

Abstract
We report the identification of βIV spectrin, a novel spectrin isolated as an interactor of the receptor tyrosine phosphatase-like protein ICA512. The βIV spectrin gene is located on human and mouse chromosomes 19q13.13 and 7b2, respectively. Alternative splicing of βIV spectrin generates at least four distinct isoforms, numbered βIVΣ1–βIVΣ4 spectrin. The longest isoform (βIVΣ1 spectrin) includes an actin-binding domain, followed by 17 spectrin repeats, a specific domain in which the amino acid sequence ERQES is repeated four times, several putative SH3-binding sites and a pleckstrin homology domain. βIVΣ2 and βIVΣ3 spectrin encompass the NH2- and COOH-terminal halves of βIVΣ1 spectrin, respectively, while βIVΣ4 spectrin lacks the ERQES and the pleckstrin homology domain. Northern blots revealed an abundant expression of βIV spectrin transcripts in brain and pancreatic islets. By immunoblotting, βIVΣ1 spectrin is recognized as a protein of 250 kD. Anti–βIV spectrin antibodies also react with two additional isoforms of 160 and 140 kD. These isoforms differ from βIVΣ1 spectrin in terms of their distribution on subcellular fractionation, detergent extractability, and phosphorylation. In islets, the immunoreactivity for βIV spectrin is more prominent in α than in β cells. In brain, βIV spectrin is enriched in myelinated neurons, where it colocalizes with ankyrinG 480/270-kD at axon initial segments and nodes of Ranvier. Likewise, βIV spectrin is concentrated at the nodes of Ranvier in the rat sciatic nerve. In the rat hippocampus, βIVΣ1 spectrin is detectable from embryonic day 19, concomitantly with the appearance of immunoreactivity at the initial segments. Thus, we suggest that βIVΣ1 spectrin interacts with ankyrinG 480/270-kD and participates in the clustering of voltage-gated Na+ channels and cell-adhesion molecules at initial segments and nodes of Ranvier.
Keywords: chromosome 19, diabetes, neuropathies, secretion, signal transduction
Introduction
The spectrins were originally identified as rod-shaped molecules that are part of the lattice-like cytoskeleton underneath the erythrocyte membrane (Marchesi and Steers 1968; Byers and Branton 1985; Bennett and Gilligan 1993). This subplasmalemmal meshwork is required to maintain the shape and lipid asymmetry of the plasma membrane. The critical role of these proteins is best revealed by the evidence that several human genetic syndromes, such as elliptocytosis and spherocytosis, are caused by mutant spectrin alleles (for review, see Marchesi 1988; Gallagher and Forget 1998). The spectrins have also been proposed to participate in the trafficking of organelles along the secretory pathways (Beck and Nelson 1998; De Matteis and Morrow 1998), and to play a role in regulated secretion (Goodman et al. 1995; Sikorski et al. 1991, Sikorski et al. 2000).
The spectrin molecule is a tetramer that consists of two α and two β subunits. A tight association between the NH2-terminal domain of an α subunit and the COOH-terminal domain of a β subunit forms a heterodimer, which assembles into a tetramer through a head-to-head association. The most distinctive feature of both subunits is the presence of multiple repeats, each consisting of ∼106 residues that assemble to form a three-helix bundle (Speicher and Marchesi 1984; Yan et al. 1993). α spectrins include 20 spectrin repeats, two of which are less conserved, an SH3 motif between spectrin repeats 9 and 11, and a COOH-terminal domain with two EF-hand motifs. β spectrins, on the other hand, contain 17 spectrin repeats in between an actin-binding domain at the NH2 terminus and a pleckstrin homology (PH) domain towards the COOH terminus.
