

Abstract
Recent work has provided new insights into the mechanism of spindle assembly. Growing evidence supports a model in which the small GTPase Ran plays a central role in this process. Here, we examine the evidence for the existence of a RanGTP gradient around mitotic chromosomes and some controversial data on the role that chromosomes play in spindle assembly. We review the current knowledge on the Ran downstream targets for spindle assembly and we focus on the multiple roles of TPX2, one of the targets of RanGTP during cell division.
A central goal of cell division in eukaryotes is to separate two identical copies of the DNA and evenly distribute them to the daughter cells. This task is performed by microtubules that assemble into a spindle-shaped apparatus around the chromosomes. Substantial evidence coming from several labs supports a model in which the small GTPase Ran in its GTP-bound form (RanGTP) coordinates spatially spindle assembly. A number of recent excellent reviews cover different aspects of Ran functions in spindle assembly and in other cellular processes (Clarke and Zhang, 2001; Moore, 2001; Dasso, 2002; Hetzer et al., 2002; Quimby and Dasso, 2003; Weis, 2003; Di Fiore et al., 2004). Here, we examine some controversial data on the role of chromosomes and the Ran model in spindle assembly. We then review our current knowledge on the downstream targets of Ran during mitosis and we focus on the multiple functions performed by the targeting protein for Xklp2 (TPX2), one central target of Ran in spindle assembly.
General mechanism of Ran action during the cell cycle: a small protein with many tricks
The function of Ran in nucleocytoplasmic transport during interphase has been extensively studied. It relies primarily on the restriction of high concentrations of its GTP form to the nucleus. The differential distribution of RanGTP between the nucleus and the cytoplasm is safeguarded by the integrity of the nuclear envelope. In the nucleus, RanGTP regulates the binding properties of the family of importin β–like nuclear transport receptors, promoting the dissociation of import receptors from their cargo proteins and participating in complex formation of export receptors with their cargos. This governs the directionality of nucleocytoplasmic transport (Mattaj and Englmeier, 1998; Görlich and Kutay, 1999). There is now substantial evidence indicating that these activities are important for spindle assembly as well. Experiments in Xenopus egg extracts have shown that the dissociation of import receptors from their cargos by RanGTP is one of the mechanisms that renders several proteins competent to function in spindle formation (Gruss et al., 2001; Nachury et al., 2001; Wiese et al., 2001). Recently, there have been some direct (Askjaer et al., 2002; Bamba et al., 2002) and indirect evidence supporting a role for the Ran system in spindle assembly in mitosis in vivo (Gruss et al., 2002; Moore et al., 2002; Trieselmann and Wilde, 2002; Li et al., 2003). In addition, the Ran system has been demonstrated to be involved in other cell cycle regulated events, such as nuclear envelope assembly (Hetzer et al., 2000; Zhang and Clarke, 2001) and kinetochore function (Arnaoutov and Dasso, 2003). Moreover, Yamaguchi and Newport (2003) have recently shown that in the presence of high levels of RanGTP the export receptor CRM1 inhibits rereplication by sequestering replication proteins inside the nucleus without exporting them to the cytoplasm. Interestingly, this situation is like a mirror image of mitosis when Ran releases proteins from inhibitory import receptors.
A gradient of RanGTP in mitosis?
As the cell enters mitosis and the nuclear envelope breaks down the nucleoplasm and the cytoplasm mix, diluting the high concentration of nuclear RanGTP. However, because RCC1, the GEF for Ran associates with the chromosomes in M-phase, the concentration of RanGTP may still be high enough in the vicinity of chromatin to dissociate complexes of importins and cargo proteins. A local high RanGTP concentration may therefore provide positional information for the formation of the spindle around the chromosomes. To test this idea Kalab et al. (2002) used FRET-based biosensors to monitor the binding of RanGTP to its partners in Xenopus egg extracts and visualized a high local concentration of RanGTP around the mitotic chromatin, providing the first experimental support for the RanGTP gradient model for spindle assembly. However, it was not clear whether this only occurred in a large cytoplasm like the Xenopus egg or if such a gradient could also exist in somatic cells. Indeed, mathematical modeling based on known kinetic parameters of the components of the Ran system (Görlich et al., 2003) predicted that a gradient of RanGTP could exist around chromosomes in a large cytoplasm but not in an average 20-μm large somatic cell. Recently, Li and Zheng (2004) have provided evidence for the existence of a gradient of RanGTP around chromosomes in somatic cells. Therefore, there is now evidence supporting the existence of a gradient in different systems. It still remains to be determined how the localization of various components of the Ran system to specific cellular compartments influence this gradient. Indeed it has been reported that Ran localizes to the chromatin (Bilbao-Cortes et al., 2002; Li and Zheng, 2004), the kinetochores (Bamba et al., 2002), the spindle (Trieselmann and Wilde, 2002) and with the centrosomes (Forgues et al., 2003; Keryer et al., 2003). In addition, RanGAP1 as well as RanBP2 colocalize with spindle microtubules and kinetochores (Joseph et al., 2002, 2004), and the export receptor CRM1 can associate with centrosomes (Forgues et al., 2003; Keryer et al., 2003). Whether and how the localization of Ran or components of the Ran system may affect the shape of the Ran gradient during mitosis and how different factors involved in spindle assembly react to a given RanGTP concentration remains to be determined.
