

Abstract
During receptor-mediated endocytosis, AP2 complexes act as a bridge between the cargo membrane proteins and the clathrin coat by binding to sorting signals via the μ2 subunit and to clathrin via the β subunit. Here we show that binding of AP2 to sorting signals in vitro is regulated by phosphorylation of the μ2 subunit of AP2. Phosphorylation of μ2 enhances the binding affinity of AP2 for sorting motifs as much as 25-fold compared with dephosphorylated AP2. The recognition of sorting signals was not affected by the phosphorylation status of the α or β2 subunit, suggesting that phosphorylation of μ2 is critical for regulation of AP2 binding to sorting signals. Phosphorylation of μ2 occurs at a single threonine residue (Thr-156) and is mediated by the newly discovered adaptor-associated kinase, AAK1, which copurifies with AP2. We propose that phosphorylation of the AP2 μ2 subunit by AAK1 ensures high affinity binding of AP2 to sorting signals of cargo membrane proteins during the initial steps of receptor-mediated endocytosis.
Keywords: clathrin; endocytosis; receptor; trafficking; coated vesicle
Introduction
Endocytosis from the plasma membrane via clathrin-coated vesicles (CCVs)* is the major pathway for entry of receptors and certain pathogens into the cell. AP2 is a key component of the endocytic machinery that links cargo membrane proteins to the clathrin lattice. AP2 is a member of the family of adaptor proteins (today comprising AP1, AP2, AP3, and AP4) and is composed of two large subunits with a molecular mass of ∼100 kD, termed α and β2, a subunit of ∼50 kD, termed μ2, and a small subunit of 19 kD, called σ2 (Kirchhausen, 1999). Considerable evidence suggests that μ2 binds to sorting signals present in the cytoplasmic tails of membrane proteins (Aguilar et al., 1997; Owen and Evans, 1998), whereas the so-called hinge region of the AP2 β2 subunit binds to clathrin (Goodman and Keen, 1995; Shih et al., 1995).
Protein kinases have long been known to associate with CCVs (Campbell et al., 1984; Pauloin et al., 1984; Bar-Zvi and Branton, 1986), and phosphorylation plays a role in regulating protein interactions in receptor-mediated endocytosis (Slepnev et al., 1998; Cousin and Robinson, 2001). For example, AP2 must be dephosphorylated to mediate clathrin assembly (Wilde and Brodsky, 1996). We recently demonstrated that phosphorylation of AP2 by an endogenous kinase(s) enhances binding to the internalization motifs of membrane proteins (Fingerhut et al., 2001). However, neither the identity of the kinase(s) involved, nor which of the four AP2 subunits required phosphorylation to mediate high affinity binding to membrane protein sorting signals was known. Here we show that μ2 phosphorylation accounts for the enhanced affinity for sorting motifs, and we identify AAK1, a newly discovered adaptor-associated kinase (Conner and Schmid, 2002) as the responsible kinase.
Results and discussion
AP2 phosphorylation by copurifying kinases
To identify the kinase and its targets in the AP2 complex required for high affinity binding to sorting motifs, we purified AP2 from pig brain according to published procedures (see Materials and methods). The final purification step during which AP2 is separated from AP1 by hydroxyapatite (HAP) chromatography is shown in Fig. 1 A. The AP2-containing fractions were collected in two pools (3 and 4) and were completely resolved from the AP1-containing pool 2, as revealed by Western blotting (Fig. 1 A, inset).
Figure 1.
