

Abstract
The dynein regulatory complex (DRC) is an important intermediate in the pathway that regulates flagellar motility. To identify subunits of the DRC, we characterized a Chlamydomonas motility mutant obtained by insertional mutagenesis. The pf2-4 mutant displays an altered waveform that results in slow swimming cells. EM analysis reveals defects in DRC structure that can be rescued by reintroduction of the wild-type PF2 gene. Immunolocalization studies show that the PF2 protein is distributed along the length of the axoneme, where it is part of a discrete complex of polypeptides. PF2 is a coiled-coil protein that shares significant homology with a mammalian growth arrest–specific gene product (Gas11/Gas8) and a trypanosome protein known as trypanin. PF2 and its homologues appear to be universal components of motile axonemes that are required for DRC assembly and the regulation of flagellar motility. The expression of Gas8/Gas11 transcripts in a wide range of tissues may also indicate a potential role for PF2-related proteins in other microtubule-based structures.
Keywords: dynein; flagella; Gas8; Gas11; TLTF
Introduction
Cilia and flagella are homologous, microtubule-based organelles found on diverse cell types ranging from spermatozoa to the epithelia lining the respiratory and reproductive tracts. The driving force for motility is supplied by the axonemal dynein ATPases. Defects in the dynein motors, accessory proteins that regulate dynein activity, or other components that control axoneme assembly have profound consequences. In vertebrates, these include infertility, respiratory and kidney disease, and defects in the determination of the left–right body axis during embryonic development (Afzelius, 1995; Nonaka et al., 1998; Sapiro et al., 2002; Pazour and Rosenbaum, 2002; for review see Supp et al., 2000). In motile axonemes, dyneins form the inner and outer arms that are bound to the outer doublet microtubules and interact transiently with adjacent doublet microtubules to generate the force for interdoublet sliding (for review see Kamiya, 2002). Because dyneins can generate sliding forces in only one direction (Sale and Satir, 1977; Fox and Sale, 1987), the activity of the dynein arms must be coordinated both spatially and temporally to generate an efficient flagellar waveform.
Both structural and genetic evidence have implicated the central pair (CP)* and radial spoke (RS) structures as key regulators of dynein activity. Mutations that disrupt the CP or RS structures result in flagellar paralysis under physiological conditions (Witman et al., 1978), by a mechanism that appears to involve global inactivation of the dynein arms (Huang et al., 1982; Smith and Sale, 1992). One hypothesis is that the CP projections act like a distributor that periodically sweeps past the RS heads during CP rotation, inducing a local signal that is ultimately transmitted to the dynein arms (Omoto and Kung, 1979; Omoto et al., 1999). In the absence of these signals, other axonemal components must inhibit dynein activity. Evidence for such an inhibitory control system has been obtained by the isolation of extragenic suppressor mutations that restore partial motility to CP/RS-defective strains without restoring the missing structures (Huang et al., 1982; Piperno et al., 1992). Two distinct groups of suppressor mutations have been identified. One group alters the activity of the outer dynein arms by mutations in dynein heavy chain domains that regulate cross-bridge activity (Huang et al., 1982; Porter et al., 1994; Rupp et al., 1996). The second group alters the assembly of the inner dynein arms and/or a subset of closely associated polypeptides known as the dynein regulatory complex (DRC) (Huang et al., 1982; Piperno et al., 1992, 1994; Porter et al., 1992; Gardner et al., 1994). The mechanism by which the DRC regulates dynein activity is still unclear.
The DRC contains at least seven polypeptides that are tightly associated with the outer doublet microtubules (Piperno et al., 1994), but until this study, the identity of any of these DRC subunits has been unknown. The DRC has also been correlated with a crescent-shaped structure located near the base of the second RS, close to the site of attachment for the nexin link (Burgess et al., 1991; Mastronarde et al., 1992; Gardner et al., 1994; Woolley, 1997). This structure appears to be ideally situated to mediate local signals between the CP/RS complex, the nexin links, and the dynein arms, and it is altered or missing in DRC-defective strains (Mastronarde et al., 1992; Gardner et al., 1994). It is unclear whether the local signals that control dynein arm activity are chemical, mechanical, or both. However, both pharmacological and biochemical evidence suggest that ciliary and flagellar motility is regulated in part by reversible phosphorylation of axonemal components (for review see Porter and Sale, 2000). Several kinases and phosphatases are anchored in the axoneme (San Agustin and Witman, 1994; Yang and Sale, 2000; Yang et al., 2000; Gaillard et al., 2001; Smith, 2002), and numerous axonemal phosphoproteins have been detected (Piperno et al., 1981). Recent studies also show that components of the dynein arms are targets of reversible phosphorylation, and that changes in their phosphorylation state can modulate dynein activity and flagellar motility (Hamasaki et al., 1991; Habermacher and Sale, 1997; King and Dutcher, 1997; Yang and Sale, 2000).
To identify axonemal components that regulate dynein activity, we used insertional mutagenesis procedures in Chlamydomonas (Tam and Lefebvre, 1993) to recover “tagged” motility mutants with characteristics similar to previously described DRC mutants (Huang et al., 1982; Gardner et al., 1994). Structural, functional, and genetic evidence indicate that we have cloned the PF2 locus, which encodes DRC subunit 4, a highly coiled-coil protein of ∼55 kD that is tightly associated with the outer doublet microtubules. Localization of an epitope-tagged PF2 construct indicates that PF2 is uniformly distributed along the length of the axoneme and also associated with the basal body region. Interestingly, homologues of PF2 have recently been identified in diverse organisms, ranging from trypanosomes (trypanin) to humans (Gas8/Gas11), where they have been proposed to play important, but poorly understood, roles in both cell motility and growth arrest (Brenner et al., 1989; Hutchings et al., 2002; Yeh et al., 2002). Our study strongly suggests that these PF2/trypanin/Gas8/Gas11 homologues are part of a conserved DRC complex involved in the regulation of axonemal motility in multiple species. The presence of Gas8/Gas11 transcripts in growth-arrested cells and tissues that do not assemble motile axonemes (Brenner et al., 1989; Whitmore et al., 1998; Yeh et al., 2002; unpublished data) may also indicate a possible role for the DRC in other microtubule-based organelles.
Results
Recovery of a tagged pf2 allele
A collection of motility mutants generated by insertional mutagenesis was screened for strains with abnormal swimming behaviors to identify new loci involved in the regulation of motility. One strain, 9B11, swam more slowly than wild-type cells (∼51 ± 14 μm/s vs. 139 ± 24 μm/s) and displayed an aberrant flagellar waveform similar to those seen in inner arm and DRC mutant strains (Brokaw and Kamiya, 1987; Gardner et al., 1994). Direct comparison between different strains by phase contrast microscopy indicated that 9B11 was most similar to the DRC mutant pf2. 9B11 was backcrossed to a nit − strain with wild-type motility to verify that the 9B11 motility defect was linked to the NIT1 gene used as a selectable marker. Analysis of the tetrad progeny confirmed that the 9B11 motility phenotype cosegregated with the nit + phenotype (<6.3 cM apart).
Cloning and characterization of the PF2 gene
To identify the gene that was disrupted in 9B11, a fragment of genomic DNA flanking the site of plasmid insertion was obtained by screening a size-fractionated minilibrary with a probe derived from the 3′ end of the NIT1 gene (see Materials and methods; Fig. 1 A). Southern blots of wild-type and 9B11 genomic DNA probed with the flanking DNA (flanking clone 1 [FC-1]) confirmed that this DNA was located near the site of the mutation in 9B11 (Fig. 1 B).
Figure 1.
