

Abstract
The present study used event-related potentials (ERPs) to examine the time-course of visual word recognition using a masked repetition priming paradigm. In two experiments participants monitored a stream of words for occasional animal names, and ERPs were recorded to non-animal critical target items that were either repetitions or were unrelated to the immediately preceding masked prime word. In Experiment 1 the onset interval between the prime and target (stimulus-onset-asynchrony – SOA) was manipulated across four levels (60, 180, 300 and 420 ms) and the duration of primes was held constant at 40 ms. In Experiment 2 the SOA between the prime and target was held constant at 60 ms and the prime duration was manipulated across four levels (10, 20 30 and 40 ms). Both manipulations were found to have distinct effects on the N250 and N400 ERP components. The results provide converging evidence that the N250 reflects processing at the level of form representations (orthography and phonology) while the N400 reflects processing at the level of meaning.
Keywords: Visual word processing, word recognition, N400, N250, masked priming
1. Introduction
Visual word recognition occurs rapidly and with little apparent mental effort in skilled readers (Rayner, 1998). For over two decades behavioral researchers have used the masked priming technique to help unravel the temporal dynamics of the mental processes underlying this skill (see Forster et al., 2003; Grainger & Jacobs, 1999; for reviews). Although there are several versions of the masked priming technique, the one typically used to study word processing involves the rapid sequential presentation of a series of visual stimuli: a forward mask (e.g., a series of hash marks #########) followed by a prime (usually a word), a backward mask (another row of hash marks) and finally a target stimulus, to which participants are asked to make a speeded judgment (e.g., lexical decision). Critical to the typical pattern of effects is that the prime be very briefly displayed (e.g., 50 ms) and that it be sandwiched between the presentation of the preceding forward and subsequent backward masking stimuli which together serve to make the prime imperceptible (i.e., participants are usually unaware of its presence). However, even with such brief exposure the prime can produce robust effects on subsequent processing as indicated by changes in reaction time and/or error rates to the target. The classic finding is that when the prime and target are the same word, responses to the target are facilitated compared to the baseline condition were the prime and target are different unrelated words. This “repetition priming” effect, which has been widely interpreted as indicating that processing of the masked prime interacts with the stream of processes involved in recognizing the target, has proven to be one of the most useful techniques for helping researchers study visual word recognition.
Recently a number of studies have used the masked repetition priming paradigm in combination with the recording of event-related potentials (ERPs – Schnyer et al., 1997; Misra & Holcomb, 2003; Holcomb et al., 2005; Holcomb & Grainger, 2006; Kiyonaga et al., in press; Grainger, Kiyonaga & Holcomb, 2006; Petit et al., 2006; Eddy et al., 2006). In one such study, Holcomb and Grainger (2006) found that the N400 component was larger to target words following unrelated primes than primes that were identical to or nearly identical to the target. This pattern is similar to that found in a multitude of unmasked repetition and semantic priming studies (e.g., Bentin et al., 1985; Holcomb et al., 2005) and have been interpreted as reflecting processing of the semantic attributes of the stimuli.
Of most interest for the present study were the findings on two earlier components not previously reported in ERP priming studies (but see Grossi & Coch, 2005). The first of these components called the N/P150, peaked in the time frame of the exogenous N1, and was relatively focal effect being most notable over right occipital sites and took the form of a larger positivity to target words preceded by orthographically and lexically unrelated masked prime words compared to targets there were repetitions of their primes. Holcomb and Grainger attributed this earliest of repetition effects to the greater overlap of visual features in the repeated compared to the unrelated targets. The second effect was a more widely but somewhat frontally distributed ERP negativity that started around 170ms and peaked near 250 ms. Holcomb and Grainger referred to this as the N250 component, and argued that it was sensitive to the degree of orthographic overlap between the prime and target stimuli, as it proved sensitive to the number of letters that the prime and target had in common, being largest when primes and targets shared no letters, intermediate when prime and target shared several letters (partial repetition) and smallest when they shared all letters (full repetition). Collectively, these findings demonstrate that ERPs are highly sensitive to the time-course of early processing in the visual word recognition system and suggests that the combination of the masked priming technique with the recording of ERPs may be very useful for examining a host of issues that are difficult to address with reaction time alone. The general picture that arises from this work so far (see Grainger & Holcomb, in press, for review) is that a series of ERP components, the N/P150, N250, and N400, reflect a cascade of processes triggered on presentation of a printed word and primarily involving feature representations (N/P150), form representations (N250), and meaning (N400). Although Holcomb and Grainger (2006) and Grainger and Holcomb (in press) provided a more detailed analysis of the mapping of ERP components onto the process of visual word recognition, the above description summarizes the essential ingredients of that analysis for the purposes of the present study.
Interestingly however, several studies prior to Holcomb and Grainger that also used ERPs in the masked repetition priming paradigm did not find evidence for repetition effects earlier than the N400 (Misra & Holcomb, 2003; Holcomb et al., 2005). The biggest difference between these earlier studies and the Holcomb and Grainger experiment was the duration of the interval between the prime and target events. In Holcomb and Grainger (2006), and most of the work done by behavioral researchers interested in word recognition, the prime-target interval (stimulus-onset-asynchrony -- SOA) has been relatively short. In Holcomb and Grainger (2006) it was 70 ms (50 ms duration of the prime and a 20 ms backward mask with the target immediately replacing the backward mask). In earlier ERP masking studies typical prime-target SOAs have been between 500 and 1000 ms. These longer SOAs have been used to avoid problems with the overlap of ERPs that occurs when multiple events are presented in a short period of time. Holcomb and Grainger suggested that many of the early ERP effects that proved sensitive to visual word processing in their study might have been refractory at the longer SOA used in the Misra and Holcomb (2003) and Holcomb et al (2005) studies. This seems reasonable given that at least one behavioral study has also found that RT is sensitive to word processing manipulations in the masked priming paradigm, but only when the SOA is shorter than 500 ms (Ferrand, 1996).
Experiment 1 of the current study is a follow-up to the Holcomb and Grainger experiment and the earlier long interval masked priming studies (Misra & Holcomb, 2003; Holcomb et al., 2005). Here we explicitly manipulated the interval between primes and targets in the masked priming paradigm between the values used in these previous studies (60, 180, 300 and 420 ms). If prior studies failed to find early onset ERP masked repetition priming effects due to the relatively long SOAs employed, then it seems reasonable to predict that earlier effects should emerge at some point along the interval of SOAs used in the earlier studies (e.g., 500 ms in Holcomb et al., 2005) and the more recent Holcomb and Grainger study (70 ms).
In Experiment 2 the SOA was held constant at 60 ms and the duration of the masked primes was varied between 10 and 40 ms. Prior work has shown that the amplitude of the N400 repetition priming effect is sensitive to prime durations in the range of increases from 40 to 80 ms (Holcomb et al., 2005). What is not known is what happens when the prime duration is reduced below 40 ms. Furthermore, the prime duration manipulation of Experiment 2 allows a within-condition analysis of priming effects as proposed by Jacobs, Grainger, and Ferrand (1995). The main effect of prime duration (or prime intensity if prime duration is held constant) is analyzed for each prime condition separately, in order to examine how increased availability of information from the prime stimulus is modulating the influence of each type of prime on processing of target stimuli. Jacobs et al. (1995) demonstrated that the overall size of masked repetition priming effects found with word stimuli in the lexical decision task was due to a combination of slowing down of response times (RTs) in the different prime condition and faster RTs in the repetition prime condition. The same logic can be applied to the amplitude of ERP components. Although it is not yet possible to unambiguously interpret the direction of ERP effects in terms of underlying processes, concerning the N400 component, it is generally assumed that increased negativity reflects more processing effort (e.g., Holcomb, 1993). Therefore, the incremental priming technique can potentially allow us to distinguish between priming effects that are mainly driven by a release from the extra processing effort caused by an unrelated prime stimulus, and those that are mainly driven by a reduced effort in target processing following related primes. We hypothesize that the former type of priming will mostly likely occur in form-based processing (where inhibitory processing mechanisms are more prevalent), while the latter type of priming will mostly likely occur in semantic-based processing. Finally, the incremental analysis can be used as a further means of distinguishing different (overlapping) components, and therefore ensuring that they reflect activity in distinct sets of neural processors.