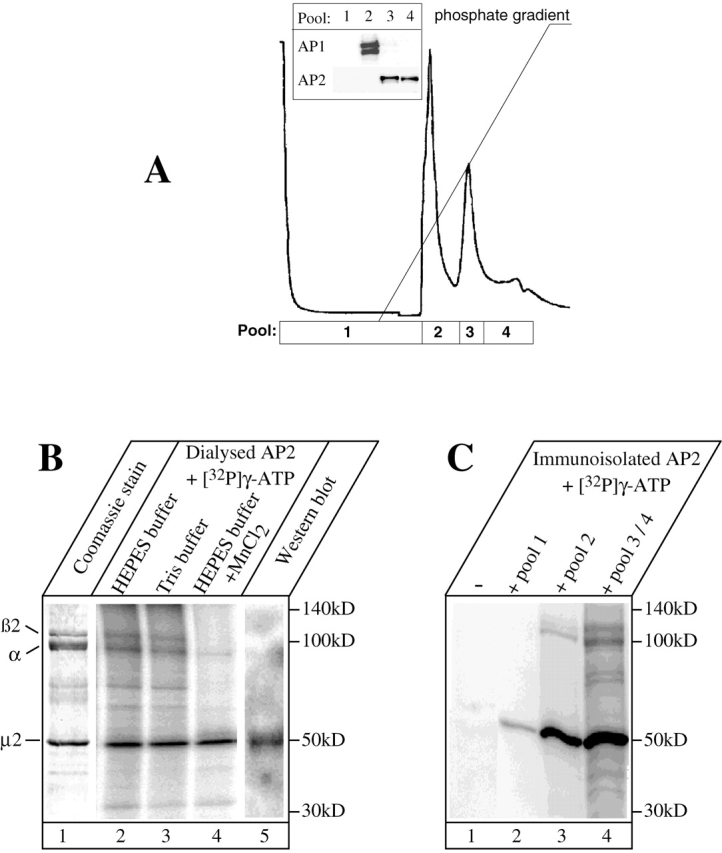
Highly purified AP2 is phosphorylated by copurifying kinases. (A) AP1 and AP2 were prepared from pig brain by differential centrifugation and gel filtration. Separation of AP2 from AP1 was achieved by HAP chromatography with AP1 eluting first. The different pools (pools 1–4) used for subsequent analysis are indicated below the chromatogram. Inset shows a Western blot analysis for AP1 and AP2, ruling out any cross-contamination between the pooled fractions. (B) AP2 from pool 4 (Coomassie stain, lane 1) was incubated in buffer A (lane 2), buffer B (lane 3), or buffer A containing divalent Mn2+ (lane 4) in the presence of [γ32P]ATP before SDS-PAGE and detection of incorporated radioactive phosphate. The 50-kD protein, predominantly phosphorylated by the copurifying kinase activity, was identified as the μ2 subunit by Western blotting (lane 5) and also by radiosequencing (Fig. S1, available online at http://www.jcb.org/cgi/content/full/jcb.200111068/DC1). (C) AP2 was immunoisolated from pool 4 (see Materials and methods), extensively washed, and incubated for 20 min with [γ32P]ATP alone (lane 2) or [γ32P]ATP together with one of the indicated HAP pools (lanes 2–5). Subsequently, the samples were washed and the incorporated phosphate was detected.
The HAP-purified AP2 fractions (Fig. 1 B, lane 1) still contained kinase activity, as incubation at 37°C in the presence of [γ32P]ATP led to the phosphorylation of a 50-kD protein and, to a lesser extent, two proteins of ∼100 kD (Fig. 1 B, lane 2). The latter two proteins most likely represent the AP2 α and β2 subunits, and the 50-kD band was identified as the μ2 subunit by Western blot analysis (Fig. 1 B, lane 5). This phosphorylation pattern was identical in two different buffer systems (Fig. 1 B, compare lanes 2 and 3); however, in the presence of Mn2+, phosphorylation of the AP2 large subunits was totally abolished, whereas that of μ2 was unaffected (lane 4). We later took advantage of the Mn2+ effect to study the sorting signal binding of AP2 selectively phosphorylated on μ2.