Spindle assembly without chromosomes?
A strong prediction of the Ran gradient model is that chromosomes are essential determinants for spindle assembly. This idea was initially supported by experiments showing that injection of chromatin or DNA into Xenopus metaphase arrested eggs triggered spindle assembly whereas injection of centrosomes did not (Karsenti et al., 1984). Later experimental support came from the observation of spindle assembly around DNA-coated beads in Xenopus egg extracts (Heald et al., 1996). These results however posed the question of whether spindle assembly around chromatin could just be a specialized pathway existing in systems that naturally lack centrosomes, like in vertebrate female meiosis and in plants. Many observations argue against this simple interpretation. Inhibition of RanGTP production in Xenopus egg extracts leads to severe defects in spindle assembly even in the presence of centrosomes (Carazo-Salas et al., 2001). Other manipulations in different cellular systems (meiotic and mitotic) also support the view that chromosomes are important determinants for spindle assembly in general (Karsenti et al., 1984; McKim and Hawley, 1995; Zhang and Nicklas, 1995; Khodjakov et al., 2000).
However, reports on spindle formation in the absence of chromosomes have challenged this idea (Brunet et al., 1998; Faruki et al., 2002; Bucciarelli et al., 2003) and have led some authors to propose that the organization of microtubules into two antiparallel arrays is an intrinsic property of mitotic microtubules (Brunet et al., 1998).
Microtubules become highly dynamic in mitosis. It is conceivable that when two or more dynamic microtubule asters come into proximity in an M-phase cytoplasm, some antiparallel interactions may be established and become stabilized through the recruitment of some mitotic factors. Spindle-like structures have indeed been reported to form in the absence of chromosomes (Faruki et al., 2002; Bucciarelli et al., 2003). This is in apparent contradiction with the Ran gradient model. However, unlike the finely temporally and spatially controlled process of spindle formation around chromosomes, spindle-like structures formed in the absence of chromosomes arise randomly in the cellular space (Brunet et al., 1998) and are often unstable (Faruki et al., 2002).
Computer simulations have recently been used to analyze theoretically the minimal requirements for the establishment of stable antiparallel interactions between two microtubule asters (Nedelec, 2002). The results suggest that relatively few components may be sufficient. Thus, it may not be surprising that in a mitotic cytoplasm two or more microtubule asters may establish antiparallel interactions. One important question is then whether these structures could be as functional as real spindles.
Recently, Drosophila secondary spermatocytes lacking chromosomes have been reported to assemble a spindle and go through anaphase and cytokinesis like wild-type cells (Bucciarelli et al., 2003). This suggests that except for the obvious lack of chromosome segregation the spindle-like structure that formed retained the other functions attributed to spindles. In fact, a closer look at microtubule organization in these cells suggests that the two microtubule asters nucleated by the centrosomes barely interact. Because of the space restriction imposed by the cell membrane most microtubules nucleated by the centrosomes can only grow toward the center of the cell and thus adopt a spindle-like topology. However, the density of microtubules in the central region is much lower than in bona fide spindles suggesting that there are no interactions between antiparallel microtubules. Although the later assembly of the central spindle and cytokinesis could suggest that a spindle had indeed previously assembled, in fact these are most certainly independent events. First, in this meiotic system the spindle checkpoint is nonexistent or very weak (Rebollo and Gonzalez, 2000). Second, the central spindle does not derive from the spindle itself but forms from newly assembled microtubules that grow in the central cortical region of the cell (C. Gonzalez, personal communication). Moreover, using live imaging on dividing spermatocytes, Rebollo et al. (2004) have shown that in this system the plane of cytokinesis is not determined by the spindle itself but by the position of the two centrosomal asters. Interestingly, these authors have also shown that spindles can assemble in Drosophila spermatocytes without any contribution from centrosome nucleated microtubules (Rebollo et al., 2004). Finally, Canman et al. (2003) have demonstrated recently that contractile ring positioning in rat kangaroo cells can even occur in the presence of just one monopolar array of microtubules, indicating that bipolar organization of antiparallel microtubules is indeed not needed for cytokinesis in general. Therefore, these data suggest that these late events do not depend on the previous formation of a spindle. In summary, although some antiparallel interactions may be established between two or more microtubule asters in an M-phase cytoplasm, chromosomes appear as important determinants to direct spatially the formation of a robust spindle.