1.1 Experiment 1
In this experiment ERP masked repetition priming effects were measured in a variable SOA prime-target paradigm designed to decompose the temporal sequence of processing steps involved in visual word recognition. Participants were presented with brief (40 ms) visual primes that were masked by a both a forward and backward pattern mask and were followed by a fully visible target word (300 ms) that was a repetition of the prime (e.g., table – TABLE, mouth-MOUTH) or a target word that was completely unrelated to the prime (e.g., mouth-TABLE, table-MOUTH). The interval between the onset of the prime and target (SOA) was manipulated across four levels: 60 ms, 180 ms, 300 ms and 420 ms. Participants were instructed to read all stimuli and to press a button to occasional probe words in a particular semantic category (animals). The critical conditions in this experiment involved the non-probe words (i.e., those that did not require a behavioral response). There were two main predictions. The first was that shorter prime-target SOAs would result in pre-N400 priming effects including the N250. However, at longer prime-target intervals, pre-N400 effects would become progressively more attenuated. This prediction is based on behavioral work demonstrating that at SOAs longer than 300 ms lexical and sub-lexical mask priming effects are attenuated (Ferrand, 1996), but that at shorter SOAs of from 50 to 100 ms robust lexical effects can be obtained. A second prediction was that unlike the earlier components, the N400 would prove to be less sensitive to prime-target SOA and would produce robust priming effects across SOAs. This is based on the findings from long and short SOA studies all of which have reported clear N400s (Holcomb et al., 2005; Holcomb & Grainger, 2006; Kiyonaga at al., 2007).
1.2 Experiment 2
In a second experiment we measured ERP masked repetition priming effects in a variable prime duration paradigm. Here the goal was to extend our study of the temporal dynamics of visual word recognition by determining the minimal amount of prime processing necessary to reveal robust priming effects. Participants were presented with brief visual primes at each of four durations (10, 20, 30 and 40 ms) that were masked by a both a forward and backward pattern mask and were followed by a fully visible target word (300 ms) that was a repetition of the prime (e.g., table – TABLE, mouth-MOUTH) or a target word that was completely unrelated to the prime (e.g., mouth-TABLE, table-MOUTH). The interval between the onset of the prime and target was held constant at 60 ms, which is the same as the short SOA condition of Experiment 1 and is close to the SOA used by Holcomb and Grainger (2006). Participants were instructed to read all stimuli and to press a button to occasional probe words in a particular semantic category (animals). ERPs were time locked to the onset of primes and recorded for 700 ms after the onset of target words. This manipulation will allow us to determine the minimal prime duration (for the stimulus luminance values used in this experiment) that is necessary for obtaining repetition priming effects in the different ERP components. It also allows us to apply a within-condition analysis of the influence of prime duration on each prime condition separately (Jacobs et al., 1995).
2. Results
2.1 Experiment 1 Results
Visual Inspection of ERPs
The ERPs time locked to targets from 29 electrode sites for the two repetition conditions at each of the four prime-target SOAs are plotted in Figure 1a–d. Figure 2 presents blowups of the CP1 site from Figure 1 (panel a) and the difference waves calculated by subtracting repeated from unrelated target ERPs and over-plotted for all four prime-target SOAs (panel b). Plotted in Figures 3 and 4 are the voltage maps for the time periods surrounding the N250 and N400. As can be seen in these plots, ERPs in the target epoch varied substantially as a function of both the REPETITION and the prime-target SOA factors. Keep in mind that at the shorter SOAs (60 and 180 ms) the ERPs time locked to the target are a composite of neural activity generated by both the target and the immediately preceding prime and masking stimuli, while the ERPs time locked to the targets at the longer prime-target SOAs (300 and 420 ms) reflect activity evoked by the offset of the backward mask and the target item itself. Therefore, at least some of the SOA differences visible in Figures 1 and 2 are likely due to differential overlap of prime/mask and target ERPs. For this reason we will not focus on the main effects of SOA, which likely vary due to differences in prime-target ERP overlap. SOA effects will only be reported when they interact with the REPETITION factor.