To obtain AP2 free of kinase activity, we immunoisolated AP2 from pool 4 using an α-adaptin–specific antibody. After immunoisolation, the immunocomplexes were extensively washed using high salt and Tween before incubation with [γ32P]ATP. Virtually no phosphorylation of any AP2 subunit was detectable (Fig. 1 C, lane 1), showing that immunoisolated AP2 was free of AP2-phosphorylating kinases. When immunoisolated AP2 was incubated with [γ32P]ATP and an aliquot of the HAP-derived pools (Fig. 1 A), the strong phosphorylation of a 50-kD protein corresponding to the AP2 μ2 subunit was detected (Fig. 1 C, lanes 2–4). The μ2-phosphorylating activity was highest in pools 3 and 4, but was also readily detectable in pool 2. In addition to μ2 phosphorylation, a weak but significant phosphorylation of the AP2 large subunits became apparent. Taken together, these experiments show that kinases that predominantly phosphorylate the μ2 subunit copurify with AP2 but can be separated by immunoadsorption and extensive washing.
AAK1, a μ2-phosphorylating kinase, copurifies with AP2
Two kinases have recently been shown to mediate phosphorylation of the μ2 subunit: GAK/auxilin-2 (Umeda et al., 2000) and the newly discovered AAK1 (Conner and Schmid, 2002). Whereas AAK1, which copurifies with adaptor fractions isolated by gel filtration chromatography, colocalizes to endocytic structures, GAK/auxilin-2 was shown to be predominantly cytosolic, with only a small fraction concentrated in perinuclear structures that colocalize with AP1 (Greener et al., 2000). Recombinant GAK/auxilin-2 can mediate μ2 phosphorylation, however, it is not known whether the kinase copurifies with AP2. We therefore analyzed the distribution of GAK/auxilin-2 and AAK1 during the AP2 purification (Fig. 2) . Both kinases can be detected in pig brain–derived CCVs (Fig. 2, lane 1); however, the two kinases were resolved by further fractionation. After extraction of CCVs with 0.5 M Tris, pH 7.8, most of GAK/auxilin-2 remained associated with the pelleted membranes (Fig. 2, lane 2), whereas ∼50% of AAK1 was recovered in the supernatant together with extracted coat proteins, including AP2. AAK1 was slightly enriched in the adaptor-containing fractions after gel filtration (lane 4), whereas GAK/auxilin-2 was further depleted from the adaptor-containing pool. After fractionation on HAP (Fig. 1 A), the remaining GAK/auxilin-2 was enriched in AP1-containing fractions (pool 2), but almost depleted from AP2-containing fractions (pool 3 and 4), consistent with its localization to the perinuclear region of cells (Greener et al., 2000). In contrast, AAK1 was detectable in pools 2–4, with a maximum in pool 4. The amount of 32P incorporated into μ2 correlated with the amount of AAK1 in pools 3 and 4, rather than with the amount of AP2 (Fig. 2 B), suggesting that AAK1 is the kinase that phosphorylates μ2 in pools 3 and 4.
Figure 2.

AAK1 copurifies with AP2 and phosphorylates μ2. (A) The presence of either GAK/auxilin-2 or AAK1 was analyzed throughout the purification procedure of AP2. Both kinases were detectable in crude CCVs. However, GAK was depleted during subsequent enrichment of AP2, whereas AAK1 copurified with AP2 to a significant degree (HAP, pools 3 and 4). (B) Incubation of pools 3 and 4, obtained after HAP chromatography (Fig. 1 A), revealed a much higher kinase activity toward μ2 in pool 4 than in pool 3, corresponding to the enrichment of AAK1 in pool 4.
AAK1 corresponds to the endogenous μ2 kinase activity associated with AP2
Our results suggest that AAK1 corresponds to the endogenous kinase activity responsible for phosphorylation of μ2. Indeed, when purified recombinant AAK1 was added to immunoisolated AP2 lacking associated kinase activity (Fig. 3 A, lane 1) and devoid of AAK1 (as revealed by Western blotting, unpublished data), μ2 was very efficiently phosphorylated (Fig. 3 A, lane 2). This demonstrates that the μ2 subunit of immobilized AP2 is a substrate for recombinant AAK1. We next compared the endogenous μ2 kinase activity associated with HAP-purified AP2 with the activity of recombinant AAK1 toward immunoisolated AP2 for their sensitivity to various kinase inhibitors. Phosphorylation of μ2 by both the endogenous kinase and purified AAK1 was abolished by N-ethylmaleimide treatment (Fig. 3 A, lane 3, NEM) and was inhibited in the presence of staurosporine, a broad spectrum inhibitor of PKC (lane 4). Additionally, the PKA-specific inhibitor KT5720 partially inhibited both endogenous kinase activity and that of purified recombinant AAK1 (Fig. 3 A, lane 7). None of the other kinase inhibitors with specificity for PKG, CaM kinase, or CKII (Fig. 3 A, lanes 5, 6, and 9, respectively), nor a more specific PKC inhibitor (lane 8) had any effect when present at concentrations known to selectively inhibit their target kinases. Thus, we were unable to distinguish AAK1 activity from that of the endogenous μ2-phosphorylating activity.
