

Abstract
Skeletal muscles display a remarkable diversity in their arrangement of fibers into fascicles and in their patterns of innervation, depending on functional requirements and species differences. Most human muscle fascicles, despite their great length, consist of fibers that extend continuously from one tendon to the other with a single nerve endplate band. Other mammalian muscles have multiple endplate bands and fibers that do not insert into both tendons but terminate intrafascicularly. We investigated whether these alternate structural features may dictate different modes of cell hypertrophy in two mouse gracilis muscles, in response to expression of a muscle-specific insulin-like growth factor (IGF)-1 transgene (mIGF-1) or to chronic exercise. Both hypertrophic stimuli independently activated GATA-2 expression and increased muscle cross-sectional area in both muscle types, with additive effects in exercising myosin light chain/mIGF transgenic mice, but without increasing fiber number. In singly innervated gracilis posterior muscle, hypertrophy was characterized by a greater average diameter of individual fibers, and centralized nuclei. In contrast, hypertrophic gracilis anterior muscle, which is multiply innervated, contained longer muscle fibers, with no increase in average diameter, or in centralized nuclei. Different modes of muscle hypertrophy in domestic and laboratory animals have important implications for building appropriate models of human neuromuscular disease.
Keywords: skeletal muscle hypertrophy; intrafascicularly terminating fibers; IGF-1; exercise; GATA-2
Introduction
Skeletal muscle hypertrophy results in an increased amount of contractile tissue. In rodents, commonly studied leg muscles such as soleus, extensor digitorum longus (EDL),* and thigh muscles contain fibers that extend continuously from one tendon to the other, with a single zone of neuromuscular junctions in their middle. Hypertrophy of these muscles reflects an increase in muscle fiber diameter without an increase in the number of muscle fibers (Gollnick et al., 1981; Timson and Dudenhoeffer, 1990).
Many muscles in larger mammals have multiple endplate bands and muscle fibers that do not extend the entire length between the two tendons but terminate intrafascicularly. Animals with multiply innervated muscles include rabbit, cat, goat, horse, cattle (Coers and Durand, 1957; Adams and MacKay, 1961; Loeb et al., 1987; Gans et al., 1989; Ryan et al., 1992; Purslow and Trotter, 1994), frogs (Cooper, 1925), and chicken (Gaunt and Gans, 1990). Multiple endplate bands are found in muscles of nonprimate animals that exceed a fascicle length of 3.5 cm and the number of endplate bands increases regularly with increasing fascicle length (Paul, 2001). Intrafascicularly terminating fibers taper over much of their length by reducing their diameter in steps characterized by folding of the sarcolemma and formation of myo–myonal junctions with neighboring fibers (Young et al., 2000).
In contrast to other mammals, most human and macaque monkey limb muscles are singly innervated up to a fascicle length of 13 cm and predominantly consist of long, singly innervated muscle fibers that insert into both tendons (Paul, 2001). In humans, only sartorius and gracilis muscles with fascicles up to 50 cm long are multiply innervated (Schwarzacher, 1959; Paul, 2001). Specific adaptations of human muscles to compensate for these very long fibers are unknown, but they may have been evolutionarily selected for increased fine motor control. The neuromuscular anatomy of human muscles is therefore distinct from that of animals used in laboratory experiments (e.g., rabbit and cat), bred for their meat (e.g., cattle), or sporting performance (e.g., horses).
In response to a hypertrophic stimulus, muscle fibers may increase in diameter, as is found in singly innervated muscles, to increase the number of myofibrils in parallel. Alternatively, intrafascicularly terminating fibers could elongate, effectively increasing the number of fibers as well as myofibrils in parallel.
Several practical issues have delayed the investigation of these two hypotheses. First, the existence of multiply innervated muscles with intrafascicularly terminating fibers has long been ignored in the general literature. Second, the total number of muscle fibers must be counted in the muscle of concern. Muscles from cattle for example may have twelve endplate bands, twelve in-series sets of intrafascicularly terminating fibers, and contain tens of thousands of muscle fibers (Swatland and Cassens, 1972).
In the present study, the response to hypertrophic stimuli was investigated in mouse gracilis anterior, one of two multiply innervated muscles in the mouse, as in the human. Gracilis anterior muscle has two endplate bands and intrafascicularly terminating fibers. This muscle particularly suited our investigations since there are only two sets of intrafascicularly terminating fibers in series, reducing the muscle's complexity. Moreover, the entire muscle contains only 500–600 fibers, easily counted in cross-section or by acid digestion. The response of gracilis anterior muscle to hypertrophic stimuli was compared with that of gracilis posterior muscle, which lies close to gracilis anterior muscles, and has a similar function, but only has a single endplate band and no intrafascicularly terminating fibers. The dramatically different modes of hypertrophy found in the two types of muscles underscores the complexity of skeletal muscle phenotypes and the need for reassessment of appropriate animal models for human myopathies.
Results
Neuromuscular anatomy of mouse gracilis muscles
Single or multiple endplate bands were visualized in mouse gracilis muscles by detecting a visible acetylcholinesterase (AChE) precipitate at neuromuscular junctions, muscle tendon junctions, and myo-myonal junctions between intrafascicularly terminating fibers. In the gracilis posterior muscle a single band of endplates was present in the center of the muscle (arrow, Fig. 1 A) and no myo–myonal junctions were found. In the gracilis anterior muscle two endplate bands were present (arrows, Fig. 1 B), as were myo–myonal junctions at the ends of intrafascicularly terminating fibers (arrowheads) between the two endplate bands. AChE staining of longitudinally sectioned tissue illustrates the unique morphologies of neuromuscular junctions (Fig. 1 C) and intrafascicular terminations (Fig. 1 D) of the gracilis anterior muscle.
Figure 1.

AChE precipitate showing pattern of innervation in singly and multiply innervated muscles. (A) Gracilis posterior muscles of the mouse have a single endplate band in the center of the muscle (arrow) and no intrafascicularly terminating fibers. (B) Gracilis anterior muscle of the mouse has two endplate bands (arrows) and intrafascicularly terminating fibers whose ends also show the AChE precipitate (arrowheads). (C) Neuromuscular junction labeled with AChE precipitate. (D) Myo–myonal junction between two intrafascicularly terminating fibers (white and black asterisks) labeled with AChE precipitate.