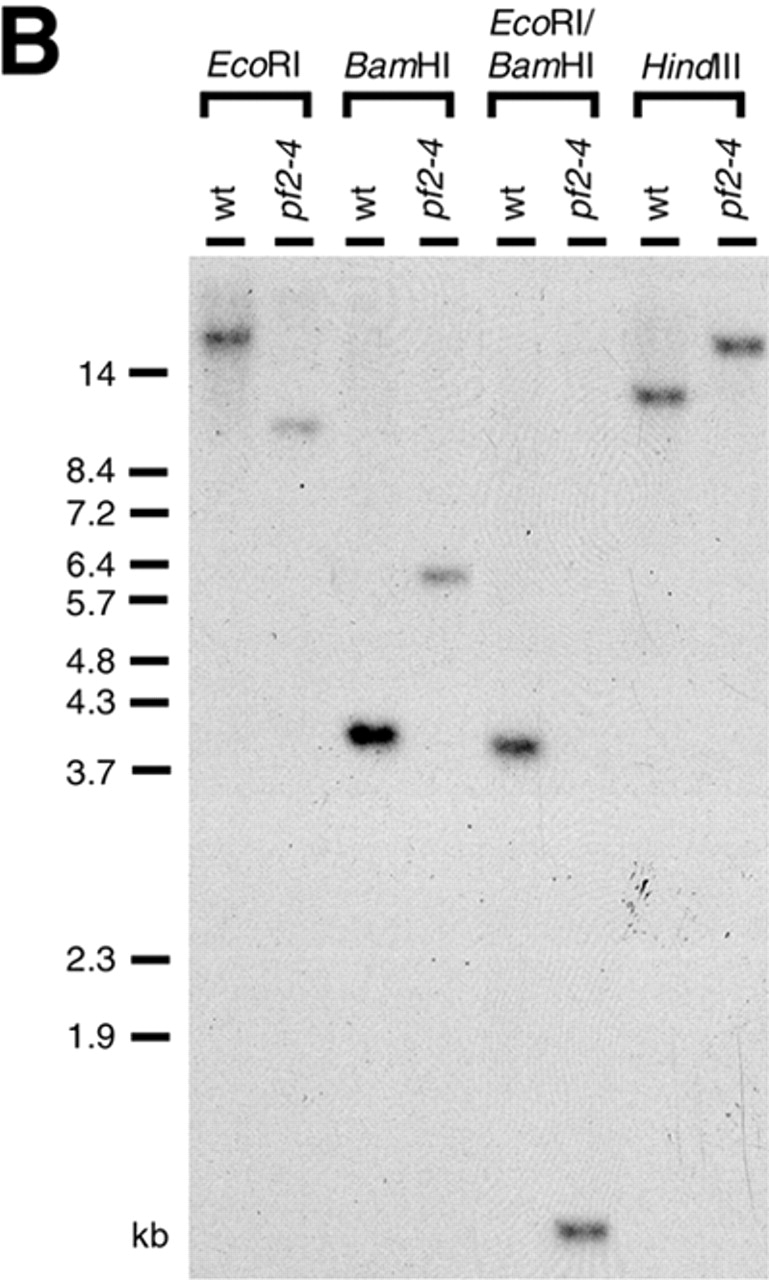


Cloning the PF2 gene. (A) Partial restriction maps of the region containing the PF2 gene from wild-type and pf2-4 (9B11). Also indicated is the location of the NIT1 plasmid insertion in 9B11, now known as pf2-4, and the position of the flanking clone FC-1 (black box) representing the NotI/BamHI restriction fragment recovered from the 9B11 minilibrary. S, SacI sites; N, NotI sites. (B) Southern blot of wild-type (wt) and pf2-4 genomic DNA digested with the indicated restriction enzymes and hybridized with FC-1. (C) Alignment of three overlapping phage clones recovered with FC-1 (black boxes) and two plasmid clones, derived from λG1, that contain the PF2 gene. Plasmid p9-3HA is the epitope-tagged PF2 construct. Also shown are the number of rescued strains obtained by cotransformation of pf2-4 and pf2-1 with the selected clones. nd, not determined.
FC-1 was then used to screen a wild-type library and recover more than 30 kb of genomic DNA (Fig. 1 C). RFLP mapping procedures revealed that this DNA was closely linked to the PF2 locus on linkage group XI (see Materials and methods). These results and the 9B11 motility phenotype described above suggested that 9B11 might represent a new pf2 mutation. To test if any of the phage clones contained a full-length copy of the PF2 gene, three clones were analyzed for their ability to rescue the 9B11 and pf2 motility defects by cotransformation. Two clones, λG2 and λJ1, were able to restore wild-type motility to either 9B11 or pf2-1 cells (Fig. 1 C). The 9B11 strain was therefore renamed pf2-4. The λM2 clone, which shares ∼50% overlap with λG2, failed to rescue the motility defects, suggesting that the PF2 gene must extend beyond the limits of this clone.
Selected restriction fragments from the phage clones were used to probe Southern and Northern blots to define the boundaries of the PF2 gene. Genomic Southern blots probed with subclones A–D indicated that the NIT1 plasmid inserted into a 3.5-kb SacI restriction fragment without significant deletion of the surrounding genomic DNA in pf2-4 (Fig. 2 A). Subclones A–D were also hybridized to Northern blots loaded with total RNA isolated from wild-type and mutant cells both before and 45 min after deflagellation to delineate the boundaries of the PF2 transcription unit and determine the size of the PF2 transcript. Deflagellation has previously been shown to induce up-regulation of transcripts that encode flagellar proteins (for review see Lefebvre and Rosenbaum, 1986). Probes B–D identified a single ∼2.5-kb transcript that was up-regulated after deflagellation in wild-type cells and missing in pf2 strains (Fig. 2 B). Given the location of the NIT1 insertion in pf2-4 and the expression pattern of the 2.5-kb transcript, we identified this region as the site of the PF2 transcription unit. Probes A and B also hybridized to a smaller ∼2.2-kb transcript that was expressed in both wild-type and pf2 cells and showed homology to a 50S ribosomal subunit (Fig. 2 A).
Figure 2.



Analysis of the PF2 transcription unit. (A) Partial restriction map of wild-type genomic DNA in the region of the PF2 transcription unit. Also shown are selected SacI fragments (A, C, and D) and a NotI fragment (B) that were used to map the boundaries of the plasmid insertion and determine the size of the transcription unit. (B) Northern blot loaded with total RNA isolated from wild-type (wt) and pf2 mutant cells before (0) and 45 min after deflagellation. This blot was hybridized with probe C, which recognized an ∼2.5-kb transcript that is up-regulated in wild-type cells and missing in pf2 mutants. (B, bottom) Loading control. The blot was rehybridized with a CRY1 probe, which encodes the ribosomal S14 protein subunit (Nelson et al., 1994). (C) Diagram of the intron–exon structure of the PF2 transcription unit. Open rectangles indicate exons and solid lines indicate introns. The predicted translation start (ATG) site and the putative polyadenylation signal (Poly A) are indicated. Also shown are the single nucleotide alterations that resulted in the pf2-1 and pf2-2 mutations, and the location of the epitope tag in exon 12.
The PF2 transcription unit contains 12 exons and 11 introns spanning ∼5.7 kb of genomic DNA (Fig. 2 C). All splice junctions were confirmed by sequence analysis of RT-PCR products derived from the wild-type PF2 transcript. A subsequent search of the Chlamydomonas EST database has also identified a cDNA clone (BE056668) whose partial sequence closely matches the predicted sequence of the PF2 transcript. PCR with gene-specific primers was then used to identify the mutations in two previously isolated pf2 alleles, pf2-1 and pf2-2 (Ebersold et al., 1962; Huang et al., 1982). Sequence analysis of pf2-1 DNA revealed a single base (G→T) mutation that results in an in-frame stop codon in exon 7 (Fig. 2 C). Interestingly, probes B and C (Fig. 2 A), which contain the DNA encoding exons 1–6, did not detect any transcript on Northern blots of pf2-1 total RNA (Fig. 2 B), which suggested that the pf2-1 transcript may be unstable and/or rapidly degraded. Sequence analysis of pf2-2 DNA revealed a single base (G→A) mutation that altered the donor splice site of exon 1 (Fig. 2 C). Failure to recognize the altered sequence as a splice site would result in translation of the adjacent intron, which contains an in-frame stop codon 30 bp downstream of the altered splice site.
