

Abstract
Ongoing neurogenesis in the adult mammalian dentate gyrus and olfactory bulb is generally accepted, but its existence in other adult brain regions is highly controversial. We labeled newly born cells in adult rats with the S-phase marker bromodeoxyuridine (BrdU) and used neuronal markers to characterize new cells at different time points after cell division. In the neocortex and striatum, we found BrdU-labeled cells that expressed each of the eight neuronal markers. Their size as well as staining for γ-aminobutyric acid (GABA), glutamic acid decarboxylase 67, calretinin and/or calbindin, suggest that new neurons in both regions are GABAergic interneurons. BrdU and doublecortin-immunoreactive (BrdU+/DCX+) cells were seen within the striatum, suggesting migration of immature neurons from the subventricular zone. Surprisingly, no DCX+ cells were found within the neocortex. NG2 immunoreactivity in some new neocortical neurons suggested that they may instead be generated from the NG2+ precursors that reside within the cortex itself.
Introduction
It is widely believed that new neurons are added to only two regions of the adult mammalian brain: the dentate gyrus, in which new granule cells arise from an in situ dividing population, and the olfactory bulb, to which new granule cells and periglomerular cells migrate from the subventricular zone (SVZ) adjacent to the lateral ventricle. Neurogenesis in the adult neocortex was reported many years ago using tritiated thymidine combined with light microscopy or electron microscopy to show that the newborn cells had axon hillocks/initial segments as well as somatic and dendritic synapses (Altman, 1962; Kaplan, 1981). More recently, ongoing neocortical neurogenesis has been observed in both rats and macaques, using BrdU labeling and confocal microscopy to show retrograde labeling of axons and staining for the mature neuronal marker NeuN (Gould et al., 1999b, 2001; Bernier et al., 2002). However, the existence of adult neurogenesis in the neocortex is not widely accepted due to negative reports from other groups using similar methods (Kornack and Rakic, 2001; Rakic, 2002; Ehninger and Kempermann, 2003; Koketsu et al., 2003). Additional studies have found no neocortical neurogenesis in normal rodents, despite the appearance of compensatory neurogenesis after cortical damage (Gu et al., 2000; Magavi et al., 2000; Jiang et al., 2001).
Coincident with recent reports of neocortical neurogenesis, several studies have found young neurons that appear to exit the SVZ or rostral migratory stream and migrate through subcortical white matter tracts (Gould et al., 1999b, 2001; Nacher et al., 2001; Bernier et al., 2002; Luzzati et al., 2003), suggesting that some neurons generated in the adult SVZ may incorporate into a region or regions other than the olfactory bulb. In addition to the neocortex, adult-generated neurons have recently been reported in the adult primate striatum (Bedard et al., 2002); however, three other groups have failed to find any new neurons in this structure under normal conditions (Benraiss et al., 2001; Pencea et al., 2001; Teramoto et al., 2003).
In this study, we used the S-phase marker BrdU along with several cell-type specific immunohistochemical markers to find evidence for or against the existence of newborn neurons in the normal rat neocortex. In addition, we examined early time points after BrdU labeling to investigate whether new neurons appear to be derived from the neuronal precursors residing in the SVZ.
Results
New neurons in neocortex
Based on the premise that new cortical neurons would migrate from the SVZ across the subcortical white matter, we examined BrdU-immunoreactive (+) cells in the deep portion of the rostral neocortex (Bregma 1.20–3.20 mm; Paxinos and Watson, 1998) at several time points after BrdU injection (Table I). Examination of brain sections from rats injected with BrdU 4–5 wk earlier revealed BrdU+ cells that were double labeled with antibodies against each of three neuronal markers: NeuN, a marker specific for mature neurons; EAAC-1, a neuronal glutamate transporter; and HuC/D, neuron-specific RNA binding proteins (Fig. 1; Tables II and III). BrdU+ cells double labeled with neuron-specific enolase (NSE), a marker of metabolically active neurons (Marangos and Schmechel, 1987), were found only at a later time point, 11–12 wk after BrdU injection (Table III). BrdU+/NeuN+ neurons were also observed at this longer survival time (Fig. 1).
Table I. BrdU injections and survival times.
| Group | N | Injections | Survival |
|---|---|---|---|
| 2 h | 3 | 1 | 2 h |
| 1–2 wk | 3 | 8 (2×/day, bidaily) | 5 d |
| 3–4 wk | 3 | 5 (1×/day) | 25 d |
| 4–5 | 10 | 4 (1×/day) | 32 d |
| 11–12 wk | 3 | 5 (1×/day) | 82 d |
Groups of rats were injected with BrdU (300 mg/kg, i.p.) between one and eight times at the frequencies indicated. The survival column indicates the amount of time between the last BrdU injection and perfusion of the animals. N indicates the number of animals in each group.
Figure 1.

Some cells born in the adult cortex express neuronal markers. (A) A 4–5-wk-old cell labeled with BrdU and the mature neuronal marker NeuN is shown within the green box in A1. Sample z-planes through the boxed area in A1 are shown with color separation in A2, demonstrating colocalization of BrdU and NeuN but not NG2; note that the green BrdU label and the red NeuN label both peak in brightness at the same level (z = 1.5 μm). The location of the cell shown in A is circled in the small diagram of a coronal section from the appropriate rostrocaudal level (Bregma 1.2); the analyzed region of cortex is shown in yellow on the diagram, and the subcortical white matter, used as a boundary for the analysis, is shown in blue. (B) A 4–5-wk-old BrdU+ (NG2−) neuron labeled with the neuronal marker HuC/D is shown within the box in the multi-channel view in B1 and in orthogonal views in B2. Also note the BrdU+/NG2+/Hu− cell in B1. (C) A 4–5-wk-old BrdU+/NeuN+ neuron that is also immunoreactive for the neuronal glutamate transporter EAAC-1 is seen in color separation in C1, with the blue nuclear counterstain Hoechst 33258 (Hch) added in the multi-color view. Higher magnification views of selected individual z-planes are shown in C2. (D) An 11–12-wk-old BrdU+/NeuN+/NG2− neuron. (E) An 11–12-wk-old BrdU+/NSE+ neuron is shown within the box in E1, at higher magnification with color separation in E2, and in orthogonal views in E3. Images show single focal planes (C1, E1, E2) or Z-axis projections of 14 × 0.64 μm, i.e., 14 planes, 0.64 μm apart (A1), 14 × 0.51 μm (B), 8 × 1.05 μm (D1), or 14 × 0.5 μm (E3). Bars: 5 μm in orthogonal views; 10 μm in all other views.
Table II. Immunostaining of BrdU-labeled cells in the adult rat neocortex and striatum.
| Age of BrdU-labeled cells
|
||||||
|---|---|---|---|---|---|---|
| Immunoreactive for | 2 h | 1–2 wk | 3–4 wk | 4–5 wk | 11–12 wk | |
| Neocortex (deep layers) | DCX | − | − | − | − | − |
| CRMP4 | na | + | + | na | na | |
| NG2 | 92.4 ± 3.8%, 140/152, n = 3 (1) | + | + | 58.5 ± 2.7%, 466/798, n = 10 (0.5) | + | |
| NeuN | − | − | na | 0.4 ± 0.13%, 33/7624, n = 10 (4) | + | |
| NeuN & NG2 | − | na | na | 0.1 ± 0.07%, 11/7624, n = 10 (4) | +/− | |
| not NeuN or NG2 | 7.6 ± 3.8%, 12/152, n = 3 (1) | na | na | 40.9 ± 2.8%, 325/798, n = 10 (0.5) | + | |
| HuC/D | na | na | na | + | na | |
| NSE | na | na | na | − | + | |
| EAAC-1 & NeuN | na | na | na | + | na | |
| GABA | na | na | na | + | + | |
| GAD-67 | na | na | na | − | + | |
| CB | na | na | na | + | + | |
| CR | na | na | na | + | + | |
| Striatum | DCX | − | + | + | + | − |
| DCX/CRMP4 | na | + | + | na | na | |
| DCX/NeuN | − | − | + | + | − | |
| NeuN | − | − | + | 5.8 ± 0.9%, 57/995, n = 8 (2) | + | |
| NSE | na | na | na | − | + | |
| GAD-67 | na | na | na | + | + | |
| CR | na | na | na | + | + | |
In cases where quantitative analysis was performed, mean % of BrdU+ cells ± SEM, d, the number of cells in category/number of BrdU+ cells analyzed, and n, the number of brains analyzed (number of sections per brain), are provided.
+, some cells of this type observed; −, not observed; na, not analyzed; +/−, very few observed.