Figure 1.
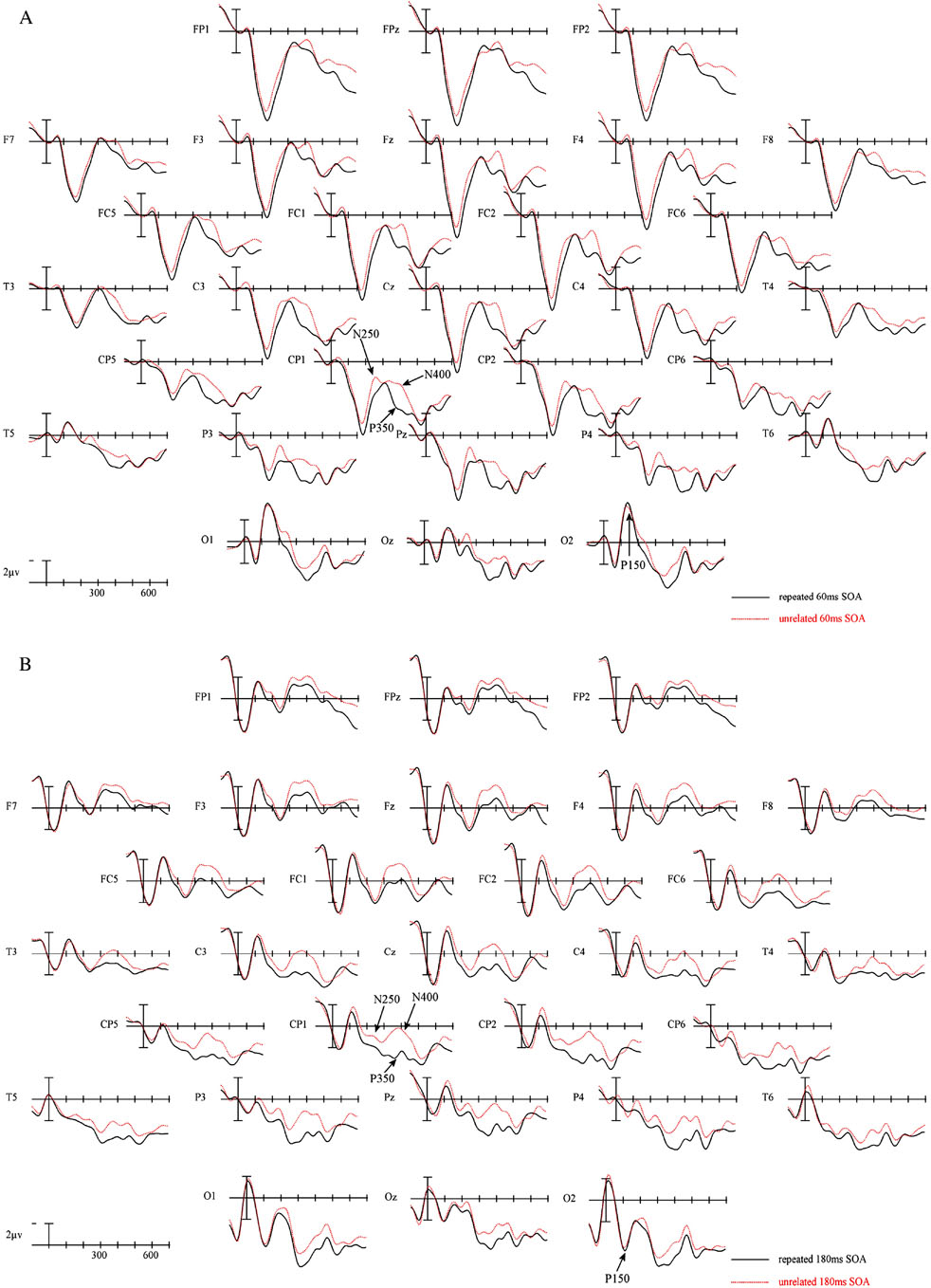
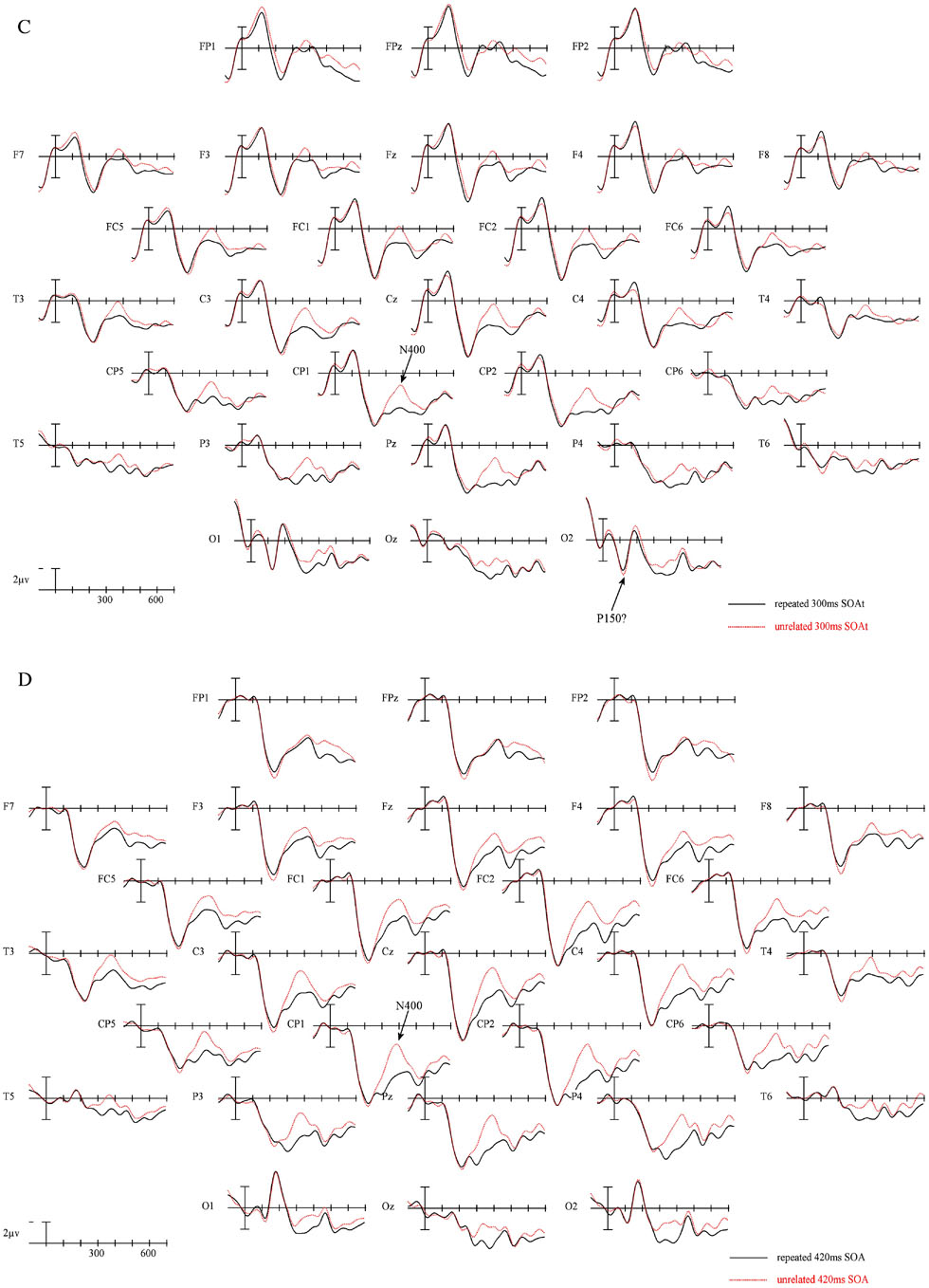
ERPs time locked to target onset in the two repetition conditions for the 60 ms SOA condition (a), 180 ms SOA condition (b) 300 ms SOA condition (C) and the 420 ms SOA condition (D). Negative voltages are plotted upward. Refer to Figure 12 for electrode locations.
Figure 2.

Panel a (top) Blowup of site CP1 from Figure 1 for all four SOAs. Panel b (bottom) Difference waves at CP1 calculated by subtracting Repeated from Unrelated Target ERPs for all four SOAs.
Figure 3.

Voltage maps calculated from difference waves (Unrelated-Repeated) for each of the four SOAs. Each row represents a different SOA with activity calculated at 50 ms intervals surrounding the N250 epoch.
Figure 4.

Voltage maps calculated from difference waves (Unrelated-Repeated) for each of the four SOAs. Each row represents a different SOA with activity calculated at 50 ms intervals surrounding the N400 epoch.
Analyses of ERP Data
N/P150 (125–175ms ms, Occipital Sites)
Not surprisingly there was a large main effect of SOA (F(3, 69) = 12.12, p < .001), but no effect of Repetition in this epoch.
N250 Epoch (150–300 ms)
Not surprisingly there was a large main effect of SOA across the head (midline: F(3, 69) = 15.20, p < .001; C1: F(3,69) = 11.92, p < .001; C2: F(3,69) = 9.32, p < .001; C3: F(3,69) = 7.05, p = .002). Most importantly, as can be seen in Figures 1, 2 and 3 this epoch produced clear signs of a REPETITION effect at the two shortest SOAs and little evidence of a similar effect at the two longest SOAs. This effect took the form of a widely distributed negativity larger for target words that were unrelated to the prior prime than those that were repeats. We confirmed this difference in priming as a function of SOA in two ways. First, in the omnibus ANOVA there was a linear trend in the SOA by REPETITION interaction (midline: F(1,23) = 4.25, p = .05; C1: F(1,23) = 4.02, p = .057; C2: F(1,23) = 3.67, p = .068; C3: F(1,23) = 3.45, p = .076) indicating that the SOAs differed in the size of the repetition priming effect. Second, in separate follow-up analyses at each SOA only the 60 and 180 ms SOA conditions produced significant REPETITION effects (main effect of REPETITION 60 ms SOA midline: F(1,23) = 8.37, p = .008; C1: F(1,23) = 7.07, p = .014; C2: F(1,23) = 5.65, p = .026; C3: F(1,23) = 6.05, p = .022; 180 ms midline: F(1,23) = 4.50, p = .045; C1: F(1,23) = 3.72, p = .066; C2: F(1,23) = 3.77, p = .064; C3: F(1,23) = 3.32, p = .082). There was little evidence of a REPETITION effect at the two longer SOAs in this epoch (all ps > .17).
N400 Epoch (400–550 ms)
As in the previous epoch Figures 1, 2 and 4 suggest that in the region of the peak of the N400 there were again robust widespread effects of SOA (midline: F(3,69) = 11.30, p < 001; C1: F(3,69) = 11.79, p < .001 ; C2: F(3,69) = 10.54, p < .001; C3: F(3,69) = 8.38, p < .001) and importantly, main effects of REPETITION (midline: F(1,23) = 12.05, p = 002; C1: F(1,23) = 13.96, p = .001 ; C2: F(1,23) = 16.09, p =.001; C3: F(1,23) = 18.33, p < .001). Unlike in the N250 epoch, the effects of REPETITION did not appear to vary as a function of SOA (REPETITION × SOA interaction, app ps > .8). This observation was also supported by significant REPETITION effects in each of the separate follow-up ANOVAs at each SOA (main effect of REPETITION 60 ms SOA, midline: F(1,23) = 5.96, p =< .023; C1: F(1,23) = 5.70, p = .026; C2: F(1,23) = 4.93, p = .037; C3: (1,23) = 6.08, p = .022; 180 ms SOA, midline: F(1,23) = 5.67, p =< .026; C1: F(1,23) = 6.28, p = .02; C2: F(1,23) = 7.91, p = .01; C3: (1,23) = 6.89, p = .015; 300 ms SOA, midline: F(1,23) = 3.83, p =< .063; C1: F(1,23) = 3.71, p < .067; C2: F(1,23) = 3.87, p = .061; C3: (1,23) = 5.43, p = .029; 420 ms SOA, midline: F(1,23) = 6.83, p =< .016; C1: F(1,23) = 10.95, p = .003; C2: F(1,23) = 10.74, p = .003; C3: (1,23) = 7.27, p = .013).
Time-course Analysis
We also performed fine grained time-course analyses on successive 50 ms epochs starting at target onset and continuing through the N400 epoch (450 ms). Reported in Table 1 are the main effects of REPETITION across nine electrode sites (F3, Fz, F4, C3, Cz, C4, P3, Pz, P4). As can be seen the effects of REPETITION had different time-courses in the different SOA conditions. At the two shortest SOAs repetition priming effects were significant in early phases of target processing (between 100 and 150 ms post-target onset), but at the two longest SOAs these priming effects emerged later during target processing.
Table 1.