Figure 3.

Sensitivity of AP2-associated kinases toward kinase inhibitors. Phosphorylation of the μ2 subunit of AP2 after incubation in the presence of [γ32P]ATP was determined for either (A) immunoisolated AP2 incubated alone (lane 1) or in the presence (lanes 2–9) of purified recombinant AAK1, or (B) for HAP-purified AP2 with its copurifying kinase activity in the absence (lane 1) or presence of N-ethylmaleimide (NEM, 1 mM, lane 3), the broad spectrum PKC inhibitor staurosporine (20 nM, lane 4), the PKG inhibitor KT 5823 (250 nM, lane 5), the CaM inhibitor KN 93 (400 nM, lane 6), the PKA inhibitor KT 5720 (60 nM, lane 7), the PKC inhibitor bisindolylmalemide (10 nM, lane 8), or the CKII inhibitor DRB (6 μM, lane 9). After incubating for 20 min at RT, the samples were resolved by SDS-PAGE, and the amount of radiolabel incorporated into the μ subunit was quantified.
It has been reported that μ2 is phosphorylated at a single residue, Thr-156, by an endogenous kinase (Pauloin and Thurieau, 1993; Olusanya et al., 2001). However, the identity of the μ2-phosphorylating kinase remained unknown. Although we cannot formally exclude the contamination of purified AP2 (pool 4) by a yet unknown kinase, it is devoid of GAK and contains significant amounts of AAK1. Incubation of purified AP2 from pool 4 with [γ32P]ATP and subsequent analysis of μ2 by reverse phase HPLC, MALDI/MALDI MS, and radiosequencing of tryptic peptides revealed a single phosphorylation site to Thr-156 (Fig. S1, available online at http://www.jcb.org/cgi/content/full/jcb.200111068/DC1). Together, these data suggest that AAK1 corresponds to the endogenous kinase responsible for phosphorylation of μ2 at Thr-156.
μ2 phosphorylation by AAK1 mediates high affinity binding of AP2 to sorting signals
As mentioned above, phosphorylation of AP2 increases its affinity for membrane protein sorting signals (Fingerhut et al., 2001); however neither the kinase(s) involved, nor the AP2 subunit(s) whose phosphorylation was responsible for the increase in affinity had been identified. Given that μ2 is known to bind to tyrosine-based sorting signals (Aguilar et al., 1997; Owen and Evans, 1998), we first analyzed whether the selective phosphorylation of μ2 modulates AP2 binding affinity. The kinetics of AP2 binding were determined by surface plasmon resonance using a synthetic peptide representing the tyrosine-based sorting motif of lysosomal acid phosphatase (LAP) immobilized on a biosensor surface. In agreement with previously published data, HAP-purified AP2 (pool 4) effectively bound to the LAP tail (Fig. 4 , sensorgram 1), but its affinity decreased more than fourfold when AP2 was treated with phosphatase before the biosensor experiment (sensorgram 2), suggesting that AP2 in pool 4 was partially phosphorylated. In contrast, AP2 incubation with ATP resulted in an eightfold increase in affinity (Fig. 4, sensorgram 3) as compared with dephosphorylated AP2. This increase in binding affinity was sensitive to the addition of staurosporine (Fig. 4, sensorgram 5). Incubation of AP2 without ATP, but in the presence of staurosporine, did not decrease the binding affinity, as compared with the untreated control (unpublished data), indicating that pool 4 does not contain a μ2-specific phosphatase.
