled from the same gene pool differ in their propensities to point tells us that pointing emerges in our nearest living relatives when, and only when, the environment provides a function for that gesture. Furthermore, the differences in pointing morphology between different groups of captive chimpanzees suggest that chimpanzees’ tendencies to adopt the communicative signals of their caregivers (i.e., the extent to which they are motivated to imitate their caregivers) depend on the amount and quality of their experience interacting with humans (Bard, 1998).
Because both the propensity to point and the morphology of pointing in humans is also variable, the study of chimpanzees’ communicative repertoires in captivity may implicate specific environmental features that are relevant to understanding the development of pointing in humans. There are parallels in the circumstances in which captive apes and human children begin to use the pointing gesture (Fig. 2). First, captive apes experience physical barriers to directly obtaining desirable but unreachable food. Human infants also experience barriers, either through being physically restrained, for example in high chairs, or through endogenous limitations on their locomotor capacities. Secondly, both captive apes and human children experience histories of daily food provisioning by adult human caretakers. Pointing emerges, then, in circumstances in which the establishment of a referential triangle becomes a viable solution to a particular kind of problem. We propose that these factors may play a role in the development of pointing to request things in both humans and chimpanzees (e.g., Leavens, 2004; Leavens et al., 1996, 2004).
Fig. 2.
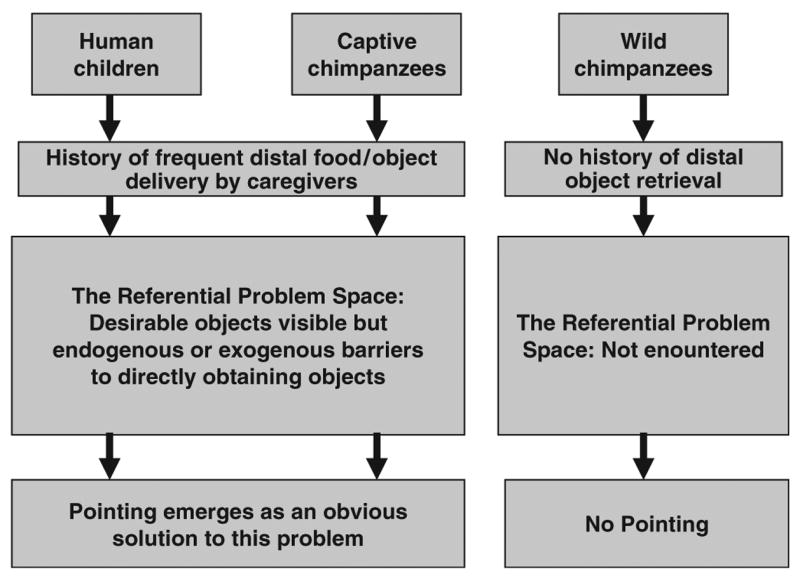
The referential problem space. A problem space is a set of related circumstances in which barriers exist to direct attainment of goals and therefore indirect means to attain those goals must be discovered or recalled and then effectively applied. Human children and captive chimpanzees experience both long histories of delivery of distal food and other objects by their human caregivers and, very frequently, barriers to directly obtaining those objects for themselves. Faced with a familiar care-giver and unreachable but desirable items, pointing emerges as an obvious solution to a particular problem space that is not encountered (or only rarely encountered) by wild chimpanzees.
The fact that chimpanzees in captivity frequently point but those in the wild almost never do is not merely a trivial consequence of raising chimpanzees in bizarre, captive environments. Rather, it suggests that pointing by humans is written neither in our genes nor in our anatomy, but in the functional characteristics of our social and physical environments. The harder argument to make is that pointing emerges in similar circumstances in two very closely related species with similar body plans and hand anatomies, yet derives from completely unrelated psychological processes (as suggested by Povinelli et al., 2003). When chimpanzees in captivity point to unreachable food, the overt meaning is obvious. It would be unfortunate if we failed to grasp the implicit meaning of this gesture: Pointing is not uniquely specified by the human genome.