Time-course Analysis Experiment 1 (SOA)
| 0–50 | 50–100 | 100–150 | 150–200 | 200–250 | 250–300 | 300–350 | 350–400 | 400–450 | 450–500 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 60 ms SOA | Ns | - | * | * | ** | - | ns | ** | ** | ns |
| 180 ms SOA | Ns | ns | - | * | - | - | ** | ** | ** | - |
| 300 ms SOA | Ns | ns | ns | - | ns | ns | * | ** | - | - |
| 420 ms SOA | Ns | ns | ns | ns | ns | ns | ** | ** | * | - |
- p < .1
p < .05
p < .01
Behavioral Data
Participants detected 88.6% (SD = 6) of animal probes in the target position and fewer than 18% (SD=17) in the prime position within a window of 200 to 800 ms post-probe onset. There was no effect of prime-target SOA on the number of probes detected.
2.2 Discussion of Experiment 1
Consistent with the behavioral results of Ferrand (1996), the earliest effects of masked repetition priming were only evident when the interval between the onset of the prime and target stimuli was less than 300 ms. At both the 60 and 180 ms SOAs there were robust effects of priming starting no later than 150 to 200 ms, while longer SOAs produced their first reliable effects of priming no earlier than 300–350 ms – during the classic N400 epoch.
Examination of Figures 1 and 2 suggests that the earliest priming effect in the 60 ms SOA condition occurred on the component identified by Holcomb Grainger (2006) as the N250. In the 180 ms SOA condition there was a similar effect in this same time frame. However, the morphology of the waveforms at the two SOAs do not appear to be the same (compare Figure 2 left two panels). One possible reason for these differences in wave shape could be due to the differential contribution of overlapping ERPs from the earlier prime and backward mask stimuli. In the 180 ms condition the N1 and P2 components from the prime and backward mask are likely overlapping with the early part of the target ERP. By comparison, in the 60 ms SOA condition, the overlap of prime/backward mask N1 and P2 with and target ERPs should be delayed by 120 ms. However, it is important to keep in mind that because the priming manipulation is the same across SOAs, any absolute differences in ERPs between unrelated and repeated targets is most likely due to the interaction of the amount of time that has elapsed between the prime and target items and the prime-target relationship (as opposed to differences in the prime or backward mask ERPs themselves).
Interestingly, there was no comparable effect to the early N250 visible in the ERPs to targets that followed their primes by 300 ms or 420 ms. In other words, when the time between prime and target onset is 300 ms or longer the process reflected by the N250 appears to be refractory. We will return to the importance of this observation in the General Discussion. However, consistent with previous masked repetition priming studies, a robust N400 repetition effect was present across the four SOAs (see Figures 2 to 4) used in the current experiment. In fact, there was little evidence that the prime target interval had a significant impact on the amplitude of this effect (the F-ratio for SOA by Repetition interaction was < 1). This strongly suggests that the processes reflected by this repetition effect are robust across a range of time intervals.
One curious omission for the current pattern of results is the lack of a significant effect at the earliest latencies reported by Holcomb and Grainger (2006). In their study the first repetition effect was a small focal occipital component which resulted in a more positive-going ERP in the Unrelated compared to Repeated targets and which peaked around 150 ms (N/P150). This region did not produce a similar significant difference in the current study.
2.3 Experiment 2 Results
Electrophysiological Data
Visual Inspection of ERPs
The ERPs time locked to targets from 29 electrode sites for the two repetition conditions at each of the four prime durations are plotted in Figure 5. Figure 6a presents blowups of the CP1 site from Figure 5 and Figure 6b presents differences waves over-plotted for all four prime durations at the CP1 site. Plotted in Figures 7 and 8 are the voltage maps for the time periods surrounding the N250 and N400. As can be seen in these plots, ERPs in the target epoch varied substantially as a function of both the REPETITION and the prime duration factors.
Figure 5.
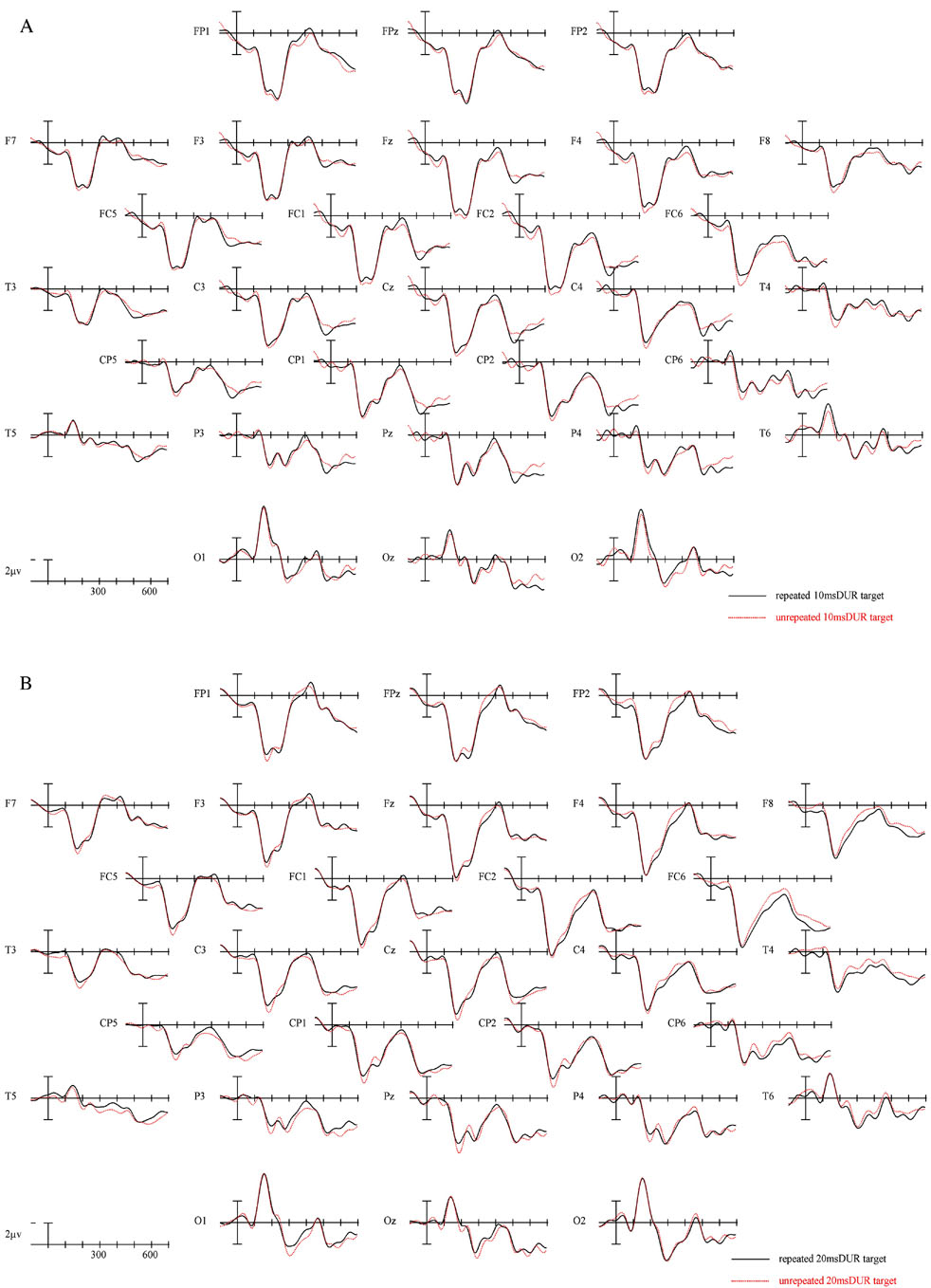
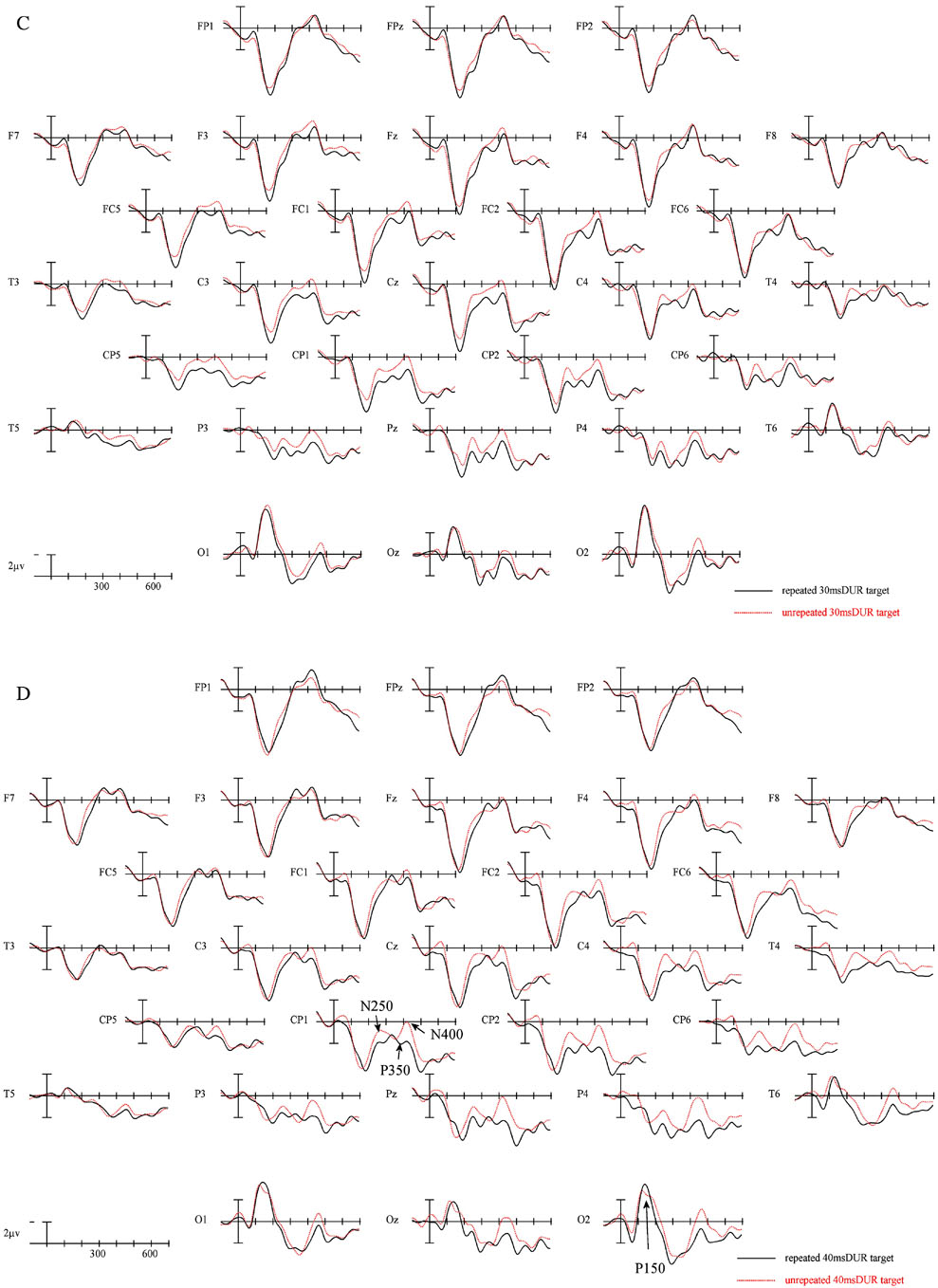
ERPs time locked to target onset in the two repetition conditions for the 10 ms Duration condition (a), 20 ms Duration condition (b) 30 ms Duration condition (C) and the 40 ms Duration condition (D).
Figure 6.

Panel a (top): Blowup of site CP1 from Figure 5 for all four SOAs. Panel b (bottom): Difference waves at CP1 calculated by subtracting Repeated from Unrelated Target ERPs for all four prime durations.
Figure 7.

Voltage maps calculated from difference waves (Unrelated-Repeated) for each of the four prime durations. Each row represents a different prime duration with activity calculated at 50 ms intervals surrounding the N250 epoch.
Figure 8.

Voltage maps calculated from difference waves (Unrelated-Repeated) for each of the four prime durations. Each row represents a different prime duration with activity calculated at 50 ms intervals surrounding the N400 epoch.