Figure 4.
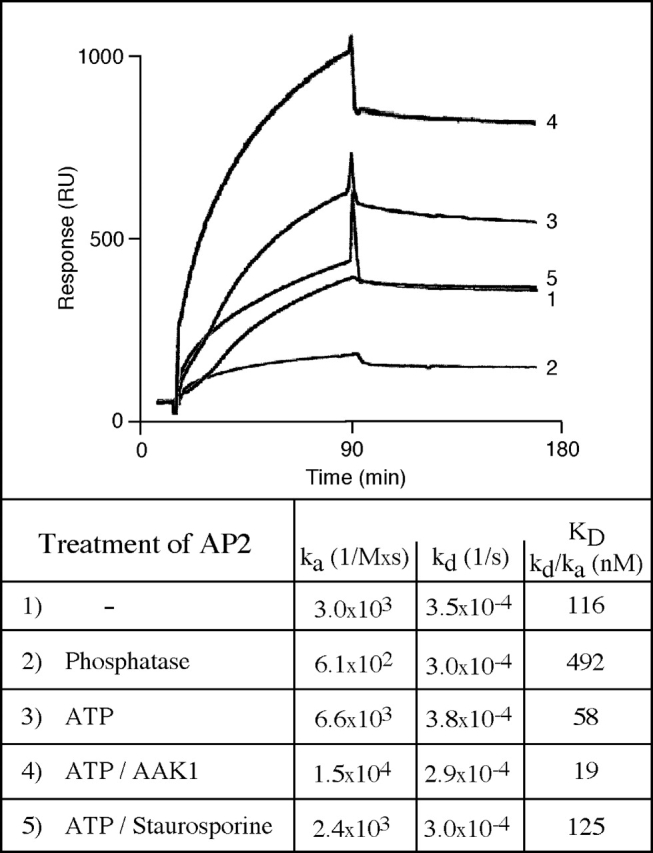
Phosphorylation of μ2 by an AP2-associated kinase or recombinant AAK1 increases the AP2 binding affinity for sorting signals. A peptide corresponding to the cytoplasmic tail of LAP was immobilized on the chip surface of a biosensor to monitor binding of AP2. AP2 was used without any treatment (sensorgram 1), after treatment with calf intestinal alkaline phosphatase (sensorgram 2), or after incubation with ATP in the absence (sensorgram 3) or presence (sensorgram 4) of recombinant AAK1 before the measurement of tail binding. The phosphorylation-dependent increase in AP2 binding affinity was sensitive to the kinase inhibitor staurosporine (sensorgram 5), resulting in values obtained for untreated AP2. Neither AAK1-phosphorylated nor untreated AP2 bound to a mutated LAP tail peptide in which the critical tyrosine residue was replaced by alanine (unpublished data).
The increase in AP2 affinity towards sorting signals, which was induced by μ2 phosphorylation, could also be, although unlikely, the result of AP2 aggregation induced by μ2 phosphorylation. To test this possibility, we incubated freshly prepared AP2 with phosphatase or ATP before analytical gel filtration on a Superdex-200 column. We found no difference in the elution profile of phosphatase- or ATP-treated AP2 compared with untreated controls (unpublished data), demonstrating that phosphorylation does not induce AP2 aggregation and arguing in favor of an increased affinity of μ2-phosphorylated versus nonphosphorylated AP2 complexes. In further experiments, we tested whether AAK1 phosphorylation of μ2 is sufficient to enhance the affinity of AP2 for sorting motifs. Incubation of AP2 with recombinant AAK1 and ATP (Fig. 4, sensorgram 4) increased the affinity about sixfold to 19 nM as compared with the untreated control (sensorgram 1). Most strikingly, these data indicate that the affinity of dephosphorylated AP2 (sensorgram 2) and AAK1-phosphorylated AP2 differ ∼25-fold. Thus, our in vitro experiments demonstrate that AP2 affinity for sorting signals can be significantly regulated by AAK1-mediated phosphorylation of μ2.
