

Abstract
Ezrin/radixin/moesin (ERM) proteins cross-link actin filaments to plasma membranes to integrate the function of cortical layers, especially microvilli. We found that in cochlear and vestibular sensory hair cells of adult wild-type mice, radixin was specifically enriched in stereocilia, specially developed giant microvilli, and that radixin-deficient (Rdx − / −) adult mice exhibited deafness but no obvious vestibular dysfunction. Before the age of hearing onset (∼2 wk), in the cochlea and vestibule of Rdx − / − mice, stereocilia developed normally in which ezrin was concentrated. As these Rdx − / − mice grew, ezrin-based cochlear stereocilia progressively degenerated, causing deafness, whereas ezrin-based vestibular stereocilia were maintained normally in adult Rdx − / − mice. Thus, we concluded that radixin is indispensable for the hearing ability in mice through the maintenance of cochlear stereocilia, once developed. In Rdx − / − mice, ezrin appeared to compensate for radixin deficiency in terms of the development of cochlear stereocilia and the development/maintenance of vestibular stereocilia. These findings indicated the existence of complicate functional redundancy in situ among ERM proteins.
Keywords: ERM; radixin; stereocilia; cochlea; deafness
Introduction
Deafness is one of the most prevalent forms of sensory impairment in humans, and it affects ∼1 in 1,000 children (for reviews see Morton, 1991; Fortnum and Davis, 1997). Various gene mutations are known to cause hereditary hearing loss in humans (for reviews see Holme and Steel, 1999; Petit et al., 2001; Tekin et al., 2001; Zuo, 2002). Hearing is achieved through a number of steps: first, acoustic stimuli reaching the cochlea deflect the stereocilia on the apical surface of the inner and outer hair cells in the organ of Corti, which in turn opens the mechanotransduction channels located at the tips of stereocilia. Potassium then floods through these open channels, resulting in plasma membrane depolarization, and an electrical signal is transmitted to the central nervous system (for reviews see Roberts et al., 1988; Hudspeth, 1989; Pickles and Corey, 1992). Thus, the stereocilia of cochlear hair cells are key elements in the transduction of acoustic stimuli into electrical signals. Stereocilia also occur in the hair cells of the vestibule to detect acceleration (Eatock et al., 1998; Corey, 2003). Deaf mouse models have provided crucial clues for understanding the molecular and cellular biology of cochlear sensory hair cells, and several forms of hearing loss are now understood to be caused by mutations of the cytoskeletal proteins of cochlear stereocilia (Probst and Camper, 1999; Gillespie and Walker, 2001; Steel and Kros, 2001; Call and Morton, 2002; Zuo, 2002; Belyantseva et al., 2003). In these mice, vestibular stereocilia were also defective, concomitantly causing imbalance.
Stereocilia are specifically developed giant microvilli (finger-like projections of plasma membrane that are underlain by actin filaments). Microvilli show significant variations in morphology, diameter, and length depending on cell types; some are formed by densely packed rigid-looking actin filaments, whereas others are formed by loosely packed fragile-looking actin filaments (Furukawa and Fechheimer, 1997; Bartles, 2000; DeRosier and Tilney, 2000). Of these microvilli, the stereocilia of cochlear sensory hair cells are the most specialized with a characteristic morphology (DeRosier et al., 1980; Gillespie and Walker, 2001; Zuo, 2002). The stereocilia of vestibular sensory hair cells show a more fragile appearance (Sobin and Flock, 1983; Denman-Johnson and Forge, 1999). Besides the general microvilli cytoskeletal proteins such as actin, fimbrin, and myosin Ic, which were immunolocalized to stereocilia (Tilney et al., 1989; Drenckhahn et al., 1991; Skowron et al., 1998; Dumont et al., 2002), the positional cloning of deafness genes in humans and mice has enabled the identification of the novel cytoskeleton-associated constituents of stereocilia such as espin, harmonin, SANS, whirlin, unconventional myosins (VI/VIIa/XV), and cadherins (cadherin 23 and protocadherin 15); these proteins are most likely implicated during stereocilia formation and/or maintenance (Avraham et al., 1995; Gibson et al., 1995; Probst et al., 1998; Liang et al., 1999; Littlewood Evans and Müller, 2000; Zheng et al., 2000; Alagramam et al., 2001; Di Palma et al., 2001; Wada et al., 2001; Boeda et al., 2002; Karolyi et al., 2003; Mburu et al., 2003; Weil et al., 2003). Despite these advances, we are still far from having a comprehensive view of how the stereocilia of hair cells are built during development and maintained throughout life.
Ezrin/radixin/moesin (ERM) proteins are three closely related proteins in the band 4.1 superfamily that are thought to function as cross-linkers between plasma membranes and actin filaments, thus integrating the actin cortical layer, especially the microvilli (Sato et al., 1992; Berryman et al., 1993; Mangeat et al., 1999; Tsukita and Yonemura, 1999; Bretscher et al., 2002). The COOH-terminal domains of these ERM proteins bind to the actin filaments (Algrain et al., 1993; Turunen et al., 1994; Martin et al., 1995), and the NH2-terminal halves, the FERM domains (band 4.1/ezrin/radixin/moesin homology domains; Algrain et al., 1993; Henry et al., 1995; Bretscheret al., 2002), associate directly with the cytoplasmic domains of several integral membrane proteins such as CD43, CD44, and ICAM-1/2/3 (Tsukita et al., 1994; Helander et al., 1996; Serrador et al., 1997; Yonemura et al., 1998, 1999; Shaw, 2001). These NH2-terminal halves are also indirectly associated with membrane transporters/channels such as the Na+/H+ exchanger 3, CFTR (cystic fibrosis transmembrane conductance regulator), β2-adrenergic receptor, and PDGF-receptor, via a cytoplasmic phosphoprotein, EBP50/NHE-RF (Reczek et al., 1997; Yun et al., 1997; Murthy et al., 1998; Short et al., 1998; Maudsley et al., 2000; Bretscher et al., 2002). The cross-linking activity of ERM proteins between actin filaments and plasma membranes is thought to be regulated by PIP2 in the downstream of Rho (Hirao et al., 1996; Mackay et al., 1997; Heiska et al., 1998; Matsui et al., 1999; Yonemura et al., 2002).
The suppression of ERM protein expression in lymphocytes using antisense oligonucleotides results in the complete disappearance of microvilli, suggesting that they play an essential role in cortical microvilli formation (Takeuchi et al., 1994). To examine the physiological functions of ERM proteins at the entire body level (Ingraffea et al., 2002), moesin- and radixin-deficient mice have been generated (Doi et al., 1999; Kikuchi et al., 2002). Moesin-deficient mice showed no abnormalities when examined under laboratory conditions (Doi et al., 1999). By contrast, radixin deficiency caused congenital conjugated hyperbilirubinemia with losses of multidrug resistance protein 2, a bilirubin transporter, from the bile canalicular membranes of the hepatocytes with concomitant decrease of microvilli in number as well as in length (Kikuchi et al., 2002). In wild-type mice, radixin was the dominant ERM protein in the hepatocytes, and was not substituted by either ezrin or moesin in Rdx −/− mice.
Taking all of the above into consideration, it would seem important that the possible involvement of ERM proteins in the formation, maintenance, and functions of stereocilia on hair cells be examined. In this work, we found that radixin was the only detectable species of ERM protein in the stereocilia of the cochlea of adult wild-type mice, whereas vestibular stereocilia appeared to contain a detectable amount of ezrin in addition to radixin. Adult Rdx −/− mice exhibited profound deafness, but not imbalance; consistent with this, stereocilia were significantly defective in the outer and inner hair cells of the cochlea, but not in the hair cells of the vestibule. Furthermore, we found that in the cochlea of Rdx −/− mice, ezrin-based stereocilia appeared to develop normally from birth to postnatal day 14 (P14), the onset of hearing, after which they progressively degenerated. Thus, the radixin knockout provides a type of mouse deafness model caused by mutation in cytoskeletal components in stereocilia, which presents with progressive degeneration of cochlear (but not vestibular) stereocilia. The present paper provides a new insight into the ERM-based molecular mechanisms for the formation and maintenance of stereocilia, and the functional specificity/redundancy among ERM proteins in situ.
Results
Radixin is specifically enriched in stereocilia in the cochlear and vestibular sensory hair cells in adult wild-type mice
As a first step to examine the possible involvement of ERM proteins in the formation/maintenance of stereocilia on hair cells, the organ of Corti and the crista ampullaris of the vestibule were isolated from adult wild-type mice aged 5 wk, and then whole-mount stained with ezrin-, radixin-, and moesin-specific mAbs (Kondo et al., 1997; Doi et al., 1999). In the organ of Corti, only radixin was concentrated both in one row of the inner hair cell stereocilia and three rows of the outer hair cell stereocilia (Fig. 1, a–c): Individual stereocilia appeared resolved both by radixin staining in immunofluorescence microscopy (Fig. 1 d) and by scanning EM (Fig. 1 e). In addition to such intense signals, diffuse weak staining for ezrin and radixin was also detected in both hair and supporting cells of the organ of Corti, whereas moesin was not detected in either; instead it was concentrated in the VE-cadherin–positive blood vessels (Berryman et al., 1993). In the crista ampullaris of the vestibule, radixin was concentrated in the stereocilia of the hair cells, but ezrin was also detectable in stereocilia, though faintly (Fig. 1, f–h). At higher magnification, individual long fragile vestibular stereocilia were also resolved both by radixin staining in immunofluorescence microscopy (Fig. 1 i) and by scanning EM (Fig. 1 j). Moesin was detected only in the VE-cadherin–positive blood vessels. These results indicated that radixin is the dominant ERM protein in the hair cell stereocilia of the inner ear, although the degree of contribution of ezrin appeared to be different between the cochlear and vestibular stereocilia.