Table III. Antibodies used for immunohistochemistry.
| Primary antibody
|
Made ina
|
Dilution
|
Source
|
Cell type labeled
|
References
|
|
|---|---|---|---|---|---|---|
| Anti-BrdU | Rat | 1:100 | Accurate | Cells in S-phase when BrdU is injected |
Dayer et al., 2003 | |
| Anti-BrdU | Mouse | 1:100 | Becton Dickinson | Cells in S-phase when BrdU is injected |
Dayer et al., 2003 | |
| Anti-DCX | Rabbit | 1:500 | a gift from C. Walsh (Harvard Medical School, Boston, MA) |
Immature neurons |
Minturn et al., 1995; Gleeson et al., 1999; Quinn et al., 1999 |
|
| Anti-DCX (COOH terminus) |
Goat | 1:100 | Santa Cruz Biotechnology, Inc. | “ | “ | |
| Anti–CRMP-4 (Ab-25) |
Rabbit | 1:2,500 | a gift from S. Hockfield (Yale University, New Haven, CT) |
“ | “ | |
| Anti-HuC/D | Mouse | 1:50 | Molecular Probes | Neurons | Okano and Darnell, 1997 | |
| Anti-NSE | Rabbit | 1:1,000 | Polysciences | Mature neurons |
Marangos and Schmechel, 1987; Mullen et al., 1992 |
|
| Anti-NeuN | Mouse | 1:250 | CHEMICON International, Inc. | |||
| Anti-EAAC1 | Rabbit | 1:500 | a gift from J. Rothstein (Johns Hopkins University, Baltimore, MD) |
Neuronal glutamate transporter |
Conti et al., 1998; Danbolt, 2001 |
|
| Anti-GABA | Rabbit | 1:2,000 | Sigma-Aldrich | GABAergic neurons |
Rosier et al., 1995; Gabbott and Bacon, 1996 |
|
| Anti-GAD-67 | Rabbit | 1:1,000 | a gift from J.-Y. Wu (Florida Atlantic University, Boca Raton, FL) |
|||
| Anti-CR | Mouse | 1:250 | CHEMICON International, Inc. | Subclasses of GABAergic neurons |
Gabbott and Bacon, 1996; Wichterle et al., 2001 |
|
| Anti-CR | Rabbit | 1:500 | CHEMICON International, Inc. | “ | “ | |
| Anti–calbindin D-28K |
Mouse | 1:5,000 | Swant | “ | “ | |
| Anti-parvalbumin | Mouse | 1:1,000 | Sigma-Aldrich | “ | “ | |
| Anti-somatostatin28 | Rabbit | 1:2,000 | CHEMICON International, Inc. | “ | “ | |
| Anti-NG2 | Rabbit | 1:250 | CHEMICON International, Inc. | Chondroitin sulfate proteoglycan | Mallon et al., 2002 | |
| Anti-GFAP | Guinea pig | 1:500 | Advanced ImmunoResearch | Astrocytes and radial glia | Palmer et al., 2000 | |
| Anti–S-100β | Mouse | 1:1,000 | Sigma-Aldrich | Astrocytes and other cell types | Vives et al., 2003 | |
| Anti-CNPase | Mouse | 1:250 | Sigma-Aldrich | Oligodendrocytes | Gould et al., 2001 | |
| Anti–Iba-1 | Rabbit | 1:250 | a gift from Y. Imai (National Institute of Neuroscience, Tokyo, Japan) |
Microglia |
Imai et al., 1996; Ehninger and Kempermann, 2003 |
|
| Secondary antibody | Made ina | Dilution | Source | Fluorescent conjugate | ||
| Anti–rat | Goat | 1:250 | Molecular Probes | Alexa 488, 568, or 647 | ||
| Anti–mouse | “ | |||||
| Anti–rabbit | “ | |||||
| Anti–rat | Donkeyb | 1:250 | Molecular Probes | Alexa 488, 568, or 647 | ||
| Anti–mouse | “ | |||||
| Anti–rabbit | “ | |||||
| Anti–goat | “ | |||||
| Anti–guinea pig | Donkeyc | 1:250 | Jackson ImmunoResearch Laboratories |
Cy3 or Cy5 | ||
| Anti–rabbit | “ | |||||
All antibodies made in rat and mouse were monoclonal; all others were polyclonal.
Secondary antibodies made in donkey were used whenever one of the primary antibodies was goat anti-DCX.
Secondary antibodies purchased from Jackson ImmunoResearch Laboratories were used for anti-GFAP staining to avoid cross-detection between anti-GFAP and Molecular Probes anti–rabbit secondary antibodies.
Systematic quantitative analysis of the deepest 500 μm of the rostral neocortex (comprising layer VI and in some areas part of layer V) was done in one half section (Bregma 1.2 mm) from each of 10 rats injected 4–5 wk earlier. This analysis revealed seven BrdU+ cells that were also NeuN+ (4.9 ± 2.9 cells/mm3; 0.6 ± 0.3% of 798 BrdU+ cells, both mean ± SEM), all with NeuN staining comparable to that of neighboring neurons. To look for additional double-labeled cells using a less time-consuming method of analysis, an additional 3.5 sections (Bregma 1.2–3.2) from each of the 10 rats were analyzed by scanning on the confocal microscope only those BrdU+ cells that appeared to be NeuN+ under epifluorescence. This second scan resulted in 26 cells BrdU+/NeuN+ cells (2.8 ± 0.9 cells/mm3; 0.4 ± 0.1% of 6826 BrdU+ cells), for an overall density of 3.1 ± 1.1 cells/mm3. Variability was seen across rats: 3 of the 10 brains analyzed contained no BrdU+/NeuN+ cells in any of the four sections sampled, whereas one brain contained as many as 11 BrdU+/NeuN+ cells in four sections. High variability is also seen across individuals in the dentate gyrus (Dayer et al., 2003), where it is likely to reflect a combination of genetic and environmentally induced differences in rates of neurogenesis (Kempermann et al., 1997a,b). BrdU+/NeuN+ cells were observed within the infralimbic cortex, cingulate cortex, somatosensory cortex, secondary motor cortex, and claustrum. No double-labeled cells were seen in the intervening primary motor cortex, which comprised ∼20% of the region analyzed.
Because of their small size and lack of cytoplasmic NeuN staining, none of the cells double labeled with BrdU and neuronal markers had the appearance of pyramidal neurons. All had small nuclei, 5–10 μm in diameter, similar in size to newborn neurons previously found in the adult mammalian cortex (Kaplan, 1981; Magavi et al., 2000; Gould et al., 2001). The 33 BrdU+/NeuN+ cells counted above had a mean nuclear diameter of 7.6 ± 0.2 μm, and five BrdU+/NSE+ cells had nuclei measuring 7.4 ± 0.7 μm. This small size suggested that newborn neurons in the neocortex were interneurons, a possibility that was examined by immunostaining for the neurotransmitter γ-aminobutyric acid (GABA). A small fraction of 4–5-wk-old BrdU+ cells were GABA+ (Fig. 2, A and B). Because GABA immunoreactivity could potentially result from uptake rather than production of GABA, sections were also stained for the GABA synthesizing enzyme glutamic acid decarboxylase 67 (GAD-67). No BrdU+/GAD67+ cells were found in the deep layers of the neocortex 4–5 wk after BrdU injection. However, several BrdU+/GAD67+ cells were encountered at the 11–12 wk time point (Fig. 2, C and D). Why 4–5-wk-old cells would label for GABA and not for GAD is unclear; however, strong GABA immunoreactivity and weaker GAD67 expression have previously been observed in small cortical neurons in the adult primate, where this pattern was thought to result from relatively low activity levels suggesting immature neurons (Hendrickson et al., 1994). All of the BrdU+/GABA+ and BrdU+/GAD67+ cells also had small nuclei, 7.4 ± 0.3 μm and 7.3 ± 0.3 μm in diameter, respectively. Calcium binding protein expression was examined, because functional classes of GABAergic interneurons are often identified using these markers (Gabbott and Bacon, 1996; Gabbott et al., 1997). Calbindin (CB) and calretinin (CR) immunoreactivity were each observed in 4–5-wk-old and 11–12-wk-old BrdU+ cells (Fig. 3). In triple-labeled sections, no BrdU+/CR+ cells appeared to be CB+, suggesting that there are two distinct classes of newly generated cortical interneurons or, alternatively, that new interneurons make a developmental switch from CR to CB expression like the adult-born dentate gyrus granule neurons (Brandt et al., 2003). BrdU+/CR+ and BrdU+/CB+ cells also had small nuclear diameters (7.0 ± 0.3) like cells labeled with other neuronal markers. No 4–5-wk-old BrdU+/parvalbumin+ cells or BrdU+/somatostatin+ cells were found.
Figure 2.

Some cells born in the adult cortex express markers of GABAergic interneurons. (A) A 4–5-wk-old BrdU+/GABA+ cell is shown within the boxed area in the multi-channel view in A1 and at higher magnification in A2. (B) An 11–12-wk-old BrdU+/GABA+ cell in color separation in B1 and in orthogonal views in B2. (C and D) 11–12-wk-old BrdU+/GAD67+ neurons in the cortex. GAD stains cell bodies of some GABAergic neurons but is also strongly expressed in nerve terminals, which appear as GAD+ specks around cell bodies of non-GABAergic neurons. The BrdU+/GABA+ and BrdU/GAD67+ cells in A and C appear to be satellite cells, based on their close apposition to other cells. Images show single focal planes (A2, D1) or Z-axis projections of 4 × 1.71 μm (A1), 5 × 1.17 μm (B), or 7 × 0.87 μm (C). Bars: 5 μm in orthogonal views; 10 μm in all other views.