Targets of Ran during mitosis
During mitosis RanGTP regulates the activity of factors involved in microtubule nucleation, stabilization and organization (Wilde and Zheng, 1999; Zhang et al., 1999; Carazo-Salas et al., 2001; Wilde et al., 2001; Hetzer et al., 2002). Importin β was found to mediate the effect of RanGTP on mitotic factors. At present, we know of three proteins that bind to importin β via the adaptor protein importin α in mitosis: the kinesin-like protein (KLP) XCTK2 and two microtubule-associated proteins, targeting protein for Xklp2 (TPX2) and nuclear protein of the mitotic apparatus (NuMA). These proteins have been proposed to be targets of RanGTP in the spindle assembly pathway (Gruss et al., 2001; Nachury et al., 2001; Wiese et al., 2001; Ems-McClung et al., 2004; Table I). The predominant interacting partners of TPX2 and XCTK2 in Xenopus M-phase extracts are indeed importins α and β (Gruss et al., 2001; Ems-McClung et al., 2004). By contrast, the majority of NuMA is bound to the dynein–dynactin complex (Merdes et al., 1996) indicating that only a small subset of NuMA could be regulated by the Ran system through the binding to importins.
Table I.
Proteins regulated by RanGTP during spindle assembly a
| Protein | Function/activity | Localization | Activity regulated by RanGTP |
|---|---|---|---|
| TPX2a–j | MAP MT nucleation Spindle pole organization Targeting of Xklp2 and Aurora A Activation of Aurora A |
Spindle poles and spindle MTs Midbody in telophase Degraded in G1 |
Binding to importins MT nucleation Interaction with Aurora A Activation of Aurora A |
| NuMAk–m | MAP Spindle pole organization |
Spindle pole Nuclear matrix in interphase |
Binding to importins |
| XCTK2n,o | Kinesin-like protein Minus end–directed motility Organization of spindle poles |
Spindle poles and spindle MTs | Binding to importins Inhibitory effect of importins on MT binding of the nonmotor domain (assay with purified components) |
| Eg5p–r | Kinesin-like protein Homotetramer Plus end–directed motility Centrosome separation Antiparallel microtubule interactions Maintenance of spindle bipolarity |
Spindle poles and spindle MTs | MT organization |
| kide, s–v | Chromokinesin Plus end–directed motility Chromosome oscillations Metaphase chromosome alignment |
Chromatin, spindle MTs | Inhibitory effect of importins on MT binding |
| Aurora Af–j, w | Mitotic kinase Activated by TPX2 Known substrates: Eg5, TPX2, TACC |
Spindle MTs centrosome | Binding to TPX2 |
Wittmann et al., 2000; bGruss et al., 2001; cGruss et al., 2002; dGarrett et al., 2002; eTrieselmann et al., 2003; fKufer et al., 2002; gEyers et al., 2003; hTsai et al., 2003; iEyers and Maller, 2004; jBayliss et al., 2003; kMerdes et al., 1996; lNachury et al., 2001; mWiese et al., 2001; n Walczak et al., 1997; oEms-McClung et al., 2004; p Sawin et al., 1992; q Blangy et al., 1995; rWilde et al., 2001; s Antonio et al., 2000; t Funabiki and Murray, 2000; u Yajima et al., 2003; v Levesque and Compton, 2001; w Nigg, 2001.
The number of proteins regulated by Ran during cell division will probably increase rapidly. Likely candidate factors involved in cell division are those sequestered into the nucleus in interphase and thus interacting with importins. The microtubule binding capacity of the nuclear KLP kid was recently found to be modulated by Ran and the importins in vitro (Trieselmann et al., 2003; see Fig. 2, and Table I). In addition, other factors may be regulated in a more indirect manner. Wilde et al. (2001) have shown that the kinesin like protein Eg5 is regulated by RanGTP and involved in the organization of microtubules into “mini-spindles”, spindle-like structures unique to the Ran pathway because they do not form after addition of taxol or centrosomes to an M-phase egg extract. A general picture is starting to emerge in which RanGTP coordinates the activities of numerous factors that collectively direct the formation of the spindle around the chromosomes (Fig. 1 and Table I). However, TPX2 probably remains the best-characterized Ran target. Moreover, many recent data suggest that the regulation of TPX2 is important for several steps in spindle assembly. We will focus on TPX2 in the last part of this review.
Figure 1.