Figure 1.

ERM proteins in the cochlea and vestibule in adult wild-type mice at the age of 5 wk. (a–c) Whole-mount immunofluorescence micrographs of the organ of Corti isolated from the cochlea with mAbs specific for ezrin, radixin, or moesin. The stereocilia of inner hair cells (IHC) and outer hair cells (OHC) are intensely stained with anti-radixin mAb (arrows). Ezrin and moesin are undetectable in the stereocilia, though moesin (moesin; green) is detected in large amounts in VE-cadherin–positive blood vessel endothelial cells (VE-cad; red) at a different focus plane (inset). Bar, 15 μm. (d and e) Comparison of the whole-mount radixin staining image with the scanning electron microscopic image of stereocilia on outer hair cells. Both radixin immunostaining (d) and scanning EM (e) reveal a highly organized array of stereocilia (arrows). Bars, 5 μm. (f–h) Whole-mount immunofluorescence micrographs of the crista ampullaris isolated from the vestibule with mAbs specific for ezrin, radixin, or moesin. Stereocilia are intensely stained with anti-radixin mAb, and weakly but reproducibly stained with anti-ezrin mAb (arrows). Moesin is detected only in the blood vessels at a different focus plane from radixin/ezrin-positive stereocilia (inset), as judged by the double-staining images for moesin (moesin; green) and VE-cadherin (VE-cad; red, inset). Bar, 15 μm. (i and j) Comparison of the whole-mount radixin staining image with the scanning electron microscopic image of stereocilia on vestibular hair cells. Radixin appears to concentrate along the entire length of long fragile stereocilia. Bars, 3 μm.
Radixin deficiency causes deafness
Thus, it would be interesting to examine the physiological roles of radixin in hearing and balance in mice. First, we focused on the hearing ability of radixin-deficient (Rdx −/−) mice. These mice showed a normal growth rate (Fig. 2 A) and were fertile, although they suffered from congenital conjugated hyperbilirubinemia (Kikuchi et al., 2002). Interestingly, when a sound stimulus was administrated in the form of a loud handclap, adult Rdx −/− mice (5–10-wk-old) showed no reflexive reaction, so-called Preyer's reflex, although Rdx +/+ and Rdx +/− mice quickly moved their heads (Fig. 2 A). The auditory-evoked brainstem response (ABR) was then measured in two sets of heterogeneous Rdx +/− intercross adult littermates aged 5 and 10 wk (12 littermates in total) in response to a stimuli with a sound pressure level (SPL) of 70 decibels (dB) (20 kHz; Fig. 2 B). Of the 12 littermates, three showed no ABR; the others all showed a typical ABR waveform. Afterwards, these mice were genotyped, and these three littermates showing no ABR were all found to be Rdx −/− mice. This perfect correlation between the Rdx −/− genotype and a lack of ABR was reproducibly obtained in different series of measurements. In Fig. 2 C, the hearing thresholds of 10-wk-old mice were measured at various sound frequencies. Wild-type and Rdx +/− mice showed normal hearing thresholds (10–50 dB SPL), whereas Rdx −/− mice showed profound deafness (hearing threshold, >70–90 dB SPL).
Figure 2.
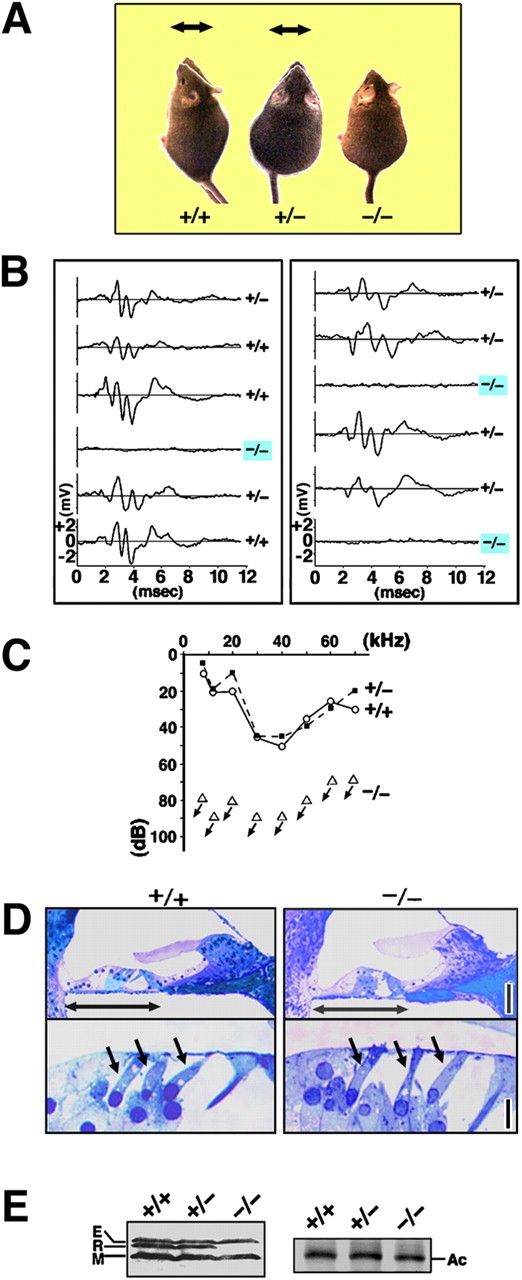
Deafness of Rdx −/− mice. (A) Loss of Preyer's reflex in Rdx −/− mice. Time-lapse photography captures Preyer's reflex in Rdx +/+ and Rdx +/− mice, head movements (arrows), but not in Rdx −/− mice. Two successive frames after a loud handclap (1-s interval) were superimposed. These 10-wk-old mice show similar growth rates. (B) ABR to stimuli of 70-dB SPL (20 kHz) in two sets of Rdx +/− intercross littermates (left, 5-wk-old; right 10-wk-old). Among 12 littermates in total, three show no ABR, and were afterwards genotyped as Rdx −/− mice (blue squares). (C) Hearing thresholds of 10-wk-old Rdx +/+, Rdx +/−, and Rdx −/− mice at various sound frequencies. Rdx +/+ and Rdx +/− mice show normal hearing thresholds (10–50 dB SPL), whereas Rdx −/− mice show profound deafness (hearing threshold, >70–90 dB SPL). (D) Toluidine blue–stained Epon semi-thin sections of the cochlea. No gross morphological difference is observed in the organ of Corti (double-headed arrows) including hair cells (single-headed arrows) between Rdx +/+ and Rdx −/− mice. Bars, 50 μm (top panels); 10 μm (bottom panels). (E) Western blot analysis of isolated cochleae of the Rdx +/+ , Rdx +/−, and Rdx −/− mice with anti-ERM pAb (TK89) that recognizes ezrin (E), radixin (R), and moesin (M) equally. In the Rdx −/− cochlea, radixin becomes undetectable without significant up-regulation of ezrin or moesin. Silver-stained bands of actin (Ac) in the same gels are present to show that an equal amount of cell lysate was applied in each lane.
Then, we compared light microscopic images of toluidine blue–stained sections prepared from Epon-embedded tissues of the inner ear between Rdx +/+ and Rdx −/− P60 mice (Fig. 2 D): No obvious gross morphological malformations were observed in Rdx −/− cochlea including hair cells, which was consistent with the scanning electron microscopic observation (see Fig. 3 A). However, as Rdx −/− mice became much older (>90 d), hair cells appeared to begin to be eliminated, leaving a fairly disorganized cell architecture of the organ of Corti (unpublished data). The expression of ERM proteins were examined in the Rdx +/+, Rdx +/−, and Rdx −/− cochleae isolated from P40 adult littermates by Western blotting with anti-ERM pAb (Fig. 2 E). Ezrin, radixin, and moesin were detected in both Rdx +/+ and Rdx +/− cochleae, but Rdx −/− cochlea lacked radixin expression specifically and showed no significant up-regulation of ezrin or moesin.
Figure 3.