Figure 3.

Some cells born in the adult cortex express CB or CR, markers of specific interneuron classes. (A) A pair of 4–5-wk-old BrdU+/CB+ neurons is shown within the box in A1 and at higher magnification color separation in A2. Pairs of BrdU+ labeled cells were frequently observed, suggesting local division from a common precursor. Note the larger BrdU−/CB+ interneuron in the lower left of A1. (B) An 11–12-wk-old BrdU+/CB+ neuron is shown within the box in the multi-channel view in B1 and in orthogonal views in B2. Note the larger BrdU−/CB+ interneuron in the upper left of B1. (C and D) 4–5-wk-old BrdU+/CR+ neurons are shown in color separations in C and (a different cell) in orthogonal views in D. In C, the BrdU+/CR+ neuron and an adjacent BrdU−/CR− cell both appear to be a satellites of a large cell on the left with a speckled nucleus visible in Hoechst staining. (E) A pair of 11–12-wk-old BrdU+/CR+ neurons are shown within the boxes in E1 and in orthogonal views in E2 and E3. Images show Z-axis projections of 6 × 4.96 μm (A1), 10 × 2.03 μm (A2), 7 × 3.17 μm (B1), 9 × 0.84 μm (B2), 14 × 1.47 μm (C), 30 × 0.64 μm (D), 6 × 2.24 μm (E1), 12 × 0.55 μm (E2), or 14 × 0.63 μm (E3). Bars: 5 μm in orthogonal views; 10 μm in all other views.
Because of their close apposition to other, generally larger, cells (Figs. 2 and 3), some BrdU+/GABA+, BrdU+/GAD67+, BrdU+/CB+, and BrdU+/CR+ cells appeared to be “satellite cells”. This suggests that some satellite cells, previously described as cells that could be mistaken for the nuclei of BrdU-labeled pyramidal cells (Gould et al., 1999b; Kornack and Rakic, 2001), may actually be newly born GABAergic neurons themselves. If this is the case, BrdU+/NeuN+ cells may have been undercounted in the quantitative analysis, because all satellite cells were classified as NeuN-nonimmunoreactive (NeuN−) in this analysis, due to the difficulty in determining the NeuN immunoreactivity of a small cell in very close proximity to a large NeuN+ cell. Even using high resolution Z-sectioning, NeuN immunoreactivity from the large neuron precludes unambiguous scoring of NeuN expression in a closely apposed BrdU+ satellite cell (Fig. 4 A).
Figure 4.
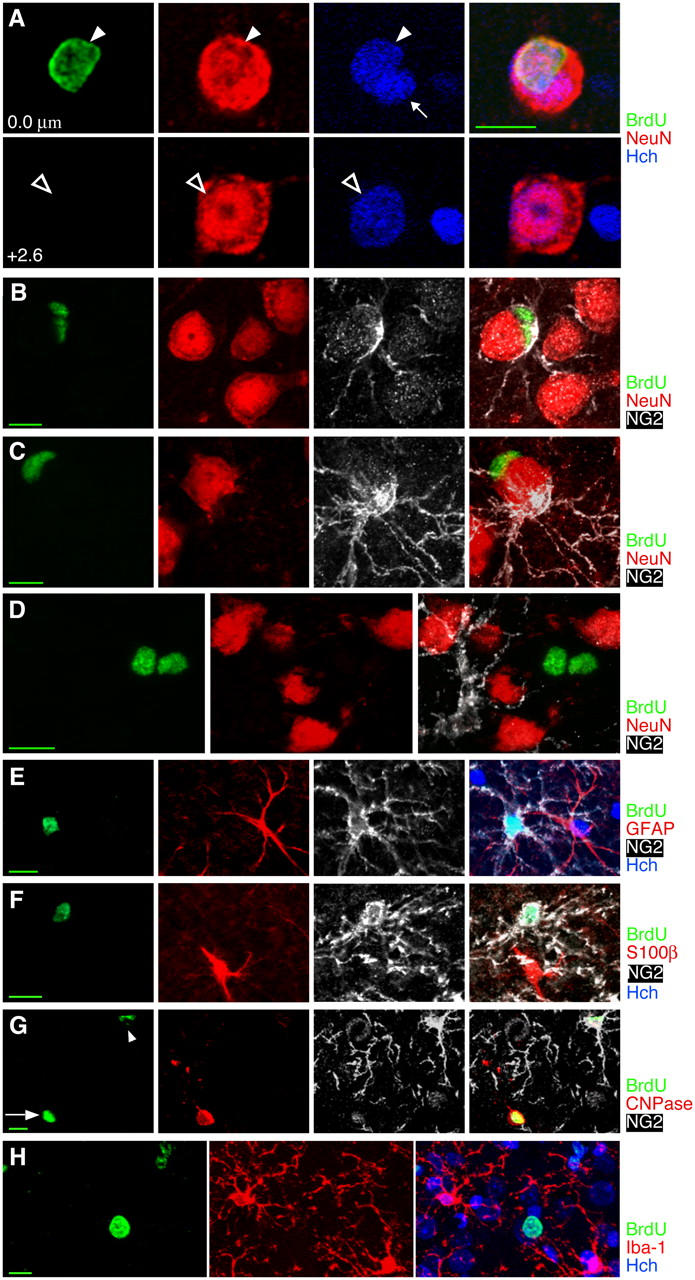
Many cells in the adult rat cortex do not appear to be neurons or commonly recognized glial types. (A) BrdU+ satellite cells closely apposed to large neurons could lead to erroneous identification of the large neurons as BrdU labeled. However, Hoechst nuclear counterstain and z-sectioning make it clear that the 4–5-wk-old BrdU+ cell nucleus (arrowhead at 0.0 μm) is not the nucleus of the large neuron (hollow arrow at 2.6 μm) and must instead belong to a satellite cell. A second, BrdU−, satellite cell can also be seen with Hoechst counterstain (small arrow at 0.0 μm). (B and C) Some but not all 4–5-wk-old BrdU+ satellite cells express the chondroitin sulfate proteoglycan NG2, characteristic of dividing or immature cells. Two satellite cells, one BrdU+/NG2− and one BrdU−/NG2+ can be seen in C. (D) A pair of 4–5-wk-old BrdU+ cells are NG2− and NeuN−, as were ∼40% of 4–5-wk-old BrdU+ cells. (E–G) Newborn NG2+ cells do not stain for astrocyte, oligodendrocyte, or microglial markers. (E) A BrdU+/NG2+ cell and a BrdU−/GFAP+ cell 2 h after BrdU injection. (F) A BrdU+/NG2+ cell and a BrdU−/S100β+ cell 2 h after BrdU injection. (G) A 3–4-wk-old BrdU+ cell in the cortex expressing the oligodendrocyte marker CNPase (arrow) shows faint nuclear staining of NG2 around the nucleus but none of the NG2+ processes that defined NG2+ cells. A second BrdU+ cell (arrowhead) shows strong NG2 staining and very faint CNPase staining. (H) A solidly stained BrdU+ cell and a pair of cells with speckled BrdU staining, all 4–5 wk old and all nonimmunoreactive for the microglial marker Iba-1. Images show single planes (B and E) or Z-axis projections of 10 × 0.69 μm (C), 7 × 1.05 μm (D), 8 × 1.72 μm (F), 9 × 0.67 μm (G), or 18 × 1.38 μm (H). Bars: 5 μm in orthogonal views; 10 μm in all other views.
Source of new neocortical neurons
To investigate whether new cortical neurons arise from precursors in the SVZ, coronal sections through the rostral forebrain (Bregma 0.48–2.20 mm) were immunolabeled for BrdU and the immature neuronal marker doublecortin (DCX). Several isolated BrdU+/DCX+ cells were observed within the subcortical white matter of the forceps minor and external capsule (Fig. 5, A–C). These cells had the appearance of migrating young neurons, with elongated cell bodies and one or two long leading processes directed away from the SVZ. Triple staining for BrdU, DCX, and the immature neuronal marker CRMP4 (sometimes referred to as TOAD-64 or TUC-4) revealed that all BrdU+/DCX+ cells were also CRMP4+ (Fig. 5 C). Surprisingly, no DCX+ cells, with or without BrdU labeling, were observed within the deep layers of the neocortex in 48 sections from 12 brains. However, a small number of 1–4-wk-old BrdU+/CRMP4+ cells that were DCX- were observed in this region (Fig. 5 D). It is possible that BrdU+/DCX+ cells enter from the subcortical white matter and down-regulate DCX as soon as they do so. However, the lack of DCX immunoreactivity in BrdU+/CRMP4+ cells, along with the observation that BrdU+/NeuN+ cells, BrdU+/CB+ cells, and BrdU+/CR+ cells often occurred in pairs <25 μm apart (4 of 33 cells, 2 of 5 cells, and 6 of 10 cells, respectively), suggested that new cortical neurons might arise from in situ progenitors rather than from the SVZ.