Cumulative effects of genetic and physiologic hypertrophy
We challenged mice with several paradigms of muscle hypertrophy to determine their effect on singly and multiply innervated muscles. Previous work has shown that the myosin light chain (MLC)/insulin-like growth factor transgene (mIGF-1) induces hypertrophy in singly innervated muscles, resulting in increased fiber diameter and muscle mass (Barton-Davis et al., 1998; Musarò et al., 2001). As described recently, voluntary wheel running in mice also increases fiber area of the singly innervated gastrocnemius and tibialis anterior muscles (Allen et al., 2001). We developed a similar voluntary exercise program using commercially available small animal running wheels (see Materials and methods). After an initial period, most mice recorded up to 6 h and 5 km of running during a 24-h time period regardless of genotype. Mice were allowed to exercise for 4 wk, after which muscle hypertrophy was analyzed in wild-type (WT), MLC/mIGF-1 transgenic (IGF-1), exercised WT (WT-EX), and exercised MLC/mIGF-1 transgenic (IGF-EX). As seen in Fig. 2 A, no change in body weight was found between WT and WT-EX mice (30 g), whereas transgenic IGF (35 g) and IGF-EX (40 g) mice were significantly heavier than their WT counterparts (P ≤ 0.05). Moreover, EX-IGF mice were significantly (P ≤ 0.01) heavier than inactive IGF mice.
Figure 2.

Skeletal muscle hypertrophy of inactive and exercised WT and mIGF transgenic mice. (A) Comparison of body weights shows a significant increase in IGF and IGF-EX compared with WT animals, with IGF-EX animals weighing the most. (B) Muscle cross-sectional area increased significantly in gracilis anterior, gracilis posterior, and soleus muscles, indicating hypertrophy in these muscles. Since the MLC/mIGF-1 transgene is expressed at very low levels in soleus (Musarò et al., 2001) the increase in the average cross-sectional area of soleus muscle may reflect a secondary effect of increased loading in neighboring muscles, resulting in a transition to a faster fiber type.
Muscle hypertrophy in each WT, IGF, WT-EX, and IGF-EX category was calculated from the cross-sectional area for gracilis anterior, gracilis posterior, and soleus muscles (Fig. 2 B). IGF, WT-EX, and IGF-EX muscles had a significantly larger cross-sectional area when compared with WT muscles, with IGF-EX muscles showing the greatest increase. WT-EX gracilis anterior muscle was the exception and showed no significant increase compared with WT muscle. These results highlight the additive effects of exercise on the increase in diameter of singly and multiply innervated muscles in MLC/mIGF-1 transgenic animals.
Induction of mIGF-1 expression in exercise-induced hypertrophy
Since overexpression of localized mIGF-1 under the control of MLC1/3 regulatory elements resulted in hypertrophy of skeletal myocytes (Musarò et al., 1999, 2001), the additive effect of exercise on the hypertrophic phenotype in MLC/mIGF-1 mice could be due to elevated expression of the mIGF-1 transgene. Northern blot analysis with a probe recognizing the mIGF-1 isoform detected little expression in WT thigh muscle, with no increase in the WT-EX muscle (Fig. 3). In contrast, the IGF-EX transgenic muscle responded to exercise by increasing transgenic mIGF-1 RNA levels. Thus, although the endogenous mIGF-1 gene is not induced in exercised muscles, the MLC regulatory elements driving the mIGF-1 transgene are further stimulated by running exercise.
Figure 3.

Northern blot showing levels of mIGF-1 in thigh muscles from WT, IGF, WT-EX, and IGF-EX using a the full-length probe against the tissue-restricted isoform of IGF-1. This week-long exposure shows no IGF-1 RNA in WT and WT-EX muscles. Only in the MLC/mIGF-1 transgenic mice is the gene expressed, with an upregulation in IGF-EX. The endogenous gene is not expressed in any specimen. The endogenous IGF-1 gene is therefore not upregulated with voluntary wheel-running exercise. The transgene is under the control of the MLC promoter and is upregulated with exercise.
Markers of exercise-induced hypertrophy
Although the GATA-2 transcription factor is normally absent from skeletal muscles at all stages of development (M. McGrew, personal communication) it is induced both in muscle cell culture and in transgenic mice in response to mIGF-1 expression (Musarò et al., 1999, 2001). GATA-2 is therefore implicated as a molecular marker of hypertrophic muscle. To determine whether GATA-2 was also expressed in exercise-induced hypertrophy, we compared GATA-2 protein levels in gracilis anterior and posterior muscles of WT, IGF, WT-EX, and IGF-EX mice. As seen in Fig. 4, IGF and WT-EX muscles showed a similar level of hypertrophy accompanied by increased GATA-2 expression, whereas the combined effects of exercise and the MLC/IGF-1 transgene in the IGF-EX mice resulted in exceptionally high GATA-2 expression. GATA-2 protein accumulated in the sarcolemma, was enriched toward intrafascicular terminations in IGF and WT-EX gracilis anterior muscles, increased in IGF-EX gracilis anterior and posterior muscles (Fig. 5), and was unexpectedly excluded from nuclei (insert Fig. 5 D). Induction of GATA-2 expression is therefore a general marker of the hypertrophic response, regardless of stimulus, in both singly and multiply innervated muscles.
Figure 4.

Western blot using an antibody against GATA-2 transcription factor, a marker for skeletal muscle hypertrophy. In both gracilis anterior and posterior the WT muscle contained the least amount of GATA-2 protein, followed by an equal level in IGF and WT-EX muscles. IGF-EX muscles had the highest expression of GATA-2 protein, indicating that exercise and the mIGF-1 transgene synergize to increase muscle hypertrophy.
Figure 5.