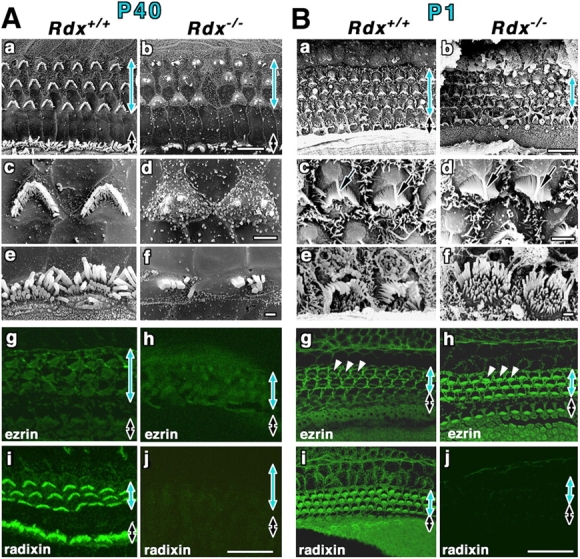
Stereocilia on Rdx +/+ and Rdx −/− cochlear hair cells of adult and newborn mice. (A) Scanning electron micrographs (a–f) and whole-mount immunofluorescence micrographs for radixin and ezrin (g–j) of the Rdx +/+ and Rdx −/− organ of Corti at 40 d of age (P40). The levels of outer hair cells (blue arrows) and inner hair cells (black arrows) are shown. At low magnification in the scanning electron micrograph of the Rdx +/+ organ of Corti (a), the luminal surface is characterized by three rows of outer hair cells (blue arrow) and one row of inner hair cells (black arrow). Outer hair cells bear stereocilia arranged regularly in the form of a letter W (c), and inner hair cells have more disorganized arrays of stereocilia (e). In the Rdx −/− organ of Corti (b), no significant abnormalities are detected in the cellular arrangements on the luminal surface, but the morphology of the stereocilia on both the outer (blue arrow) and inner (black arrow) hair cells is severely affected. In Rdx −/− outer hair cells, instead of regularly arranged stereocilia, 1–3 residual knoblike protrusions are observed on their apical surface (d). Rdx −/− inner hair cells bear several shorter fused irregularly shaped protrusions instead of long stereocilia (f). By immunostaining, in the Rdx −/− organ of Corti (h and j), no staining for radixin is detected without any increase in the staining intensities for ezrin. Bars, 10 μm (a and b); 2 μm (c and d); 1 μm (e and f); 20 μm (g–j). (B) Scanning electron micrographs (a–f) and whole-mount immunofluorescence micrographs for radixin and ezrin (g–j) of the Rdx +/+ and Rdx −/− organ of Corti at 1 d of age (P1). Scanning electron microscopic images of the cochlea of P1 Rdx −/− mice (b, d, and f) are indistinguishable from those in the Rdx +/+ mice (a, c, and e). In addition to stereocilia arrays carrying central tubulin-based kinocilium (arrows in c and d), hair cells as well as supporting cells are covered with large numbers of short conventional microvilli that have mostly disappeared in adult mice. Whole-mount immunostaining reveals that in the P1 Rdx +/+ organ of Corti, radixin is highly enriched in stereocilia of inner and outer hair cells as well as the apical surface of hair and supporting cells (i), but in contrast to adult mice ezrin is also detected in stereocilia, weakly but clearly (g, arrowheads). In the Rdx −/− organ of Corti, instead of radixin, ezrin is highly concentrated at stereocilia on both inner and outer hair cells (h, arrowheads). Bars, 10 μm (a and b); 2 μm (c and d); 1 μm (e and f); 20 μm (g–j).
Radixin deficiency causes progressive degeneration of stereocilia of cochlear hair cells after the onset of hearing
The question then naturally arose as to what is the consequence of radixin deficiency in the cochlear stereocilia in Rdx −/− mice. Isolated cochleae of P40 Rdx +/+ and Rdx −/− littermates were examined by scanning EM (Fig. 3 A, a–f). When removing the lateral wall and tectorial membrane of the cochlea in P40 Rdx −/− mice to expose the organ of Corti, the cellular arrangement of the outer and inner hair cells as well as the supporting cells appeared normal, but very interestingly, the stereocilia in both outer and inner hair cells were significantly defective compared with those in Rdx +/+ organ of Corti. Instead of being regularly arranged in a “W” shape, stereocilia of the outer hair cells of Rdx −/− mice were deformed to 1–3 residual knoblike protrusions on the apical surfaces (Fig. 3 A, d). Inner hair cells had several shorter fused protrusions instead of stereocilia (Fig. 3 A, f). When the organ of Corti isolated from these Rdx −/− cochleae was whole-mount stained with ezrin-, radixin-, and moesin-specific mAbs, ezrin, radixin, or moesin was not detected in the residual structures of outer and inner stereocilia, i.e., knoblike protrusions and shorter fused protrusions, respectively (Fig. 3 A, g–j). Diffuse ezrin staining in the organ of Corti and intense moesin staining in blood vessels did not appear to be affected. Considering that stereocilia play a central role in transducing acoustic stimuli into electrical signals in the cochlea, it is likely that these defects in cochlear stereocilia are responsible for the hearing impairments observed in adult Rdx −/− mice.
Cochlear hair cells of mice were reported to continue to develop after birth, and to become functionally mature at the onset of hearing around the age of 14 d (Lim and Anniko, 1985; Erven et al., 2002; Zuo, 2002). Therefore, we next attempted to distinguish whether in Rdx −/− mice cochlear stereocilia are defective in their development or degenerate after they have developed normally. As previously reported (Lim and Anniko, 1985; Zuo, 2002), in the cochleae of wild-type P1 mice, scanning EM showed that both the inner and outer hair cells already bore well-developed stereocilia in a characteristic arrangement (Fig. 3 B, a, c, and e). The central tubulin-based kinocilium of the W-shaped array of stereocilia on inner hair cells were especially prominent at this early stage. In addition to stereocilia, at this stage hair cells as well as supporting cells were covered with a large number of short conventional microvilli that disappeared in adult mice (Fig. 3 A). To our surprise, the scanning electron microscopic images of the cochlea of P1 Rdx −/− mice were indistinguishable from those in the Rdx +/+ mice (Fig. 3 B, b, d, and f). The question is how stereocilia were developed normally in the absence of radixin in P1 Rdx −/− mice. Whole-mount immunostaining was then performed (Fig. 3 B, g–j). In P1 Rdx +/+ mice, radixin was highly enriched in stereocilia of inner and outer hair cells, but in contrast to adult mice, ezrin was also detected in stereocilia, weakly but clearly (Fig. 3 B, g and i). Furthermore, the P1 Rdx +/+ cochlea was characterized by punctate radixin and ezrin signals on the apical surface of hair cells and supporting cells, which may correspond to numerous conventional microvilli. Of course, in the organ of Corti in P1 Rdx −/− littermates, radixin was undetectable (Fig. 3 B, j), but instead of radixin, ezrin was markedly increased in stereocilia of both inner and outer hair cells as well as in conventional microvilli on hair and supporting cells (Fig. 3 B, h). Therefore, ezrin appeared to compensate for radixin deficiency in the development of hair cell stereocilia, at least up to 1 d after birth. Subsequently, these ezrin-based stereocilia degenerated postnatally.
We then pursued the possible postnatal degeneration process of stereocilia in Rdx −/− mice (Fig. 4). At P6, scanning EM did not detect any significant differences between Rdx +/+ and Rdx −/− cochlea, indicating that there was still no sign for stereocilia degeneration (Fig. 4 A, a–f). However, in Rdx +/+ cochlea, ezrin became weaker during P1 to P14, almost undetectable from stereocilia at ∼P14, and radixin was dominantly detected among ERM proteins in stereocilia (Fig. 4 A, g and i). Supporting cells were intensely stained for ezrin and radixin. In P6 Rdx −/− cochlea, ezrin was highly concentrated in stereocilia on both inner and outer hair cells (Fig. 4 A, h) with no radixin staining (Fig. 4 A, j). Around age P14 (onset of hearing in mice), in Rdx −/− cochlea, the sign for stereocilia degeneration began to be detected on outer hair cells by scanning EM (Fig. 4 B, a–f): The central part of the W-shaped row of stereocilia was lost, leaving discontinuous and disorganized arrays of stereocilia. Ezrin was still concentrated in these degenerating stereocilia in large amounts, though in Rdx +/+ hair cells, radixin, but not ezrin, was enriched at stereocilia (Fig. 4 B, g–j). We concluded that in Rdx −/− cochleae, ezrin can counterbalance radixin deficiency to normally develop stereocilia up to the onset of hearing, but not to maintain these once-developed stereocilia to transmit acoustic stimuli into electrical signals.
Figure 4.