Figure 5.
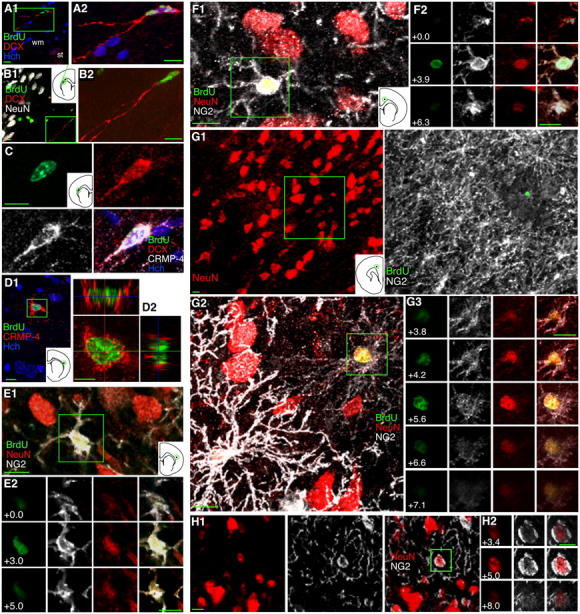
Young cells in the adult cortex expressing CRMP4, NG2, and NeuN, but not DCX, suggest in situ neurogenesis. (A–C) Isolated 1–2-wk-old BrdU+/DCX+ and BrdU+/DCX+/CRMP4+ neurons in the subcortical white matter (wm) dorsal to the striatum (st), have long processes suggesting leading processes. DCX+ cells were relatively easy to find in the SVZ and subcortical white matter but were never seen in the neocortex. (D) A 3–4-wk-old BrdU+/CRMP4+ cell in the cortex shown in the box in D1 and in orthogonal views in D2. (E–G) 4–5-wk-old BrdU+/NeuN+/NG2+ neurons in the cortex suggest a transition from NG2+ cells to NeuN+ cells. NG2 immunoreactivity in processes of BrdU+/NeuN+ cells was either moderate (E and F) or faint (G) and appeared to be inversely correlated with the intensity of NeuN staining. A pair of BrdU+ cells is shown in G2; the strong NG2 staining of the cell on the left nearly obscures its green nucleus. (H) A NeuN+/NG2+ neuron in a section not immunostained for BrdU indicates that this coexpression is not related to BrdU labeling. Images show single a focal plane (H) or Z-axis projections of 7 × 0.92 μm (A1), 14 × 0.58 μm (A2), 10 × 2.28 μm (B1), 15 × 1.5 μm (B2), 8 × 0.96 μm (C), 9 × 1.08 μm (D1), 16 × 0.57 μm (D2), 12 × 0.45 μm (E1), 9 × 0.78 μm (F1), 16 × 3.35 μm (G1), or 16 × 0.47 μm (G2). Bars: 5 μm in orthogonal views; 10 μm in all other views.
To explore this possibility, the identity of proliferating cells in the deep cortical layers was examined 2 h after BrdU injection. 92.4% ± 3.8 of 152 BrdU+ cells scanned in the deepest 500 μm of the neocortex had cell bodies and processes that were immunoreactive for the chondroitin sulfate proteoglycan NG2. The majority of BrdU+/NG2− cells could be identified as endothelial cells based on their distinctive morphology, i.e., a flattened and curved shape consistent with wrapping around a blood vessel (Palmer et al., 2000). None of the BrdU+ neocortical cells scanned at this 2 h time point were NeuN+. However, at the 4–5 wk survival time point, processes with faint to moderate NG2 immunoreactivity were observed in 2 of the 7 BrdU+/NeuN+ cells in one sample and in 9 of the 26 BrdU+/NeuN+ cells in the second sample (Fig. 5, E–G). NeuN+/NG2+ cells were also found in sections not stained for BrdU, indicating that colabeling was not an artifact of BrdU staining (Fig. 5 H). In rare cases, 11–12-wk-old BrdU+/NeuN+ neurons still displayed faint NG2 immunoreactivity.
Nearly 60% of 4–5-wk-old BrdU+ cells were NG2+/NeuN−. Many of these cells appeared to be satellites of large neurons (Fig. 4, B and C), similar to some of the BrdU+ cells labeled with neuronal markers. NG2+ cells were further characterized using several glial markers (Fig. 4). A recent study showed NG2 staining of EGFP+ cells in S100β-EGFP expressing transgenic mice (Vives et al., 2003). However, we found no NG2+/S100β+ cells among 130 NG2+ and 118 S100β+ cells. Triple labeling for BrdU, NG2, and either GFAP or CNPase showed that none of the BrdU+/NG2+ cells expressed these astrocyte or oligodendrocyte markers. A few cells were double labeled with BrdU and either CNPase, S100β, or the microglial marker Iba-1, but the number of these new glial cells was low and could not account for the ∼40% of BrdU+ cells that were NG2− and NeuN− in the systematic analysis (Table II; Fig. 4 D).
New neurons in the striatum
To investigate the fate of BrdU+/DCX+ cells in the SVZ that do not follow the rostral migratory stream and do not appear to enter the cortex, we examined the region surrounding the SVZ 1–2 wk after BrdU injection. A large number of 1–2-wk-old BrdU+/DCX+ cells were observed in the portion of the SVZ stretching laterally from the dorsolateral corner of the lateral ventricle, between the striatum and corpus callosum (Fig. 6, A and B). Examination of the striatum revealed BrdU+/DCX+/CRMP4+ cells whose location and morphology suggested that they could be migrating within the fiber tracts that course through the striatum (Fig. 6, B and C). A few BrdU+/DCX+/CRMP4+ cells were also found within the striatal matrix, i.e., the portion of the striatum outside the fiber tracts, at this time point. 2 wk later (3–4 wk after BrdU injection), BrdU+/DCX+ cells had more branched processes (Fig. 6, E and F). BrdU+/DCX+/NeuN+ neurons were also found in both the dorsomedial striatum and nucleus accumbens at this time point (Fig. 6, G–I). 4–5 wk after BrdU injection, BrdU+/NeuN+ cells in the striatum had rounder nuclei than BrdU+/NeuN+ cells observed at 3–4 wk (Fig. 7 A). 11–12 wk after BrdU injection, BrdU+/NSE+ as well as BrdU+/NeuN+ cells were found in the striatum (Fig. 7, B–D).
Figure 6.
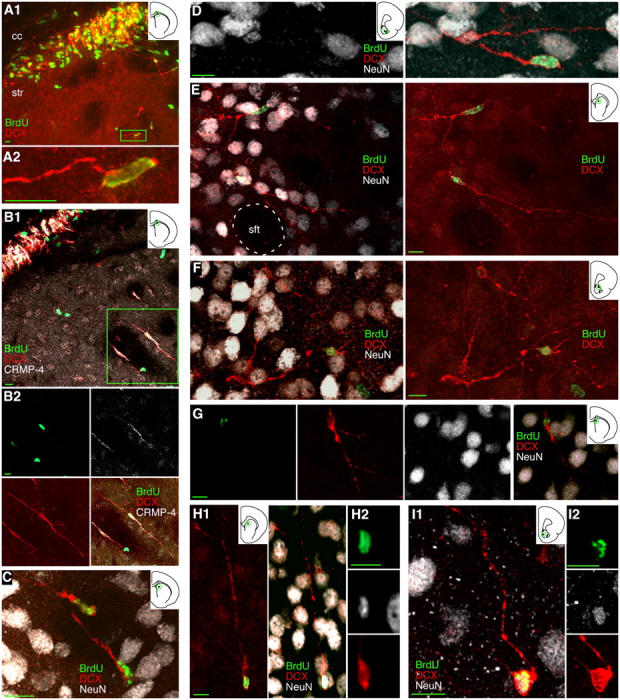
Young neurons appear to migrate from the adult SVZ into the striatum. (A and B) A band of DCX+ and CRMP4+ 1–2-wk-old BrdU+ neurons appears in the extension of the SVZ between the striatum (str) and the corpus callosum (cc). Scattered immature neurons can be seen outside of this band, either within the striatal matrix as the cell boxed in A1 or in a striatal fiber tract as one of the boxed cells in B1. (C) 1–2-wk-old BrdU+/DCX+ neurons in a striatal fiber tract. (D) A 1–2-wk-old BrdU+/DCX+ neuron in the core of the nucleus accumbens. (E and F) 3–4-wk-old BrdU+/DCX+ neurons extending complex processes are seen in the striatal matrix outside striatal fiber tracts (sft) as in E and in the shell of the nucleus accumbens as in F. (G–I) 3–4-wk-old BrdU+/DCX+/NeuN+ neurons can be found in the striatum as in G and H as well as in the shell of the nucleus accumbens as in I. Images show Z-axis projections of 12 × 0.90 μm (A1); 16 × 0.85 μm (A2); 18 × 0.72 μm (B1), 30 × 0.91 μm (B2), 18 × 1.44 μm (C), 16 × 0.78 μm (D), 36 × 0.48 μm (E), 20 × 0.98 μm (F); 14 × 1.04 μm (G); 30 × 0.78 μm (H1), 11 × 0.78 μm (H2), 16 × 0.62 μm (I1 and I2). Bars: 5 μm in orthogonal views; 10 μm in all other views.