Until recently, four genes encoding two α spectrins (αI and αII) and two β spectrins (βI and βII) were known in mammals. αIΣ1 and βIΣ1 spectrin are primarily expressed in erythrocytes, whereas αII Σ1 and βIIΣ1 spectrin have a wide tissue distribution and are particularly abundant in the brain (Goodman et al. 1981; Burridge et al. 1982; Glenney et al. 1982; Ma et al. 1993). An alternative spliced isoform of βI spectrin, termed βIΣ2 spectrin is highly expressed in muscle and brain (Riederer et al. 1986; Zagon et al. 1986; Winkelmann et al. 1990b; Malchiodi-Albedi et al. 1993). Similarly, an alternative spliced form has been found for βII spectrin (Hayes et al. 2000). The number of β spectrins has recently expanded with the discovery of βIII (Ohara et al. 1998) and βV (Stabach and Morrow 2000) spectrin. βIII spectrin, which is most abundant in brain and kidney, is concentrated at the Golgi apparatus (Stankewich et al. 1998), and may therefore represent one of the Golgi spectrins previously reported (Beck et al. 1994; for review, see Beck and Nelson 1998; De Matteis and Morrow 1998). βV spectrin is an exception among mammalian spectrins, as it includes 30 repeats, similar to the Drosophila β heavy (βH) spectrin (Dubreuil et al. 1990). This large spectrin is most prominent in the outer segments of the retina and in gastric epithelial cells, while low levels are found in many tissues.
Besides with α spectrins, β spectrins interact directly with many additional proteins, including actin, protein 4.1, and ankyrins (Bennett and Gilligan 1993). Through the association with protein 4.1, βI spectrin interacts with the membrane proteins glycophorin C and the Cl−/HCO− 3 exchanger (for review, see Conboy 1993). The binding to ankyrins provides β spectrins with an additional linkage to integral membrane proteins, such as the voltage-gated Na+ channel, CD44, and the Na+/K+ ATPase (Bennett and Gilligan 1993). Finally, β spectrins can directly associate with membrane proteins via a domain located at the end of spectrin repeat 1 and with phosphoinositides through the PH domain (Davis and Bennett 1994; Lombardo et al. 1994; Godi et al. 1998; Yao et al. 1999). Thus, β spectrins are essential in the generation of organized cytoplasmic and membrane microdomains.
Here we report the cloning and characterization of a novel human spectrin, termed βIV spectrin, which was isolated in a two-hybrid screening in yeast as an interactor of the autoantigen of type I diabetes ICA512 (Rabin et al. 1992, Rabin et al. 1994; Lan et al. 1994). ICA512 (also known as IA-2, PTP-35, PTPLP, and PTPN) is a receptor tyrosine phosphatase–like protein of unknown function, which is associated with the secretory granules of neurons and peptide-secreting endocrine cells (Solimena et al. 1996; Lee et al. 1998; Solimena 1998; Hermel et al. 1999). The association of ICA512 with βIV spectrin is consistent with previous studies suggesting that ICA512 interacts with the cytoskeleton (Hermel et al. 1999; Ort et al. 2000). Our data indicate that βIV spectrin is most abundant in brain and pancreatic islet cells and undergoes extensive alternative splicing. In brain, βIV spectrin is colocalized with ankyrinG 480/270-kD at initial segments and nodes of Ranvier of myelinated neurons, the axonal compartments responsible for the generation and regeneration of action potentials. Thus, we propose that βIV spectrin, in concert with ankyrinG 480/270, is required for the anchoring of voltage-gated Na+ channels and cell adhesion molecules to the actin cytoskeleton and may play an important role in the physiology of nerve conduction.
Materials and Methods
Antibodies
βIV spectrin antisera were raised against two synthetic peptides corresponding to amino acids 2237–2256 (specific domain antiserum, SD) and 2542–2559 (COOH-terminal domain antiserum, CT) of human βIV spectrin, respectively. Both peptides were coupled to keyhole limpet hemocyanin through an added cysteine at their NH2 terminus and were injected into rabbits to generate polyclonal antisera. Both antisera were affinity purified on their corresponding peptides that had been covalently linked to ECH-Sepharose beads (Amersham Pharmacia Biotech). Fractions of the antisera were eluted with 0.2 M glycine (pH 3.0), collected and dialyzed with Tris buffer, pH 7.5. The specificity of these antisera and affinity purified antibodies was tested by Western blot on Triton X-100 extracts from COS cells expressing a partial fragment of βIV spectrin (residues 2068–2559), termed B8.