If this interpretation of epigenetic effects on pointing by chimpanzees is correct, then we should find micro-ecological effects in our own species similar to those in chimpanzees. For example, we predict that babies raised in cultures with relatively less physical restraint might exhibit reduced propensities in middle infancy to manipulate their social partners through manual gesture; such children would experience fewer instances of the referential problem space. In terms of language evolution, the fact that pointing emerges so easily in chimpanzees suggests that this capacity for explicit reference was present in the common ancestor of humans and chimpanzees. Both relatively immobile human babies and chimpanzees housed in cages use pointing as a tactical solution to a particular kind of problem: the referential problem. No other primate physically restrains its offspring to the degree that humans do, and no other primate experiences such a prolonged epoch of locomotor immaturity; these factors put our offspring into the referential problem space. That pointing emerges in this problem space may, therefore, reflect not the influence of our unique adaptation for symbolic reference, but the causative influence of this problem space on our species’ evolutionary and developmental trajectories into referential communication.
Recommended Reading
de Waal, F.B.M. (2001, January 19). Pointing primates: Sharing knowledge . . . without language. Chronicle of Higher Education, B7–B9.
Leavens, D.A. (2004). (See References)
Leavens, D.A., Russell, J.L., & Hopkins, W.D. (2005). (See References)
Acknowledgments
We thank the late George Butterworth and Mark A. Krause for discussions of some of the issues raised in this manuscript.
References
- Bard KA. Social-experiential contributions to imitation and emotion in chimpanzees. In: Bråten S, editor. Intersubjective communication and emotion in early ontogeny: A source book. Cambridge, UK: Cambridge University Press; 1998. pp. 208–227. [Google Scholar]
- Butterworth G. Pointing is the royal road to language for babies. In: Kita S, editor. Pointing: Where language, culture, and cognition meet. Mahwah, NJ: Erlbaum; 2003. pp. 9–33. [Google Scholar]
- Gottlieb G. Individual development & evolution: The genesis of novel behavior. New York: Oxford University Press; 1992. [Google Scholar]
- Humle T, Matsuzawa T. Ant dipping among the chimpanzees of Boussou, Guinea, and some comparisons with other sites. American Journal of Primatology. 2002;58:133–148. doi: 10.1002/ajp.10055. [DOI] [PubMed] [Google Scholar]
- Leavens DA. Manual deixis in apes and humans. Interaction Studies. 2004;5:387–408. [Google Scholar]
- Leavens DA, Hopkins WD. Intentional communication by chimpanzees: A cross-sectional study of the use of referential gestures. Developmental Psychology. 1998;34:813–822. doi: 10.1037//0012-1649.34.5.813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leavens DA, Hopkins WD. The whole hand point: The structure and function of pointing from a comparative perspective. Journal of Comparative Psychology. 1999;113:417–425. doi: 10.1037/0735-7036.113.4.417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leavens DA, Hopkins WD, Bard KA. Indexical and referential pointing in chimpanzees (Pan troglodytes) Journal of Comparative Psychology. 1996;110:346–353. doi: 10.1037/0735-7036.110.4.346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leavens DA, Hopkins WD, Thomas RK. Referential communication by chimpanzees (Pan troglodytes) Journal of Comparative Psychology. 2004;118:48–57. doi: 10.1037/0735-7036.118.1.48. [DOI] [PubMed] [Google Scholar]
- Leavens DA, Russell JL, Hopkins WD. Intentionality as measured in the persistence and elaboration of communication by chimpanzees (Pan troglodytes) Child Development. 2005;76:291–306. doi: 10.1111/j.1467-8624.2005.00845.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Povinelli D, Bering J, Giambrone S. Chimpanzee ‘pointing’: Another error of the argument by analogy? In: Kita S, editor. Pointing: Where language, culture, and cognition meet. Hillsdale, NJ: Erlbaum; 2003. pp. 35–68. [Google Scholar]
- Veà JJ, Sabater-Pi J. Spontaneous pointing behaviour in the wild pygmy chimpanzee (Pan paniscus) Folia Primatologica. 1998;69:289–290. doi: 10.1159/000021640. [DOI] [PubMed] [Google Scholar]
- Wilkins D. Why pointing with the index finger is not a universal (in sociocultural and semiotic terms) In: Kita S, editor. Pointing: Where language, culture, and cognition meet. Hillsdale, NJ: Erlbaum; 2003. pp. 171–215. [Google Scholar]