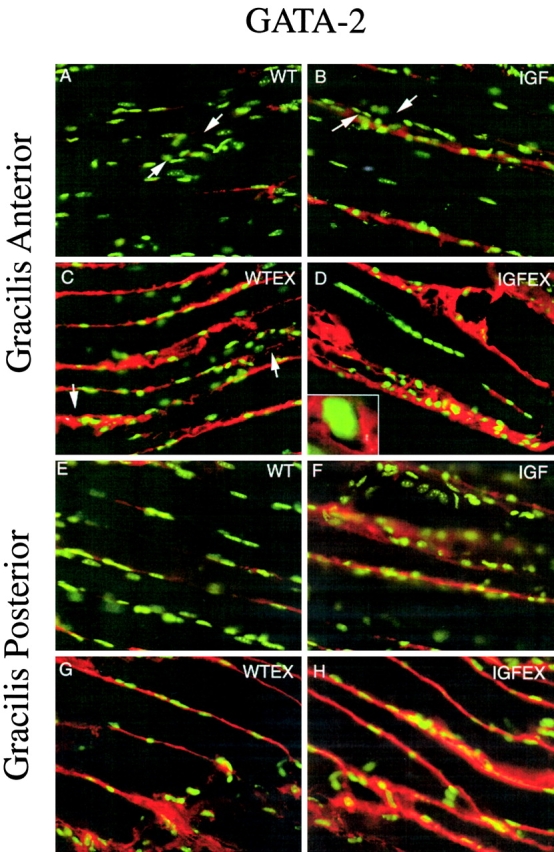
GATA-2 localization on gracilis anterior or posterior muscles with Hoechst (green fluorescence) and AChE (brightfield) stains. (A–D) Gracilis Anterior with AChE precipitate between arrows. (A) WT, (B) MLC/mIGF-1 transgenic, (C) WT-EX, (D) MLC/mIGF-1 transgenic exercised, and (D, insert) GATA-2 is excluded from the nucleus. (E–H) Gracilis posterior. (E) WT, (F) MLC/mIGF-1 transgenic, (G) WT-EX, (H) MLC/mIGF-1 transgenic exercised. GATA-2 protein is upregulated in IGF and WT-EX, with a further increase in IGF-EX.
Neonatal myosin, a standard marker for hypertrophy in singly innervated muscles (Walro et al., 1991), was also up-regulated along the entire length of fibers in WT-EX (Fig. 6 C) and IGF-EX (Fig. 6 D) gracilis anterior, suggesting that induction of muscle myosins appears to be a general response to the hypertrophic process.
Figure 6.

Neonatal myosin localization on gracilis anterior muscles with Hoechst (green fluorescence) and AChE (brightfield) stains. (A–D) Gracilis anterior with AChE precipitate between arrows. (A) WT, (B) MLC/mIGF-1 transgenic, (C) WT-EX, (D) MLC/mIGF-1 transgenic exercised. Neonatal myosin is upregulated along the length of fibers in WT-EX and IGF-EX muscles.
Muscle fiber type changes with exercise-induced hypertrophy
The effects of transgenic or exercise-induced hypertrophy on shifts in muscle fiber type were investigated by scoring the percentage of type I, type IIB, and type IIA/X myosin heavy chain–positive fibers in gracilis anterior and posterior muscles. Minimal fiber type changes have been observed previously in the MLC/mIGF-1 transgenic mice (Musarò et al., 2001), whereas significant fiber type changes have been observed with voluntary exercise (Allen et al., 2001). Gracilis anterior muscles (Fig. 7 A) showed decreased type I and type IIA/X MyHC–positive fibers, with an increase in type IIB MyHC–positive fibers, although the trend was not statistically significant. Exercise, rather than the expression of the MLC/mIGF-1 transgene appears to be the determinant of fiber type changes in this muscle, since only muscles from the WT-EX and IGF-EX showed a significant increase in type IIB MyHC–positive fibers.
Figure 7.
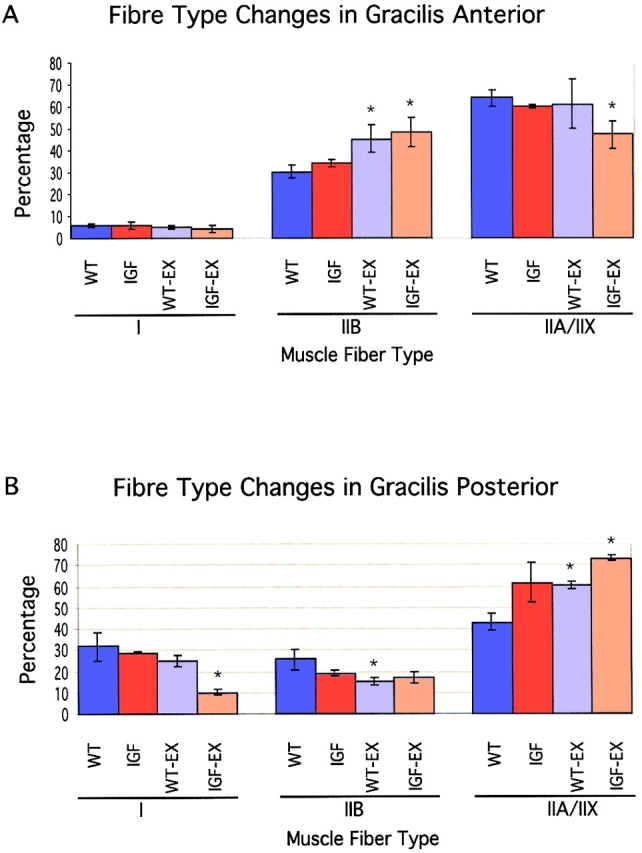
Trend in fiber type changes in gracilis anterior and posterior muscle influenced by exercise not transgene. (A) Gracilis anterior muscle showed a trend of increasing type IIB MyHC at the cost of type IIA/X MyHC–positive fibers. A significant difference was only found in WT-EX and transgenic mice. (B) Fiber type changes in gracilis posterior muscle showed a trend toward type IIA/X MyHC–positive fibers at the cost of fibers expressing type IIB and type I MyHC. Exercise appears to be the main influence on these trends.
Gracilis posterior muscle (Fig. 7 B) also showed a slight decrease in the number of type I MyHC–positive fibers with a trend toward a greater number of type IIA/X MyHC–positive fibers at the cost of type IIB fibers. The preferential increase in type IIA/X over type IIB–positive fibers in this muscle compared with gracilis anterior muscle likely reflects the specific activity patterns and loading of these muscles. These results indicate that the proportion of fiber phenotype is predominantly influenced by exercise in both the singly and the multiply innervated muscle.
Distinct modes of hypertrophy in singly and multiply innervated muscles
In response to a hypertrophic stimulus, the fibers in singly innervated muscles such as soleus and gracilis posterior increase in diameter (Fig. 8 B). However, intrafascicularly terminating fibers in multiply innervated muscles such as gracilis posterior could also elongate, effectively increasing the overall diameter of the muscle. To distinguish between these two modes of hypertrophy in multiply innervated muscles, we compared the number of muscle fibers in cross-sections of gracilis anterior, gracilis posterior, and soleus muscles from WT, WT-EX, IGF, and IGF-EX (Fig. 8 A) and calculated the average fiber area in each group (Fig. 8 B). Note that the fibers of gracilis anterior and posterior muscles are arranged in parallel, thus allowing perpendicular transverse sections of all fascicles, whereas soleus muscle has a pinnate arrangement of fibers.