Suppression of flagellar paralysis by pf2-4
pf2 mutants belong to a group of bypass suppressor mutations that are capable of restoring motility to paralyzed RS or CP mutants without restoring the missing RS or CP structures (Huang et al., 1982; Piperno et al., 1994). pf2 mutations also alter the assembly of the DRC polypeptides, which are thought to be regulatory components located between the RS and the dynein arms (Table I ; Piperno et al., 1992; Gardner et al., 1994). To test whether pf2-4 can also act as a suppressor, pf2-4 was crossed to a paralyzed, RS mutant, pf14, and the motility phenotypes of the resulting progeny were analyzed. The pf2-4 pf14 cells had a motility phenotype that differed from either parent strain or wild-type cells (Table II). The double mutants displayed vigorous flagellar movements, but they could generate little or no forward progress. This type of motility is similar to that seen with other suppressed strains containing defects in the DRC and RS (Huang et al., 1982; Piperno et al., 1992; unpublished data).
Table I. Dynein regulatory complex subunits.
| Subunit | Molecular massa | Polypeptide deficiency ina , b | Gene product of |
|---|---|---|---|
|
|
kD |
|
|
| 1 | 83 | pf3, sup-pf-5 | PF3 c |
| 2 | 70 | pf3, sup-pf-5 | Unknown |
| 3 | 62 | pf2, sup-pf-3 | Unknown |
| 4 | 55 | pf2, sup-pf-3 | PF2 c , d |
| 5 | 40 | pf2, pf3, sup-pf-3, sup-pf-4, sup-pf-5 | SUP-PF-4 b , c |
| 6 | 29 | pf2, pf3, sup-pf-3, sup-pf-4, sup-pf-5 | Unknown |
| 7 | 192 | pf2 | Unknown |
Piperno et al., 1994.
Huang et al., 1982.
Putative gene products based on dikaryon rescue (Piperno et al., 1994).
Identified in this work.
Table II. Strains used in this work.
| Strain | Structural defect | Motility phenotype | Reference |
|---|---|---|---|
| Wild-type (137c) | None | Fast forward swimming | Harris, 1989 |
| nit1Δ (A54e18) | None | Fast forward swimming | Nelson et al., 1994 |
| pf2-1 (CC1025) | DRC, inner arm region | Slow forward swimming | Ebersold et al., 1962 |
| pf2-2 (pf2A, CC2503) | DRC, inner arm region | Slow forward swimming | Huang et al., 1982 |
| pf2-4 a (9B11) | DRC, inner arm region | Slow forward swimming | This study |
| pf2-1 rescue | None | Fast forward swimming | This study |
| pf2-4 rescue | None | Fast forward swimming | This study |
| arg2 | None | Fast forward swimming | Harris, 1989 |
| arg7 | None | Fast forward swimming | Harris, 1989 |
| pf2-1 arg2 | DRC, inner arm region | Slow forward swimming | This study |
| pf2-4 arg7 | DRC, inner arm region | Slow forward swimming | This study |
| pf14 | Radial spokes | Paralyzed | Piperno et al., 1981 |
| pf2-4 pf14 | Radial spokes and DRC | Limited motilityb | This study |
9B11 is a new pf2 allele identified in this study. The pf2-3 (pf2B) strain (Huang et al., 1982) is no longer available.
See Results.
Structural defects in the DRC
To determine if the pf2-4 motility phenotype was also associated with structural defects in the DRC, axonemes were isolated from wild-type and pf2-4 cells and prepared for EM. Analysis of longitudinal sections with image-averaging procedures revealed a defect in a crescent-shaped structure located close to the second RS, within the 96-nm axoneme repeat (Fig. 3, A and B). This defect is very similar to the defect previously observed in pf2-1 (Fig. 3 C; Mastronarde et al., 1992; Gardner et al., 1994). The defects can be more clearly seen in the difference plots (Fig. 3, F and G), where the darkened regions represent the statistically significant differences between wild-type and pf2 axonemes. The missing densities correspond to the structure previously identified as the likely location of the DRC polypeptides (Gardner et al., 1994). Significantly, the recovery of wild-type motility after transformation of pf2-4 with the wild-type PF2 gene was also associated with the reassembly of the DRC structure in the rescued strain (Fig. 3 D).
Figure 3.

Structural defects in pf2 axonemes. Longitudinal images of wild-type and mutant axonemes were aligned and computer image averaged. Shown here are grand averages of axonemes isolated from (A) wild type, (B) pf2-4, (C), pf2-1, and (D) a rescued pf2 strain (pf2-4r). The arrows in A–D indicate the region of the 96-nm repeat that is occupied by the DRC structure. (E) Model of axoneme structures within the 96-nm axoneme repeat. The proximal and distal RSs are labeled S1 and S2, respectively; the outer dynein arms (OA) are shown on top, and the DRC is indicated as a crescent-shaped structure above the distal RS (S2). Difference plots between wild type (wt) and pf2-4 (F), and wt and pf2-1 (G) demonstrate that the DRC is missing in both mutants. Note that the DRC is restored in the rescued strain, pf2-4r (D).
Localization of the PF2 polypeptide and molecular complex formation
Previous studies have shown that pf2 axonemes lack five axonemal polypeptides known as DRC subunits 3–7 (Table I; Huang et al., 1982; Piperno et al., 1992, 1994). Dikaryon rescue experiments tentatively identified DRC subunit 4 as the gene product of the PF2 locus (Piperno et al., 1994). To determine if PF2 encodes a structural component of the axoneme or a polypeptide extrinsic to the axoneme that is required for assembly of the DRC, an epitope-tagged construct was used to rescue the pf2 mutant phenotype. PF2 is predicted to contain numerous coiled-coil domains throughout its length (Fig. 4 A). The epitope tag was therefore inserted into the 12th exon of the PF2 gene to avoid possible disruption of the coiled-coil domains and their potential functions (Fig. 2 C; Fig. 4 A). The resulting construct (p9-3HA) is predicted to encode a 60-kD polypeptide with a triple HA epitope tag inserted between aa 458 and 459 of the PF2 sequence. Cotransformation with the epitope-tagged transgene rescued the aberrant motility phenotype of pf2-4 cells with high efficiency, and the rescued strains swam forward at wild-type velocities (e.g., 127 ± 14 μm/s), demonstrating that the presence of the epitope tag does not interfere with the function of PF2 (Fig. 1 C). Axoneme samples from wild-type, pf2-4, and rescued pf2-4 strains were analyzed on Western blots probed with an antibody directed against the HA epitope. A single polypeptide migrating at ∼60 kD was observed exclusively in the epitope-tagged, rescued strain (Fig. 4 B), which indicates that PF2 is a structural component of the isolated axoneme. The size of the epitope-tagged PF2 is consistent with the hypothesis that PF2 encodes DRC subunit 4 (Piperno et al., 1994).
Figure 4.