Analyses of ERP Data
P/N150 Epoch (125–175 ms Occipital and Frontal Sites)
As with the SOA manipulation in Experiment 1, there was a main effect of prime DURATION in Experiment 2 (F(3,69) = 3.66, p = .027). However, there were no main effects of Repetition in this epoch.
N250 Epoch (150–300 ms)
There were large main effects of prime DURATION across the head in Experiment 2 (midline: F(3,69) = 12.33, p < .001; C1: F(3,69) = 14.17, p < .001; C2: F(3,69) = 10.02, p < .001; C3: F(3,69) = 4.63, p = .006). As suggested by Figures 5 and 6 this was due to the two longest prime durations producing more negative going ERPs in this epoch than the two shortest prime durations. To confirm this observation we used the same analysis strategy used in Experiment 1 analyzing the four levels of prime duration separately. Only the two longest duration prime conditions produced a significant REPETITION effect (10 and 20 ms DURATION all ps > .49; 30 ms DURATION, midline: F(1,23) = 4.41, p = 047; C1: F(1,23) = 3.66, p = .068; C2: F(1,23) = 4.76, p = .04; C3: F(1,23) = 5.85, p = .024; 40 ms DURATION, midline: F(1,23) = 2.97, p = 098; C1: F(1,23) = 4.40, p = 047; C2: F(1,23) = 4.27, p = .05; C3: F(1,23) = 4.34, p = .049) with targets unrelated to the preceding prime word being more negative-going than targets that repeated across the prime and target positions. As in Experiment 1 this effect was widespread across the scalp.
N400 Epoch (400–550 ms)
In this final epoch there were again significant main effects of REPETITION in the overall ANOVA (midline: F(1,23) = 7.31, p = .013; C1: F(1,23) = 10.99, p = .003; C2: F(1,23) = 9.04, p = .006; C3: F(1,23) = 4.69, p = .041) and this effect tended to be larger at more posterior sites (REPETITION × ELECTRODE SITE interaction, midline: F(4,92) = 2.86, p = .092; C1: F(2,46) = 5.61, p = .019; C2: F(3,69) = 4.52, p = .038; C3: F(4,92) = 3.24, p = .073). Follow-up analyses at each separate duration again showed that the REPETITION effects was reliable only at the longer prime durations. At the two shortest durations there was no hint of a REPETITION effect (all ps > .25). At the 30 ms prime duration there were REPETITION effects at the more midline posterior and left hemisphere central posterior sites (midline, REPETITION × ELECTRODE SITE interaction: F(4,92) = 6.88, p = .007; C1, REPETITION × ELECTRODE × HEMISPHERE interaction: F(2,46) = 5.33, p = .013) but at the 40 ms duration REPETITION effects tended to be more robust across the scalp although they still showed a more posterior focus (REPETITION × ELECTRODE SITE interaction, midline: F(4,92) = 6.88, p = .007; C1: F(2,46) = 5.26, p = .015; C2: F(3,69) = 6.10, p = .017; C3: F(4,92) = 5.55, p = .02).
Time-course Analysis
As in Experiment 1 we also performed a time-course analyses on successive 50 ms epochs starting at target onset and continuing through the N400 epoch (450 ms). Reported in Table 2 are the main effects of REPETITION across nine electrode sites (F3, Fz, F4, C3, Cz, C4, P3, Pz, P4). As can be seen the effects of REPETITION had similar time-courses at the two longest stimulus durations, and as suggested by the above epoch analyses the two shortest duration produced little evidence of a REPETITION effect at any time point.
Table 2.
Time-course Analysis Experiment 2 (Prime Duration)
| 0–50 | 50–100 | 100–150 | 150–200 | 200–250 | 250–300 | 300–350 | 350–400 | 400–450 | 450–500 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 10 ms Duration | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| 20 ms Duration | ns | ns | ns | ns | ns | ns | - | ns | ns | ns |
| 30 ms Duration | ns | ns | ns | - | * | - | * | * | * | ns |
| 40 ms Duration | ns | ns | ns | - | * | * | ns | ** | * | - |
- p < .1
p < .05
p < .01
Behavioral Data
Participants detected 84.6% (SD = 8) of animal probes in the target position and fewer than 20% (SD=25) in the prime position within a window of 200 to 800 ms post-probe onset.
2.4 Experiment 2 Discussion
Consistent with Experiment 1, robust effects of repetition priming were apparent in the N250 and N400 ERP components when the duration of the prime was 40 ms. In addition a similar pattern was obtained when the duration of the prime was reduced to 30 ms. However, there was little evidence of priming effects with prime durations of 10 and 20 ms (cf. Figure 5a–d and Table 2). A similar pattern was obtained during the N400 epoch, with the 30 and 40 ms prime duration conditions producing robust priming effects and the 20 and 10 ms conditions showing no evidence of priming. This pattern of results defines the minimal amount of time that a word must be present, with standard luminance values (see procedure), for the visual word recognition system to extract critical orthographic and semantic information. Furthermore, the within-condition analysis of priming effects showed a very distinct pattern for priming in the N250 and priming in the N400 epochs. The N250 showed an increase in negativity with increasing prime duration that was most evident in the unrelated prime. On the other hand, the N400 showed an increased positivity in the repetition condition, and no significant change in the unrelated condition.