The following antibodies were from commercial sources: rabbit anti–LexA antiserum and mouse anti–V5 antibody (Invitrogen), mouse monoclonal antibody against ankyrin G 480/270-kD (Zymed Laboratories), mouse monoclonal antibody against glucagon (Sigma-Aldrich), mouse monoclonal antibody against myelin basic protein (Sternberger Monoclonal Antibodies), anti–rabbit and anti–mouse goat IgG antibodies conjugated to Alexa 488 or Alexa 568 (Molecular Probes, Inc.). A mouse monoclonal antibody against hemagglutinin (HA) was a kind gift of R. Collins (Cornell University, Ithaca, NY). Affinity-purified antibodies against the cytoplasmic or the ectodomain of ICA512 have been described previously (Solimena et al. 1996; Hermel et al. 1999).
Two-Hybrid Screening in Yeast
A yeast two-hybrid screening was executed according to the protocol of Vojtek et al. 1993. Briefly, the cDNAs encoding the cytoplasmic domains of human ICA512 (amino acids 601–979, ICA512cyt), human PHOGRIN (amino acids 640–1015, PHOGRINcyt) and ICA512 mutant allele (ICA512cyt AD/DA) were amplified by PCR and subcloned into the pLexA vector to create fusion proteins with the LexA DNA binding domain. The ICA512cyt AD/DA mutant was constructed using site-directed mutagenesis on human ICA512 in pRC/RSV (Invitrogen) with the QuickChange mutagenesis kit (Stratagene) according to the manufacturer's instructions (A877D, 5′ primer: CCTCAGCTGGCCGGACGAGGGCACACCGG, 3′ primer: CCGGTGTGCCCTCGTCCGGCCAGCTGAGG; D911A, 5′ primer: CGTGCACTGCTCGGCCGGTGCGGGGAG; 3′ primer: CTCCCCGCACCGGCCGAGCAGTGCAC). The cytoplasmic domain of human PHOGRIN was amplified by PCR from a human adult brain cDNA library in pACT2 (CLONTECH Laboratories, Inc.). Automated sequencing of all constructs was carried out by the Keck Biotechnology Resource Laboratory at Yale University School of Medicine.
pLexA-ICA512cyt, pLexA-ICA512cyt AD/DA, and pLexA-PHOGRINcyt were independently transformed into the yeast strain L40 [partial genotype MATa trp1 leu2 his3 LYS2::(lexAop)-HIS3 URA3::(lexAop)-lacZ GAL4] and AMR70 [partial genotype: MATα his3 lys2 trp1 leu2 URA3::(lexAop)-lacZ GAL4]. Expression of the LexA-fusion proteins (baits) in yeast cells was verified by Western blotting using the anti-LexA antiserum (1:1,000). L40 cells expressing pLexA-ICA512cyt were cotransformed with 500 μg of a human adult brain cDNA library in pACT2 (Clontech) that included an in-frame HA tag (Clontech). Double transformants were selected for His+ and LacZ+ phenotypes. After purging of the bait plasmid, L40 His+ LacZ+ cells were mated separately with AMR70 cells transformed with the following baits: LexA-ICA512cyt, LexA-ICA512cyt AD/DA, LexA-PHOGRINcyt, LexA-lamin, and LexA-MSS4. Since lamin and MSS4 are proteins unrelated to ICA512, they were not expected to share interactors with ICA512. Accordingly, only those pACT2 plasmids that reconstituted His+ auxotrophy with LexA-ICA512 cyt, but not LexA-lamin or LexA-MSS4, in the mating assay were isolated and their cDNA inserts characterized by sequence analysis. The stringency conditions of the mating assay were increased by adding 3-amino-1,2,4-triazole (3-AT) (Sigma-Aldrich) to the plate media at concentrations ranging from 0 to 10 mM. To assess expression of bait and prey proteins in the mating assay, diploid cells were lysed by incubation for 10 min in 0.2 N NaOH/0.5% β-mercaptoethanol on ice. Lysates were incubated with 10% trichloroacetic acid for 30 min on ice, and then spun for 10 min at 10,000 g at 4°C. After precipitation in 100% acetone, the proteins were resuspended in 2% SDS sample buffer (New England Biolabs, Inc.) to a final concentration of 200–400 μg/OD. Protein concentration in tissue extracts was determined using the BCA assay procedure (Pierce Chemical Co.). 100 μg protein from each lysate was run on 12% SDS-PAGE and blotted with anti-HA (1:10) and anti-LexA (1:1,000) antibodies followed by 125I-Protein A (105 cpm/ml) and autoradiography.