Figure 7.
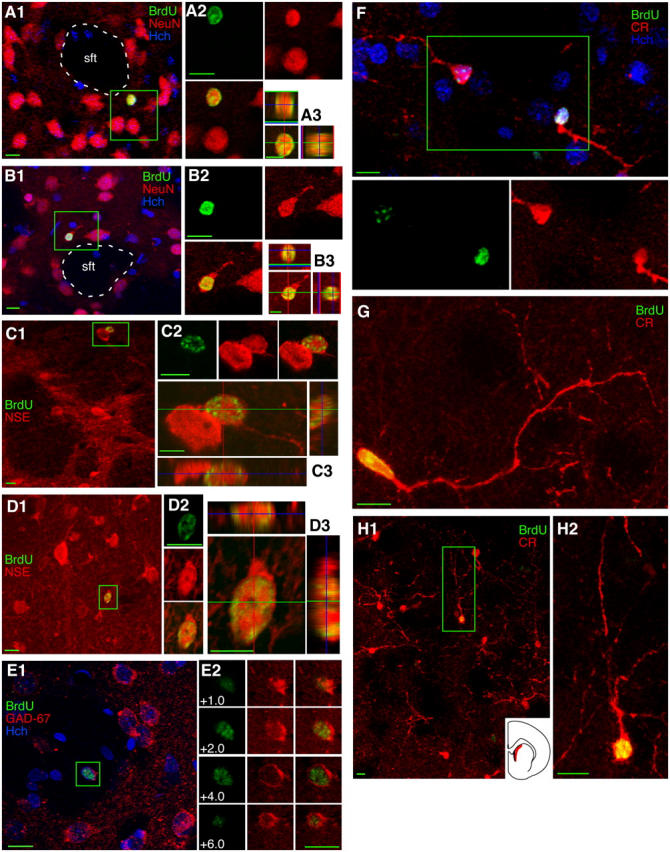
BrdU-labeled cells in the adult striatum express markers of GABAergic interneurons. (A) A 4–5-wk-old BrdU+ cell expressing the neuronal marker NeuN in the striatum just outside a striatal fiber tract (sft) is seen in the box in A1, in color separations in A2, and in orthogonal views in A3. (B) An 11–12-wk-old striatal BrdU+/NeuN+ neuron is shown in the same three formats as the cell in A. (C and D) 11–12-wk-old striatal BrdU+/NSE+ neurons were seen near the border of the white matter, as the boxed cell in C1, or deeper within the striatum, as the boxed cell in D1. (E) An 11–12-wk-old BrdU+/GAD67+ neuron in the striatum is shown with surrounding neurons in E1 and in color and z-plane separations in E2. (F–H) 4–5-wk-old BrdU+/CR+ neurons were found in the striatum, and nearly all newborn striatal neurons were found in a region known to have a high density of CR+ neurons, seen in red on the section outline (H1, inset). Images show single focal planes (A1, A2, B2, C1, C2, D2) or Z-axis projections of 10 × 1.02 μm (A3), 10 × 0.92 μm (B1, B3), 10 × 0.65 μm (C3), 6 × 0.71 μm (D1, D3), 7 × 1.20 μm (E1), 18 × 0.74 μm (F), 20 × 0.63 μm (G), 14 × 2.63 μm (H1), 18 × 0.69 μm (H2). Bars: 5 μm in orthogonal views; 10 μm in all other views.
BrdU+/GAD67+ and BrdU+/CR+ cells were both observed 4–5 wk after BrdU injection (Fig. 7, E–H), suggesting that newborn neurons in the striatum were GABAergic interneurons as in the neocortex. No double labeling of 4–5-wk-old BrdU+ cells was found with CB, parvalbumin, or somatostatin.
The vast majority of the new neurons in the striatum, identified with NeuN, GAD-67, or CR, were found either in the nucleus accumbens or within a restricted dorsomedial region of the striatum, within 650 μm of the lateral edge of the SVZ, ∼10–15 times the width of the SVZ (Fig. 7 H, inset). This portion of the striatum has been found to contain large numbers of CR+ interneurons and is believed to be an associative region of the striatum, corresponding to the caudate region in nonrodents (Riedel et al., 2002). 36.4 ± 7.0 BrdU+/NeuN+ cells/mm3 (5.8 ± 0.9% of 995 BrdU+ cells) were counted in the dorsomedial striatum at this time point. The location, immunoreactivity, and nuclear diameter (5–10 μm) of these BrdU+ cells were all consistent with striatal CR+ interneurons (Riedel et al., 2002).
Discussion
Similarity to other neurons born in adulthood
The findings of this study suggest that new neurons are generated in the adult rat neocortex and striatum. The relatively small, 5–10 μm, nuclear diameter of these new neurons is strikingly similar to that of granule cells in the dentate gyrus and olfactory bulb, two neuronal populations widely accepted as being generated in adulthood. Although the size of BrdU+/NeuN+ cells was not stated in three recent studies of adult cortical and striatal neurogenesis, the examples shown in these reports also had nuclear diameters in this same range (Magavi et al., 2000; Gould et al., 2001; Bedard et al., 2002), as did the cells in a much earlier electron microscopy study (Kaplan, 1981). The findings of these previous studies therefore appear consistent with the idea that the new neurons in the neocortex are nonpyramidal, although the techniques used did not allow identification of specific classes of neurons. In the current study, immunoreactivity for GABA/GAD-67 and calcium-binding proteins provide evidence that new cortical and striatal cells are in fact GABAergic interneurons. Distinct functional and morphological groups of GABAergic interneurons can be identified by their expression of particular calcium-binding proteins (Gabbott et al., 1997). These groups can be further subdivided into classes based on size and shape; eight classes of CB+ interneurons and five classes of CR+ interneurons have been described in the rat neocortex (Gabbott et al., 1997). Class 7 CB+ interneurons are small, with cell body diameters of 8–14 μm, and are found in cortical layers 5 and 6 (Gabbott et al., 1997), which is consistent with cortical BrdU+/CB+ cells observed in the current study. Based on their location and shape, the cortical CR+ cells in the current study appear to be class 4 CR+ interneurons (Gabbott et al., 1997); although no specific size range was given for this particular class by Gabbott et al. (1997) the entire CR+ population had relatively small cell bodies, 6–13 μm in diameter. Dentate gyrus granule cells born in the adult undergo a transition from early CR expression to later CB expression (Brandt et al., 2003), suggesting that these two classes of new interneurons found in the adult cortex might instead represent two stages of maturation of a single class of interneurons. The immunoreactivity and location of the new striatal neurons within a small region of the dorsolateral striatum and the nucleus accumbens match closely with descriptions of striatal CR+ interneurons (Riedel et al., 2002); striatal interneurons have not been subdivided into multiple subclasses expressing each calcium-binding protein.
The observed density of BrdU+/NeuN+ cells in the adult rat neocortex is remarkably consistent with that previously found in macaque cortex (Gould et al., 1999b, 2001) but seems low relative to what is seen in the adult dentate gyrus, where new neurons are known to have functional effects (Shors et al., 2001). However, the total population of dentate gyrus granule cells is very large compared with that of cortical or striatal interneurons in these classes. Additionally, the volume of the dentate gyrus granule cell layer is small compared with that of the cortex or striatum. Because these cortical and striatal interneuron populations are so small and spread out compared with granule cells, lower densities of newborn neurons do not necessarily indicate a significantly lower rate of neuron replacement or addition. Based on the differences in population size and volume, the cortex would need 0.9 BrdU+/NeuN+ cells/mm3 and the striatum would need 1.3 BrdU+/NeuN+ cells/mm3 to have a turnover rate equivalent to the 850 BrdU+/NeuN+ cells/mm3 in the adult rat dentate gyrus (Table SI, available at http://www.jcb.org/cgi/content/full/jcb.200407053/DC1). The current data suggest that one BrdU injection would label 0.8 BrdU+/NeuN+ cells/mm3 in the deep neocortex and 9.1 BrdU+/NeuN+ cells/mm3 in the striatum (Table SI), indicating that the turnover rates in the neocortex and striatum may in fact be very similar to that in the dentate gyrus.
Technical considerations
Our results are unlikely to result from BrdU labeling of neurons undergoing DNA repair or abortive mitosis for several reasons. First, BrdU immunostaining, using a staining protocol nearly identical to that used here, is very specific for dividing cells even after higher intraperitoneal doses of BrdU or induction of very high levels of DNA repair (Parent et al., 1999; Palmer et al., 2000; Cameron and McKay, 2001). Second, if cells undergoing DNA repair incorporated detectable amounts of BrdU, BrdU labeling should have been observed in cells of all types, including the very numerous pyramidal neurons. Third, any neurons undergoing abortive mitosis should be cleared away within three days (Hu et al., 1997; Thomaidou et al., 1997), i.e., long before the 4–5 wk and 11–12 wk time points at which we observed double-labeled cells. Fourth, BrdU+ cells showed evidence of maturation of their morphology and marker expression, including late expression of NSE and GAD-67 and changes in the morphology and location of double-labeled cells within the striatum, rather than degeneration.