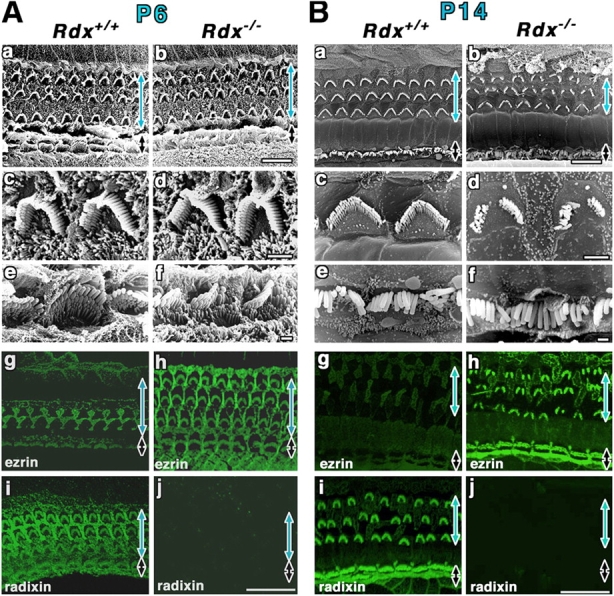
Postnatal degeneration of stereocilia on Rdx −/− cochlear hair cells. (A) Scanning electron micrographs (a–f) and whole-mount immunofluorescence micrographs for radixin and ezrin (g–j) of the organ of Corti isolated from Rdx +/+ and Rdx −/− mice at 6 d of age (P6). The levels of outer hair cells (blue arrows) and inner hair cells (black arrows) are shown. Scanning EM does not detect any significant differences between Rdx +/+ (a, c, and e) and Rdx −/− cochlea (b, d, and f). In contrast to P1 Rdx +/+ cochlea, in P6 Rdx +/+ cochlea, whole-mount immunostaining detects a weak signal of ezrin at radixin-enriched stereocilia on inner hair cells (black arrow), and a trace of ezrin in outer hair cells (blue arrow) (g and i). In Rdx −/− cochlea (h and j), ezrin is still highly concentrated at stereocilia on both inner hair cells (black arrow) and outer hair cells (blue arrow). Bars, 10 μm (a and b); 2 μm (c and d); 1 μm (e and f); 20 μm (g–j). (B) Scanning electron micrographs (a–f) and whole-mount immunofluorescence micrographs for radixin and ezrin (g–j) of the organ of Corti isolated from Rdx +/+ and Rdx −/− mice at 14 d of age (P14). Scanning EM detects the initial sign for the degeneration of stereocilia on outer hair cells (blue arrow) (a–d). The central part of the W-shaped row of stereocilia is lost, leaving discontinuous and disorganized arrays of stereocilia in Rdx −/− cochlea. At this stage, no significant defects can be observed in stereocilia on the inner hair cells (black arrow) (b and f). In P14 Rdx +/+ cochlea, ezrin is only weakly detected at radixin-enriched stereocilia on inner hair cells (black arrow), and is not detected in outer hair cells (blue arrow) (g and i). In Rdx −/− cochlea, ezrin was still concentrated in the degenerating stereocilia in large amounts in outer and inner hair cells (h). Bars, 10 μm (a and b); 2 μm (c and d); 1 μm (e and f); 20 μm (g–j).
To clarify the interior structure of ezrin-based stereocilia in Rdx −/− cochleae, the cochleae of Rdx −/− mice aged 3 wk were examined by ultrathin-section EM (Fig. 5). In these mice, stereocilia at various stages of degeneration were observed. Ezrin-based stereocilia contained highly organized bundles of actin filaments as cores that were indistinguishable in appearance from those in wild-type radixin-based stereocilia, and during the process of degeneration these stereocilia as well as core bundles appeared to be fused to form abnormal thick and short protrusions.
Figure 5.
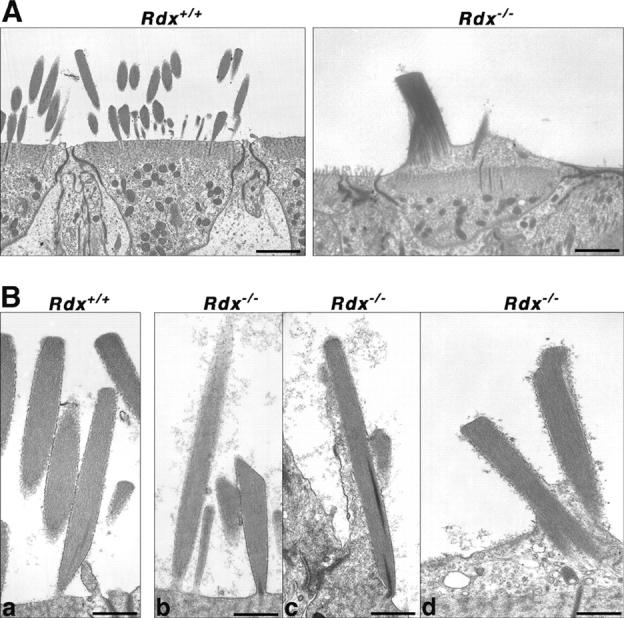
Ultrathin-section electron micrographs of stereocilia of cochlear outer hair cells of Rdx +/+ and Rdx −/− mice aged 3 wk. (A) Low power electron micrographs. All of the Rdx +/+ hair cells bear numerous stereocilia with normal appearance, whereas Rdx −/− hair cells were frequently characterized by abnormal 1–3 residual knoblike cellular protrusions on their apical surfaces. These images may correspond to the scanning electron micrographs of Fig. 3 A, c and d, respectively. Bars, 2 μm. (B) Rdx +/+ radixin-based stereocilia (a) and Rdx −/− ezrin-based stereocilia at various degeneration stages (b–d). Rdx −/− ezrin-based stereocilia contained highly-organized bundles of actin filaments as cores (b) that were indistinguishable in appearance from those in Rdx +/+ radixin-based stereocilia (a), and during the process of degeneration, these stereocilia as well as core bundles appeared to be fused to form abnormal thick and short protrusions (c and d). Bars, 0.5 μm.
Finally, a natural question is what are the membrane-binding partners for radixin/ezrin in cochlear stereocilia. To elucidate an answer, we first performed immunoprecipitation experiments with anti-radixin or anti-ezrin mAb using isolated Rdx +/+ or Rdx −/− cochlea; however, the results were inconclusive, probably due to the limitation of the material and the low insolubility of radixin and ezrin. We also performed a gel overlay blot with labeled radixin/ezrin, but again failed to detect any bands specifically associated with radixin/ezrin.
Ezrin compensates radixin deficiency in the development/maintenance of stereocilia in the vestibular hair cells
As shown in Fig. 1, in the vestibule of wild-type mice aged 5–10 wk, radixin was predominantly concentrated in stereocilia among ERM proteins, but distinct from cochlear stereocilia, these stereocilia contained detectable amounts of ezrin. The question then arose as to the structure and function of vestibular stereocilia in adult Rdx −/− mice. Structurally, scanning EM identified no abnormality in the number/density and morphology of stereocilia at the crista ampullaris of the vestibule of adult Rdx −/− mice (Fig. 6 A). Also, scanning EM identified no abnormality in the otolith organs (the utricle and saccule) of adult Rdx −/− mice (unpublished data). Functionally, Rdx −/− vestibular stereocilia appeared to be normal because adult Rdx −/− mice showed no signs of imbalance in their behavior as far as we examined up to 100 d after birth. Then, their vestibulo-ocular reflex (VOR) was measured, in which the function of horizontal semicircular canals, but not otolith organs (the utricle and saccule), can be evaluated. Heads of Rdx +/+, Rdx +/−, or Rdx −/− mice aged ∼5–10 wk were rotated sinusoidally and eye positions were recorded using a CCD camera at various head rotation frequencies. As shown in Fig. 6 B, the VOR gains of Rdx −/− mice were normal at all head rotation frequencies.
Figure 6.

The vestibule and the balance function of 5-wk-old Rdx −/− mice. (A) Scanning EM of the Rdx +/+ and Rdx −/− crista ampullaris of the vestibule. The appearance of stereocilia (arrows) was indistinguishable between Rdx +/+ and Rdx −/− mice. Bar, 5 μm. (B) VOR. A mouse head was rotated sinusoidally and the eye position recorded by a CCD camera (left). The eye velocity was calculated from the change of eye position, and was fitted with sinusoidal curves (red lines). The gain was obtained by dividing the peak eye velocity by the peak head velocity. No difference is detected in the VOR between Rdx +/+ and Rdx −/− mice. The VOR gains of Rdx −/− mice are also normal at any frequency of the head rotation stimulus in Rdx +/+ and Rdx −/− mice (right). (C) Western blot analysis of isolated Rdx +/+ and Rdx −/− vestibule (mainly crista ampullaris) with anti-ERM pAb (TK89) that recognizes ezrin, radixin, and moesin with almost the same affinity. In Rdx −/− vestibule, radixin (R) became undetectable without significant up-regulation of ezrin (E) or moesin (M). Silver-stained bands of actin (Ac) are presented on the right to show that equal amounts of cell lysate were applied in each lane. (D) Whole-mount immunostaining of the crista ampullaris isolated from the Rdx +/+ and Rdx −/− vestibules with ezrin-, radixin- and moesin-specific antibodies. In the Rdx +/+ crista ampullaris, radixin is highly concentrated in the stereocilia where ezrin is detected weakly but reproducibly (inset, a frozen section stained with anti-ezrin antibody). Moesin is detected only in the blood vessels at a different focus plane from stereocilia (inset), as judged by the double staining images for moesin (moesin; green) and VE-cadherin (VE-cad; red) (inset). By contrast, in the Rdx −/− crista ampullaris, the concentration of ezrin in the stereocilia is significantly increased. All samples were treated under completely identical conditions, paying special attention to ensure that the signals were not saturated. Bar, 40 μm.
Western blotting of the isolated vestibule with anti-ERM pAb revealed no significant up-regulation of ezrin or moesin in adult Rdx −/− mice (Fig. 6 C). We then whole-mount stained the crista ampullaris isolated from adult Rdx +/+ and Rdx −/− vestibule with ezrin-, radixin-, and moesin-specific mAbs (Fig. 6 D). In the vestibule of Rdx +/+ mice, radixin was specifically and highly enriched in the stereocilia of the crista ampullaris with a small amount of ezrin, whereas in the Rdx −/− crista ampullaris, instead of radixin, ezrin was highly concentrated in stereocilia, although this up-regulation of ezrin in stereocilia could not be detected by immunoblotting of the whole vestibule (Fig. 6 C). Therefore, in the vestibule, ezrin appears to be able to compensate radixin deficiency not only in developing stereocilia, but also in maintaining them to transmit acceleration stimuli into electrical signals.