Cloning of βIV Spectrin
Cloning of the full length βIV spectrin cDNA from adult human brain was performed using standard molecular procedures (Sambrook et al. 1989). Briefly, a 325-bp fragment (6381–6705 bp in βIV spectrin) within the partial clone isolated in the two-hybrid screening was amplified by PCR (5′ primer: AGCCGGGAGCTGGGCAGCAGCGTG and 3′ primer: GGCGCACATACCCCACCCGAGTCC) and labeled with [α-32P-dCTP] (3,000 Ci/mmol; Amersham Pharmacia Biotech) using the Random Prime Labeling Kit (Amersham Pharmacia Biotech). This probe was used to screen 2 × 106 independent phage clones of a λgt11 human brain cDNA library (Clontech) on nitrocellulose filters. Three independent phages were isolated whose sequences partially overlapped that of the original clone. Six additional screenings, each of 2 × 106 phages from a λgt10 human cerebellum cDNA library (Clontech), yielded 20 independent phages. Contig alignment of their inserts led to the assembly of 7,779-bp cDNA. The remaining 1,010 bp at the 5′ end βIV spectrin were cloned by PCR on a human brain Marathon-Ready cDNA library (Clontech) according to the manufacturer protocol using the following reverse primer: 5′-CTCCCGAGGCACAAAGAGGCGACG.
A 444-bp cDNA fragment of βIV spectrin was amplified by PCR from mouse pancreatic islet cDNA (gift of Dr. R. Flavell, Yale University) using the following primers: (5′ primer: GGCTCAGAATAAGGAGTGGCTGGAGAAGAT; 3′ primer: AACGGCTCGAGCAGGCGCACCAGGCGC). Computer-assisted analysis of all cDNAs was performed using the following software programs: BLAST (Altschul et al. 1990), Lasergene (DNASTAR Inc.) and PFSCAN (ISREC Bioinformatics Groups). Intron-exon boundaries of the human βIV spectrin gene were established by comparing the cDNA sequence with the corresponding genomic sequence.
Chromosomal Localization
Human and mouse BAC clones encompassing the βIV spectrin gene were isolated at Genome Systems according to established protocols (Shizuya et al. 1992) using a fragment of the human βIV spectrin cDNA (nucleotides 4094–4502) as a probe. Automated sequence analysis of these clones confirmed the presence of exon–intron boundaries within the βIV spectrin gene. The locus of the βIV spectrin gene on human and mouse chromosomes was also established at Genome Systems by fluorescent in situ hybridization using human and mouse genomic probes (>10 kb) originating from the corresponding BAC clones (Stokke et al. 1995).