It is important to note that all biosensor experiments described here were performed in the presence of divalent Mn2+ ions to establish conditions that only allow μ2 phosphorylation, but selectively abolish the phosphorylation of α and β2 (Fig. 1 B). In parallel to the affinity measurements, we verified the selective phosphorylation of μ2 in identical samples under identical conditions, however in the presence of [γ32P]ATP (unpublished data). When the same biosensor experiments were performed without the addition of Mn2+, resulting in the weak but significant phosphorylation of α and β2, we did not observe any difference with the results presented here (unpublished data). Taken together, our in vitro data clearly show that the AAK1-mediated phosphorylation of the AP2 μ subunit alone is sufficient to modulate the affinity of AP2 towards sorting signals, whereas the phosphorylation status of the adaptor large subunits is of no relevance.
Our data is in line with and provide a convincing explanation for the recent findings that phosphorylation of μ2 on Thr-156 is critical for AP2 function in receptor-mediated endocytosis both in vitro and in vivo (Olusanya et al., 2001). Using a perforated A431 cell assay, the authors demonstrated that μ2 phosphorylation was required for AP2-stimulated transferrin sequestration into coated pits, and also showed that overexpression of μ2 (T156A) mutants inhibited receptor-mediated endocytosis. Our results now demonstrate that the inhibition of endocytosis can be attributed to the loss of efficient AP2 binding to the membrane protein sorting signals due to inactivation of the μ2 phosphorylation site Thr-156.
Interestingly, using the same perforated A431 cell assay, AAK1 was shown to inhibit AP2-dependent transferrin sequestration into constricted coated pits and coated vesicles in a kinase activity–dependent manner (Conner and Schmid, 2002). Taken together, these data are compatible with the view that continuous endocytosis requires cycles of phosphorylation/dephosphorylation of AP2, allowing the utilization of AP2 for multiple rounds of cargo recruitment. Alternatively, the inhibitory effect of excess AAK1 in this assay could reflect phosphorylation of other as yet unidentified substrates. Further work will be necessary to define the respective roles of AAK1 and GAK and to determine how these kinases are themselves regulated and how they regulate AP2 function and receptor-mediated endocytosis.
Materials and methods
Antibodies
Antibodies specific for AP1 and AP2 were obtained from Sigma-Aldrich, Transduction Laboratories, and Alexis Biochemicals Corp. A rabbit serum specific for μ2 was provided by M.S. Robinson (University of Cambridge, Cambridge, UK). GAK was identified by a monoclonal antibody (MoBiTec) or a rabbit serum provided by E. Ungewickell (Hannover Medical School, Hannover, Germany) and L. Greene (National Institutes of Health, Bethesda, MD). AAK was identified by a rabbit serum raised against the COOH-terminal AP2 binding domain (Conner and Schmid, 2002).
Preparation of AP2
The preparation of CCVs from porcine brain, the Tris extraction of coat proteins from CCVs, the Superose 6 gel filtration during which adaptors are enriched, and the HAP chromatography for the separation of AP2 from AP1 were performed exactly as previously described (Fingerhut et al., 2001). The purity of AP1 and AP2 was analyzed by SDS-PAGE and Western blotting, using specific antibodies for AP1 and AP2. Fractions containing AP2 were pooled and dialyzed into either buffer A (25 mM Hepes, 125 mM KOAc, 10 mM NaCl, 5 mM MgCl2, 1 mM DTT, pH 7.2) or buffer B (100 mM Tris-Cl, 1 mM DTT, pH 7.2), as indicated. In some experiments, buffer A or B was supplemented with 2 mM MnCl2.
Immunoisolation of AP2 was performed by first absorbing the anti–α-adaptin antibody AP.6 (Alexis Biochemicals Corp.) to protein A–Sepharose (Sigma-Aldrich) followed by incubation with AP2 for 4 h at 4°C. After immunoisolation, the immune complexes were washed with 0.5 M NaCl in PBS and 0.1% Tween 20 in PBS before they were used for further experiments.