Figure 8.

Number and diameter of muscle fibers. (A) The number of muscle fibers increased significantly only in gracilis anterior muscle and not gracilis posterior or soleus muscles. (B) The average fiber area increased significantly in gracilis posterior muscle and in IGF-EX gracilis anterior muscle only. Singly innervated muscles therefore respond to hypertrophic signals by increasing average fiber area and not number. Gracilis anterior muscle, which contains intrafascicularly terminating fibers, responds to hypertrophic signals by increasing the number of muscle fibers in cross-section.
Gracilis posterior showed a significant increase (P ≤ 0.05) in the average fiber area without an increase in fiber number. Fiber area from IGF-EX mice showed the largest increase (Fig. 8 B), confirming that exercise and overexpression of mIGF-1 have a synergistic effect on fiber hypertrophy. Soleus muscles only had a slight, but not significant increase in either muscle fiber number or in average fiber area (Fig. 8 A). However, these small increases, enhanced by increased pinnation of fibers with hypertrophy, resulted in an overall significant increase in muscle diameter.
In contrast, counts of muscle fibers in cross-sections of gracilis anterior were increased in all three modes of hypertrophy (WT-EX, IGF, or IGF-EX). Additional muscle fibers could either derive from the formation of new muscle fibers, or from the elongation of existing intrafascicular fibers. To distinguish between these two possibilities, we counted the total number of muscle fibers in WT and IGF gracilis anterior muscles by acid digestion, which removes connective tissue (Fig. 9 A) and allows physical separation of individual fibers within a muscle, including short intrafascicularly terminating fibers. As expected, the fiber count in acid-digested WT gracilis anterior muscles exceeded the fibers counted in transverse sections, since not all fibers could be scored in a single cross-sectional area. However, the total number of fibers was equivalent in WT and IGF gracilis anterior muscle (Fig. 9 B). Moreover, the fiber count for IGF gracilis anterior muscles closely matched the number of muscle fibers counted in transverse sections (Fig. 8 A). The increase in muscle fiber number in hypertrophic gracilis anterior muscle therefore must result from elongation of existing fibers and not from the formation of new fibers (Fig. 10).
Figure 9.
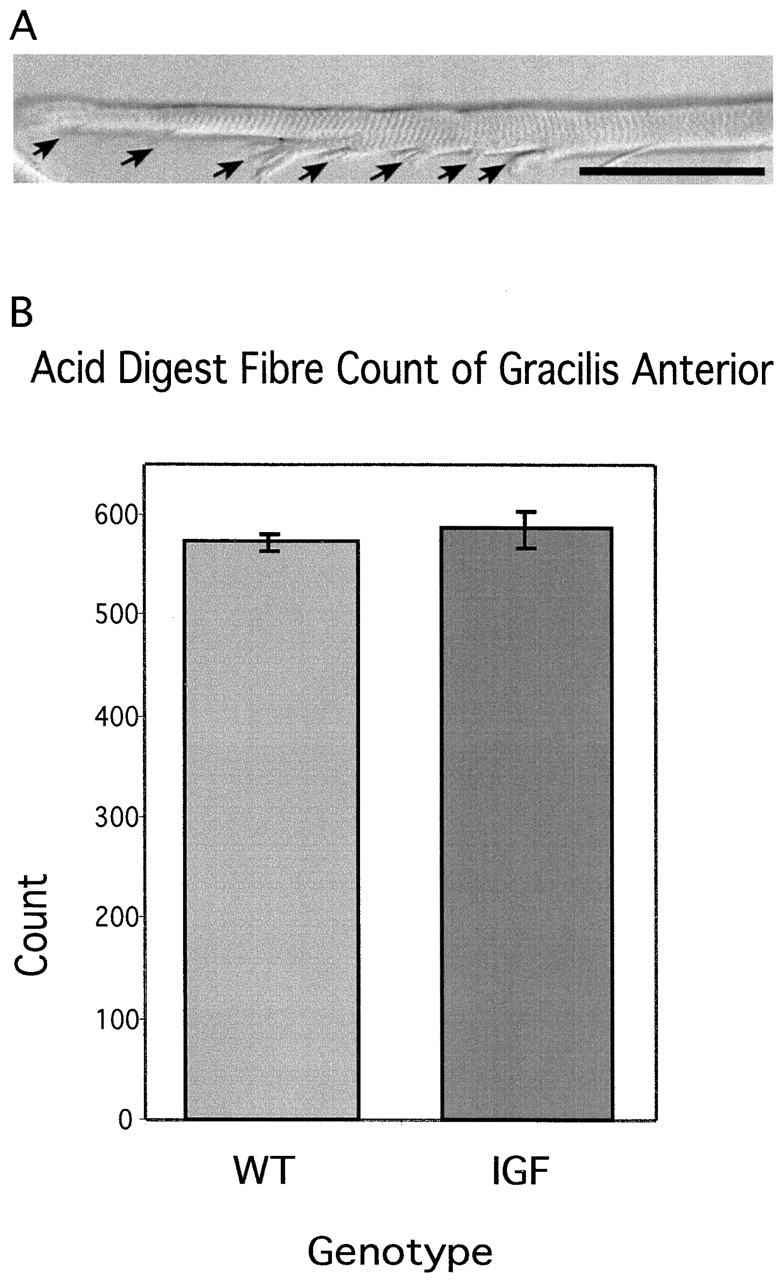
Counts of total fiber number in gracilis anterior after acid digestion. (A) Acid digestion of gracilis anterior muscle separates intrafascicularly terminating fibers that taper in steps (arrows). (B) Counts of all muscles fibers in WT and mIGF-1 transgenic mice after acid digest does not show a difference in total fiber number. Therefore, the increase in fiber number seen in cross-section is the result of elongation of existing intrafascicularly terminating muscle fibers.
Figure 10.

Model of modes of skeletal muscle hypertrophy. (A) Skeletal muscles with a single endplate band hypertrophy by increasing the diameter of individual fibers. (B) Intrafascicularly terminating fibers (IFTs) in a muscle with two endplate bands hypertrophy by elongating, causing an increase in the number of fibers in transverse section (TS).