Finally, similar to Experiment 1 there was no significant effect in the P150 region at any stimulus duration. We will return to this point below.
3. General Discussion
The present experiments provide a further exploration of the temporal dynamics of masked repetition priming by recording ERPs to target words and manipulating prime-target SOA (with prime duration held constant) in Experiment 1, and prime duration (with SOA held constant) in Experiment 2. The results provide important information concerning the precise timing necessary for obtaining repetition priming effects in different ERP components, and provide an explanation for why some prior studies failed to find such priming effects. Moreover, the pattern of priming effects found as a function of SOA (Experiment 1) and prime duration (Experiment 2) help further constrain an interpretation of the processing reflected in the principal ERP components isolated in our prior work on visual word recognition (Grainger & Holcomb, in press; Holcomb & Grainger, 2006; Kiyonaga et al., 2007).
Concerning the SOA manipulation of Experiment 1, all four SOAs resulted in robust N400 repetition effects, but only the 60 and 180 ms conditions produced clear N250 repetition effects (see Figure 2). By the 300 ms SOA condition there was little evidence of this earlier component. This finding suggests that the N250 repetition effect rapidly dissipates at some time point between 180 and 300 ms after presentation of the prime, which likely explains its absence in previous unmasked repetition priming experiments where SOAs > 500 ms are the norm. Another interesting finding was that at an SOA of 180 ms the N250 component began as early as 100 ms, which is about 50 ms earlier than in the 60 ms SOA condition. This result suggests that when increasing prime-target ISI there is an initial increase in priming effects possibly due to more time being available for information extracted from the prime stimulus to continue to feed-forward activation to representations that will subsequently be involved in target word processing. However, the function relating ISI to priming effects is clearly non-monotonic, with some form of decay eventually offsetting the initial benefits of increasing ISI.
Concerning the prime duration manipulation of Experiment 2, there was little evidence for any priming at the two shortest prime durations (10 and 20 ms), and when priming effects did emerge at the two longest prime durations (30 and 40 ms) then it was equally evident in the N250 and N400 components. This pattern suggests that a minimal amount of processing of the prime stimulus is necessary to generate observable priming effects, and that once these conditions are satisfied, information is propagated through the entire system devoted to recognizing printed words. We examine the within-condition priming effects below.
3.1 Features, form, and meaning
The primary theoretical goal of the present study was to further constrain possible interpretations of the ERP components observed in our prior work on masked repetition priming (Holcomb & Grainger, 2006; Kiyonaga et al., 2007). This interpretation is couched in a model of visual word recognition that proceeds from visual features to orthographic and phonological representations and on to semantics. Three ERP components were isolated as being particularly relevant for the present study: The N/P150, the N250, and the N400. The N/P150 component is hypothesized to reflect feature-level processing, the N250 component is hypothesized to reflect form-level processing, and the N400 component is hypothesized to reflect meaning-level processing. In Figure 9 we have mapped the ERP time course data onto the putative cascade of processes described in the bimodal interactive-activation model of word recognition (Grainger, Diependaele, Spinelli, Ferrand, & Farioli, 2003; Grainger, Muneaux, Farioli, & Ziegler, 2005). Figure 9 illustrates our hypothesis that the N250 reflects the mapping of prelexical orthographic representations onto whole-word orthographic representations, as well as the prelexical mapping of orthography onto phonology (Grainger, Kiyonaga, & Holcomb, 2006). The earliest part of the N400, on the other hand, is thought to reflect the mapping of whole-word representations onto semantics. In a recent paper, Barber and Kutas (2006) provided a similar time course analysis of effects of different independent variables on ERP components generated during visual word recognition. Barber and Kutas’ analysis, based on a large number of published studies, fits well with the above proposal. Prelexical orthographic effects appear mainly in the window between 150 ms and 200 ms post-stimulus onset, whereas lexical influences are mainly visible in the window between 250 ms and 400 ms.
Figure 9.

Theoretical mapping of masked ERP priming effects onto the bimodal interactive-activation model. The model distinguishes featural, prelexical (O-units, P-units), and lexical (O-words, P-words) levels of representation for visual and auditory words, that map onto a common semantic/conceptual level of representation (O stands for “orthographic” and P for “phonological”). Since the present focus is on visual word recognition, the spoken word pathway has been dimmed.
The pattern of results reported in the present study also fit well within this framework. In language comprehension, individual word meanings must remain activated long enough to allow integration within a sentence-level interpretation. We therefore expected the priming effects that are subtended by semantic representations to be relatively insensitive to our SOA manipulation. This was indeed the case, with the N400 priming effect being unaffected by the interval between prime offset and target onset in Experiment 1. Word form representations (orthographic and phonological), on the other hand, must be rapidly suppressed as each new word is encountered during language comprehension. In this view, once a lexical entry has activated its conceptual representation(s) its lower level orthographic and phonological representations need to be quickly suppressed to make way for the next word (concepts on the other hand need to stay active to participate in comprehension). SOA was therefore expected to have a strong influence on priming effects that are subtended by form representations. This is exactly what was found in Experiment 1, with the N250 priming effect diminishing as a function of increasing SOA. The fact that the N250 effect disappeared between the 180 ms and the 300 ms SOA is consistent with estimates of the time required to complete form-level processing and to activate semantic representations.
Finally, feature-level representations were expected to be even more fragile than higher-level orthographic and phonological representations, and therefore to be even more affected by our SOA manipulation. Unfortunately, neither Experiment 1 nor Experiment 2 showed any significant priming effect on the N/P150 ERP component. This is a component reported by Holcomb and Grainger (2006) taking the form of a larger positivity to repeated compared to unrelated targets at occipital sites, primarily over the right hemisphere in an epoch starting near 100 ms post-target onset. A similar pattern was reported by Petit et al (2006) and Eddy et al., (2006) using letter and picture repetition priming tasks. Together these findings suggest that this earliest of priming effects reflects an early feature extraction process. So, it is peculiar that a similar effect was not replicated in either experiment of the current study. One possible reason for this might be that due to its small size this effect did not survive the interaction with SOA or Duration. In other words, the effect may be too small and/or restricted in time course that the longer SOA and shorter duration conditions prevented it from producing a reliable effect in the current experiments. Another related possibility is that we had relatively less power in the current experiments to extract such a small effect. Holcomb and Grainger used 80 stimuli per condition and 48 subjects. The current study used 24 subjects per experiment and 40 items per condition. To more carefully test for the presence of early effects we collapsed across the two conditions in Experiment 1 and 2 which where procedurally the most similar and which most closely match those used by Holcomb and Grainger; the 60 ms SOA condition of Experiment 1 and the 40 ms duration condition of Experiment 2. This effectively doubled the N for analysis purposes. With this approach we now found some evidence for a similar N/P150 effect (see Figure 10). Analyzing the mean amplitude between 125 and 175 ms (the same epoch used by Holcomb & Grainger, 2006) at the two most posterior sites (O1 and O2) there was a marginal Repetition effect (F(1,47) = 3.72, p = .06) with a larger negativity to Repeated compared to Unrelated items. This is the same pattern reported by Holcomb and Grainger (2006).
Figure 10.

ERPs averaged over Experiment 1 and 2 for Repeated and Unrelated items (40 ms duration and 60 ms SOA).
3.2 Incremental analysis
Another goal of the present study was to provide an incremental analysis of the influence of prime duration on each prime condition separately (Jacobs et al., 1995). This was achieved in Experiment 2 by holding SOA constant and increasing prime duration in steps of 10 ms. The results of this experiment suggest that the N250 component reflects an increase in negativity as prime duration increases that is offset in the repetition condition. It would appear that increasing the duration of the prime stimulus results in an increased difficulty in processing target stimuli that follow in rapid succession (i.e., with a short ISI, as was the case in Experiment 2). This fits well with the principle implemented in the interactive-activation model of word recognition, that simultaneously activated form representations inhibit each other (McClelland & Rumelhart, 1981; Grainger & Jacobs, 1996). Thus the prime stimulus activates a set of form representations that compete with the representations generated by the target stimulus. This competition is maximal when primes and targets are completely unrelated, and reduces as a function of the amount of orthographic and/or phonological overlap between prime and target word.
The N400 component, on the other hand, appears to reflect a decrease in negativity with increasing prime duration in the repetition prime condition, accompanied by little change in the unrelated prime condition. This fits with the principle that semantic representations associated with different words can be simultaneously activated with no cost.