In vitro phosphorylation and dephosphorylation of AP2
AP2 dialyzed into buffer A, was incubated with 1 μl [γ32P]ATP (3,000 Ci/mmol; Amersham Pharmacia Biotech) for 20 min at room temperature (RT). The 20-μl reaction was stopped by the addition of sample buffer, boiled for 5 min at 95°C, and analyzed by SDS-PAGE, autoradiography, and Western blotting with antibodies against the μ2 subunit. The phosphorylation of immunoisolated AP2 (20 μl) was performed in 50 μl of buffer A or in 50 μl of pooled kinase-containing fractions (Fig. 1). In some experiments, recombinant AAK1, purified as previously described (Conner and Schmid, 2002), was added to AP2 under the same buffer conditions at a molar ratio of 1:20 (AAK1/AP2). The reaction was stopped by washing three times with ice-cold PBS, containing 0.1% Tween 20. Samples were analyzed as above. The kinase inhibitor experiments are described in the legend of Fig. 3. All inhibitors (Alexis Biochemicals Corp.) were used at concentrations recommended to abolish their target kinases. For phosphorylation with unlabeled ATP, AP2 was incubated for 20 min at 37°C in the presence of 2 mM ATP in buffer A or B, with or without Mn2+.
Dephosphorylation of AP2 was performed in buffer A or B with various amounts of calf intestinal alkaline phosphatase (Boehringer) for 20 min at 37°C.
Biosensor-based interaction analysis
The interaction between AP2 and the cytoplasmic tail peptides was analyzed in real time by surface plasmon resonance, using a BIAcore3000 biosensor (BIAcore AB). Immobilization of peptides corresponding to the tail sequence from LAP to the biosensor surface was exactly as described by Fingerhut et al. (2001). The experiments were performed at a flow rate of 20 μl/min, and AP2 was used at concentrations of 50 and 250 nM. Association (1–2 min) was followed by dissociation (2 min), during which buffer A was perfused. Subsequently, bound AP2 was removed by a short pulse injection (15 s) of 10 mM NaOH containing 0.5% SDS.
The rate constants (k a for association, k d for dissociation, and the equilibrium rate constant K D = k d/k a) for the interaction between the tail peptides and AP2 were calculated as previously described (Höning et al., 1997; Fingerhut et al., 2001).
Online supplemental material
Fig. S1 (available online at http://www.jcb.org/cgi/content/full/jcb.200111068/DC1) shows results from reverse phase HPLC purification of phosphopeptides after tryptic digestion of phosphorylated μ2, which was used together with MALDI/MALDI MS and radiosequencing to map the phosphorylation site.
Supplemental Material
Acknowledgments
We acknowledge E. Smythe (University of Dundee, Dundee, UK) for sharing unpublished results. We thank Klaus Neifer and Jianhe Peng for their help during the identification of the μ2 phosphorylation site.
This work was supported by grants from the European Union (XCT 960058), the Deutsche Forschungsgemeinschaft (SFB 523/TP A5), the Fonds der Chemischen Industrie, and the National Institutes of Health (R37-MH61345 to S.L. Schmid and GM20632 to S.D. Conner).
The online version of this article includes supplemental material.
Footnotes
Abbreviations used in this paper: AAK, adaptor-associated kinase; CCV, clathrin-coated vesicle; HAP, hydroxyapatite; LAP, lysosomal acid phosphatase; RT, room temperature.