One of the hallmarks of regenerating skeletal muscle is the occurrence of centralized nuclei within affected myofibers. A general increase in centralized nuclei has been reported in singly innervated muscles after infection with a viral vector carrying the MLC/mIGF-1 transgene (Barton-Davis et al., 1998). To determine whether multiply innervated muscles displayed a similar response to hypertrophic stimuli, we compared the average number of centralized nuclei in gracilis anterior and posterior muscles of WT, IGF, WT-EX, and IGF-EX mice. As seen in Fig. 11, gracilis posterior muscles from IGF animals showed a modest increase in centralized nuclei, which rose substantially with exercise. In contrast, gracilis anterior muscles did not accumulate centralized nuclei in any of the hypertrophic models tested. Thus, modes of hypertrophy in singly and multiply innervated muscles can be distinguished by both morphological and subcellular criteria.
Figure 11.

Percentage of muscle fibers with centralized nuclei in gracilis anterior and posterior muscles. In the gracilis posterior the number of centralized nuclei was increased in mIGF animals, with a large increase in the IGF-EX muscle, indicating hypertrophy along the length of fibers. In contrast, the number of centralized nuclei was not increased in gracilis anterior muscle.
Discussion
Not all muscles are created equal, nowhere illustrated more clearly than in a comparison of the distinct architectures of singly and multiply innervated muscles. Multiply innervated muscles with their short, in-series fibers are found in a large range of species, though only two such muscles have been found in humans. Since skeletal muscle is a composite tissue, multiple structural components need to be modified when muscle fibers elongate. Intrafascicularly terminating fibers are embedded and interlinked to their extracellular matrix by integrin- and dystrophin-associated complexes, closely interdigitating with other fibers and forming myo-myonal junctions with additional trans-membrane links (unpublished data). To facilitate elongation, the relationship between neighboring fibers must necessarily change and involve the breaking of old connections and the formation of new junctions, with their associated trans-membrane and morphological specializations. An appreciation of the variations in anatomical and physiological characteristics of different muscle types is therefore critical to the design of appropriate models for animal growth and human muscle diseases.
Hypertrophic stimuli have similar and additive effects
Hypertrophy was induced in mouse skeletal muscle by voluntary exercise and by expression of the MLC/mIGF-1 gene. The response to these two types of hypertrophic signals was similar in both singly and multiply innervated muscles, producing equivalent changes in fiber diameter, number, and phenotype, and similar induction of GATA-2 protein expression. Either model can therefore be used to induce hypertrophy, although wheel-running exercise does not appear to induce hypertrophy by increasing endogenous mIGF-1 RNA levels. Further experiments will be necessary to determine whether exercised and MLC/mIGF-1 transgenic mice respond similarly to age, unloading, injury, and disease.
The combination of exercise and the increased action of the MLC/mIGF transgene significantly enhanced the hypertrophic effects of MLC/mIGF-1 expression in transgenic mice with the activation of similar markers. These results have several interesting ramifications. First, they suggest that the two forms of hypertrophic stimuli are cooperative. Although the endogenous IGF-1 gene is activated in skeletal muscles under conditions of injury (Musarò et al., 2001), the combination of mechanical stress produced by exercise and supplemental mIGF-1 expression resulted in a more robust anabolic response than either stimulus alone could induce. Second, it confirms that the hypertrophy observed in MLC/mIGF-1 transgenic lines does not represent a physical limit on muscle growth. Exercised MLC/mIGF-1 transgenic animals did not exhibit muscle dysmorphology associated with over-exercise or injury. Finally, the cumulative effects of exercise and mIGF-1 transgene expression were distinct in singly and multiply innervated muscles, suggesting that the mode of hypertrophy employed by each muscle group is an inherent characteristic that is independent of the form of hypertrophic stimulus. Further delineation of these inherent differences will undoubtedly yield important insights into the cell biology of muscle hypertrophy.
Shifting definitions of skeletal muscle hypertrophy
Hypertrophy has the overall effect of enhancing muscle strength by increasing the number of contractile myofibrils. The source of the additional myofibrils remains the cause of some debate, as is the nature of the accompanying increase in muscle fiber size. In tissues composed of mononucleated cells, such as cardiac muscle, hypertrophy is defined as an increase in the ratio of cytoplasm to nucleus, whereas hyperplasia is defined as nuclear division and an increase in the number of cells. In contrast, hypertrophy of skeletal muscle is thought to involve an increase in the number of nuclei, as well as an increase in cytoplasmic volume. Hypertrophy of skeletal muscle therefore incorporates part of the definition for hyperplasia, an increase in the number of nuclei. As a further complication, the additional nuclei may come from internal nuclear division or fused satellite cells and their donated nuclei. Thus, evidence of DNA synthesis, a hallmark of hyperplasia in other tissues, is not always illuminating in skeletal muscle as the increase in muscle fiber size is greater than the volume contributed by fusing mononucleated cells. In addition, myoblasts could fuse with each other to form new muscle fibers.
It is therefore suggested that the definitions of hyperplasia and hypertrophy in skeletal muscle should pertain to features over which there is no confusion, e.g., the number of muscle fibers. According to this definition, hypertrophy of skeletal muscle is an increase in the size of fibers without an increase in their number, irrespective of any increase in the number of nuclei per fiber. Consistent with this definition, we demonstrate here that intrafascicularly terminating fibers are elongated in response to hypertrophic stimuli without an increase in fiber number.
The present study also raises the possibility that some reports have misinterpreted the intricate arrangements of elongating fibers as increases in fiber number. Indeed, a recent review of seventeen investigations into skeletal muscle growth concluded that “in several animal species certain forms of mechanical overload increases muscle fiber number” (Kelley, 1996). Notably, studies reporting an increase in the number of muscle fibers used avian or cat muscles, both species where muscles with intrafascicularly terminating fibers predominate (Gaunt and Gans, 1990; Scott et al., 1992; Brown et al., 1998). Other reports have commented on the technical difficulties of discerning between increases in fiber size or number (Taylor and Wilkinson, 1986; Timson, 1990; Tamaki and Uchiyama, 1995). It is admittedly difficult to analyze the number of fibers in pinnate muscles, such as soleus, where muscle fibers do not all lie in the plane of sectioning, or in intrafascicularly terminating muscles where sections through the mid-belly will miss short muscle fibers. The problem is compounded when larger muscles are used with many endplate bands and many intrafascicularly terminating fibers in series. Moreover, counting muscle fiber number by acid digestion is laborious in large muscles, and confounded by splitting, overlapping, and tapering fibers. It remains to be determined whether the growth of muscles with intrafascicularly terminating fibers in larger mammals arises from new fiber formation as assumed previously, or from elongation of existing fibers as documented here.