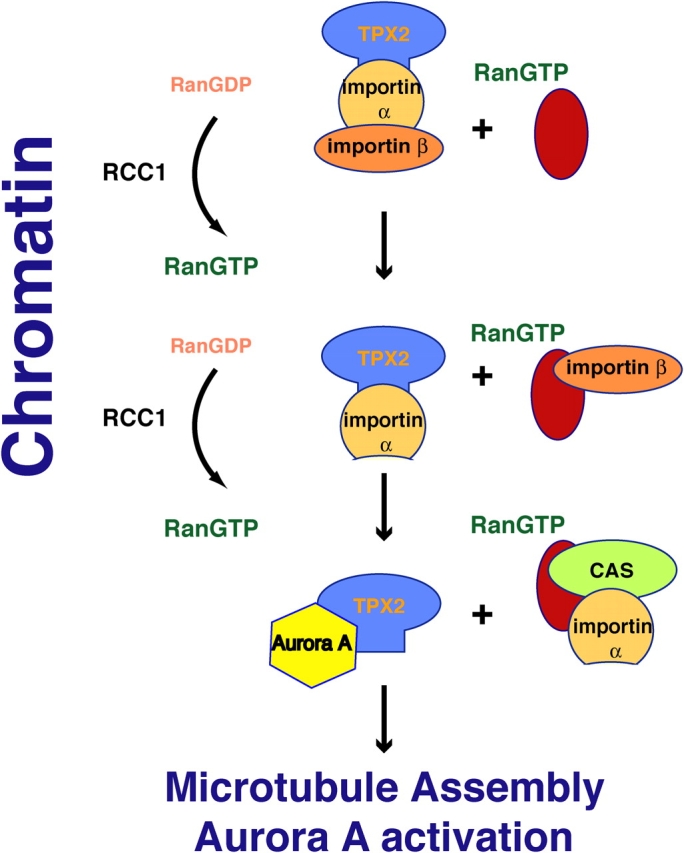
During mitosis RanGDP is converted into RanGTP in the proximity of the chromosomes by the nucleotide exchange factor RCC1. In a first step, RanGTP binds to importin β releasing it from importin α. The affinity of importin α for NLS sequences is reduced and a complex of importin α CAS and RanGTP forms releasing TPX2. CAS functions to reexport importin α to the cytoplasm. TPX2 triggers microtubule nucleation and binds to the mitotic kinase Aurora A activating it.
Role of TPX2 in RanGTP induced microtubule nucleation
Although TPX2 was first identified as an antigen specific for mitosis (Heidebrecht et al., 1997) and as the TPX2 (Wittmann et al., 2000), a major breakthrough came with the discovery that it is one of the targets of RanGTP during mitosis (Gruss et al., 2001). TPX2 is bound to importins α and β in Xenopus M-phase extracts. This complex is dissociated by RanGTP and once released TPX2 promotes microtubule assembly (Gruss et al., 2001; Fig. 2). How does TPX2 trigger microtubule assembly? TPX2 behaves as a microtubule associated protein (MAP) in all biochemical assays and it colocalizes with spindle microtubules in M-phase in vivo and in vitro (Heidebrecht et al., 1997; Wittmann et al., 2000; Garrett et al., 2002; Gruss et al., 2002; Trieselmann et al., 2003). However, TPX2 does not stabilize microtubules like other MAPs but probably plays a yet poorly understood role in the early steps of microtubule nucleation (Gruss et al., 2002). In fact, bacterially expressed TPX2 can initiate microtubule polymerization in a solution of pure tubulin, an activity regulated by importins (Schatz et al., 2003). Together, these data indicate strongly that an essential early role of TPX2 is to promote microtubule assembly in the vicinity of chromosomes. Accordingly, TPX2 has been found to be crucial for spindle formation in Xenopus egg extracts and in tissue culture cells (Gruss et al., 2001, 2002; Garrett et al., 2002). These studies indicate that microtubule assembly induced by TPX2 around the chromosomes is a general requirement for spindle assembly—a conclusion in sharp contrast with the idea that two centrosomal asters are sufficient to organize a spindle (see Spindle assembly without chromosomes? section).
Figure 2.

Factors proposed to be regulated by RanGTP during mitosis and their role in spindle assembly. (A) Nucleation: in the initial phases chromosomes generate an environment enriched in RanGTP. TPX2 is released from the importins and triggers microtubule nucleation. (B) Organization: microtubules assembled around the chromosomes coalesce and start to organize. Two proteins were suggested to be regulated by RanGTP and to participate in these events: Eg5 and Kid. (C) Pole formation: microtubule minus ends focus into spindle poles. This process involves two proteins regulated by RanGTP: NuMA and XCTK2. TPX2 may also play a role in this process.