PF2 protein structure and epitope tagging. (A) Diagrammatic representation of the domain structure of the PF2 polypeptide. Indicated are predicted coiled-coil domains (gray, shaded boxes) and the location of the epitope tag. (B) Western blot analysis of whole axonemes isolated from wild type, pf2-4, and a pf2-4 strain rescued with the epitope-tagged PF2 gene (pf2-4r:3HA). Blots were visualized with a reversible total protein stain (left) before immunolabeling with an antibody directed against the HA epitope (right). The HA antibody recognized a single band in pf2-4r:3HA migrating at ∼60 kD.
Mutant and rescued cells were stained with antibodies against the HA epitope to analyze the subcellular distribution of PF2. Analysis of pf2 mutant and pf2 rescued cells by differential interference contrast microscopy demonstrated that both strains assemble full-length flagella (Fig. 5, A, C, and E). Staining with the HA antibody revealed that the tagged PF2 is present along the entire length of the flagella in the pf2-4 rescued strain (Fig. 5, B and D) but missing in control samples (Fig. 5, F and H). The tagged PF2 was also visible in two spots at the anterior end of the cell in the rescued strain (Fig. 5, B and D). This site corresponds to the predicted location of the two basal bodies. The faint cell body staining seen in pf2 and wild-type cells (Fig. 5, F and H) appeared to result from autofluorescence, as a similar pattern was observed after treatment with secondary antibody alone.
Figure 5.

Localization of the epitope-tagged PF2 protein in Chlamydomonas flagella. The epitope-tagged, rescued strain, pf2-4r:3HA, was stained with an antibody against the HA tag and then imaged by differential interference contrast (A and C) or indirect immunofluorescence light microscopy (B and D). The HA-tagged PF2 is present along the entire length of the flagella in pf2-4r:3HA cells and can also been seen in two spots in the basal body region (B and D). pf2-4 mutant (E and F) or wild-type cells (G and H) labeled with the HA antibody or secondary antibody alone (not depicted) show only background cell body autofluorescence.
Based on the high incidence of coiled-coil domains in the predicted tertiary structure of PF2, it is likely that PF2 interacts with other axonemal polypeptides, specifically other components of the DRC and outer doublet microtubules. To identify those polypeptides, isolated axonemes from the HA-tagged strain were treated with varying concentrations of the zero-length cross-linker 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride (EDC) and then analyzed on Western blots. EDC has previously proven useful in the identification of interactions between subunits of several other axonemal structures (for examples see King et al., 1991; Yang et al., 2000). After a 1-h exposure of isolated axonemes to 1 mM EDC, three new bands migrating at ∼125, 130, and 145 kD, as well as a number of higher molecular mass (>250 kD) bands, could be detected (Fig. 6). The PF2 cross-linked products did not react on Western blots with antibodies to α- and β-tubulin (Fig. 6). Rather, the sizes of the PF2 cross-linked products suggest instead a close association of PF2 with three polypeptides of ∼65, 70, and 85 kD. These results are consistent with the proposal that PF2 is part of a discrete complex of axonemal polypeptides.
Figure 6.

Identification of polypeptides that interact with PF2. Axonemes from the HA-tagged pf2-4 rescued strain were treated with 0, 1, 5, or 10 mM EDC and then used to prepare four identical Western blots. The first blot was stained for total protein. The remaining blots were labeled with antibodies directed against either the HA epitope (HA), α-tubulin (YOL 1/34), or β-tubulin (Tu27B). Asterisks mark three cross-linked products formed as a result of treating the HA-tagged PF2 axonemes with 1 mM EDC.
PF2 encodes the Chlamydomonas homologue of Gas11/Gas8
The Chlamydomonas PF2 gene is predicted to encode a 471-aa polypeptide (Fig. 7 A) with a molecular mass of ∼55 kD and pI of 6.66. Database searches reveal that PF2 is closely related to the growth arrest–specific gene product known as Gas11 in humans and Gas8 in mice (Whitmore et al., 1998; Yeh et al., 2002). PF2 shares >53% identity and 73% similarity with Gas11 and Gas8 (Fig. 7 A). Significant homology is also observed between PF2 and Gas11/Gas8 orthologues found in zebrafish (Danio rerio), flies (Drosophila melanogaster), and trypanosomes (Trypanosoma brucei) (Fig. 7 A). PF2 homologues have also been identified in ESTs obtained from cow, pig, and schistosomes. Phylogenetic analysis indicates that PF2 is more closely related to the vertebrate Gas11/Gas8 sequences than to the fly or trypanosome homologues (Fig. 7 B). No PF2 homologues have been identified in either yeast or Caenorhabditis elegans. However, high levels of Gas11/Gas8 transcripts have been detected in numerous vertebrate cells and tissues that do not assemble motile axonemes (Whitmore et al., 1998; Yeh et al., 2002; unpublished data). PF2 homologues may therefore have additional functions in differentiated cells of higher organisms (see Discussion).
Figure 7.

PF2 homologues. (A) Clustal W alignment of the deduced amino acid sequence of the Chlamydomonas reinhardtii (Cr) PF2 gene (AY309087–AY309089) with homologues identified by BLAST. The shaded areas represent identical or conservatively substituted amino acids in >50% of the sequences. The underlined region represents a putative microtubule-binding domain identified in the trypanosome and human orthologues (Hill et al., 2000). The abbreviation and GenBank/EMBL/DDBJ accession nos. for the sequences are as follows: Danio rerio (Dr), AI584969; Drosophila melanogaster (Dm), AE003427 and AA440118; Homo sapiens (Hs), AF050079; Mus musculus (Mm), U19859; Trypanosoma bruzi (Tb), AAB81499; and Trypanosoma cruzi (Tc), AQ44061. Gas11, growth arrest–specific 11; Gas8, growth arrest–specific 8. (B) Phylogenetic analysis of the Cr PF2 sequence and its homologues.
Discussion
The PF2 locus encodes an axonemal component required for assembly of the DRC
The CP, RS, and DRC are key elements in a signal transduction pathway that regulates dynein arm activity to produce the complex waveforms characteristic of eukaryotic cilia and flagella (for reviews see Porter and Sale, 2000; Kamiya, 2002). To better understand the role of the individual DRC subunits in this pathway, we used insertional mutagenesis strategies to isolate tagged mutations and thereby cloned the PF2 locus. This is the first DRC gene and gene product to be characterized at both the molecular and cellular levels.
pf2 mutants fail to assemble five of the seven DRC-associated subunits (Table I; Piperno et al., 1994) and display both reduced swimming velocities and aberrant waveforms with reduced shear amplitudes (Brokaw and Kamiya, 1987; Gardner et al., 1994). Our work demonstrates that PF2 encodes an ∼55-kD polypeptide that is uniformly distributed along the length of the axoneme and in the basal body region (Figs. 4 and 5). These results are consistent with the localization of the DRC (Gardner et al., 1994) and a prediction based on dikaryon rescue experiments that PF2 might encode DRC subunit 4 (Piperno et al., 1994). In addition, transformation of pf2 mutants with the wild-type PF2 gene restores both wild-type flagellar motility and the missing DRC structure (Figs. 1 and 3). Thus, PF2 is required for proper assembly of DRC components and the regulation of flagellar motility.
The predicted structure of the PF2 polypeptide contains several coiled-coil domains (Fig. 4) that could play a role in protein–protein interactions. The most likely candidates for such interactions would be other components of the DRC and/or tubulin subunits of the outer doublets. Preliminary experiments using the zero-length cross-linker EDC suggest that PF2 may not directly interact with α- or β-tubulin (Fig. 6), but whether an interaction might be identified using other cross-linking agents remains to be determined. However, a close association between PF2 and at least three polypeptides has been identified, based on the relative mobility of three cross-linked products that react with an antibody specific for the HA-tagged PF2 (Fig. 6). The identities of the three polypeptides are unknown, but their apparent sizes are similar to those reported for three DRC subunits, DRC1, DRC2, and DRC3 (Table I; Piperno et al., 1994). Further confirmation will require the development of specific probes for these other DRC subunits. Nonetheless, these results, together with previous observations that pf2 axonemes lack five different polypeptides (DRC3–DRC7) (Huang et al., 1982; Piperno et al., 1992, 1994), indicate that the DRC is a discrete axonemal structure.