Distinguishing features of muscle fiber hypertrophy
The elongation of intrafascicularly terminating, multiple innervated muscle fibers resembles hypertrophy of singly innervated fibers in several respects. The accumulation of neonatal myosin, a hallmark of muscle regeneration, is similar in both types of fibers. Another shared feature is the activation of GATA-2, a unique molecular marker of the hypertrophic response. The extranuclear accumulation of GATA-2 in hypertrophic muscles represents an unusual subcellular distribution for a transcription factor, and may reflect a different role for this protein in the later stages of skeletal muscle hypertrophy. However, other transcription factors, such as NFATs, with which GATA factors interact (Musarò et al., 1999), are also excluded from the nucleus under certain conditions (Rao et al., 1997). It remains to be seen whether the nuclear exclusion of GATA-2 in late stages of hypertrophy is related to its interaction with NFAT factors.
Another distinction of fiber elongation in gracilis anterior muscle is the maintenance of peripheral nuclei in the IGF-EX model, whereas the same stimulus induces centralized nuclei in hypertrophic gracilis posterior muscle. In elongating fibers, new sarcomeres are likely to be added in series at the fiber ends, where the structure of the myofibrils will be least disturbed, to retain efficient contractions. Nuclei are therefore more likely to become centralized at the ends of hypertrophying intrafascicularly terminating fibers outside of these mid-belly sections. Notably, nuclei are centralized at the ends of elongating embryonic intrafascicular fibers before they move to the periphery (Duxson and Sheard, 1995). This mode of hypertrophy represents a recapitulation of the mechanism of fiber growth in the embryo and may therefore serve as a useful model for embryonic muscle growth.
Mouse as a model for other animals
Can the modes of hypertrophy documented in the mouse be extended to other species? The muscles of many mammals, with the exception of primates, are multiply innervated when their fascicle length exceeds 3.5 cm (Paul, 2001). Some investigators have commented on the existence of these intrafascicularly terminating fibers in other animals (Swatland and Cassens, 1972; Swatland, 1976) and have suggested a similar mode of hypertrophy to that proposed in the present study. It is important to stress the physical limitations on multiply innervated muscles in larger species, however. The intrafascicularly terminating fibers in mouse gracilis anterior muscle have two endplate bands and two sets of overlapping intrafascicularly terminating fibers, all of which have at least one termination in the tendon matrix. Three bands of endplates and three sets of intrafascicularly terminating fibers exist in muscles such as the guinea pig caudofemoralis. Those fibers centered around the outer endplate bands still have one tendinous insertion, but the fibers originating at the center endplate band have two intrafascicular terminations (unpublished data). It is not clear how such fibers without a tendinous insertion respond to hypertrophic signals, but logic would suggest that elongation would also occur at the ends to retain the source of innervation at the center of the fiber. Thus, multiply innervated mouse muscles, such as the gracilis anterior, are likely to recapitulate the mode of hypertrophy found in large animals such as cattle, sheep, horses, dogs, cats, and rabbits, rather than that found in primates. It remains to be determined whether the modes of myofiber atrophy are equally divergent in singly and multiply innervated muscles, which may limit extrapolation of data from large animal models to human myopathies.
Materials and methods
Animals
Transgenic FVB mice carrying a rat mIGF-1 cDNA driven by skeletal muscle–specific regulatory elements from the rat MLC-1/3 locus have been described previously (Musarò et al., 2001). Expression of the MLC/mIGF-1 transgene in these mice is restricted to skeletal muscle and elevated in muscles enriched in fast fibers, such as gracilis anterior and posterior muscles. Transgene expression is reduced in slow muscles such as the soleus, where the MLC regulatory cassette is expressed at very low levels. Five three-month-old mice in each of the four categories were used: WT, MLC/mIGF-1, WT-EX, and exercised MLC/mIGF-1 (IGF-EX).
Exercise
WT and transgenic mice were exercised to investigate skeletal muscle hypertrophy (n = 5 in each group) (Allen et al., 2001). Two male mice were exercised simultaneously in a S.A.M. Palace Kit™ cage fitted with an additional wheel. The distance and time spent exercising on the wheels was measured with a Sigma Sport BC 600 bicycle computer and a magnet glued to the periphery of each wheel. Exercised mice had free access to the wheels, food, and water. The weight of WT-EX mice transiently dropped 2–3 g several days after the beginning of exercise (unpublished data), presumably due to loss of fat before muscle hypertrophy. After 4 wk, exercised or sedentary age- and sex-matched controls were killed and muscles were processed for histology, immunohistochemistry, RNA expression, and protein analysis by Western blotting and immunohistochemistry.
AChE staining
The patterns of mouse gracilis muscle innervation were investigated as described previously (Schwarzacher, 1957). Gracilis anterior and gracilis posterior mouse muscles were dissected and stored in 1% paraformaldehyde solution. Muscles were washed three times in PBS and permeabilized by soaking in 0.2% Triton/PBS solution followed by three washes in PBS. Neuromuscular and myo–myonal junctions were stained by incubating in AChE staining solution for up to 1 h (Karnovsky and Roots, 1964), rinsed in PBS, and photographed with a Leica Wild M10 microscope. Longitudinal 8-μm cryosections of fixed gracilis anterior muscle were also stained for AChE and neuromuscular and myo–myonal junctions were photographed using a Nikon Optiphot-2 microscope.
Muscle fiber number and average area
Skeletal muscle hypertrophy was investigated in gracilis posterior muscle (no intrafascicularly terminating fibers), in gracilis anterior muscle (containing intrafascicularly terminating fibers), and soleus muscle from WT, IGF, WT-EX, and IGF-EX mice. Five muscles of each type were embedded in TBS tissue freezing medium and snap frozen in liquid nitrogen–cooled isopentane. Transverse 8-μm sections were cut through the middle of the muscle. Sections were stained with the Fast Green modification of the Van Gieson stain (Clark, 1981), resulting in green muscle fibers and black nuclei. The sections were photographed on a Nikon Optiphot-2 microscope and all muscle fibers counted in cross-section. The total area of the muscle cross-section was calculated by weighing a paper cut-out of the silhouette of the muscle and adjusting the area according to the known weight of 1 mm2 paper. The average area of individual muscle fibers was then calculated by dividing the muscle area by the number of fibers.