Expression of ICA512 and βIV Spectrin in COS Cells and Coimmunoprecipitations
A full-length human ICA512 construct in the mammalian expression vector pRC/RSV (Invitrogen) has been described previously (Hermel et al. 1999). A starting methionine was introduced by PCR at the 5′ end of the βIV spectrin partial clone B8. The resulting cDNA, including the NH2-terminus HA tag derived from the pACT2 cDNA library, was subcloned as a HindIII–XbaI insert into pRC/RSV. Both ICA512 and B8 constructs (5 μg) were transiently transfected into COS cells by the Ca2+-phosphate procedure, as described previously (Dirkx et al. 1995). 72 h after transfection, transfected or mock-transfected (no cDNA) COS cells were harvested in PBS, pelleted, and then solubilized for 90 min on ice with homogenization buffer (158 mM NaCl, 10 mM HEPES, pH 7.4, 1mM PMSF, 10 mM benzamidine, 1 μg/ml aprotinin, 1 μg/ml leupeptin, 1 μg/ml pepstatin A, 1 μg/ml antipain) including 2% Triton X-100. One confluent dish (∼1.5 × 107 cells) was used per immunoprecipitation. Insoluble material was removed by centrifugation at 15,000 g for 10 min at 4°C. Triton X-100 soluble material was precleared with 125 μl of 50% slurry protein G sepharose beads (Amersham Pharmacia Biotech). Next, extracts were incubated overnight at 4°C with 10 μl of a monoclonal antibody against the cytoplasmic domain of ICA512 or 50 μl of the βIV-SD antibody. 100 μl of a 50% slurry protein G sepharose beads was added to each sample and incubated for 1 h on ice. Beads were pelleted by centrifugation, washed three times with 2% Triton X-100 in homogenization buffer without protease inhibitors, and then resuspended in 100 μl of 1× SDS sample buffer. 20 μl of each immunoprecipitate was separated on a 8% SDS-polyacrylamide gel, transferred on nitrocellulose, and immunoblotted with the following antibodies: anti–ICA512 ectodomain (1:1,000), anti–HA (1:10), and anti–βIV-SD (1:250). Immunoreactivity was detected with alkaline phosphatase–conjugated goat anti–mouse (Sigma-Aldrich) or anti–rabbit IgG (Roche) (1:5,000).
Northern Blotting
A nitrocellulose filter with ∼2 μg polyA+ RNA from various human tissues (Multi Tissue Northern blot; Clontech) was hybridized with an α-32P-dCTP–labeled probe including nucleotides 6202–6526 within the predicted open reading frame of βIV spectrin. The hybridization was performed according to the manufacturer's instructions. After x-ray exposure for 5 d, the blot was stripped by immersion in boiling water containing 0.5% SDS and rehybridized with a probe specific for human β-actin provided by the manufacturer.
10 μg polyA+ RNA was purified with an Oligotex Direct mRNA kit (Qiagen) from ∼4 × 104 human pancreatic islets kindly provided by the JDF Human Islet Distribution Centers at the University of Miami (Miami, FL), and Washington University (St. Louis, MO). ∼2 μg polyA+ RNA was separated on a 1% agarose gel containing 5% formaldehyde, blotted overnight on nitrocellulose by capillarity and hybridized with a probe including nucleotides 1805–2356 in the open reading frame of human βIV spectrin. The autoradiography was developed after a 5-d exposure.
Transfection of β-Spectrin Constructs in CHO Cells
The cDNAs encoding the carboxy termini of human βIΣ2, βII, βIII, and βIVΣ1 spectrin were amplified by PCR using the following templates: a partial clone of human βIΣ2 spectrin cDNA (Winkelmann et al. 1990b; gift of Dr. Bertrand Forget, Yale University) (5′ primer: ACCATGGAAGAAGAGGGAACGTGG, 3′ primer: CTTCTTTTTGGGGAAGAAGCTGAA); the pACT2 human brain cDNA library (Clontech) for βIIΣ1 spectrin (5′ primer: GCACCATGCGGATGGCAGAAACGGTGGAC, 3′ primer: TTTCTTTTTGCCAAAAAGGCTGAACCGCTT) and βIII spectrin (5′ primer: GCACCATGGGACAACAGAGACTTGAGCAC, 3′ primer: CTTGTTCTTCTTAAAGAAGCTGAA); our partial clone B8 for βIV spectrin (GCACCATGGTGCGGCCACGACCGGAGCGCCAGGAG, 3′primer: CTTCCTGCGCCCGCTGGCCCTGCGATCTCCGCCTTCCC). All cDNAs were subcloned upstream of the V5 epitope tag into pcDNA3.1/V5-His-TOPO/lacZ (Invitrogen) and transfected into CHO cells using the Lipofectin reagent (Life Technologies, Inc.) as described previously (Solimena et al. 1993).