These findings are also unlikely to result from staining of nonneuronal cells by neuronal markers. Although no antibody should be viewed as a perfect marker of cell identity, BrdU+ cells were labeled with eight independent neuronal markers, i.e., DCX (in striatum), CRMP-4, NeuN, NSE, HuC/D, EAAC-1, GABA/GAD-67, CR, and CB (in cortex), in a time course consistent with neuronal differentiation during development and in other neurogenic regions in the adult brain. Because each of these markers is highly specific for neurons within the normal rat brain (Marangos and Schmechel, 1987; Mullen et al., 1992; Minturn et al., 1995; Rosier et al., 1995; Gabbott et al., 1997; Okano and Darnell, 1997; Gleeson et al., 1999; Danbolt, 2001; Wichterle et al., 2001), and because the probability of nonspecific labeling decreases exponentially with each additional marker, it is improbable that eight neuronal markers would aberrantly label newly born glial cells within the neocortex and striatum.
It is not clear why double-labeled cells were not found in the adult rodent and primate neocortex in some previous reports (Kornack and Rakic, 2001; Ehninger and Kempermann, 2003; Koketsu et al., 2003). It is possible that investigators focused on pyramidal cells, the large and easily recognized primary neurons that constitute nearly 70% of the neurons in the cortex (Douglas et al., 2004), and which were also not found to be double labeled in the current study. An additional issue is that nuclear NeuN fluorescence can be obscured by BrdU fluorescence if dual excitation filters are used to search for cells to scan with confocal lasers. This is not a concern in pyramidal cells, because they have a significant amount of NeuN+ perinuclear cytoplasm. However, the NeuN expression in small interneurons is restricted to the nucleus (Mullen et al., 1992), making them very difficult to detect using dual filters. The difficulty in determining the NeuN-immunoreactivity of satellite cells adds another possible obstacle. Similar problems could occur in examining the striatum, where it might be assumed that any cell body smaller than those of the medium spiny neurons belongs to a glial cell. In addition to these issues of methods or expectations, differences in the animals' environments could also potentially affect the ability to detect new neurons. Stress, learning, and housing conditions all have significant effects on neurogenesis in the dentate gyrus (Gould et al., 1997, 1999a; Kempermann et al., 1997b). Although it is not known whether these factors also affect neocortical or striatal neurogenesis, the inhibition of cortical NG2+ cell proliferation by corticosteroids (Alonso, 2000) and the inter-animal variability observed in the current study suggest that they may.
Source of new neurons
Both the morphology of BrdU+/DCX+ cells seen in the current study and their change in location with increasing survival time are consistent with recent reports indicating that immature neurons or neuronal precursors in the SVZ do not all join the rostral migratory stream and may instead migrate into the striatum (Gould et al., 1999b, 2001; Nacher et al., 2001; Bernier et al., 2002; Luzzati et al., 2003; Yang et al., 2004). In the neocortex, however, the lack of DCX+ cells, pairing of many BrdU+ cells, and coexpression of NeuN and NG2 in some newborn cells suggest that new neocortical neurons may be generated in situ from NG2+ precursors rather than from precursors in the SVZ. This is somewhat surprising, because NG2 has commonly been considered to be a specific marker for oligodendrocyte precursors. However, several recent studies suggest that not all NG2+ cells are committed to this oligodendrocyte lineage. First, two distinct subpopulations of NG2+ cells that remain stable throughout development can be distinguished by the presence or absence of proteolipid protein expression (Mallon et al., 2002). Second, immature GABAergic neurons and NG2+ cells in the developing cortex both express dlx and appear to be derived from a common progenitor in the embryonic ventral forebrain (He et al., 2001). Third, and probably the strongest evidence, is that in the postnatal hippocampus and neocortex, some cells expressing CNPase and NG2 express neuronal markers (CRMP-4, class III β-tubulin, NeuN, and GAD-67), have spontaneous synaptic currents, and fire action potentials (Belachew et al., 2003; Aguirre et al., 2004; Chittajallu et al., 2004)—all indicators of a neuronal phenotype. These findings all provide support for the idea that GABAergic cortical neurons could be derived from the NG2+ cells that reside within the cortex itself. Although the current findings suggest that new neurons in the adult cortex and striatum have different immediate precursors in adulthood, they likely share a common ancestor: the dlx+ cells in the medial ganglionic eminence, which give rise to both cortical and striatal GABAergic interneurons during development (Marin et al., 2000; Wichterle et al., 2001). Together, these findings suggest that some medial ganglionic eminence-derived precursor cells residing in different regions of the brain during adulthood may retain their ability to divide and generate new GABAergic interneurons.
Materials and methods
Animal treatments
Adult male Sprague-Dawley rats (300–350 g, ∼9–10 wk old; Charles River or Taconic Labs) were housed two per cage under a 12:12 light/dark cycle with food and water ad libitum. All rats were injected with BrdU (Boehringer-Mannheim; 300 mg/kg body weight, 10 mg/ml in 0.007N NaOH/0.9% NaCl) and killed at different survival times after BrdU injection (Table I). Three rats were given a single BrdU injection and perfused 2 h later (2 h old BrdU+ cells); this short survival time labels cells that are still dividing (Cameron and McKay, 2001). Because previous studies indicate that the density of newborn neocortical neurons is very low compared with that of newborn dentate gyrus neurons (Gould et al., 2001), rats in all other groups received multiple BrdU injections to increase the numbers of labeled cells (Table I). All animal treatments conformed to NIH guidelines and were approved by the NIMH Institutional Animal Care and Use Committee.
Immunohistochemistry
At the selected survival time points, rats were deeply anesthetized with isoflurane inhalation and transcardially perfused with 4% PFA, pH 7.4. Brains were dissected from the skulls and fixed overnight in 4% PFA, pH 7.4. Coronal sections (50 μm) between Bregma 0.48 and 4.20 mm (Paxinos and Watson, 1998) were cut on an Oscillating Tissue Slicer 4000 (Electron Microscopy Sciences).
Sections were immunofluorescently stained using the following standard procedure (exceptions are listed below): Free-floating sections were pretreated with 2 N HCl for 30 min to denature DNA; rinsed well in PBS to return pH to 7.4; incubated in blocking solution (PBS, pH 7.4, with 0.5% Tween-20 and 3% donkey or goat normal serum, depending on the secondary antibodies) for 20 min at RT; incubated in a combination of primary antibodies (Table III) in blocking serum at 4°C for 48–72 h; washed in PBS; incubated with secondary antibodies against the appropriate species (Table III); counterstained with Hoechst 33258 (bisbenzimide; Molecular Probes); and coverslipped with aqueous hardening mounting medium (Immu-mount, Shandon). Sections stained with rat anti-BrdU antibody were heated at 90°C in citric acid for 25 min (0.1 M, pH 6.0) before HCl treatment. Staining for NSE, GABA or GAD-67 combined with BrdU was performed in two steps: sections were first processed for marker staining (through primary and secondary antibodies) before proceeding to HCl treatment and anti-BrdU staining. DMSO (0.3% vol/vol) was used instead of Tween-20 for GAD-67 staining.
Data analysis
Fluorescently immunolabeled sections were analyzed on a Meta confocal microscope (model LSM 510; Carl Zeiss MicroImaging, Inc.) equipped with three lasers (Argon 488, HeNe 543, and HeNe 633) and a coherent laser set at 720 nm for 2 photon UV excitation. Each channel was separately scanned using a multitrack PMT configuration to avoid cross-talk between fluorescent labels. All secondary antibody combinations were carefully examined to ensure that there was no cross-talk between fluorescent dyes or cross-reactivity between secondary antibodies. To evaluate double labeling, confocal Z sectioning was performed at 0.5–1.0-μm intervals using Plan-Neofluar 40× oil-immersion (NA = 1.30), or Plan-Apochromat 63× oil-immersion (NA = 1.40) objectives. To see labeled structures in relation to other cells, Z sectioning at wider intervals was done at 40× as above, or at 25× (Plan-Neofluar, multi-immersion, NA = 0.8). Images were acquired and three-dimensionally reconstructed using the Zeiss LSM software, cropped and optimized in Photoshop 7.0 by making minor adjustments to contrast, and arranged with line drawings in Canvas 8.0.