TPX2 targeting activities and its role in spindle pole organization
TPX2 accumulates at the poles of metaphase spindles in a dynein–dynactin-dependent way. This localization does not depend on RanGTP because TPX2 accumulates at the center of microtubule asters induced in Xenopus egg extracts by addition of centrosomes, or agents like taxol or DMSO and binds to pure microtubules in the presence of importins (Wittmann et al., 2000; Trieselmann et al., 2003). The precise mechanism involved in this localization is still unclear. In contrast to NuMA (Merdes et al., 1996), that has a very similar localization at spindle poles, a direct interaction between TPX2 and the dynein–dynactin complex has not been described. In any case, the spindle pole localization of TPX2 may be important functionally because TPX2 is required for spindle pole integrity (Wittmann et al., 2000; Garrett et al., 2002). The function of TPX2 at spindle poles could also be related to its targeting activities. Two proteins have been shown to be targeted by TPX2, the KLP Xklp2 (Wittmann et al., 2000; Garrett et al., 2002) and the kinase Aurora A (Kufer et al., 2002). The targeting of Xklp2 by TPX2 to microtubule minus ends of asters assembled in Xenopus M-phase extracts occurs in the absence of RanGTP (Wittmann et al., 2000; Garrett et al., 2002) suggesting that its targeting to the spindle poles is independent of the Ran system as well. By contrast, the interaction between TPX2 and Aurora A is stimulated by RanGTP (Kufer et al., 2002; Eyers et al., 2003; Tsai et al., 2003; Fig. 1). Interestingly, this interaction leads to the activation of the kinase and, in turn, the phosphorylation of TPX2 by Aurora A (Eyers et al., 2003; Tsai et al., 2003). Structural and biochemical studies have helped to understand the precise mechanism by which TPX2 activates the kinase (Bayliss et al., 2003; Eyers and Maller, 2004). The binding of a short NH2-terminal sequence of TPX2 to the catalytic domain of Aurora A locks the kinase into an active conformation. First, it positions the activation segment into a more favorable conformation for substrate binding and second by inducing the swinging of a critical phosphorylated residue into a buried position protecting it from dephosphorylation by PP1 and thus inactivation (Bayliss et al., 2003). Therefore, the interaction of TPX2 with Aurora A could be a mechanism to translate the RanGTP signaling network into a phosphorylation network. Although few substrates for Aurora A have been identified, one of them is the KLP Eg5 (Giet et al., 1999) and the other is TPX2 itself (Bayliss et al., 2003; Eyers and Maller, 2004). Interestingly, Eg5 is activated by RanGTP and it has been suggested that it is regulated by Aurora kinase phosphorylation (Giet and Prigent, 2000). In contrast, it is not clear whether and how Aurora A–mediated phosphorylation of TPX2 affects its activities. In any case, the RanGTP-dependent microtubule assembly activity of TPX2 as well as its RanGTP-mediated interaction with Aurora A that leads to the activation of the kinase reveal important mechanisms by which RanGTP can control directly and indirectly the activity of factors required for spindle assembly.
Concluding remarks
The central role of Ran in spindle formation reflects the general importance of the regulation of nuclear proteins in this process. In fact, nuclear proteins, all set free early in mitosis upon nuclear envelope breakdown, are involved at all stages of M-phase. Thus, it will be interesting to see whether and how Ran works to regulate the accessibility of nuclear proteins for late functions in cell division like chromosome segregation and cytokinesis.
Acknowledgments
We thank Eric Karsenti and Iain Mattaj for useful comments on the manuscript.
Abbreviations used in this paper: KLP, kinesin-like protein; MAP, microtubule associated protein; NuMA, nuclear protein of the mitotic apparatus; RanGTP, GTPase Ran in its GTP-bound form; TPX2, targeting protein for Xklp2.
References
- Antonio, C., I. Ferby, H. Wilhelm, M. Jones, E. Karsenti, A.R. Nebreda, and I. Vernos. 2000. Xkid, a chromokinesin required for chromosome alignment on the metaphase plate. Cell. 102:425–435. [DOI] [PubMed] [Google Scholar]
- Arnaoutov, A., and M. Dasso. 2003. The Ran GTPase regulates kinetochore function. Dev. Cell. 5:99–111. [DOI] [PubMed] [Google Scholar]