The total number of muscle fibers with centralized nuclei was counted in these cross-sections of each muscle.
Fiber type analysis
Myosin heavy chain isoform changes were investigated in gracilis anterior and gracilis posterior muscles in WT, IGF, WT-EX, and IGF-EX mice. Transverse 8-μm sections were cut through the center of each muscle and blocked in 10% goat serum for 15 min. The sections were incubated with either anti-MyHC I antibody (American Type Culture Collection A4.951; 1:10 dilution in PBS/1% BSA) or anti-MyHC IIB antibody (American Type Culture Collection BF-F3; 1:10 dilution in PBS/1% BSA), washed three times in PBS, and incubated in secondary antibody (Jackson ImmunoResearch Laboratory; peroxidase-conjugated goat anti–mouse IgG 115-035-164, or peroxidase-conjugated goat anti–mouse IgM 115-035-075, both 1:500 in PBS/1% BSA). The peroxidase was visualized with Sigma-Aldrich FAST 3,3′-diaminobenzidine tablets, D-4168. The fibers with each phenotype were counted and the number of MyHC IIA/IIX calculated by subtraction from the total number of muscle fibers.
Acid digestion of muscle fibers
The total number of fibers was counted in three WT and IGF-1 gracilis anterior muscles by acid digestion. Muscles were dissected, fixed for 1 h in 4% paraformaldehyde and stored in 80% glycerol for at least 1 wk. Muscles were then briefly rinsed in PBS and placed in 8 N HCl at 60°C for up to 1 h until individual fibers began to separate. The HCl was removed and the muscles placed in PBS. The total number of muscle fibers could then be counted by peeling off individual fibers under a dissecting microscope.
RNA preparation and Northern blot
Total RNA was obtained from thigh muscles of exercised and unexercised WT and MLC/mIgf-1 transgenic muscles by RNA-TRIZOL extraction (GIBCO-BRL). Total RNA (2 μg) was fractionated by electrophoresis on 1.3% agarose gels, transferred to GeneScreen Plus Membrane overnight, and hybridized with full-length cDNA probes, including Exon-1 (muscle specific localized form) of the rat IGF-1 gene. 25 ng of probe were labeled with P32 isotope using the Megaprime DNA labeling system RPN1604 from Amersham Pharmacia Biotech.
Protein analysis by Western blot
Protein was extracted from gracilis anterior and gracilis posterior muscles from WT, IGF, WT-EX, and IGF-EX muscles. Muscles were snap frozen directly in liquid nitrogen, macerated, and placed in RIPA buffer (25 mM Tris, pH 8.2, 50 mM NaCl, 0.5% sodium deoxycholate, 0.1% SDS, 0.1% sodium azide, 0.5% NP-40) with 5% protease inhibitor cocktail (Sigma-Aldrich; P 2714). 30 μg of total protein was added to an equal volume of 2× reducing SDS sample buffer, boiled, and separated on a 9% SDS-PAGE gel. Proteins were transferred to Immobilon PVDF membrane using the Bio-Rad Laboratories semidry transfer system at 100 mA for 1 h. The membrane was blocked in MTTBS (TBS, 0.1% Triton, 5% nonfat milk powder) for 1 h, followed by incubation in the primary antibody (mouse monoclonal anti–GATA-2, Santa Cruz Biotechnology, Inc. CG2-96, dilution 1:500 in MTTBS). The membrane was washed three times in MTTBS and incubated in secondary antibody (Jackson ImmunoResearch Laboratory; peroxidase-conjugated goat anti–mouse IgG 115-035-164, dilution 1:10,000 in MTTBS). The HRP-conjugated protein was detected using chemiluminescence according to the manufacturer's protocol (Renaissance; NEN Life Science Products) and visualized on Kodak Biomax ML film.
Protein analysis
Expression patterns of GATA-2, neonatal myosin, and proliferating cell nuclear antigen (PCNA) were investigated on longitudinal 8-μm frozen sections of WT, IGF, WT-EX, and IGF-EX gracilis anterior and posterior muscles with immunohistochemistry. The procedure was identical to that described earlier. Primary antibodies used were anti-GATA-2 (Santa Cruz Biotechnology, Inc. CG2-96, dilution 1:100) in PBS/BSA (PBS with 1% bovine serum albumin), antineonatal myosin (gift from Schiaffino, dilution 1:100), and PCNA (Santa Cruz Biotechnology, Inc. PC10, dilution 1:100). The antibodies were detected with anti–mouse IgG-Cy3, Amersham Pharmacia Biotech PA43002 used at 1:1,000 dilution resulting in red fluorescence. Sections were costained with Hoechst 33258 dye (blue fluorescence) and the acetylthiocholinesterase method described earlier. For clearer presentation, the blue filter was processed into a green image using Adobe Photoshop® 5.
Acknowledgments
We thank A. Musarò, F. Depreux, C. Neville, E. Slonimsky, S. Zaltsman, and J. Downer for helpful advice and discussion, and Leslie Leinwand and Michael McGrew for sharing unpublished data.
This work was supported by grants to N. Rosenthal from the Muscular Dystrophy Association.
Footnotes
Abbreviations used in this paper: AChE, acetylcholinesterase; EDL, extensor digitorum longus; EX, exercised; IGF, insulin-like growth factor; MLC, myosin light chain; WT, wild-type.
References
- Adams, D., and B. MacKay. 1961. The distribution of motor end-plates in mammalian muscles. Bibl. Anat. 2:153–154. [Google Scholar]
- Allen, D.L., B.C. Harrison, A. Maass, M.L. Bell, W.C. Byrnes, and L.A. Leinwand. 2001. Cardiac and skeletal muscle adaptations to voluntary wheel running in the mouse. J. Appl. Physiol. 90:1900–1908. [DOI] [PubMed] [Google Scholar]
- Barton-Davis, E.R., D.I. Shoturma, A. Musaro, N. Rosenthal, and H.L. Sweeney. 1998. Viral mediated expression of insulin-like growth factor I blocks the aging-related loss of skeletal muscle function. Proc. Natl. Acad. Sci. USA. 95:15603–15607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown, I.E., T. Satoda, F.J. Richmond, and G.E. Loeb. 1998. Feline caudofemoralis muscle. Muscle fibre properties, architecture, and motor innervation. Exp. Brain Res. 121:76–91. [DOI] [PubMed] [Google Scholar]
- Clark, G. 1981. Methods for connective tissue. Staining Procedures. G. Clark, editor. Williams & Wilkins, Baltimore. 512.