The proportion of 4–5-wk-old BrdU+ cells in the cortex that were immunoreactive for NeuN and/or NG2 was determined in one half-section, i.e., the right or left half of a coronal section, (Bregma 1.20 mm) from each of 10 brains. All BrdU+ cells located with a single row of 400× magnification optical fields (500 μm in diameter in a 50-μm-thick section) were systematically scanned to determine NeuN and NG2 immunoreactivity–starting at the midline dorsal, to the corpus callosum, and moving laterally through the deep cortical layers with the bottom edge of the field at the dorsal border of the white matter–scanning adjacent fields until the border between the white matter and cortex became difficult to establish with sufficient clarity (14–15 fields). The total volume analyzed was 14.7 fields × 3.14 × 0.25 mm × 0.25 mm × 0.05 mm = 0.14 mm3 in each of 10 brains. Each optical field extended through layers VIa and VIb, and part of layer V, depending on the cortical subregion. The scanned region of interest contained the cingulate cortex, secondary motor cortex, primary motor cortex, and primary somatosensory cortex, and ended partway through the dorsal portion of the insular claustrum. The claustrum has developmental and phylogenetic origins very close to cortex (Kowianski et al., 1999; Reblet et al., 2002) despite its past classification in some texts as part of the basal ganglia. An additional 3.5 sections (from Bregma 1.2 to 3.2 mm; Paxinos and Watson, 1998) per brain were analyzed using the same method to define the region of interest (94.2 ± 3.2 optical fields per brain; 0.92 mm3 total volume per brain). This region contained the same cortical areas as the initial sections, with the addition of the prelimbic and infralimbic cortices (Paxinos and Watson, 1998). These additional sections were first surveyed on an epifluorescence microscope equipped with separate filters for rhodamine, FITC, and DAPI. Any BrdU+ cells that appeared to be NeuN+ were then scanned on the confocal microscope to confirm NeuN immunoreactivity and determine NG2 immunoreactivity. The proportion of 2-h-old BrdU+ cells in the neocortex that expressed NeuN and/or NG2 was quantified in one section (Bregma 1.20 mm) from each brain (28 optical fields per brain).
For quantification of striatal neurogenesis, the percentage of BrdU+ cells that were NeuN+ was determined in the dorsomedial region of the striatum near the SVZ, where most BrdU+ cells double labeled with neuronal markers were found in qualitative examination. All 4–5-wk-old BrdU+ cells located within a single row of five 400× magnification optical fields in each of two sections (20 optical fields per brain between Bregma 1.60 and 1.20 mm) were surveyed on an epifluorescence microscope, starting at the ventral edge of the SVZ and progressing dorsolaterally keeping one edge of the field at the border of the SVZ. Only BrdU+ and BrdU+/NeuN+ cells located within the striatum were counted, i.e., BrdU+ cells touching the SVZ were not included.
Online supplemental material
Table S1 shows a comparison of the replacement rates of newborn neurons in the adult rat cortex, striatum, and dentate gyrus based on BrdU+/NeuN+ cell densities found in the current study and neuronal population sizes, region volumes, and dentate gyrus BrdU+/NeuN+ cell density from other studies. Online supplemental material is available at http://www.jcb.org/cgi/content/full/jcb.200407053/DC1.
Acknowledgments
This article was funded by the NIMH Division of Intramural Research. A.G. Dayer was supported by the Swiss National Fund.
Abbreviations used in this paper: +, immunoreactive; −, nonimmunoreactive; CB, calbindin; CR, calretinin; DCX, doublecortin; GABA, γ-aminobutyric acid; GAD-67, glutamic acid decarboxylase 67; NSE, neuron-specific enolase; SVZ, subventricular zone.
References
- Aguirre, A.A., R. Chittajallu, S. Belachew, and V. Gallo. 2004. NG2-expressing cells in the subventricular zone are type C-like cells and contribute to interneuron generation in the postnatal hippocampus. J. Cell Biol. 165:575–589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alonso, G. 2000. Prolonged corticosterone treatment of adult rats inhibits the proliferation of oligodendrocyte progenitors present throughout white and gray matter regions of the brain. Glia. 31:219–231. [DOI] [PubMed] [Google Scholar]
- Altman, J. 1962. Are new neurons formed in the brains of adult mammals? Science. 135:1127–1128. [DOI] [PubMed] [Google Scholar]
- Bedard, A., M. Cossette, M. Levesque, and A. Parent. 2002. Proliferating cells can differentiate into neurons in the striatum of normal adult monkey. Neurosci. Lett. 328:213–216. [DOI] [PubMed] [Google Scholar]
- Belachew, S., R. Chittajallu, A.A. Aguirre, X. Yuan, M. Kirby, S. Anderson, and V. Gallo. 2003. Postnatal NG2 proteoglycan-expressing progenitor cells are intrinsically multipotent and generate functional neurons. J. Cell Biol. 161:169–186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benraiss, A., E. Chmielnicki, K. Lerner, D. Roh, and S.A. Goldman. 2001. Adenoviral brain-derived neurotrophic factor induces both neostriatal and olfactory neuronal recruitment from endogenous progenitor cells in the adult forebrain. J. Neurosci. 21:6718–6731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernier, P.J., A. Bedard, J. Vinet, M. Levesque, and A. Parent. 2002. Newly generated neurons in the amygdala and adjoining cortex of adult primates. Proc. Natl. Acad. Sci. USA. 99:11464–11469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brandt, M.D., S. Jessberger, B. Steiner, G. Kronenberg, K. Reuter, A. Bick-Sander, W. von der Behrens, and G. Kempermann. 2003. Transient calretinin expression defines early postmitotic step of neuronal differentiation in adult hippocampal neurogenesis of mice. Mol. Cell. Neurosci. 24:603–613. [DOI] [PubMed] [Google Scholar]
- Cameron, H.A., and R.D. McKay. 2001. Adult neurogenesis produces a large pool of new granule cells in the dentate gyrus. J. Comp. Neurol. 435:406–417. [DOI] [PubMed] [Google Scholar]
- Chittajallu, R., A.A. Aguirre, and V. Gallo. 2004. NG2-positive cells in the white and grey matter display distinct physiological properties. J. Physiol. 561:109–122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conti, F., S. DeBiasi, A. Minelli, J.D. Rothstein, and M. Melone. 1998. EAAC1, a high-affinity glutamate tranporter, is localized to astrocytes and gabaergic neurons besides pyramidal cells in the rat cerebral cortex. Cereb. Cortex. 8:108–116. [DOI] [PubMed] [Google Scholar]
- Danbolt, N.C. 2001. Glutamate uptake. Prog. Neurobiol. 65:1–105. [DOI] [PubMed] [Google Scholar]
- Dayer, A.G., A.A. Ford, K.M. Cleaver, M. Yassaee, and H.A. Cameron. 2003. Short-term and long-term survival of new neurons in the rat dentate gyrus. J. Comp. Neurol. 460:563–572. [DOI] [PubMed] [Google Scholar]
- Douglas, R., H. Markram, and K. Martin. 2004. Neocortex. The Synaptic Organization of the Brain. G.M. Shepherd, editor. Oxford University Press, New York. 499–558.