Discussion
The physiological functions of ERM proteins, ezrin, radixin, and moesin, have attracted increasing interest, but the relevance of the existence of these three closely related proteins in mammalian cells remains elusive (Berryman et al., 1993; Takeuchi et al., 1994; Doi et al., 1999; Tsukita and Yonemura, 1999; Ingraffea et al., 2002; Kikuchi et al., 2002). In most cultured mammalian cells, all ERM proteins are coexpressed and codistributed (Sato et al., 1992; Berryman et al., 1993). Using antisense oligonucleotides, ERM proteins were shown to be functionally redundant, without a quantitatively compensatory increase in their expression levels, in cell–cell/cell–substrate adhesion and the formation of microvilli in cultured cells (Takeuchi et al., 1994). At the whole-body level, the expression levels and combinations of ERM proteins varied significantly depending on cell type (Sato et al., 1992; Berryman et al., 1993; Franck et al., 1993; Doi et al., 1999; Ingraffea et al., 2002), and to date no clear example has been found to indicate the functional redundancy among ERM proteins. Radixin deficiency causes hyperbilirubinemia due to the loss of multidrug resistance protein 2 from the bile canalicular membranes, where microvilli were decreased in number as well as in length (Kikuchi et al., 2002). Radixin was the dominant ERM protein in the liver of wild-type mice, but no compensatory up-regulation of expression level of ezrin or moesin was detected in the Rdx −/− liver.
To discuss the functions of ERM proteins and their possible redundancy in situ, the inner ear provides an advantageous system, as in it there are two distinct types of stereocilia, cochlear and vestibular stereocilia, which play central roles in transmitting acoustic and acceleration stimuli, respectively, into electrical signals. (Roberts et al., 1988; Hudspeth, 1989; Pickles and Corey, 1992; Eatock et al., 1998; Corey, 2003). In wild-type cochlea, before ∼P14, ezrin was detectable in addition to large amounts of radixin in stereocilia, and then progressively disappeared, leaving purely radixin-based stereocilia. In the Rdx −/− cochlea, ezrin was significantly up-regulated, and purely ezrin-based stereocilia developed normally up to ∼P14 and then progressively degenerated. By contrast, in the Rdx +/+ vestibule, stereocilia contained a small amount of ezrin in addition to large amounts of radixin throughout life, whereas in the Rdx −/− vestibule, stereocilia containing up-regulated ezrin developed and was maintained normally throughout the life. These findings would provide the first clear example that these members of the ERM family, ezrin and radixin, function redundantly in situ at the whole-body level: ezrin compensates for radixin deficiency functionally in stereocilia. In Rdx +/+ mice, the cochlear hair cells up to ∼P14 and the vestibular hair cells throughout the life expressed both radixin and ezrin in large and small amounts, respectively, and probably due to the difference in their expression levels or affinity to plasma membranes, radixin was the main contributor to the formation of stereocilia. In these hair cells, in the absence of radixin (Rdx −/− mice), a larger amount of ezrin was recruited, which appeared to compensate for radixin deficiency in the formation of stereocilia.
The molecular mechanism behind the progressive degeneration of the cochlear stereocilia after the onset of hearing (∼P14) remained elusive. At present, two explanations appear possible. First, if the expression of ezrin is genetically programmed to be suppressed in cochlear hair cells after the onset of hearing in the wild-type mice, and if this program is not changed by radixin deficiency, in Rdx −/− mice, cochlear hair cells lack both ezrin and radixin after the onset of hearing, resulting in progressive degeneration of stereocilia. As the total lack of ERM proteins was reported to cause cell death (Kondo et al., 1997), this mechanism can explain why cochlear hair cells were eliminated in older Rdx −/− mice. The second possibility is that sound stimulation induces the progressive degeneration of ezrin-based stereocilia in the Rdx −/− cochlea. It would be possible that these ezrin-based stereocilia are more labile against the noise stimulation (i.e., frequent deflection) than the wild-type radixin-based stereocilia due to the qualitative or quantitative difference between radixin in wild-type stereocilia and ezrin in Rdx −/− stereocilia. Consistently, the noise-induced loss of stereocilia was reported in the wild-type mice as well as humans (Ou et al., 2000). As deflection frequency for vestibular stereocilia is expected to be much lower than for cochlear stereocilia, Rdx −/− vestibule ezrin-based stereocilia could be maintained without degeneration. According to Western blotting of the whole cochlea and vestibule, there were no signs for up-regulation of the expression of ezrin (or moesin), but as it was technically difficult to examine the expression of ERM proteins in individual sensory hair cells, the two above explanations cannot be further evaluated for now. Furthermore, to understand the molecular mechanism behind the progressive degeneration of the cochlear stereocilia, identification of the specific molecular partners of radixin and ezrin, including integral membrane proteins, within stereocilia would be important, although it is again technically difficult due to the small number of stereocilia in the cochlea. Indeed, as described in the results, we performed immunoprecipitation and gel overlay experiments, but were unable to identify the radixin- or ezrin-binding partners in stereocilia. To be successful, any future work will need to overcome several technical difficulties.
Mutations in the actin-bundling protein espin have been reported in jerker mice and DFNB36, a form of nonsyndromic deafness in humans (Grüneberg et al., 1941; Zheng et al., 2000; Belyantseva et al., 2003). Adult jerker mice exhibit impaired hearing and balance dysfunctions, as well as short cochlear and vestibular stereocilia. Analyses of various aged jerker mice revealed apparently normal cochlear stereocilia development until the onset of hearing, after which the stereocilia progressively degenerate (Sjöström and Anniko, 1992). This aspect of jerker mice is very similar to Rdx −/− mice, but jerker mice are completely different from Rdx −/− mice in terms of the balance dysfunction. Because espin is likely to play a role in bundling actin filaments in stereocilia (Loomis et al., 2003), it was suggested that the organization of core actin filaments is critical for the maintenance of stereocilia after the onset of hearing. Furthermore, genetic studies of deafness in mice and humans have identified several genes for other cytoskeleton-related proteins, harmonin, myosin VI/VIIa, whirlin, and SANS, and for cadherin-related proteins, cadherin 23 and protocadherin 15, as causal genes (Avraham et al., 1995; Gibson et al., 1995; Probst et al., 1998; Self et al., 1998; Littlewood Evans and Müller, 2000; Zheng et al., 2000; Alagramam et al., 2001; Di Palma et al., 2001: Petit et al., 2001; Boeda et al., 2002; Mustapha et al., 2002; Karolyi et al., 2003; Kikkawa et al., 2003; Mburu et al., 2003; Weil et al., 2003). In these mice, the formation of not only cochlear, but also vestibular stereocilia was affected, resulting in the deafness associated with imbalance. All these cytoskeleton-related proteins, including ERM proteins and epsin, may act in cooperative ways to form and maintain dynamic stereocilia in cochlear and vestibular sensory hair cells.
To date, three mouse models in which there is profound hearing loss and no vestibular dysfunction have been reported: the deafness (dn) gene mutant mice (Bock and Steel, 1983), Beethoven mice (Vreugde et al., 2002), and the claudin 14-knockout mice (Ben-Yosef et al., 2003). Rdx −/− mice can now be added as a new member of this list. Rdx −/− mice should provide a valuable resource for further analysis of the functional redundancy of ERM proteins at the whole-body level, and of the molecular mechanisms behind the formation and maintenance of highly specialized microvilli, stereocilia; the elucidation of which will shed light on the pathogenesis of human deafness and imbalance.
Materials and methods
Antibodies
Rabbit anti-ERM pAb, TK89, recognized COOH-terminal domains of all ERM proteins almost equally (Kondo et al., 1997; Doi et al., 1999). We previously raised and characterized rat anti-ezrin (M11), anti-radixin (R21), and anti-moesin mAb (M22) (Kondo et al., 1997; Doi et al., 1999). Goat anti-VE-cadherin (C-19) was obtained from Santa Cruz Biotechnology, Inc.
Generation of Rdx−/− mice
Rdx −/− mice were generated as previously reported (Kikuchi et al., 2002). Two independent mouse J1 ES clones (129/Sv) in which the radixin gene was correctly disrupted were injected into C57BL/6 blastocysts, and the resulting chimeras were mated with C57BL/6 mice (Doi et al., 1999).
ABR measurements
ABR measurements were performed in a soundproof room (Zheng et al., 1999). In general, the ABR waveforms were recorded for 12.8 ms at a sampling rate of 40,000 Hz using 50–5,000-Hz filter settings; waveforms recorded from 1,024 stimuli at a frequency of 9 Hz were averaged. ABR waveforms were recorded in decreasing 5-dB SPL intervals from a maximum amplitude until no waveforms could be visualized.
VOR measurements
VOR was measured as described by Iwashita et al. (2001). Head movements were transduced to DC signals using a small angular velocity sensor (Gyrostar; Murata Corporation) that was fixed on the turntable. Eye movements were detected by LED and a CCD camera (C53500; Tokyo Electronic Industry), and eye velocities were calculated online by downloading them onto a computer through a video capture board. Both the head and eye velocity curves were fitted with sinusoidal curves using the least squares criterion, and the gain of eye velocity relative to the head velocity was obtained.