- Askjaer, P., V. Galy, E. Hannak, and I.W. Mattaj. 2002. Ran GTPase cycle and importins alpha and beta are essential for spindle formation and nuclear envelope assembly in living Caenorhabditis elegans embryos. Mol. Biol. Cell. 13:4355–4370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bamba, C., Y. Bobinnec, M. Fukuda, and E. Nishida. 2002. The GTPase Ran regulates chromosome positioning and nuclear envelope assembly in vivo. Curr. Biol. 12:503–507. [DOI] [PubMed] [Google Scholar]
- Bayliss, R., T. Sardon, I. Vernos, and E. Conti. 2003. Structural basis of Aurora-A activation by TPX2 at the mitotic spindle. Mol. Cell. 12:851–862. [DOI] [PubMed] [Google Scholar]
- Bilbao-Cortes, D., M. Hetzer, G. Langst, P.B. Becker, and I.W. Mattaj. 2002. Ran binds to chromatin by two distinct mechanisms. Curr. Biol. 12:1151–1156. [DOI] [PubMed] [Google Scholar]
- Blangy, A., H.A. Lane, P. d'Herin, M. Harper, M. Kress, and E.A. Nigg. 1995. Phosphorylation by p34cdc2 regulates spindle association of human Eg5, a kinesin-related motor essential for bipolar spindle formation in vivo. Cell. 83:1159–1169. [DOI] [PubMed] [Google Scholar]
- Brunet, S., Z. Polanski, M.H. Verlhac, J.Z. Kubiak, and B. Maro. 1998. Bipolar meiotic spindle formation without chromatin. Curr. Biol. 8:1231–1234. [DOI] [PubMed] [Google Scholar]
- Bucciarelli, E., M.G. Giansanti, S. Bonaccorsi, and M. Gatti. 2003. Spindle assembly and cytokinesis in the absence of chromosomes during Drosophila male meiosis. J. Cell Biol. 160:993–999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canman, J.C., L.A. Cameron, P.S. Maddox, A. Straight, J.S. Tirnauer, T.J. Mitchison, G. Fang, T.M. Kapoor, and E.D. Salmon. 2003. Determining the position of the cell division plane. Nature. 424:1074–1078. [DOI] [PubMed] [Google Scholar]
- Carazo-Salas, R.E., O.J. Gruss, I.W. Mattaj, and E. Karsenti. 2001. Ran-GTP coordinates regulation of microtubule nucleation and dynamics during mitotic-spindle assembly. Nat. Cell Biol. 3:228–234. [DOI] [PubMed] [Google Scholar]
- Clarke, P.R., and C. Zhang. 2001. Ran GTPase: a master regulator of nuclear structure and function during the eukaryotic cell division cycle? Trends Cell Biol. 11:366–371. [DOI] [PubMed] [Google Scholar]
- Dasso, M. 2002. The Ran GTPase: theme and variations. Curr. Biol. 12:R502–R508. [DOI] [PubMed] [Google Scholar]
- Di Fiore, B., M. Ciciarello, and P. Lavia. 2004. Mitotic functions of the Ran GTPase network: the importance of being in the right place at the right time. Cell Cycle. 3:305–313. [PubMed] [Google Scholar]
- Ems-McClung, S.C., Y. Zheng, and C.E. Walczak. 2004. Importin alpha/beta and Ran-GTP regulate XCTK2 microtubule binding through a bipartite nuclear localization signal. Mol. Biol. Cell. 15:46–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eyers, P.A., and J.L. Maller. 2004. Regulation of Xenopus Aurora A activation by TPX2. J. Biol. Chem. 279:9008–9015. [DOI] [PubMed] [Google Scholar]
- Eyers, P.A., E. Erikson, L.G. Chen, and J.L. Maller. 2003. A novel mechanism for activation of the protein kinase aurora a. Curr. Biol. 13:691–697. [DOI] [PubMed] [Google Scholar]
- Faruki, S., R.W. Cole, and C.L. Rieder. 2002. Separating centrosomes interact in the absence of associated chromosomes during mitosis in cultured vertebrate cells. Cell Motil. Cytoskeleton. 52:107–121. [DOI] [PubMed] [Google Scholar]
- Forgues, M., M.J. Difilippantonio, S.P. Linke, T. Ried, K. Nagashima, J. Feden, K. Valerie, K. Fukasawa, and X.W. Wang. 2003. Involvement of Crm1 in hepatitis B virus X protein-induced aberrant centriole replication and abnormal mitotic spindles. Mol. Cell. Biol. 23:5282–5292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Funabiki, H., and A.W. Murray. 2000. The Xenopus chromokinesin Xkid is essential for metaphase chromosome alignment and must be degraded to allow anaphase chromosome movement. Cell. 102:411–424. [DOI] [PubMed] [Google Scholar]
- Garrett, S., K. Auer, D.A. Compton, and T.M. Kapoor. 2002. hTPX2 is required for normal spindle morphology and centrosome integrity during vertebrate cell division. Curr. Biol. 12:2055–2059. [DOI] [PubMed] [Google Scholar]
- Giet, R., and C. Prigent. 2000. The Xenopus laevis aurora/Ip11p-related kinase pEg2 participates in the stability of the bipolar mitotic spindle. Exp. Cell Res. 258:145–151. [DOI] [PubMed] [Google Scholar]
- Giet, R., R. Uzbekov, F. Cubizolles, K. Le Guellec, and C. Prigent. 1999. The Xenopus laevis aurora-related protein kinase pEg2 associates with and phosphorylates the kinesin-related protein XlEg5. J. Biol. Chem. 274:15005–15013. [DOI] [PubMed] [Google Scholar]
- Görlich, D., and U. Kutay. 1999. Transport between the cell nucleus and the cytoplasm. Annu. Rev. Cell Dev. Biol. 15:607–660. [DOI] [PubMed] [Google Scholar]