- Ehninger, D., and G. Kempermann. 2003. Regional effects of wheel running and environmental enrichment on cell genesis and microglia proliferation in the adult murine neocortex. Cereb. Cortex. 13:845–851. [DOI] [PubMed] [Google Scholar]
- Gabbott, P.L., and S.J. Bacon. 1996. Local circuit neurons in the medial prefrontal cortex (areas 24a,b,c, 25 and 32) in the monkey: I. Cell morphology and morphometrics. J. Comp. Neurol. 364:567–608. [DOI] [PubMed] [Google Scholar]
- Gabbott, P.L., B.G. Dickie, R.R. Vaid, A.J. Headlam, and S.J. Bacon. 1997. Local-circuit neurones in the medial prefrontal cortex (areas 25, 32 and 24b) in the rat: morphology and quantitative distribution. J. Comp. Neurol. 377:465–499. [DOI] [PubMed] [Google Scholar]
- Gleeson, J.G., P.T. Lin, L.A. Flanagan, and C.A. Walsh. 1999. Doublecortin is a microtubule-associated protein and is expressed widely by migrating neurons. Neuron. 23:257–271. [DOI] [PubMed] [Google Scholar]
- Gould, E., B.S. McEwen, P. Tanapat, L.A. Galea, and E. Fuchs. 1997. Neurogenesis in the dentate gyrus of the adult tree shrew is regulated by psychosocial stress and NMDA receptor activation. J. Neurosci. 17:2492–2498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gould, E., A. Beylin, P. Tanapat, A. Reeves, and T.J. Shors. 1999. a. Learning enhances adult neurogenesis in the hippocampal formation. Nat. Neurosci. 2:260–265. [DOI] [PubMed] [Google Scholar]
- Gould, E., A.J. Reeves, M.S. Graziano, and C.G. Gross. 1999. b. Neurogenesis in the neocortex of adult primates. Science. 286:548–552. [DOI] [PubMed] [Google Scholar]
- Gould, E., N. Vail, M. Wagers, and C.G. Gross. 2001. Adult-generated hippocampal and neocortical neurons in macaques have a transient existence. Proc. Natl. Acad. Sci. USA. 98:10910–10917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu, W., T. Brannstrom, and P. Wester. 2000. Cortical neurogenesis in adult rats after reversible photothrombotic stroke. J. Cereb. Blood Flow Metab. 20:1166–1173. [DOI] [PubMed] [Google Scholar]
- He, W., C. Ingraham, L. Rising, S. Goderie, and S. Temple. 2001. Multipotent stem cells from the mouse basal forebrain contribute GABAergic neurons and oligodendrocytes to the cerebral cortex during embryogenesis. J. Neurosci. 21:8854–8862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hendrickson, A.E., N.J. Tillakaratne, R.D. Mehra, M. Esclapez, A. Erickson, L. Vician, and A.J. Tobin. 1994. Differential localization of two glutamic acid decarboxylases (GAD65 and GAD67) in adult monkey visual cortex. J. Comp. Neurol. 343:566–581. [DOI] [PubMed] [Google Scholar]
- Hu, Z., K. Yuri, H. Ozawa, H. Lu, and M. Kawata. 1997. The in vivo time course for elimination of adrenalectomy-induced apoptotic profiles from the granule cell layer of the rat hippocampus. J. Neurosci. 17:3981–3989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imai, Y., I. Ibata, D. Ito, K. Ohsawa, and S. Kohsaka. 1996. A novel gene iba1 in the major histocompatibility complex class III region encoding an EF hand protein expressed in a monocytic lineage. Biochem. Biophys. Res. Commun. 224:855–862. [DOI] [PubMed] [Google Scholar]
- Jiang, W., W. Gu, T. Brannstrom, R. Rosqvist, and P. Wester. 2001. Cortical neurogenesis in adult rats after transient middle cerebral artery occlusion. Stroke. 32:1201–1207. [DOI] [PubMed] [Google Scholar]
- Kaplan, M.S. 1981. Neurogenesis in the 3-month-old rat visual cortex. J. Comp. Neurol. 195:323–338. [DOI] [PubMed] [Google Scholar]
- Kempermann, G., H.G. Kuhn, and F.H. Gage. 1997. a. Genetic influence on neurogenesis in the dentate gyrus of adult mice. Proc. Natl. Acad. Sci. USA. 94:10409–10414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kempermann, G., H.G. Kuhn, and F.H. Gage. 1997. b. More hippocampal neurons in adult mice living in an enriched environment. Nature. 386:493–495. [DOI] [PubMed] [Google Scholar]
- Koketsu, D., A. Mikami, Y. Miyamoto, and T. Hisatsune. 2003. Nonrenewal of neurons in the cerebral neocortex of adult macaque monkeys. J. Neurosci. 23:937–942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kornack, D.R., and P. Rakic. 2001. Cell proliferation without neurogenesis in adult primate neocortex. Science. 294:2127–2130. [DOI] [PubMed] [Google Scholar]
- Kowianski, P., J. Dziewiatkowski, J. Kowianska, and J. Morys. 1999. Comparative anatomy of the claustrum in selected species: A morphometric analysis. Brain Behav. Evol. 53:44–54. [DOI] [PubMed] [Google Scholar]
- Luzzati, F., P. Peretto, P. Aimar, G. Ponti, A. Fasolo, and L. Bonfanti. 2003. Glia-independent chains of neuroblasts through the subcortical parenchyma of the adult rabbit brain. Proc. Natl. Acad. Sci. USA. 100:13036–13041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magavi, S.S., B.R. Leavitt, and J.D. Macklis. 2000. Induction of neurogenesis in the neocortex of adult mice. Nature. 405:951–955. [DOI] [PubMed] [Google Scholar]
- Mallon, B.S., H.E. Shick, G.J. Kidd, and W.B. Macklin. 2002. Proteolipid promoter activity distinguishes two populations of NG2-positive cells throughout neonatal cortical development. J. Neurosci. 22:876–885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marangos, P.J., and D.E. Schmechel. 1987. Neuron specific enolase, a clinically useful marker for neurons and neuroendocrine cells. Annu. Rev. Neurosci. 10:269–295. [DOI] [PubMed] [Google Scholar]
- Marin, O., S.A. Anderson, and J.L. Rubenstein. 2000. Origin and molecular specification of striatal interneurons. J. Neurosci. 20:6063–6076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minturn, J.E., D.H. Geschwind, H.J. Fryer, and S. Hockfield. 1995. Early postmitotic neurons transiently express TOAD-64, a neural specific protein. J. Comp. Neurol. 355:369–379. [DOI] [PubMed] [Google Scholar]
- Mullen, R.J., C.R. Buck, and A.M. Smith. 1992. NeuN, a neuronal specific nuclear protein in vertebrates. Development. 116:201–211. [DOI] [PubMed] [Google Scholar]
- Nacher, J., C. Crespo, and B.S. McEwen. 2001. Doublecortin expression in the adult rat telencephalon. Eur. J. Neurosci. 14:629–644. [DOI] [PubMed] [Google Scholar]
- Okano, H.J., and R.B. Darnell. 1997. A hierarchy of Hu RNA binding proteins in developing and adult neurons. J. Neurosci. 17:3024–3037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmer, T.D., A.R. Willhoite, and F.H. Gage. 2000. Vascular niche for adult hippocampal neurogenesis. J. Comp. Neurol. 425:479–494. [DOI] [PubMed] [Google Scholar]
- Parent, J.M., E. Tada, J.R. Fike, and D.H. Lowenstein. 1999. Inhibition of dentate granule cell neurogenesis with brain irradiation does not prevent seizure-induced mossy fiber synaptic reorganization in the rat. J. Neurosci. 19:4508–4519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paxinos, G., and C. Watson. 1998. The Rat Brain in Stereotaxic Coordinates. Academic Press, Inc., San Diego. 256 pp.
- Pencea, V., K.D. Bingaman, S.J. Wiegand, and M.B. Luskin. 2001. Infusion of brain-derived neurotrophic factor into the lateral ventricle of the adult rat leads to new neurons in the parenchyma of the striatum, septum, thalamus, and hypothalamus. J. Neurosci. 21:6706–6717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quinn, C.C., G.E. Gray, and S. Hockfield. 1999. A family of proteins implicated in axon guidance and outgrowth. J. Neurobiol. 41:158–164. [PubMed] [Google Scholar]
- Rakic, P. 2002. Neurogenesis in adult primate neocortex: an evaluation of the evidence. Nat. Rev. Neurosci. 3:65–71. [DOI] [PubMed] [Google Scholar]
- Reblet, C., A. Alejo, R.I. Blanco-Santiago, J. Mendizabal-Zubiaga, M. Fuentes, and J.L. Bueno-Lopez. 2002. Neuroepithelial origin of the insular and endopiriform parts of the claustrum. Brain Res. Bull. 57:495–497. [DOI] [PubMed] [Google Scholar]
- Riedel, A., W. Hartig, G. Seeger, U. Gartner, K. Brauer, and T. Arendt. 2002. Principles of rat subcortical forebrain organization: a study using histological techniques and multiple fluorescence labeling. J. Chem. Neuroanat. 23:75–104. [DOI] [PubMed] [Google Scholar]
- Rosier, A.M., L. Arckens, H. Demeulemeester, G.A. Orban, U.T. Eysel, Y.J. Wu, and F. Vandesande. 1995. Effect of sensory deafferentation on immunoreactivity of GABAergic cells and on GABA receptors in the adult cat visual cortex. J. Comp. Neurol. 359:476–489. [DOI] [PubMed] [Google Scholar]
- Shors, T.J., G. Miesegaes, A. Beylin, M. Zhao, T. Rydel, and E. Gould. 2001. Neurogenesis in the adult is involved in the formation of trace memories. Nature. 410:372–376. [DOI] [PubMed] [Google Scholar]
- Teramoto, T., J. Qiu, J.C. Plumier, and M.A. Moskowitz. 2003. EGF amplifies the replacement of parvalbumin-expressing striatal interneurons after ischemia. J. Clin. Invest. 111:1125–1132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomaidou, D., M.C. Mione, J.F. Cavanagh, and J.G. Parnavelas. 1997. Apoptosis and its relation to the cell cycle in the developing cerebral cortex. J. Neurosci. 17:1075–1085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vives, V., G. Alonso, A.C. Solal, D. Joubert, and C. Legraverend. 2003. Visualization of S100B-positive neurons and glia in the central nervous system of EGFP transgenic mice. J. Comp. Neurol. 457:404–419. [DOI] [PubMed] [Google Scholar]
- Wichterle, H., D.H. Turnbull, S. Nery, G. Fishell, and A. Alvarez-Buylla. 2001. In utero fate mapping reveals distinct migratory pathways and fates of neurons born in the mammalian basal forebrain. Development. 128:3759–3771. [DOI] [PubMed] [Google Scholar]
- Yang, H.K., N.L. Sundholm-Peters, G.E. Goings, A.S. Walker, K. Hyland, and F.G. Szele. 2004. Distribution of doublecortin expressing cells near the lateral ventricles in the adult mouse brain. J. Neurosci. Res. 76:282–295. [DOI] [PubMed] [Google Scholar]