The function of PF2-related proteins in other species
Close homologues of PF2 have been found in a variety of organisms (Fig. 7) and include the growth arrest–specific gene product known as Gas8 (mice) or Gas11 (humans) (Brenner et al., 1989; Whitmore et al., 1998; Yeh et al., 2002), the trypanosome protein known as the T lymphocyte triggering factor (TLTF) or trypanin (Vaidya et al., 1997; Hill et al., 2000; Hutchings et al., 2002), and several Gas8/Gas11-related sequences in insects, fish, protozoa, and other mammals. The remarkably high degree of sequence conservation (Fig. 7 A) suggests a highly conserved cellular function.
Gas8 was first described as a gene whose expression in mouse fibroblasts was activated during growth arrest induced by serum starvation (Brenner et al., 1989; Lih et al., 1996). Recent work has shown that both Gas8 mRNA and Gas8 protein are highly enriched in adult mouse testes (Yeh et al., 2002). Gas8 has been localized along the length of sperm flagella, but it is also found in the apical region of ciliated epithelia that line the respiratory and female reproductive tracts (Yeh et al., 2002). The pattern of localization for Gas8 is similar to that observed for PF2 (Fig. 5) and indicates that both polypeptides are conserved components of motile axonemes.
The human homologue, Gas11, was tentatively identified as a candidate tumor suppressor gene based on its location at a chromosome locus (16q24.3) commonly deleted in breast and prostate carcinomas, and the expression of its mouse orthologue during growth arrest (Whitmore et al., 1998). Gas11 has since been localized in developing sperm, but its distribution in other human tissues has not been analyzed (Yeh et al., 2002). Interestingly, however, overexpression of a highly conserved region of Gas11 (aa 115–228; Fig. 7) resulted in the colocalization of a Gas11–GFP fusion protein with the plus ends of cytoplasmic microtubules in both trypanosomes and cultured mammalian cells (Hill et al., 1999, 2000). These observations prompted the suggestion that the conserved region might serve as a novel microtubule-binding domain and that Gas11 may function as a microtubule-specific cytoskeletal linker protein (Hill et al., 1999, 2000). A similar domain is present in PF2 (aa 114–227), but its function is unknown (Fig. 7).
Gas8 and Gas11 transcripts have previously been detected in a variety of tissues that do not assemble motile axonemes (Whitmore et al., 1998; Yeh et al., 2002). We repeated these experiments using a mouse Gas8 probe and found high levels of Gas8 transcripts not only in mouse testis, lung, and brain, but also in heart, liver, and kidney (unpublished data). What might be the origin of PF2-related transcripts in these tissues? One possibility might be that all of these tissues contain cells that assemble immotile, primary cilia (see www.wadsworth.org/bms/SCBlinks/cilial1.html). Although primary cilia typically lack the CP, RS, and dynein arms, it is possible that some DRC subunits may have been retained within the axoneme structure. If so, it would be reasonable to identify PF2-related transcripts in those tissues that assemble significant levels of primary cilia. Further work is needed to determine if Gas8/Gas11 is found in primary cilia or centrioles in mammalian cells. If present, this may account for the activation of Gas8 expression observed in growth-arrested fibroblasts (Brenner et al., 1989; Yeh et al., 2002). Previous studies have shown that maximal assembly of primary cilia in these cells is closely correlated with withdrawal from the cell cycle and/or growth to confluence (Tucker et al., 1979; Wheatley et al., 1996). Recent work has also revealed that primary cilia can serve as sensory organelles, relaying signals from receptors in the cilium membrane and thereby allowing cells to monitor their local environment and respond appropriately (Nauli et al., 2003). An exciting possibility is that signals received by primary cilia may influence the growth arrest process (Wheatley et al., 1996; Pazour and Witman, 2003).
An alternative hypothesis would be that the mammalian homologues Gas8 and Gas11 have other, yet unidentified, cytoplasmic functions distinct from their role in the axoneme. The observation that a conserved domain can direct a Gas11 fusion protein to colocalize with the plus ends of microtubules in tissue culture cells (Hill et al., 2000) may support this hypothesis. Additional work is clearly needed to more firmly establish the subcellular distribution of Gas8/Gas11 in mammalian cells.
PF2 is also closely related to a trypanosome protein, trypanin (formerly called TLTF). TLTF was first identified as a polypeptide released by T. brucei that triggers CD8+ T lymphocytes to proliferate and secrete γ-interferon (Vaidya et al., 1997). Initial studies of a GFP-tagged TLTF fusion protein suggested that TLTF was localized to the flagellar pocket, a region formed by an invagination of the cell membrane at the site where the flagellum emerges from the cell body (Hill et al., 1999). More recent work has shown that TLTF/trypanin is a component of the trypanosome cytoskeleton (Hill et al., 2000), where it is distributed along the length of the flagellum (similar to both Gas8 and PF2) and may be associated with a region known as the flagellar attachment site (Hutchings et al., 2002).
Interestingly, knockout of trypanin expression by RNA interference has shown that trypanin is required for directional cell motility of the trypanosome (Hutchings et al., 2002). Trypanin(−) mutants do not have paralyzed flagella, but they have lost the ability to coordinate their flagellar beat and thus cannot generate productive cell motility. Analysis by transmission EM failed to identify a clear defect in axoneme structure. However, scanning EM indicated that the flagellum was partially detached from the cell in ∼28% of the trypanin(−) strains as compared with ∼8% in control strains. Hutchings et al. (2002) therefore proposed that trypanin is a novel cytoskeletal linker protein critical for linking the flagellum to the underlying cytoskeleton.
Based on the similarities between PF2 and trypanin, we favor an alternative interpretation of the trypanin(−) phenotype. First, trypanin is likely to be a component of the DRC in the trypanosome axoneme. The EM methods used by Hutchings et al. (2002) to analyze trypanin(−) axonemes would probably not be sensitive enough to resolve subtle defects in DRC structure. The identification of structural defects in pf2 axonemes required the use of image-averaging procedures (Mastronarde et al., 1992; Gardner et al., 1994). Second, the apparent defect in flagellar attachment to the cell body could be a consequence of the aberrant flagellar motility in the trypanin(−) cells. This interpretation is consistent with the swimming behavior of the trypanin(−) strains (Hutchings et al., 2002), the motility phenotypes of pf2 mutants in Chlamydomonas (Brokaw and Kamiya, 1987; Mastronarde et al., 1992; Gardner et al., 1994), and the present study.
Role of the DRC and its constituent polypeptides
The absence of a functional PF2 protein results in the inability of pf2 strains to assemble a major portion of the DRC (Gardner et al., 1994; Piperno et al., 1994). The mutant cells also display an unusual motility phenotype (Brokaw and Kamiya, 1987). When combined with a CP- or RS-associated paralyzed flagellar mutation, pf2 mutations restore partial flagellar motility without restoring the missing CP or RS structures (Huang et al., 1982; Piperno et al., 1992). The suppression of flagellar paralysis has led to the hypothesis that the DRC is a part of a signal transduction pathway that controls motility by regulating dynein activity (for review see Porter and Sale, 2000). The DRC apparently serves this function by acting as an intermediate in the pathway, receiving signals from the CP/RS structures and then distributing them appropriately to the dynein arms.