- Görlich, D., M.J. Seewald, and K. Ribbeck. 2003. Characterization of Ran-driven cargo transport and the RanGTPase system by kinetic measurements and computer simulation. EMBO J. 22:1088–1100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gruss, O.J., R.E. Carazo-Salas, C.A. Schatz, G. Guarguaglini, J. Kast, M. Wilm, N. Le Bot, I. Vernos, E. Karsenti, and I.W. Mattaj. 2001. Ran induces spindle assembly by reversing the inhibitory effect of importin alpha on TPX2 activity. Cell. 104:83–93. [DOI] [PubMed] [Google Scholar]
- Gruss, O.J., M. Wittmann, H. Yokoyama, R. Pepperkok, T. Kufer, H. Sillje, E. Karsenti, I.W. Mattaj, and I. Vernos. 2002. Chromosome-induced microtubule assembly mediated by TPX2 is required for spindle formation in HeLa cells. Nat. Cell Biol. 4:871–879. [DOI] [PubMed] [Google Scholar]
- Heald, R., R. Tournebize, T. Blank, R. Sandaltzopoulos, P. Becker, A. Hyman, and E. Karsenti. 1996. Self-organization of microtubules into bipolar spindles around artificial chromosomes in Xenopus egg extracts. Nature. 382:420–425. [DOI] [PubMed] [Google Scholar]
- Heidebrecht, H.J., F. Buck, J. Steinmann, R. Sprenger, H.H. Wacker, and R. Parwaresch. 1997. p100: a novel proliferation-associated nuclear protein specifically restricted to cell cycle phases S, G2, and M. Blood. 90:226–233. [PubMed] [Google Scholar]
- Hetzer, M., D. Bilbao-Cortes, T.C. Walther, O.J. Gruss, and I.W. Mattaj. 2000. GTP hydrolysis by Ran is required for nuclear envelope assembly. Mol. Cell. 5:1013–1024. [DOI] [PubMed] [Google Scholar]
- Hetzer, M., O.J. Gruss, and I.W. Mattaj. 2002. The Ran GTPase as a marker of chromosome position in spindle formation and nuclear envelope assembly. Nat. Cell Biol. 4:E177–E184. [DOI] [PubMed] [Google Scholar]
- Joseph, J., S.H. Tan, T.S. Karpova, J.G. McNally, and M. Dasso. 2002. SUMO-1 targets RanGAP1 to kinetochores and mitotic spindles. J. Cell Biol. 156:595–602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joseph, J., S.T. Liu, S.A. Jablonski, T.J. Yen, and M. Dasso. 2004. The RanGAP1-RanBP2 complex is essential for microtubule-kinetochore interactions in vivo. Curr. Biol. 14:611–617. [DOI] [PubMed] [Google Scholar]
- Kalab, P., K. Weis, and R. Heald. 2002. Visualization of a Ran-GTP gradient in interphase and mitotic Xenopus egg extracts. Science. 295:2452–2456. [DOI] [PubMed] [Google Scholar]
- Karsenti, E., J. Newport, and M. Kirschner. 1984. Respective roles of centrosomes and chromatin in the conversion of microtubule arrays from interphase to metaphase. J. Cell Biol. 99:47s–54s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keryer, G., B. Di Fiore, C. Celati, K.F. Lechtreck, M. Mogensen, A. Delouvee, P. Lavia, M. Bornens, and A.M. Tassin. 2003. Part of Ran is associated with AKAP450 at the centrosome: involvement in microtubule-organizing activity. Mol. Biol. Cell. 14:4260–4271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khodjakov, A., R.W. Cole, B.R. Oakley, and C.L. Rieder. 2000. Centrosome-independent mitotic spindle formation in vertebrates. Curr. Biol. 10:59–67. [DOI] [PubMed] [Google Scholar]
- Kufer, T.A., H.H. Sillje, R. Korner, O.J. Gruss, P. Meraldi, and E.A. Nigg. 2002. Human TPX2 is required for targeting Aurora-A kinase to the spindle. J. Cell Biol. 158:617–623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levesque, A.A., and D.A. Compton. 2001. The chromokinesin Kid is necessary for chromosome arm orientation and oscillation, but not congression, on mitotic spindles. J. Cell Biol. 154:1135–1146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, H.Y., and Y. Zheng. 2004. Phosphorylation of RCC1 in mitosis is essential for producing a high RanGTP concentration on chromosomes and for spindle assembly in mammalian cells. Genes Dev. 18:512–527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, H.Y., D. Wirtz, and Y. Zheng. 2003. A mechanism of coupling RCC1 mobility to RanGTP production on the chromatin in vivo. J. Cell Biol. 160:635–644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mattaj, I.W., and L. Englmeier. 1998. Nucleocytoplasmic transport: the soluble phase. Annu. Rev. Biochem. 67:265–306. [DOI] [PubMed] [Google Scholar]
- McKim, K.S., and R.S. Hawley. 1995. Chromosomal control of meiotic cell division. Science. 270:1595–1601. [DOI] [PubMed] [Google Scholar]
- Merdes, A., K. Ramyar, J.D. Vechio, and D.W. Cleveland. 1996. A complex of NuMA and cytoplasmic dynein is essential for mitotic spindle assembly. Cell. 87:447–458. [DOI] [PubMed] [Google Scholar]
- Moore, J.D. 2001. The Ran-GTPase and cell-cycle control. Bioessays. 23:77–85. [DOI] [PubMed] [Google Scholar]