Immunofluorescence microscopy
Temporal bones were removed from 1-, 6-, 14-, or 40-d-old mice, and together with the small holes in the cochlear apical turn and superior semicircular canal, the round and oval windows were opened. The lymphatic space was gently perfused with 10% TCA through a round window (Hayashi et al., 1999; Kitajiri et al., 2004). Samples were then immersed in 10% TCA for 1 h at 4°C, washed three times with PBS, and decalcified with 5% EDTA in PBS for 3 d at 4°C. The cochlea and vestibule were then carefully microdissected, treated with 0.2% Triton X-100 in PBS for 15 min, and soaked in 1% BSA in PBS at RT. They were then incubated with rat anti-ezrin (M11), anti-radixin (R11), or anti-moesin (M22) mAb for 30 min at RT. Samples were washed three times with PBS, followed by a 30-min incubation with Cy3- (Jackson ImmunoResearch Laboratories, Inc.) or Alexa Fluor® 488–conjugated secondary antibody (Molecular Probes, Inc.). After a wash with PBS, they were embedded in 95% glycerol-PBS containing 0.1% paraphenylendiamine and 1% n-propylgalate. Fluorescence images were obtained with a confocal microscope (model LSM 510 META; Carl Zeiss MicroImaging, Inc.) or with a DeltaVision optical sectioning microscope (version 2.10; Applied Precision, Inc.), equipped with an Axioplan2 (Plan Apochromat 63×/1.40 NA oil immersion objective; Carl Zeiss MicroImaging, Inc.) or IX70 (PlanApo 60×/1.40 NA oil immersion objective; Olympus) microscope, respectively.
Immunoblotting
The membranous labyrinths of 5-wk-old mice were dissected under a microscope. Whole organ of Corti and vestibules isolated from each mouse were sonicated in 100 μl SDS sample buffer (50 mM Tris-HCl, pH 6.8, 2% SDS, 20% glycerol, 2% 2-mercaptoethanol, and 0.01% bromophenol blue), applied to SDS-PAGE, and immunoblotted by a blotting detection kit with biotinylated Ig and streptavidin-conjugated alkaline phosphatase (Amersham Biosciences).
Scanning EM
Temporal bones obtained from 1-, 6-, 14-, or 40-d-old mice were fixed using perilymphatic perfusion as described above with 1% glutaraldehyde in 0.1 M phosphate buffer (pH 7.2). They were then washed with phosphate buffer and post-fixed in 1% OsO4 for 2 h, after which they were once again treated with perilymphatic perfusion. The organ of Corti or crista ampullaris was microdissected, dehydrated, critical-point dried, sputter coated, and observed by scanning EM (model S-800 microscope; Hitachi Co.).
Ultrathin-section EM
Samples were processed as previously described, using 2% formaldehyde, 2.5% glutaraldehyde, and 0.1 M sodium cacodylate buffer (pH 7.4) as a fixative (Yonemura et al., 2002).
Acknowledgments
We would like to thank Drs. E. Kikuchi, T.S. Kim, H. Saito, and N. Yamamoto (Kyoto University) for technical help and advice.
This work was supported in part by a Grant-in-Aid for Cancer Research and a Grant-in-Aid for Scientific Research (A) from the Ministry of Education, Science and Culture, Japan to Shoichiro Tsukita, and in part by a Grant-in-Aid for Scientific Research (B) from the Ministry of Education, Science and Culture, Japan, to Sachiko Tsukita.
Note added in proof. While this paper was being reviewed, Pataky et al. presented evidence that radixin is a constituent of stereocilia in hair cells (Pataky, F., R. Pironkova, and A.J. Hudspeth. 2004. Proc. Natl. Acad. Sci. USA. 101:2601–2606).
Abbreviations used in this paper: ABR, auditory-evoked brainstem response; dB, decibel; ERM, ezrin/radixin/moesin; SPL, sound pressure level; VOR, vestibulo-ocular reflex.
References
- Alagramam, K.N., C.L. Murcia, H.Y. Kwon, K.S. Pawlowski, C.G. Wright, and R.P. Woychik. 2001. The mouse Ames waltzer hearing-loss mutant is caused by mutation of Pcdh15, a novel protocadherin gene. Nat. Genet. 27:99–102. [DOI] [PubMed] [Google Scholar]
- Algrain, M., O. Turunen, A. Vaheri, D. Louvard, and M. Arpin. 1993. Ezrin contains cytoskeleton and membrane binding domains accounting for its proposed role as a membrane-cytoskeletal linker. J. Cell Biol. 120:129–140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Avraham, K.B., T. Hasson, K.P. Steel, D.M. Kingsley, L.B. Russell, M.S. Mooseker, N.G. Copeland, and N.A. Jenkins. 1995. The mouse Snell's waltzer deafness gene encodes an unconventional myosin required for structural integrity of inner ear hair cells. Nat. Genet. 11:369–375. [DOI] [PubMed] [Google Scholar]
- Bartles, J.R. 2000. Parallel actin bundles and their multiple actin-bundling proteins. Curr. Opin. Cell Biol. 12:72–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belyantseva, I.A., V. Labay, E.T. Boger, A.J. Griffith, and T.B. Friedman. 2003. Stereocilia: the long and the short of it. Trends Mol. Med. 9:458–461. [DOI] [PubMed] [Google Scholar]
- Ben-Yosef, T., I.A. Belyantseva, T.L. Saunders, E.D. Hughes, K. Kawamoto, C.M. Van Itallie, L.A. Beyer, K. Halsey, D.J. Gardner, E.R. Wilcox, et al. 2003. Claudin 14 knockout mice, a model for autosomal recessive deafness DFNB29, are deaf due to cochlear hair cell degeneration. Hum. Mol. Genet. 12:2049–2061. [DOI] [PubMed] [Google Scholar]
- Berryman, M., Z. Franck, and A. Bretscher. 1993. Ezrin is concentrated in the apical microvilli of a wide variety of epithelial cells whereas moesin is found primarily in endothelial cells. J. Cell Sci. 105:1025–1043. [DOI] [PubMed] [Google Scholar]
- Bock, G.R., and K.P. Steel. 1983. Inner ear pathology in the deafness mutant mouse. Acta Otolaryngol. 96:39–47. [DOI] [PubMed] [Google Scholar]
- Boeda, B., A. El-Amraoui, A. Bahloul, R. Goodyear, L. Daviet, S. Blanchard, I. Perfettini, K.R. Fath, S. Shorte, J. Reiners, et al. 2002. Myosin VIIa, harmonin and cadherin 23, three Usher I gene products that cooperate to shape the sensory hair cell bundle. EMBO J. 21:6689–6699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bretscher, A., K. Edwards, and R.G. Fehon. 2002. ERM proteins and merlin: integrators at the cell cortex. Nat. Rev. Mol. Cell Biol. 3:586–599. [DOI] [PubMed] [Google Scholar]
- Call, L.M., and C.C. Morton. 2002. Continuing to break the sound barrier: genes in hearing. Curr. Opin. Genet. Dev. 12:343–348. [DOI] [PubMed] [Google Scholar]
- Corey, D. 2003. Sensory transduction in the ear. J. Cell Sci. 116:1–3. [DOI] [PubMed] [Google Scholar]
- Denman-Johnson, K., and A. Forge. 1999. Establishment of hair bundle polarity and orientation in the developing vestibular system of the mouse. J. Neurocytol. 28:821–835. [DOI] [PubMed] [Google Scholar]
- DeRosier, D.J., and L.G. Tilney. 2000. F-actin bundles are derivatives of microvilli: What does this tell us about how bundles might form? J. Cell Biol. 148:1–6. [PMC free article] [PubMed] [Google Scholar]
- DeRosier, D.J., L.G. Tilney, and E. Egelman. 1980. Actin in the inner ear: the remarkable structure of the stereocilium. Nature. 287:291–296. [DOI] [PubMed] [Google Scholar]
- Di Palma, F., R.H. Holme, E.C. Bryda, I.A. Belyantseva, R. Pellegrino, B. Kachar, K.P. Steel, and K. Noben-Trauth. 2001. Mutations in Cdh23, encoding a new type of cadherin, cause stereocilia disorganization in waltzer, the mouse model for Usher syndrome type 1D. Nat. Genet. 27:103–107. [DOI] [PubMed] [Google Scholar]
- Doi, Y., M. Itoh, S. Yonemura, S. Ishihara, H. Takano, T. Noda, Sh. Tsukita, and Sa. Tsukita. 1999. Normal development of mice and unimpaired cell adhesion/cell motility/actin-based cytoskeleton without compensatory up-regulation of ezrin or radixin in moesin gene knockout. J. Biol. Chem. 274:2315–2321. [DOI] [PubMed] [Google Scholar]
- Drenckhahn, D., K. Engel, D. Höfer, C. Merte, L. Tilney, and M. Tilney. 1991. Three different actin filament assemblies occur in every hair cell: each contains a specific actin crosslinking protein. J. Cell Biol. 112:641–651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dumont, R.A., Y.D. Zhao, J.R. Holt, M. Bahler, and P.G. Gillespie. 2002. Myosin-I isozymes in neonatal rodent auditory and vestibular epithelia. J. Assoc. Res. Otolaryngol. 3:375–389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eatock, R.A., A. Rusch, A. Lysakowski, and M. Saeki. 1998. Hair cells in mammalian utricles. Otolaryngol. Head Neck Surg. 119:172–181. [DOI] [PubMed] [Google Scholar]