Biochemical Procedures on Human Islets and Rat Brain Tissues
Approximately 2.5 × 104 purified human pancreatic islets were sonicated on ice in 200 μl homogenization buffer (HB; 10 mM HEPES, pH 7.4, 5 mM EDTA, 1 mM EGTA, 1 mM NaCl, 1 mM PMSF, 10 mM benzamidine, 1 μg/ml aprotinin, 1 μg/ml leupeptin, 1 μg/ml pepstatin A, 1 μg/ml antipain). The islet lysates were then solubilized in 2% SDS sample buffer to a final protein concentration of ∼2.5 mg/ml. SDS insoluble material was removed by centrifugation.
Brain tissues were collected from adult rats, 15- (E15) and 19- (E19) d-old rat embryos and 1- (P1) and 10- (P10) d-old newborn rats. Tissues were homogenized on ice in HB (1:10 wt/vol). Post-nuclear supernatant (PNS) was obtained by centrifugation at 1,000 g for 10 min and solubilized in 2% SDS sample buffer to a final concentration of ∼1.2 mg/ml. PNS proteins (40 μg/lane) were separated by 6% SDS-PAGE and immunoblotted with βIV-SD (1:500) or βIV-CT (1:500) antibodies, followed by peroxidase-conjugated goat anti–rabbit IgG (1:5,000; Sigma-Aldrich) and enhanced chemiluminescence (Amersham Pharmacia Biotech). As controls, PNS proteins from adult rat brain were blotted with the preimmune rabbit sera as well as with βIV spectrin antibodies that had been preincubated with 100 μg of the corresponding antigenic peptide.
High-speed pellet (HSP) and high-speed supernatant (HSS) from PNS of adult rat brain were obtained by centrifugation at 100,000 g for 1 h at 4°C. The HSS was collected, while the HSP was brought back to the original volume in HB containing 1% Nonidet P-40 (Boehringer) and 0.5% deoxycholic acid (Sigma-Aldrich). The HSP was solubilized for 2 h at 4°C and centrifuged at 100,000 g for 1 h. After the recovery of the soluble fraction, the HSP insoluble fraction was resuspended and sonicated in an equal volume of HB. PNS, HSS, and HSP soluble and insoluble fractions were dissolved in SDS sample buffer, separated by 6% SDS-PAGE, and blotted with the affinity purified βIV-CT and βIV-SD antibodies.
For alkaline phosphatase treatment, adult rat brain tissue was homogenized in HB without EDTA and EGTA. 300 μl of the resulting PNS (∼1.8 mg/ml) was incubated with 50 U of calf-intestine alkaline phosphatase (Roche) in phosphatase buffer for 1 h at 37°C. Alternatively, subcellular fractions of rat brain were prepared as described above, and then treated with alkaline phosphatase before SDS-PAGE and immunoblotting.
In Situ Hybridization
35S-labeled riboprobes were prepared with a transcription kit (Promega) using as a template a partial cDNA of βIV spectrin (bp's 6384–8792) that had been subcloned into pBluescript SK and KS vectors (Stratagene) as a HindIII-XbaI fragment. The antisense and control sense cRNAs were transcribed in the presence of 35S-UTP (Amersham Pharmacia Biotech) from the XbaI-linearized SK plasmid and the HindIII-linearized KS plasmid, respectively. In situ hybridization on cryosections of rat brain (10 μm) was performed as previously described (Dunbar et al. 1998). Dark-field images were acquired with a microscope (BX50; Olympus) connected to an Olympix CCD camera driven by IP-Lab Software 3.0 (Scanalytics).
Immunocytochemistry
Males Sprague-Dawley rats (150–175 g) were fixed by trans-cardiac perfusion with 1% paraformaldehyde in 120 mM sodium phosphate buffer. Tissues of interest were collected, fixed for an additional 3 h, and then infiltrated with 30% sucrose in PBS. Tissues from rats at days E15, E19, and P10 were collected, fixed in 1% paraformaldehyde for 3 h, and then infiltrated with 30% sucrose. Teased fibers from rat sciatic nerve were prepared and fixed with 1% paraformaldehyde as previously described (Fjell et al. 2000). Single and double immunolabeling on 12-μm cryostat tissue sections and transfected CHO cells was performed as previously described (De Camilli et al. 1983; Cameron et al. 1991). Primary antibodies were diluted as follows: anti–βIV-CT and anti–βIV-SD, 1:500; anti–ankyrin G 480/270, 1:25; anti–ICA512 ectodomain, 1:10; anti–glucagon, 1:200, anti–myelin basic protein, 1:200; anti–V5, 1:200. As controls, sections were incubated with the preimmune serum of the rabbit immunized with the βIV spectrin COOH-terminal peptide (1:50) and with anti–βIV-CT antibody that had been preincubated with the antigenic peptide. Confocal microscopy was performed using an MRC 1024 station (Bio-Rad Laboratories) attached to an Axiovert microscope (Carl Zeiss, Inc.).