3.3 Relation to previous ERP studies of early lexical processing
A number of previous studies using single word and sentence reading tasks have reported effects on ERP components which have been interpreted as reflecting early lexical processing. For example, Sereno et al (1998) reported effects of lexicality (words vs. nonwords) on the P1 component as early as 100 ms and effects of word frequency between 132 and 192 ms (on the N1 component). which they interpreted as indicating that lexical access is largely complete prior to 200 ms. A second study by Hauk and Pulvermuller (2004) found a similarly early effect of frequency and drew similar conclusions. More recently Dambacher et al (2006) found slightly later effects of frequency on the P200 component. Although it is not clear how the effects reported in these sentence and single word presentation paradigms map onto the masked priming results found in the current study, it is the case that roughly similar estimates of the time course of lexical processing are being made. One prediction based on our proposed mapping of ERP components onto component processes in word recognition (Figure 9), is that word frequency would influence processing in the masked priming paradigm as early as 200 ms post-stimulus onset due to the interactions between prelexical and whole-word representations. Note that this prediction relies on the assumption of cascaded processing inherent in the interactive activation model in which feedback from the fastest feedforward processes can affect on-going feedforward processing. Consistent with this prediction we have recently found a word frequency by repetition interaction on the N250 in the masked priming paradigm (Grainger & Holcomb, in preparation).
3.4 Conclusions
The different impact of our SOA and prime duration manipulations on the priming effects seen in the N250 and N400 ERP components provides further confirmation that these components do indeed reflect distinct underlying processing. The manner in which these components were affected by the manipulations of the present study, fits well with the hypothesis that the N250 reflects form-level processing, and the N400 primarily reflects semantic-level processing. The relatively weak effects found in the N/P150 ERP component further suggests that this component reflects processing at the level of visual features that is particularly sensitive to the masking procedure applied in the present study.
4. Experimental Procedure
4.1 Experiment 1
Participants
Twenty-four volunteers (13 female, mean age = 20.2, SD = 1.9) who were all undergraduate students at Tufts University were paid $25 to participate in this experiment. All were right-handed native speakers of English with normal or corrected to normal visual acuity.
Stimuli
The critical stimuli for this experiment were 320 pairs of five letter English words with a mean CELEX log frequency of 1.43 (SD = .58, range 0–2.97), a mean summed bigram frequency of 8069 (SD = 3108, range =1,690–19,763) and a mean neighborhood size of 2.9 (SD=2.49, range=0–15). The noncritical stimulus pairs were formed by combining 80 five letter animal names with 80 unrelated non-animal five letter words. For both the critical and non-critical items, the first member of each pair was referred to as the prime and the second member as the target. Stimulus lists consisted of 480 trials each containing a prime-TARGET pair of items, with the prime presented in lower case letters and the target in upper case (this was done in order to minimize the physical similarity between repeated items). In each of eight lists there were 160 trials where the target was a full repetition of the prime (e.g. table-TABLE) and 160 trials where the target was completely unrelated to the prime (e.g. mouth-TABLE). Forty items of each type (repeated and unrelated) were presented at each of four SOAs: 60 ms, 180s ms, 300 ms and 420 ms. Across lists (and participants), critical target words appeared in each of the four SOAs in both the repeated condition and the unrelated condition conditions, but within lists each target stimulus was presented only once. An important feature of this design is that the prime and target ERPs in the repeated and unrelated conditions and at each of the four SOAs are formed from exactly the same physical stimuli (across subjects) which should reduce the possibility of ERP effects across conditions due to differences in physical features or lexical properties.
For the 160 non-critical trials, 80 contained animal names in the prime position and a filler word in the target position while another 80 trials contained unrelated filler words in the prime position and animal names in the target position (20 in each position were presented at each of the four SOAs). The same 80 animal names were used once as primes and once as targets in each list. The animal names served as probe items in a go/no-go semantic categorization task in which participants were instructed to rapidly press a single button whenever they detected an animal name. Participants were told to read all other words passively (i.e., critical stimuli did not require an overt response). A practice session was administered before the main experiment to familiarize the participant with the procedure.
Procedure
Visual stimuli were presented on a 19” monitor set to a refresh rate of 100 Hz (which allows 10 ms resolution of stimulus control) and located 143 cm directly in front of the participant. Stimuli were displayed at high contrast as white letters on a black background in the Arial font (letter matrix 30 pixels tall × 15 pixels wide). Each trial began with the termination of a fixation stimulus in the middle of the screen. Five hundred milliseconds later, a forward mask of seven hash marks (#######) was presented for a duration of 500ms. The forward mask was replaced at the same location on the screen by a five letter lower case prime item for 40ms. The prime was then immediately replaced by a backward mask (#######). The backward mask remained on the screen for either 20ms, 140ms, 260 ms or 380 ms and was immediately replaced by the visual target in upper case letters for a duration of 300 ms. All target words were followed by a 900ms black screen which was replaced by a 2000ms fixation stimulus marking the end of the trial (see Figure 11 for a schematic of typical trial). Participants were asked to refrain from blinking and moving their eyes except when the fixation stimulus appeared on the screen.
Figure 11.

A typical trial with examples of the various types of items in the prime and target positions. Note that no participant was presented with any word on more than one trial. The bottom row of example items illustrate the two types of probe trials.
EEG recording procedure
Participants were seated in a comfortable chair in a sound attenuated darkened room. The electroencephalogram (EEG) was recorded from 29 active tin electrodes held in place on the scalp by an elastic cap (Electrode-Cap International – see Figure 12). In addition to the 29 scalp sites, additional electrodes were attached to below the left eye (to monitor for vertical eye movement/blinks), to the right of the right eye (to monitor for horizontal eye movements), over the left mastoid bone (reference) and over the right mastoid bone (recorded actively to monitor for differential mastoid activity). All EEG electrode impedances were maintained below 5 kΩ (impedance for eye electrodes was less than 10 kΩ). The EEG was amplified by an SA Bioamplifier with a bandpass of 0.01 and 40 Hz and the EEG was continuously sampled at a rate of 200 Hz throughout the experiment.
Figure 12.

Electrode montage and four analysis columns used for ANOVAs.
Data analysis
Averaged ERPs were formed off-line from trials free of ocular and muscular artifact (less than 10% of trials) and included eight types of targets formed from two levels of REPETITION (repeated vs. unrelated) and four levels of SOA (60 ms vs. 180 ms vs. 300 ms vs. 420 ms). Because the pre-target period contained systematically different timing of the prime and backward mask across the four SOAs, we decided to use the last 20 ms of the backward mask (i.e., the 20 ms before target onset) to baseline all target ERPs. Note that while this is a shorter period that normally used for baselining, the ERPs in Figure 1 do not suggest that there are worrisome differential effects carrying over from the pre-target period at any of the SOAs. The main analysis approach involved measuring mean amplitudes in two temporal epochs surrounding the two primary repetition ERP effects reported by Holcomb and Grainger: 175–300 for the N250, and 400 to 550 for the N400. Separate repeated measures analyses of variance (ANOVAs) were used to analyze the data in each of these epochs.1 The Geisser-Greenhouse (1959) correction was applied to all repeated measures with more than one degree of freedom. In order to thoroughly analyze the full montage of 29 scalp sites we employed an approach to data analysis that we have successfully applied in a number of previous studies (e.g., Holcomb et al., 2005). In this scheme the 29 channel electrode montage is divided up into seven separate parasagittal columns along the antero-posterior axis of the head (see Figure 12). The electrodes in each of three pairs of lateral columns and one midline column are analyzed in four separate ANOVAs. Three of these analyses (referred to as Column 1, Column 2 or Column 3) involved an anterior/posterior ELECTRODE SITE factor with either three, four or five levels, as well as a HEMISPHERE factor (Left vs. Right). The forth “midline” analysis included a single anterior/posterior ELECTRODE SITE factor with five levels. While the epoch measures capture effects on specific ERP components, in order to more carefully quantify the time-course of repetition effects, mean amplitudes in successive 50 ms epochs starting at target onset and running through the end of the typical N400 window (550 ms) were also used. Finally, because Holcomb and Grainger (2006) also found evidence for a small focal effect of repetition over occipital sites, we also measured the mean amplitude between 125 and 175 ms post-target onset at the two lateral occipital sites. These data were analyzed in a two Repetition by four SOA by two Hemisphere analysis.