- Moore, W., C. Zhang, and P.R. Clarke. 2002. Targeting of RCC1 to chromosomes is required for proper mitotic spindle assembly in human cells. Curr. Biol. 12:1442–1447. [DOI] [PubMed] [Google Scholar]
- Nachury, M.V., T.J. Maresca, W.C. Salmon, C.M. Waterman-Storer, R. Heald, and K. Weis. 2001. Importin beta is a mitotic target of the small GTPase Ran in spindle assembly. Cell. 104:95–106. [DOI] [PubMed] [Google Scholar]
- Nedelec, F. 2002. Computer simulations reveal motor properties generating stable antiparallel microtubule interactions. J. Cell Biol. 158:1005–1015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nigg, E.A. 2001. Mitotic kinases as regulators of cell division and its checkpoints. Nat. Rev. Mol. Cell Biol. 2:21–32. [DOI] [PubMed] [Google Scholar]
- Quimby, B.B., and M. Dasso. 2003. The small GTPase Ran: interpreting the signs. Curr. Opin. Cell Biol. 15:338–344. [DOI] [PubMed] [Google Scholar]
- Rebollo, E., and C. Gonzalez. 2000. Visualizing the spindle checkpoint in Drosophila spermatocytes. EMBO Rep. 1:65–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rebollo, E., S. Llamazares, J. Reina, and C. Gonzalez. 2004. Contribution of noncentrosomal microtubules to spindle assembly in Drosophila spermatocytes. PLoS Biol. 2:E8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawin, K.E., K. LeGuellec, M. Philippe, and T.J. Mitchison. 1992. Mitotic spindle organization by a plus-end-directed microtubule motor. Nature. 359:540–543. [DOI] [PubMed] [Google Scholar]
- Schatz, C.A., R. Santarella, A. Hoenger, E. Karsenti, I.W. Mattaj, O.J. Gruss, and R.E. Carazo-Salas. 2003. Importin alpha-regulated nucleation of microtubules by TPX2. EMBO J. 22:2060–2070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trieselmann, N., and A. Wilde. 2002. Ran localizes around the microtubule spindle in vivo during mitosis in Drosophila embryos. Curr. Biol. 12:1124–1129. [DOI] [PubMed] [Google Scholar]
- Trieselmann, N., S. Armstrong, J. Rauw, and A. Wilde. 2003. Ran modulates spindle assembly by regulating a subset of TPX2 and Kid activities including Aurora A activation. J. Cell Sci. 116:4791–4798. [DOI] [PubMed] [Google Scholar]
- Tsai, M.Y., C. Wiese, K. Cao, O. Martin, P. Donovan, J. Ruderman, C. Prigent, and Y. Zheng. 2003. A Ran signalling pathway mediated by the mitotic kinase Aurora A in spindle assembly. Nat. Cell Biol. 5:242–248. [DOI] [PubMed] [Google Scholar]
- Walczak, C.E., S. Verma, and T.J. Mitchison. 1997. XCTK2: a kinesin-related protein that promotes mitotic spindle assembly in Xenopus laevis egg extracts. J. Cell Biol. 136:859–870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weis, K. 2003. Regulating access to the genome: nucleocytoplasmic transport throughout the cell cycle. Cell. 112:441–451. [DOI] [PubMed] [Google Scholar]
- Wiese, C., A. Wilde, M.S. Moore, S.A. Adam, A. Merdes, and Y. Zheng. 2001. Role of importin-{beta} in coupling Ran to downstream targets in microtubule assembly. Science. 291:653–656. [DOI] [PubMed] [Google Scholar]
- Wilde, A., and Y. Zheng. 1999. Stimulation of microtubule aster formation and spindle assembly by the small GTPase Ran. Science. 284:1359–1362. [DOI] [PubMed] [Google Scholar]
- Wilde, A., S.B. Lizarraga, L. Zhang, C. Wiese, N.R. Gliksman, C.E. Walczak, and Y. Zheng. 2001. Ran stimulates spindle assembly by altering microtubule dynamics and the balance of motor activities. Nat. Cell Biol. 3:221–227. [DOI] [PubMed] [Google Scholar]
- Wittmann, T., M. Wilm, E. Karsenti, and I. Vernos. 2000. TPX2, a novel Xenopus MAP involved in spindle pole organization. J. Cell Biol. 149:1405–1418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yajima, J., M. Edamatsu, J. Watai-Nishii, N. Tokai-Nishizumi, T. Yamamoto, and Y.Y. Toyoshima. 2003. The human chromokinesin Kid is a plus end-directed microtubule-based motor. EMBO J. 22:1067–1074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi, R., and J. Newport. 2003. A role for Ran-GTP and Crm1 in blocking re-replication. Cell. 113:115–125. [DOI] [PubMed] [Google Scholar]
- Zhang, C., and P.R. Clarke. 2001. Roles of Ran-GTP and Ran-GDP in precursor vesicle recruitment and fusion during nuclear envelope assembly in a human cell-free system. Curr. Biol. 11:208–212. [DOI] [PubMed] [Google Scholar]
- Zhang, C., M. Hughes, and P.R. Clarke. 1999. Ran-GTP stabilises microtubule asters and inhibits nuclear assembly in Xenopus egg extracts. J. Cell Sci. 112:2453–2461. [DOI] [PubMed] [Google Scholar]
- Zhang, D., and R.B. Nicklas. 1995. The impact of chromosomes and centrosomes on spindle assembly as observed in living cells. J. Cell Biol. 129:1287–1300. [DOI] [PMC free article] [PubMed] [Google Scholar]