We propose that PF2 encodes a DRC polypeptide that is involved in stabilizing the complex by interacting with other DRC subunits. This proposal is supported by the observations that pf2 axonemes lack four additional DRC subunits (Piperno et al., 1994), PF2 can be cross-linked to at least three axonemal polypeptides (Fig. 6), and coiled-coil domains are present within the PF2 structure (Fig. 4). We suggest that PF2 serves as a molecular scaffold, whose function is to appropriately localize other components within the axoneme, in a manner similar to that suggested for the CP-associated protein PF6 (Rupp et al., 2001) and the RS-associated protein RSP3 (Gaillard et al., 2001). Other DRC subunits could therefore be positioned to interact with the network of regulatory enzymes and thus regulate motility by controlling the phosphorylation state of key axonemal polypeptides. This model is also consistent with observations indicating that the DRC plays an important role in stabilizing the assembly of specific inner arm isoforms (Gardner et al., 1994; Piperno et al., 1994). Alternatively, PF2 might also serve as a mechanical sensor or molecular spring that senses tension during axonemal bending and thereby influences the activity of nearby dynein isoforms. Future studies to identify and characterize other subunits of the DRC that interact with PF2 will be needed to test these various models.
Materials and methods
Mutant strains and genetic analyses
Strains listed in Table II were maintained as previously described (Rupp et al., 2001). The motility mutant 9B11 was generated by transformation of a nit1-305 strain with the plasmid pMN24 containing a wild-type copy of the NIT1 gene (Fernandez et al., 1989). The NIT1 plasmid was inserted into the PF2 locus (Fig. 1), and so 9B11 is now known as pf2-4. Other strains were obtained from the Chlamydomonas Genetics Center.
To determine whether the motility phenotype of 9B11 was linked to the NIT1 plasmid used as a selectable marker, 9B11 was backcrossed to a nit − strain, and the resulting progeny were analyzed for cosegregation of their motility phenotypes and their ability to grow on selective media. Out of 56 progeny obtained from seven complete octads, 28 were nit + and swam slowly, whereas all 28 nit − strains had wild-type motility.
Isolation of genomic sequence flanking the NIT1 plasmid
Purification of genomic DNA, restriction enzyme digests, agarose gels, isolation of total RNA, preparation of cDNA, PCR reactions, and Southern and Northern blots were performed as previously described (Rupp et al., 2001). To identify genomic DNA flanking the site of the plasmid insertion, wild-type and 9B11 genomic DNA were analyzed on Southern blots, and a 6.2-kb BamHI fragment unique to 9B11 was identified by hybridization with a 3.2-kb probe derived from the 3′ end of NIT1. The 6.2-kb BamHI fragment was recovered by screening a size-fractionated minilibrary with the NIT1 probe as previously described (Rupp et al., 2001). Positive clones were digested with several restriction enzymes and probed with NIT1 to identify a unique 0.45-kb NotI/BamHI fragment containing only 9B11 genomic DNA. Southern blot analysis confirmed that this fragment, designated FC-1, was located close to the site of NIT1 insertion (Fig. 1).
Recovery of the wild-type PF2 gene
A large-insert, wild-type (21gr), genomic library constructed in λFIX II (Schnell and Lefebvre, 1993) was screened with FC-1 as previously described (Rupp et al., 2001). 13 phage clones were recovered and restriction mapped. Selected subclones were sequenced and analyzed for potential open reading frames as previously described (Rupp et al., 2001). 9.3 kb of genomic sequence has been deposited under GenBank/EMBL/DDBJ accession nos. AY309087–AY309089. An incomplete version of the sequence is also present in the recently released draft of the Chlamydomonas genome (http://genome.jgi-psf.org/chlre1/chlre1.home.html).
To place the sequence on the genetic map, a genomic fragment was used to identify a PvuII restriction fragment length polymorphism between two strains, 137c and S1-D2. The fragment was then hybridized to a series of mapping filters containing DNA isolated from tetrad progeny of crosses between multiply marked C. reinhardtii strains and S1-D2. The segregation of the PvuII polymorphism was analyzed relative to the segregation of >42 genetic and molecular markers (Porter et al., 1996). The sequence is linked to the molecular marker CRY1 (∼9.4 cM) and the motility mutation pf2-1 (<5.5 cM).
To test whether any of the clones could rescue the motility defects in pf2-4 and other pf2 mutants, pf2 arg strains were cotransformed with pARG7.8 (containing a wild-type copy of the ARG7 gene; Debuchy et al., 1989) and the clone in question. Transformants were selected on media lacking arginine and then screened for recovery of wild-type motility (Rupp et al., 2001).
Construction of an epitope-tagged gene construct
To determine whether PF2 was a structural component of the axoneme, a modified PF2 gene containing three consecutive HA epitopes was constructed. A 6.9-kb XmaI fragment containing the complete PF2 transcription unit was subcloned from the rescuing phage clone, λG2, into pUC119 to form the plasmid p9B11-X1.2. A sequence encoding a triple HA epitope (p3HA) was PCR amplified from a second plasmid provided by M. Lavoie and C. Silflow (University of Minnesota, St. Paul, MN) using primers with BstEII restriction sites. A 250-bp product containing the triple HA epitope was ligated into pGEM-T Easy (Promega), released with BstEII, and then religated into a unique BstEII site in p9B11-X1.2, yielding the epitope-tagged PF2 gene, p9-3HA. Sequence analysis confirmed that the epitope tag was inserted into the 12th exon in the proper orientation and reading frame.
Fractionation of flagella and biochemical analyses
Axonemes were isolated from vegetative cells (Porter et al., 1992; Rupp et al., 2001) and analyzed on either 7.5% polyacrylamide or 5–15% acrylamide, 0–2.4 M glycerol gradient gels using the Laemmli (1970) buffer system. Gels were stained directly or transferred to Immobilon-P (Millipore). Total protein on the blots was visualized with Blot FastStain (Chemicon International). Western blots were probed with a high affinity rat antibody directed against the HA epitope (clone 3F10; Roche Molecular Biochemicals). Immunoreactive bands were detected using an alkaline phosphatase–conjugated secondary antibody (Rupp et al., 2001).
To identify polypeptides that associate with PF2, axonemes from the HA-tagged, pf2-4 rescued strain were prepared in buffer lacking DTT. Chemical cross-linking was performed by treating axoneme samples with EDC (Pierce Chemical Co.) for 1 h at room temperature (King et al., 1991). The reaction was terminated by the addition of a 10-fold molar excess of β-mercaptoethanol. Western blots were probed with antisera directed against either the HA epitope, α-tubulin (YOL 1/34; Accurate Chemical Co.), or β-tubulin (Tu27B; a gift of L.I. Binder, Northwestern University, Evanston, IL), and the appropriate secondary antibody.
Electron and light microscopy
Axonemes were prepared for EM as described by Porter et al. (1992). The methods for digitization and image averaging were as previously described (Mastronarde et al., 1992; O'Toole et al., 1995). Analysis of axoneme cross sections indicated that the structural defects in pf2-4 were similar to those seen previously with pf2-1, but potential differences between proximal and distal regions were not examined. Control and HA-tagged strains were prepared for immunofluorescence using an ice-cold methanol fixation protocol described by Sanders and Salisbury (1995). Fixed cells were stained with a primary antibody directed against the HA epitope and an Alexa®488 secondary antibody as previously described (Rupp et al., 2001; Perrone et al., 2003).
Acknowledgments
We thank members of the Porter laboratory for their support, especially Cathy Perrone. We are also grateful to members of the Lefebvre, Linck, and Silflow laboratories for helpful advice. We extend special thanks to Dave Mitchell (State University of New York Health Sciences Center, Syracuse, NY) for providing the 9B11 strain, Eileen O'Toole for expert assistance with image analysis, Matthew Lavoie for supplying the plasmid encoding the triple HA epitope, Andrew Bostrom for analysis of the pf2 alleles, and Darryl Kreugger for assistance with EM.