4.2 Experiment 2
Participants
Twenty-four volunteers (12 female, mean age = 21.0, SD = 1.6) who were all undergraduate students at Tufts University were paid $25 to participate in this experiment (none were previously in Experiment 1). All were right-handed native speakers of English with normal or corrected to normal visual acuity.
Stimuli and procedure
The critical and non-critical stimuli for this experiment were the same five letter English words used in Experiment 1. All procedures were also identical with the exception of the SOA and prime durations. In Experiment 2 the SOA was held constant at 60 ms and the duration of primes was varied across four levels: 10 ms, 20 ms, 30 ms and 40 ms. A typical trial is illustrated in Figure 13.
Figure 13.

A typical trial in Experiment 2. Note that prime duration was systematically manipulated across 40 levels while prime-target SOA was held constant at 60 ms. All else was as in Experiment 1.
Acknowledgments
This Research was supported by grant numbers HD25889 and HD043251. The authors would like to thanks Lauren Dennis and Kristi Kiyonaga for her help in collecting data.
Footnotes
We did not measure the interval between the N250 and N400, a region that Holcomb & Grainger (2006) referred to as the P325 and hypothesized might reflect lexical selection. To get differential activity in this region Holcomb & Grainger included a partial repetition condition. Because the current experiments only had a simple word repetition priming manipulation it seemed unlikely that a P325 effect would emerge in the current experiments.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Contributor Information
Phillip J. Holcomb, Tufts University, Medford, Ma USA
Jonathan Grainger, CNRS & Aix-Marseille University, Marseille, France.
References
- Bentin S, McCarthy G, Wood CC. Event-related potentials, lexical decision and semantic priming. Electroencephalography and Clinical Neurophysiology. 1985;60:343–355. doi: 10.1016/0013-4694(85)90008-2. [DOI] [PubMed] [Google Scholar]
- Dambacher M, Kliegl R, Hofmann M, Jacobs AM. Frequency and predictability effects on event-related potentials during reading. Brain Research. 2006;1084:89–103. doi: 10.1016/j.brainres.2006.02.010. [DOI] [PubMed] [Google Scholar]
- Eddy M, Schmid A, Holcomb PJ. A new approach to tracking the time-course of object perception: masked repetition priming and event-related brain potentials. Psychophysiology. 2006;43:564–568. doi: 10.1111/j.1469-8986.2006.00455.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrand L. The masked repetition priming effect dissipates when increasing the interstimulus interval: Evidence from word naming. Acta Psychologica. 1996;91:15–25. [Google Scholar]
- Ferrand L, Grainger J. Phonology and orthography in visual word recognition: Evidence from masked nonword priming. Quarterly Journal of Experimental Psychology. 1992;45A:353–372. doi: 10.1080/02724989208250619. [DOI] [PubMed] [Google Scholar]
- Ferrand L, Grainger J. Effects of orthography are independent of phonology in masked form priming. Quarterly Journal of Experimental Psychology. 1994;47A:365–382. doi: 10.1080/14640749408401116. [DOI] [PubMed] [Google Scholar]
- Forster KI, Mohan K, Hector J. The mechanics of masked priming. In: Kinoshita S, Lupker SJ, editors. Masked Priming: The state of the art. NY: Psychology Press; 2003. [Google Scholar]
- Geisser S, Greenhouse S. On methods in the analysis of profile data. Psychometrica. 1959;24:95–112. [Google Scholar]
- Grainger J, Diependaele K, Spinelli E, Ferrand L, Farioli F. Masked repetition and phonological priming within and across modalities. Journal of Experimental Psychology: Learning, Memory and Cognition. 2003;29:1256–1269. doi: 10.1037/0278-7393.29.6.1256. [DOI] [PubMed] [Google Scholar]
- Grainger J, Ferrand L. Phonology and orthography in visual word recognition: Effects of masked homophone primes. Journal of Memory and Language. 1994;33:218–233. [Google Scholar]
- Grainger J, Jacobs AM. Masked partial-word priming in visual word recognition: Effects of positional letter frequency. Journal of Experimental Psychology: Human Perception and Performance. 1993;19:951–964. doi: 10.1037//0096-1523.19.5.951. [DOI] [PubMed] [Google Scholar]
- Grainger J, Jacobs AM. Orthographic processing in visual word recognition: A multiple read-out model. Psychological Review. 1996;103:518–565. doi: 10.1037/0033-295x.103.3.518. [DOI] [PubMed] [Google Scholar]
- Grainger J, Jacobs AM. Temporal integration of information in orthographic priming. Visual Cognition. 1999;6:461–492. [Google Scholar]
- Grainger J, Muneaux M, Farioli F, Ziegler J. Effects of phonological and orthographic neighborhood density interact in visual word recognition. Quarterly Journal of Experimental Psychology. 2005;58A:981–998. doi: 10.1080/02724980443000386. [DOI] [PubMed] [Google Scholar]
- Grainger J, van Heuven W. Modeling letter position coding in printed word perception. In: Bonin P, editor. The mental lexicon. New York: Nova Science; 2003. pp. 2–23. [Google Scholar]
- Grainger J, Kiyonaga K, Holcomb PJ. The time-course of orthographic and phonological code activation. Psychological Science. 2006;17:1021–1026. doi: 10.1111/j.1467-9280.2006.01821.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grainger J, Holcomb PJ. Neural constraints on a functional architecture for word recognition. In: Cornelissen P, Hansen P, Kringelbach M, Pugh K, editors. The neural basis of reading. Oxford University Press; Oxford: in press. [Google Scholar]
- Grainger J, Holcomb PJ. Early ERP masked priming effects are modulated by repetition and word frequency in preparation. [Google Scholar]
- Grossi G, Coch D. Automatic word form processing in masked priming: An ERP study. Psychophysiology. 2005;42:343–355. doi: 10.1111/j.1469-8986.2005.00286.x. [DOI] [PubMed] [Google Scholar]
- Hauk O, Pulvermüller F. Effects of word length and frequency on the human event-related potential. Clinical Neurophysiology. 2004;115:1090–1103. doi: 10.1016/j.clinph.2003.12.020. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ. Semantic priming and stimulus degradation: Implications for the role of the N400 in language processing. Psychophysiology. 1993;30:47–61. doi: 10.1111/j.1469-8986.1993.tb03204.x. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ, Reder L, Misra M, Grainger J. Masked priming: An event-related brain potential study of repetition and semantic effects. Cognitive Brain Research, Cognitive Brain Research. 2005;24:155–172. doi: 10.1016/j.cogbrainres.2005.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holcomb PJ, Grainger J. On the time course of visual word recognition: An event-related potential investigation using masked repetition priming. Journal of Cognitive Neuroscience. 2006;18:1631–1643. doi: 10.1162/jocn.2006.18.10.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobs AM, Grainger J, Ferrand L. The incremental priming technique: A method for determining within-condition priming effects. Perception & Psychophysics. 1995;57:1101–1110. doi: 10.3758/bf03208367. [DOI] [PubMed] [Google Scholar]
- Kiyonaga K, Midgley KJ, Holcomb PJ, Grainger J. Masked cross-modal repetition priming: An ERP investigation. Language and Cognitive Processes. 2007;22 doi: 10.1080/01690960600652471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McClelland JL, Rumelhart DE. An interactive activation model of context effects in letter perception: I. An account of basic findings. Psychological Review. 1981;88:375–407. [PubMed] [Google Scholar]
- Misra M, Holcomb PJ. The electrophysiology of word-level masked repetition priming. Psychophysiology. 2003;40:115–130. doi: 10.1111/1469-8986.00012. [DOI] [PubMed] [Google Scholar]
- Petit J, Grainger J, Midgley KJ, Holcomb PJ. On the time-course of processing in letter perception: A masked priming ERP investigation. Psychonomic Bulletin and Review. 2006;13:674–681. doi: 10.3758/bf03193980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rayner K. Eye movements in reading and information processing: 20 years of research. Psychological Bulletin. 1998;124:372–422. doi: 10.1037/0033-2909.124.3.372. [DOI] [PubMed] [Google Scholar]
- Schnyer DM, Allen JB, Forster KI. Event-related brain potential examination of implicit memory processes: Masked and unmasked repetition priming. Neuropsychology. 1997;11:243–260. doi: 10.1037//0894-4105.11.2.243. [DOI] [PubMed] [Google Scholar]
- Sereno SC, Rayner K, Posner MI. Establishing a time-line of word recognition: evidence from eye movements and event-related potentials. NeuroReport. 1998;9:2195–2200. doi: 10.1097/00001756-199807130-00009. [DOI] [PubMed] [Google Scholar]