- Coers, C., and J. Durand. 1957. La repartition des appareils cholinesteasiques en cupule dans divers muscles stries. Archiev. Biologie. 68:209–215. [PubMed] [Google Scholar]
- Cooper, S. 1925. The relation of active to inactive fibres in functional contraction of muscle. J. Physiol. (Lond.). 67:1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duxson, M.J., and P.W. Sheard. 1995. Formation of new myotubes occurs exclusively at the multiple innervation zones of an embryonic large muscle. Dev. Dyn. 204:391–405. [DOI] [PubMed] [Google Scholar]
- Gans, C., G.E. Loeb, and F.D. Vree. 1989. Architecture and consequent physiological properties of the semitendinosus muscle in domestic goats. J. Morphol. 199:287–297. [DOI] [PubMed] [Google Scholar]
- Gaunt, A.S., and C. Gans. 1990. Architecture of chicken muscles: short-fibre patterns and their ontogeny. Proc. R. Soc. Lond. B Biol. Sci. 240:351–362. [DOI] [PubMed] [Google Scholar]
- Gollnick, P.D., B.F. Timson, R.L. Moore, and M. Riedy. 1981. Muscular enlargement and number of fibers in skeletal muscles of rats. J. Appl. Physiol. 50:936–943. [DOI] [PubMed] [Google Scholar]
- Karnovsky, M.J., and L. Roots. 1964. A “direct coloring” thiocholine method for cholinesterases. J. Histochem. Cytochem. 12:219–221. [DOI] [PubMed] [Google Scholar]
- Kelley, G. 1996. Mechanical overload and skeletal muscle fiber hyperplasia: a meta-analysis. J. Appl. Physiol. 81:1584–1588. [DOI] [PubMed] [Google Scholar]
- Loeb, G.E., C.A. Pratt, C.M. Chanaud, and F.J. Richmond. 1987. Distribution and innervation of short, interdigitated muscle fibers in parallel-fibered muscles of the cat hindlimb. J. Morphol. 191:1–15. [DOI] [PubMed] [Google Scholar]
- Musarò, A., K.J. McCullagh, F.J. Naya, E.N. Olson, and N. Rosenthal. 1999. IGF-1 induces skeletal myocyte hypertrophy through calcineurin in association with GATA-2 and NF-ATc1. Nature. 400:581–585. [DOI] [PubMed] [Google Scholar]
- Musarò, A., K. McCullagh, A. Paul, L. Houghton, G. Dobrowolny, M. Molinaro, E.R. Barton, E.L. Sweeney, and N. Rosenthal. 2001. Localized IGF-I transgene expression sustains hypertrophy and regeneration in senescent skeletal muscle. Nat. Genet. 27:195–200. [DOI] [PubMed] [Google Scholar]
- Paul, A.C. 2001. Muscle length affects the architecture and pattern of innervation differently in leg muscles of mouse, guinea pig, and rabbit compared to those of human and monkey muscles. Anat. Rec. 262:301–309. [DOI] [PubMed] [Google Scholar]
- Purslow, P.P., and J.A. Trotter. 1994. The morphology and mechanical properties of endomysium in series-fibred muscles: variations with muscle length. J. Muscle Res. Cell Motil. 15:299–308. [DOI] [PubMed] [Google Scholar]
- Rao, A., C. Luo, and P.G. Hogan. 1997. Transcription factors of the NFAT family: regulation and function. Annu. Rev. Immunol. 15:707–747. [DOI] [PubMed] [Google Scholar]
- Ryan, J.M., M.A. Cobb, and J.W. Hermanson. 1992. Elbow extensor muscles of the horse: postural and dynamic implications. Acta Anat. (Basel). 144:71–79. [DOI] [PubMed] [Google Scholar]
- Schwarzacher, H.G. 1957. Zur lage der motorischen endplatten in den skeletmuskeln. Acta Anat. (Basel). 30:758–774. [PubMed] [Google Scholar]
- Schwarzacher, H.G. 1959. Uber die lange und anordnung der muskelfasern in menschlichen skeletmuskeln. Acta Anat. (Basel). 37:217–231. 14444083 [Google Scholar]
- Scott, S.H., D.B. Thomson, F.J. Richmond, and G.E. Loeb. 1992. Neuromuscular organization of feline anterior sartorius: II. Intramuscular length changes and complex length-tension relationships during stimulation of individual nerve branches. J. Morphol. 213:171–183. [DOI] [PubMed] [Google Scholar]
- Swatland, H.J. 1976. Effect of growth and plane of nutrition on apparent muscle fiber numbers in the pig. Growth. 40:285–292. [PubMed] [Google Scholar]
- Swatland, H.J., and R.G. Cassens. 1972. Muscle growth: the problem of muscle fibers with an intrafascicular termination. J. Anim. Sci. 35:336–344. [DOI] [PubMed] [Google Scholar]
- Tamaki, T., and S. Uchiyama. 1995. Absolute and relative growth of rat skeletal muscle. Physiol. Behav. 57:913–919. [DOI] [PubMed] [Google Scholar]
- Taylor, N.A., and J.G. Wilkinson. 1986. Exercise-induced skeletal muscle growth. Hypertrophy or hyperplasia? Sports Med. 3:190–200. [DOI] [PubMed] [Google Scholar]
- Timson, B.F. 1990. Evaluation of animal models for the study of exercise-induced muscle enlargement. J. Appl. Physiol. 69:1935–1945. [DOI] [PubMed] [Google Scholar]
- Timson, B.F., and G.A. Dudenhoeffer. 1990. Skeletal muscle fibre number in the rat from youth to adulthood. J. Anat. 173:33–36. [PMC free article] [PubMed] [Google Scholar]
- Walro, J.M., J. Kucera, and R. Narvy. 1991. Non-neural and neural expression of myosin heavy chains by regenerated intrafusal fibers of rats. Neurosci. Lett. 122:213–217. [DOI] [PubMed] [Google Scholar]
- Young, M., A.C. Paul, J. Rodda, M.J. Duxson, and P.W. Sheard. 2000. An examination of intrafascicular muscle fibres terminations: implications for tension delivery in series-fibred muscles. J. Morphol. 245:130–145. [DOI] [PubMed] [Google Scholar]