This work was supported by grants from the National Institute of General Medical Sciences (GM-55667) and Minnesota Medical Foundation to M.E. Porter. G. Rupp was supported in part by a National Institutes of Health (NIH) postdoctoral fellowship (F32-GM17901), a training grant from the National Science Foundation for Interdisciplinary Studies on the Cytoskeleton (DIR-9113444), and start-up funds from Southern Illinois University School of Medicine. We also acknowledge the assistance of the Boulder Laboratory for 3-Dimensional Fine Structure, which is supported by NIH grant RR00592.
Footnotes
Abbreviations used in this paper: CP, central pair; DRC, dynein regulatory complex; EDC, 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride; RS, radial spoke; TLTF, T lymphocyte triggering factor.
References
- Afzelius, B.A. 1995. Role of cilia in human health. Cell Motil. Cytoskeleton. 32:95–97. [DOI] [PubMed] [Google Scholar]
- Brenner, D.G., S. Lin-Chao, and S.N. Cohen. 1989. Analysis of mammalian cell genetic regulation in situ by using retrovirus-derived “portable exons” carrying the Escherichia coli lacZ gene. Proc. Natl. Acad. Sci. USA. 86:5517–5521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brokaw, C.J., and R. Kamiya. 1987. Bending patterns of Chlamydomonas flagella: IV. Mutants with defects in inner and outer dynein arms indicate differences in dynein arm function. Cell Motil. Cytoskeleton. 8:68–75. [DOI] [PubMed] [Google Scholar]
- Burgess, S.A., D.A. Carter, S.D. Dover, and D.M. Woolley. 1991. The inner dynein arm complex: compatible images from freeze-etch and thin section methods of microscopy. J. Cell Sci. 100:319–328. [DOI] [PubMed] [Google Scholar]
- Debuchy, R., S. Purton, and J.-D. Rochaix. 1989. The arginosuccinate lyase gene of Chlamydomonas reinhardtii: an important tool for nuclear transformation and for correlating the genetic and molecular maps of the ARG7 locus. EMBO J. 8:2803–2809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebersold, W.T., R.P. Levine, E.E. Levine, and M.A. Olmsted. 1962. Linkage maps in Chlamydomonas reinhardtii. Genetics. 47:531–543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez, E., R. Schnell, L.P.W. Ranum, S.C. Hussey, C.D. Silflow, and P.A. Lefebvre. 1989. Isolation and characterization of the nitrate reductase structural gene in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA. 86:6449–6453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox, L.A., and W.S. Sale. 1987. Direction of force generated by the inner row of dynein arms on flagellar microtubules. J. Cell Biol. 105:1781–1787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaillard, A.R., D.R. Diener, J.L. Rosenbaum, and W.S. Sale. 2001. Flagellar radial spoke protein 3 is an A-kinase anchoring protein (AKAP). J. Cell Biol. 153:443–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardner, L.C., E. O'Toole, C.A. Perrone, T. Giddings, and M.E. Porter. 1994. Components of a “dynein regulatory complex” are located at the junction between the radial spokes and the dynein arms in Chlamydomonas flagella. J. Cell Biol. 127:1311–1325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Habermacher, G., and W. Sale. 1997. Regulation of flagellar dynein by phosphorylation of a 138-kD inner arm dynein intermediate chain. J. Cell Biol. 136:167–176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamasaki, T., K. Barkalow, J. Richmond, and P. Satir. 1991. cAMP-stimulated phosphorylation of an axonemal polypeptide that copurifies with the 22S dynein arm regulates microtubule translocation velocity and swimming speed in Paramecium. Proc. Natl. Acad. Sci. USA. 88:7918–7922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris, E. 1989. The Chlamydomonas Sourcebook. Academic Press, San Diego, CA. 780 pp. [DOI] [PubMed]
- Hill, K.L., N.R. Hutchings, D.G. Russell, and J.E. Donelson. 1999. A novel protein targeting domain directs proteins to the anterior cytoplasmic face of the flagellar pocket in African trypanosomes. J. Cell Sci. 112:3091–3101. [DOI] [PubMed] [Google Scholar]
- Hill, K.L., N.R. Hutchings, P.M. Grandgenett, and J.E. Donelson. 2000. T lymphocyte-triggering factor of African trypanosomes is associated with the flagellar fraction of the cytoskeleton and represents a new family of proteins that are present in several divergent eukaryotes. J. Biol. Chem. 275:39369–39378. [DOI] [PubMed] [Google Scholar]
- Huang, B., Z. Ramanis, and D.J. Luck. 1982. Suppressor mutations in Chlamydomonas reveal a regulatory mechanism for flagellar function. Cell. 28:115–125. [DOI] [PubMed] [Google Scholar]
- Hutchings, N.R., J.E. Donelson, and K.L. Hill. 2002. Trypanin is a cytoskeletal linker protein and is required for cell motility in African trypanosomes. J. Cell Biol. 156:867–877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamiya, R. 2002. Functional diversity of axonemal dyneins as studied in Chlamydomonas mutants. Int. Rev. Cytol. 219:115–155. [DOI] [PubMed] [Google Scholar]
- King, S.J., and S.K. Dutcher. 1997. Phosphoregulation of an inner dynein arm complex in Chlamydomonas reinhardtii is altered in phototactic mutant strains. J. Cell Biol. 136:177–191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- King, S.M., C.G. Wilkerson, and G.B. Witman. 1991. The Mr 78,000 intermediate chain of Chlamydomonas outer arm dynein interacts with α-tubulin in situ. J. Biol. Chem. 266:8401–8407. [PubMed] [Google Scholar]
- Laemmli, U.K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 227:680–685. [DOI] [PubMed] [Google Scholar]
- Lefebvre, P.A., and J.L. Rosenbaum. 1986. Regulation of the synthesis and assembly of ciliary and flagella proteins during regeneration. Annu. Rev. Cell Biol. 2:517–546. [DOI] [PubMed] [Google Scholar]
- Lih, C.-J., S.N. Cohen, C. Wang, and S. Lin-Chao. 1996. The platelet-derived growth factor α-receptor is encoded by a growth-arrest-specific (gas) gene. Proc. Natl. Acad. Sci. USA. 93:4617–4622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mastronarde, D.N., E.T. O'Toole, K.L. McDonald, J.R. McIntosh, and M.E. Porter. 1992. Arrangement of inner dynein arms in wild-type and mutant flagella of Chlamydomonas. J. Cell Biol. 118:1145–1162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nauli, S.M., F.J. Alenghat, Y. Luo, E. Williams, P. Vassilev, X. Li, A.E. Elia, W. Lu, E.M. Brown, S.J. Quinn, et al. 2003. Polycystins 1 and 2 mediate mechanosensation in the primary cilium of kidney cells. Nat. Genet. 33:129–137. [DOI] [PubMed] [Google Scholar]
- Nelson, J.A.E., P.B. Savereide, and P.A. Lefebvre. 1994. The Cry1 gene in Chlamydomonas reinhardtii: structure and use as a dominant selectable marker for nuclear transformation. Mol. Cell. Biol. 14:4011–4019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nonaka, S., Y. Tanaka, Y. Okada, S. Takeda, A. Harada, Y. Kanai, M. Kido, and N. Hirokawa. 1998. Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell. 95:829–837. [DOI] [PubMed] [Google Scholar]
- O'Toole, E., D. Mastronarde, J.R. McIntosh, and M.E. Porter. 1995. Computer-assisted analysis of flagellar structure. Methods Cell Biol. 47:183–191. [DOI] [PubMed] [Google Scholar]