- Erven, A., M.J. Skynner, K. Okumura, S. Takebayashi, S.D.M. Brown, K.P. Steel, and N.D. Allen. 2002. A novel stereocilia defect in sensory hair cells of the deaf mouse mutant Tasmanian devil. Eur. J. Neurosci. 16:1433–1441. [DOI] [PubMed] [Google Scholar]
- Fortnum, H., and A. Davis. 1997. Epidemiology of permanent childhood hearing impairment in Trent Region, 1985–1993. Br. J. Audiol. 31:409–446. [DOI] [PubMed] [Google Scholar]
- Franck, Z., R. Gary, and A. Bretscher. 1993. Moesin, like ezrin, colocalizes with actin in the cortical cytoskeleton in cultured cells, but its expression is more variable. J. Cell Sci. 105:219–231. [DOI] [PubMed] [Google Scholar]
- Furukawa, R., and M. Fechheimer. 1997. The structure, function, and assembly of actin filament bundles. Int. Rev. Cytol. 175:29–90. [DOI] [PubMed] [Google Scholar]
- Gibson, F., J. Walsh, P. Mburu, A. Varela, K.A. Brown, M. Antonio, K.W. Beisel, K.P. Steel, and S.D. Brown. 1995. A type VII myosin encoded by the mouse deafness gene shaker-1. Nature. 374:62–64. [DOI] [PubMed] [Google Scholar]
- Gillespie, P.G., and R.G. Walker. 2001. Molecular basis of mechanosensory transduction. Nature. 413:194–202. [DOI] [PubMed] [Google Scholar]
- Grüneberg, H., J.B. Burnett, and G.D. Snell. 1941. The origin of jerker, a new gene mutation of the house mouse, and linkage studies made with it. Proc. Natl. Acad. Sci. USA. 27:562–565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi, K., S. Yonemura, T. Matsui, Sa. Tsukita, and Sh. Tsukita. 1999. Immunofluorescence detection of ezrin/radixin/moesin (ERM) proteins with their carboxyl-terminal threonine phosphorylated in cultured cells and tissues. J. Cell Sci. 112:1149–1158. [DOI] [PubMed] [Google Scholar]
- Heiska, L., K. Alfthan, M. Gröholm, P. Vilja, A. Vaheri, and O. Carpén. 1998. Association of ezrin with intercellular adhesion molecule-1 and -2 (ICAM-1 and ICAM-2). Regulation by phosphatidylinositol 4,5-bisphosphate. J. Biol. Chem. 273:21893–21900. [DOI] [PubMed] [Google Scholar]
- Helander, T.S., O. Carpén, O. Turunen, P.E. Kovanen, A. Vaheri, and T. Timonen. 1996. ICAM-2 redistributed by ezrin as a target for killer cells. Nature. 382:265–268. [DOI] [PubMed] [Google Scholar]
- Henry, M.D., C. Gonzalez Agosti, and F. Solomon. 1995. Molecular dissection of radixin: distinct and interdependent functions of the amino- and carboxy-terminal domains. J. Cell Biol. 129:1007–1022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirao, M., N. Sato, T. Kondo, S. Yonemura, M. Monden, T. Sasaki, Y. Takai, Sh. Tsukita, and Sa. Tsukita. 1996. Regulation mechanism of ERM (ezrin/radixin/moesin) protein/plasma membrane association: possible involvement of phosphatidylinositol turnover and Rho-dependent signaling pathway. J. Cell Biol. 135:37–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holme, R.H., and K.P. Steel. 1999. Genes involved in deafness. Curr. Opin. Genet. Dev. 9:309–314. [DOI] [PubMed] [Google Scholar]
- Hudspeth, A.J. 1989. How the ear's works work. Nature. 341:397–404. [DOI] [PubMed] [Google Scholar]
- Ingraffea, J., D. Reczek, and A. Bretscher. 2002. Distinct cell type-specific expression of scaffolding proteins EBP50 and E3KARP: EBP50 is generally expressed with ezrin in specific epithelia, whereas E3KARP is not. Eur. J. Cell Biol. 81:61–68. [DOI] [PubMed] [Google Scholar]
- Iwashita, M., R. Kanai, K. Funabiki, K. Matsuda, and T. Hirano. 2001. Dynamic properties, interactions and adaptive modifications of vestibulo-ocular reflex and optokinetic response in mice. Neurosci. Res. 39:299–311. [DOI] [PubMed] [Google Scholar]
- Karolyi, I.J., F.J. Probst, L. Beyer, H. Odeh, G. Dootz, K.B. Cha, D.M. Martin, K.B. Avraham, D. Kohrman, D.F. Dolan, et al. 2003. Myo15 function is distinct from Myo6, Myo7a and pirouette genes in development of cochlear stereocilia. Hum. Mol. Genet. 12:2797–2805. [DOI] [PubMed] [Google Scholar]
- Kikkawa, Y., H. Shitara, S. Wakana, Y. Kohara, T. Takada, M. Okamoto, C. Taya, K. Kamiya, Y. Yoshikawa, H. Tokano, et al. 2003. Mutations in a new scaffold protein Sans cause deafness in Jackson shaker mice. Hum. Mol. Genet. 12:453–461. [DOI] [PubMed] [Google Scholar]
- Kikuchi, S., M. Hata, K. Fukumoto, Y. Yamane, T. Matsui, A. Tamura, S. Yonemura, H. Yamagishi, D. Keppler, Sh. Tsukita, and Sa. Tsukita. 2002. Radixin deficiency causes conjugated hyperbilirubinemia with loss of Mrp2 from bile canalicular membranes. Nat. Genet. 31:320–325. [DOI] [PubMed] [Google Scholar]
- Kitajiri, S., M. Furuse, K. Morita, Y. Saishin-Kiuchi, H. Kido, J. Ito, and Sh. Tsukita. 2004. Expression patterns of claudins, tight junction adhesion molecules, in the inner ear. Hear. Res. 187:25–34. [DOI] [PubMed] [Google Scholar]
- Kondo, T., K. Takeuchi, Y. Doi, S. Yonemura, S. Nagata, Sh. Tsukita, and Sa. Tsukita. 1997. ERM (ezrin/radixin/moesin)-based molecular mechanism of microvillar breakdown at an early stage of apoptosis. J. Cell Biol. 139:749–758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang, Y., A. Wang, I.A. Belyantseva, D.W. Anderson, F.J. Probst, T.D. Barber, W. Miller, J.W. Touchman, L. Jin, S.L. Sullivan, et al. 1999. Characterization of the human and mouse unconventional myosin XV genes responsible for hereditary deafness. Genomics. 61:243–258. [DOI] [PubMed] [Google Scholar]
- Lim, D.J., and M. Anniko. 1985. Developmental morphology of the mouse inner ear. A scanning electron microscopic observation. Acta Otolaryngol. Suppl. 422:1–69. [PubMed] [Google Scholar]
- Littlewood Evans, A., and U. Müller. 2000. Stereocilia defects in the sensory hair cells of the inner ear in mice deficient in integrin α8β1. Nat. Genet. 24:424–428. [DOI] [PubMed] [Google Scholar]
- Loomis, P.A., L. Zheng, G. Sekerkova, B. Changyaleket, E. Mugnaini, and J.R. Bartles. 2003. Espin cross-links cause the elongation of microvillus-type parallel actin bundles in vivo. J. Cell Biol. 163:1045–1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackay, D.J.G., F. Esch, H. Furthmayr, and A. Hall. 1997. Rho- and rac-dependent assembly of focal adhesion complexes and actin filaments in permeabilized fibroblasts: an essential role for ezrin/radixin/moesin proteins. J. Cell Biol. 138:927–938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mangeat, P., C. Roy, and M. Martin. 1999. ERM proteins in cell adhesion and membrane dynamics. Trends Cell Biol. 9:187–192. [DOI] [PubMed] [Google Scholar]
- Martin, M., C. Andréoli, A. Sahuquet, P. Montcourrier, M. Algrain, and P. Mangeat. 1995. Ezrin NH2-terminal domain inhibits the cell extension activity of the COOH-terminal domain. J. Cell Biol. 128:1081–1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsui, T., S. Yonemura, Sh. Tsukita, and Sa. Tsukita. 1999. Activation of ERM proteins in vivo by Rho involves phosphatidyl-inositol 4-phosphate 5-kinase and not ROCK kinases. Curr. Biol. 9:1259–1262. [DOI] [PubMed] [Google Scholar]
- Maudsley, S., A.M. Zamah, N. Rahman, J.T. Blitzer, L.M. Luttrell, R.J. Lefkowitz, and R.A. Hall. 2000. Platelet-derived growth factor receptor association with Na+/H+ exchanger regulatory factor potentiates receptor activity. Mol. Cell. Biol. 20:8352–8363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mburu, P., M. Mustapha, A. Varela, D. Weil, A. El-Amraoui, R.H. Holme, A. Rump, R.E. Hardisty, S. Blanchard, R.S. Coimbra, et al. 2003. Defects in whirlin, a PDZ domain molecule involved in stereocilia elongation, cause deafness in the whirler mouse and families with DFNB31. Nat. Genet. 34:421–428. [DOI] [PubMed] [Google Scholar]
- Morton, N.E. 1991. Genetic epidemiology of hearing impairment. Ann. NY Acad. Sci. 630:16–31. [DOI] [PubMed] [Google Scholar]