References
- Aguilar, R., H. Ohno, K. Roche, and J. Bonifacino. 1997. Functional domain mapping of the clathrin-associated adaptor medium chains μ1 and μ2. J. Biol. Chem. 272:27160–27166. [DOI] [PubMed] [Google Scholar]
- Bar-Zvi, D., and D. Branton. 1986. Clathrin-coated vesicles contain two protein kinase activities. Phosphorylation of clathrin beta-light chain by casein kinase II. J. Biol. Chem. 261:9614–9621. [PubMed] [Google Scholar]
- Campbell, C., J. Squicciarini, M. Shia, P.F. Pilch, and R.E. Fine. 1984. Identification of a protein kinase as an intrinsic component of rat liver coated vesicles. Biochemistry. 23:4420–4426. [DOI] [PubMed] [Google Scholar]
- Conner, S.D., and S.L. Schmid. 2002. Identification of an adaptor-associated kinase, AAK1, as a regulator of clathrin-mediated endocytosis. J. Cell Biol. 156:921–929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cousin, M.A., and P.J. Robinson. 2001. The dephosphins: dephosphorylation by calcineurin triggers synaptic vesicle endocytosis. Trends Neurosci. 24:659–665. [DOI] [PubMed] [Google Scholar]
- Fingerhut, A., K. von Figura, and S. Höning. 2001. Binding of AP2 to sorting signals is modulated by AP2 phosphorylation. J. Biol. Chem. 276:5476–5482. [DOI] [PubMed] [Google Scholar]
- Goodman, O.B., and J.H. Keen. 1995. The alpha chain of the AP-2 adaptor is a clathrin binding subunit. J. Biol. Chem. 270:23768–23773. [DOI] [PubMed] [Google Scholar]
- Greener, T., X. Zhao, H. Nojima, E. Eisenberg, and L.E. Greene. 2000. Role of cyclin G-associated kinase in uncoating clathrin-coated vesicles from non-neuronal cells. J. Biol. Chem. 275:1365–1370. [DOI] [PubMed] [Google Scholar]
- Höning, S., M. Sosa, A. Hille-Rehfeld, and K. von Figura. 1997. The 46-kDa mannose 6-phosphate receptor contains multiple binding sites for clathrin adaptors. J. Biol. Chem. 272:19884–19890. [DOI] [PubMed] [Google Scholar]
- Kirchhausen, T. 1999. Adaptors for clathrin-mediated traffic. Annu. Rev. Cell Dev. Biol. 15:705–732. [DOI] [PubMed] [Google Scholar]
- Olusanya, O., P.D. Andrews, J.R. Swedlow, and E. Smythe. 2001. Phosphorylation of threonine 156 of the μ2 subunit of the AP2 complex is essential for endocytosis in vitro and in vivo. Curr. Biol. 11:896–900. [DOI] [PubMed] [Google Scholar]
- Owen, D., and P. Evans. 1998. A structural explanation for the recognition of tyrosine-based endocytotic signals. Science. 282:1327–1332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pauloin, A., and C. Thurieau. 1993. The 50 kDa protein subunit of assembly polypeptide (AP) AP-2 adaptor from clathrin-coated vesicles is phosphorylated on threonine-156 by AP-1 and a soluble AP50 kinase which co-purifies with the assembly polypeptides. Biochem. J. 296:409–415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pauloin, A., J. Loeb, and P. Jolles. 1984. Protein kinase(s) in bovine brain coated vesicles. Biochim. Biophys. Acta. 799:238–245. [DOI] [PubMed] [Google Scholar]
- Shih, W., A. Gallusser, and T. Kirchhausen. 1995. A clathrin-binding site in the hinge of the beta 2 chain of mammalian AP-2 complexes. J. Biol. Chem. 270:31083–31090. [DOI] [PubMed] [Google Scholar]
- Slepnev, V., G. Ochoa, M. Butler, D. Grabs, and P. De Camilli. 1998. Role of phosphorylation in regulation of the assembly of endocytic coat complexes. Science. 281:821–824. [DOI] [PubMed] [Google Scholar]
- Umeda, A., A. Meyerholz, and E. Ungewickell. 2000. Identification of the universal cofactor (auxilin 2) in clathrin coat dissociation. Eur. J. Cell Biol. 79:336–342. [DOI] [PubMed] [Google Scholar]
- Wilde, A., and F.M. Brodsky. 1996. In vivo phosphorylation of adaptors regulates their interaction with clathrin. J. Cell Biol. 135:635–645. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}