- Omoto, C.K., and C. Kung. 1979. The pair of central tubules rotates during ciliary beat in Paramecium. Nature. 279:532–534. [DOI] [PubMed] [Google Scholar]
- Omoto, C.K., I.R. Gibbons, R. Kamiya, C. Shingyoji, K. Takahashi, and G.B. Witman. 1999. Rotation of the central pair microtubules in eukaryotic flagella. Mol. Biol. Cell. 10:1–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pazour, G.J., and J.L. Rosenbaum. 2002. Intraflagellar transport and cilia-dependent diseases. Trends Cell Biol. 12:551–555. [DOI] [PubMed] [Google Scholar]
- Pazour, G.J., and G.B. Witman. 2003. The vertebrate primary cilium is a sensory organelle. Curr. Opin. Cell Biol. 15:105–110. [DOI] [PubMed] [Google Scholar]
- Perrone, C.A., D. Tritschler, P. Taulman, R. Bower, B.K. Yoder, and M.E. Porter. 2003. A novel dynein light intermediate chain co-localizes with the retrograde motor for intraflagellar transport at sites of axoneme assembly in Chlamydomonas and mammalian cells. Mol. Biol. Cell. 14:2041–2056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piperno, G., B. Huang, Z. Ramanis, and D.J.L. Luck. 1981. Radial spokes of Chlamydomonas flagella: polypeptide composition and phosphorylation of stalk components. J. Cell Biol. 88:73–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piperno, G., K. Mead, and W. Shestak. 1992. The inner dynein arms I2 interact with a “dynein regulatory complex” in Chlamydomonas flagella. J. Cell Biol. 118:1455–1463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piperno, G., K. Mead, M. LeDizet, and A. Moscatelli. 1994. Mutations in the “dynein regulatory complex” alter the ATP-insensitive binding sites for inner arm dyneins in Chlamydomonas axonemes. J. Cell Biol. 125:1109–1117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porter, M.E., J. Power, and S.K. Dutcher. 1992. Extragenic suppressors of paralyzed flagellar mutations in Chlamydomonas reinhardtii identify loci that alter the inner dynein arms. J. Cell Biol. 118:1163–1176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porter, M.E., J.A. Knott, L.C. Gardner, D.K. Mitchell, and S.K. Dutcher. 1994. Mutations in the SUP-PF-1 locus of Chlamydomonas reinhardtii identify a regulatory domain in the β-dynein heavy chain. J. Cell Biol. 126:1495–1507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porter, M.E., J.A. Knott, S.H. Myster, and S.J. Farlow. 1996. The dynein gene family in Chlamydomonas reinhardtii. Genetics. 144:569–585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porter, M.P., and W.S. Sale. 2000. The 9 + 2 axoneme anchors multiple inner arm dyneins and a network of kinases and phosphatases that control motility. J. Cell Biol. 151:F37–F42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rupp, G., E. O'Toole, L.C. Gardner, B.F. Mitchell, and M.P. Porter. 1996. The sup-pf-2 mutations of Chlamydomonas alter the activity of the outer dynein arm by modification of the γ-dynein heavy chain. J. Cell Biol. 135:1853–1865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rupp, G., E. O'Toole, and M.P. Porter. 2001. The Chlamydomonas PF6 locus encodes a large alanine/proline-rich polypeptide that is required for assembly of a central pair projection and regulated flagellar motility. Mol. Biol. Cell. 12:739–751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sale, W.S., and P. Satir. 1977. The direction of active sliding of microtubules in Tetrahymena cilia. Proc. Natl. Acad. Sci. USA. 74:2045–2049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- San Agustin, J.T., and G.B. Witman. 1994. Role of cAMP in the reactivation of demembranated ram spermatozoa. Cell Motil. Cytoskeleton. 27:206–218. [DOI] [PubMed] [Google Scholar]
- Sanders, M.A., and J.L. Salisbury. 1995. Immunofluorescence microscopy of cilia and flagella. Methods Cell Biol. 47:163–169. [DOI] [PubMed] [Google Scholar]
- Sapiro, R., I. Kostetskii, P. Olds-Clarke, G.L. Gerton, G.L. Radice, and J.F. Strauss III. 2002. Male infertility, impaired sperm motility, and hydrocephalus in mice deficient in sperm-associated antigen 6. Mol. Cell Biol. 22:6298–6305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schnell, R.A., and P.A. Lefebvre. 1993. Isolation of the Chlamydomonas regulatory gene NIT2 by transposon tagging. Genetics. 134:737–747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith, E.F. 2002. Regulation of flagellar dynein by calcium and a role for an axonemal calmodulin and calmodulin-dependent kinase. Mol. Biol. Cell. 13:3303–3313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith, E.F., and W.S. Sale. 1992. Regulation of dynein-driven microtubule sliding by the radial spokes in flagella. Science. 257:1557–1559. [DOI] [PubMed] [Google Scholar]
- Supp, D.M., S.S. Potter, and M. Bruekner. 2000. Molecular motors: the driving force behind mammalian left-right development. Trends Cell Biol. 10:41–45. [DOI] [PubMed] [Google Scholar]
- Tam, L.-W., and P.A. Lefebvre. 1993. Cloning of flagellar genes in Chlamydomonas reinhardtii by DNA insertional mutagenesis. Genetics. 135:375–384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tucker, R.W., C.D. Scher, and C.D. Stiles. 1979. Centriole deciliation associated with the early response of 3T3 cells to growth factors but not to SV40. Cell. 18:1065–1072. [DOI] [PubMed] [Google Scholar]
- Vaidya, T., M. Bakhiet, K.L. Hill, T. Olsson, K. Kristensson, and J.E. Donelson. 1997. The gene for a T lymphocyte triggering factor from African trypanosomes. J. Exp. Med. 186:433–438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheatley, D.N., A.M. Wang, and G.E. Strugnell. 1996. Expression of primary cilia in mammalian cells. Cell Biol. Int. 20:73–81. [DOI] [PubMed] [Google Scholar]
- Whitmore, S.A., C. Settasatian, J. Crawford, K.M. Lower, B. McCallum, R. Seshadri, C.J. Cornelisse, E.W. Moerland, A.-M. Cleton-Jansen, A.J. Tipping, et al. 1998. Characterization and screening for mutations of the growth arrest-specific 11 (GAS11) and C16orf3 genes at 16q24.3 in breast cancer. Genomics. 53:325–331. [DOI] [PubMed] [Google Scholar]
- Witman, G.B., J. Plummer, and G. Sander. 1978. Chlamydomonas flagellar mutants lacking radial spokes and central tubules. Structure, composition, and function of specific axonemal components. J. Cell Biol. 76:729–747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woolley, D.M. 1997. Studies on the eel sperm flagellum I. The structure of the inner dynein arm complex. J. Cell Sci. 110:85–94. [DOI] [PubMed] [Google Scholar]
- Yang, P., and W.S. Sale. 2000. Casein kinase I is anchored on axonemal doublet microtubules and regulates flagellar dynein phosphorylation and activity. J. Biol. Chem. 275:18905–18912. [DOI] [PubMed] [Google Scholar]
- Yang, P., L. Fox, R.J. Colbran, and W.S. Sale. 2000. Protein phosphatases PP1 and PP2A are located in distinct positions in the Chlamydomonas flagellar axoneme. J. Cell Sci. 113:91–102. [DOI] [PubMed] [Google Scholar]
- Yeh, S.-D., Y.-J. Chen, A.C.Y. Chang, R. Ray, B.-R. She, W.-S. Lee, H.-S. Chiang, S.N. Cohen, and S. Lin-Chao. 2002. Isolation and properties of Gas8, a growth arrest-specific gene regulated during male gametogenesis to produce a protein associated with the sperm motility apparatus. J. Biol. Chem. 277:6311–6317. [DOI] [PubMed] [Google Scholar]