- Murthy, A., C. Gonzalez-Agosti, E. Cordero, D. Pinney, C. Candia, F. Solomon, J. Gusella, and V. Ramesh. 1998. NHE-RF, a regulatory cofactor for Na+-H+ exchange, is a common interactor for merlin and ERM (MERM) proteins. J. Biol. Chem. 273:1273–1276. [DOI] [PubMed] [Google Scholar]
- Mustapha, M., E. Chouery, D. Torchard-Pagnez, S. Nouaille, A. Khrais, F.N. Sayegh, A. Megarbane, J. Loiselet, M. Lathrop, C. Petit, and D. Weil. 2002. A novel locus for Usher syndrome type I, USH1G, maps to chromosome 17q24-25. Hum. Genet. 110:348–350. [DOI] [PubMed] [Google Scholar]
- Ou, H.C., B.A. Bohne, and G.W. Harding. 2000. Noise damage in the C57BL/CBA mouse cochlea. Hear. Res. 145:111–122. [DOI] [PubMed] [Google Scholar]
- Petit, C., J. Levilliers, and J.P. Hardelin. 2001. Molecular genetics of hearing loss. Annu. Rev. Genet. 35:589–646. [DOI] [PubMed] [Google Scholar]
- Pickles, J.O., and D.P. Corey. 1992. Mechanoelectrical transduction by hair cells. Trends Neurosci. 15:254–259. [DOI] [PubMed] [Google Scholar]
- Probst, F.J., and S.A. Camper. 1999. The role of mouse mutants in the identification of human hereditary hearing loss genes. Hear. Res. 130:1–6. [DOI] [PubMed] [Google Scholar]
- Probst, F.J., R.A. Fridell, Y. Raphael, T.L. Saunders, A. Wang, Y. Liang, R.J. Morell, J.W. Touchman, R.H. Lyons, K. Noben-Trauth, et al. 1998. Correction of deafness in shaker-2 mice by an unconventional myosin in a BAC transgene. Science. 280:1444–1447. [DOI] [PubMed] [Google Scholar]
- Reczek, D., M. Berryman, and A. Bretscher. 1997. Identification of EBP50: A PDZ-containing phosphoprotein that associates with members of the ezrin-radixin-moesin family. J. Cell Biol. 139:169–179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts, W.M., J. Howard, and A.J. Hudspeth. 1988. Hair cells: transduction, tuning, and transmission in the inner ear. Annu. Rev. Cell Biol. 4:63–92. [DOI] [PubMed] [Google Scholar]
- Sato, N., N. Funayama, A. Nagafuchi, S. Yonemura, Sa. Tsukita, and Sh. Tsukita. 1992. A gene family consisting of ezrin, radixin and moesin. Its specific localization at actin filament/plasma membrane association sites. J. Cell Sci. 103:131–143. [DOI] [PubMed] [Google Scholar]
- Self, T., M. Mahony, J. Fleming, J. Walsh, S.D. Brown, and K.P. Steel. 1998. Shaker-1 mutations reveal roles for myosin VIIA in both development and function of cochlear hair cells. Development. 125:557–566. [DOI] [PubMed] [Google Scholar]
- Serrador, J. M., J.L. Alonso-Lebrero, M.A. del Pozo, H. Furthmayr, R. Schwartz-Albiez, R.J. Calvo, F. Lozano, and F. Sánchez-Madrid. 1997. Moesin interacts with the cytoplasmic region of intercellular adhesion molecule-3 and is redistributed to the uropod of T lymphocytes during cell polarization. J. Cell Biol. 138:1409–1423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaw, A.S. 2001. FERMing up the synapse. Immunity. 15:683–686. [DOI] [PubMed] [Google Scholar]
- Short, D.B., K.W. Trotter, D. Reczek, S.M. Kreda, A. Bretscher, R.C. Boucher, M.J. Stutts, and S.L. Milgram. 1998. An apical PDZ protein anchors the cystic fibrosis transmembrane conductance regulator to the cytoskeleton. J. Biol. Chem. 273:19797–19801. [DOI] [PubMed] [Google Scholar]
- Sjöström, B., and M. Anniko. 1992. Genetically induced inner ear degeneration. A structural and functional study. Acta Otolaryngol. Suppl. 493:141–146. [PubMed] [Google Scholar]
- Skowron, J.F., W.M. Bement, and M.S. Mooseker. 1998. Human brush border myosin-I and myosin-Ic expression in human intestine and Caco-2BBe cells. Cell Motil. Cytoskeleton. 41:308–324. [DOI] [PubMed] [Google Scholar]
- Sobin, A., and A. Flock. 1983. Immunohistochemical identification and localization of actin and fimbrin in vestibular hair cells in the normal guinea pig and in a strain of the waltzing guinea pig. Acta Otolaryngol. 96:407–412. [DOI] [PubMed] [Google Scholar]
- Steel, K.P., and C.J. Kros. 2001. A genetic approach to understanding auditory function. Nat. Genet. 27:143–149. [DOI] [PubMed] [Google Scholar]
- Takeuchi, K., N. Sato, H. Kasahara, N. Funayama, A. Nagafuchi, S. Yonemura, Sa. Tsukita, and Sh. Tsukita. 1994. Perturbation of cell adhesion and microvilli formation by antisense oligonucleotides to ERM family members. J. Cell Biol. 125:1371–1384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tekin, M., K.S. Arnos, and A. Pandya. 2001. Advances in hereditary deafness. Lancet. 358:1082–1090. [DOI] [PubMed] [Google Scholar]
- Tilney, M.S., L.G. Tilney, R.E. Stephens, C. Merte, D. Drenckhahn, D.A. Cotanche, and A. Bretscher. 1989. Preliminary biochemical characterization of the stereocilia and cuticular plate of hair cells of the chick cochlea. J. Cell Biol. 109:1711–1723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsukita, Sa., and S. Yonemura. 1999. Cortical actin organization: lessons from ERM (ezrin/radixin/moesin) proteins. J. Biol. Chem. 274:34507–34510. [DOI] [PubMed] [Google Scholar]
- Tsukita, Sa., K. Oishi, N. Sato, J. Sagara, A. Kawai, and Sh. Tsukita. 1994. ERM family members as molecular linkers between the cell surface glycoprotein CD44 and actin-based cytoskeletons. J. Cell Biol. 126:391–401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turunen, O., T. Wahlström, and A. Vaheri. 1994. Ezrin has a COOH-terminal actin-binding site that is conserved in the ezrin protein family. J. Cell Biol. 126:1445–1453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vreugde, S., A. Erven, C.J. Kros, W. Marcotti, H. Fuchs, K. Kurima, E.R. Wilcox, T.B. Friedman, A.J. Griffith, R. Balling, et al. 2002. Beethoven, a mouse model for dominant, progressive hearing loss DFNA36. Nat. Genet. 30:257–258. [DOI] [PubMed] [Google Scholar]
- Wada, T., Y. Wakabayashi, S. Takahashi, T. Ushiki, Y. Kikkawa, H. Yonekawa, and R. Kominami. 2001. A point mutation in a cadherin gene, Cdh23, causes deafness in a novel mutant, Waltzer mouse niigata. Biochem. Biophys. Res. Commun. 283:113–117. [DOI] [PubMed] [Google Scholar]
- Weil, D., A. El-Amraoui, S. Masmoudi, M. Mustapha, Y. Kikkawa, S. Laine, S. Delmaghani, A. Adato, S. Nadifi, Z.B. Zina, et al. 2003. Usher syndrome type I G (USH1G) is caused by mutations in the gene encoding SANS, a protein that associates with the USH1C protein, harmonin. Hum. Mol. Genet. 12:463–471. [DOI] [PubMed] [Google Scholar]
- Yonemura, S., M. Hirao, Y. Doi, N. Takahashi, T. Kondo, Sa. Tsukita, and Sh. Tsukita. 1998. Ezrin/radixin/moesin (ERM) proteins bind to a positively charged amino acid cluster in the juxta-membrane cytoplasmic domain of CD44, CD43, and ICAM-2. J. Cell Biol. 140:885–895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yonemura, S., Sa. Tsukita, and Sh. Tsukita. 1999. Direct involvement of ezrin/radixin/moesin (ERM)-binding membrane proteins in the organization of microvilli in collaboration with activated ERM proteins. J. Cell Biol. 145:1497–1509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yonemura, S., T. Matsui, Sh. Tsukita, and Sa. Tsukita. 2002. Rho-dependent and -independent activation mechanisms of ezrin/radixin/moesin proteins: an essential role for polyphosphoinositides in vivo. J. Cell Sci. 115:2569–2580. [DOI] [PubMed] [Google Scholar]
- Yun, C.H.C., S. Oh, M. Zizak, D. Steplock, S. Tsao, C.-M. Tse, E.J. Weinman, and M. Donowitz. 1997. cAMP-mediated inhibition of the epithelial brush border Na+/H+ exchanger, NHE3, requires an associated regulatory protein. Proc. Natl. Acad. Sci. USA. 94:3010–3015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng, L., G. Sekerkova, K. Vranich, L.G. Tilney, E. Mugnaini, and J.R. Bartles. 2000. The deaf jerker mouse has a mutation in the gene encoding the espin actin-bundling proteins of hair cell stereocilia and lacks espins. Cell. 102:377–385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng, Q.Y., K.R. Johnson, and L.C. Erway. 1999. Assessment of hearing in 80 inbred strains of mice by ABR threshold analyses. Hear. Res. 130:94–107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zuo, J. 2002. Transgenic and gene targeting studies of hair cell function in mouse inner ear. J. Neurobiol. 53:286–305. [DOI] [PubMed] [Google Scholar]