Results
The COOH-terminal Region of a Novel β Spectrin Binds the Cytoplasmic Domain of ICA512
ICA512 belongs to the receptor protein tyrosine phosphatase (RPTP) family, but does not display phosphatase activity towards conventional PTP substrates, because its cytoplasmic PTP homology domain differs in two critical residues from that of active PTPs. Specifically, in ICA512, the catalytic aspartate of conventional PTPs is replaced by an alanine (A877, Fig. 1 A), while an aspartate (D911) rather than the obligatory alanine is present at position +2 from the catalytic cysteine of active PTPs. The replacement of A877 and D911 with aspartate and alanine, respectively, is sufficient to confer PTP activity to the corresponding ICA512 mutant (ICA512 AD/DA) (Magistrelli et al. 1996). To gain insight into ICA512's function, we searched for proteins interacting with its cytoplasmic domain by performing a two-hybrid screening in yeast.
Figure 1.
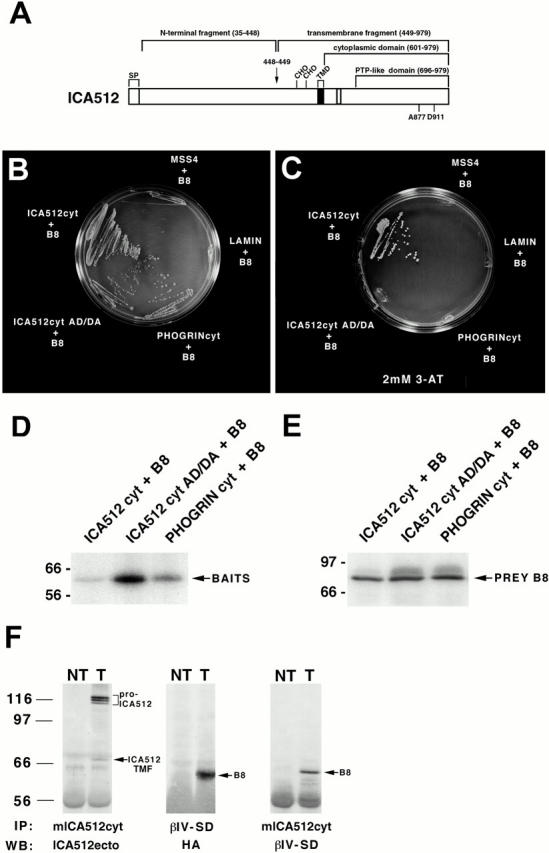
(A) Schematic representation of human ICA512. The protein includes 979 amino acids, including a signal peptide (SP), a single transmembrane domain (TMD), and a cytoplasmic domain with an inactive PTP domain. Cleavage at a consensus site for furin-like convertases between amino acids 448 and 449 (arrow) gives rise to an NH2-terminal fragment and a transmembrane fragment of 60–66 kD that includes the cytoplasmic domain. CHO, glycosylation sites. The residues in the PTP domain of ICA512 that distinguish it from active PTPs are indicated (A877, D911). (B–F) Interaction of the partial clone of βIV spectrin, B8, with ICA512cyt. (B and C) Yeast two-hybrid mating assay. L40 cells transformed with the prey plasmid pACTII-B8 were mated with AMR70 cells transformed with bait plasmids pLexA-ICA512cyt, pLexA-ICA512cyt AD/DA, pLexA-PHOGRINcyt, pLexA-lamin, or pLexA-MSS4 and grown on selective media in the absence (B) and presence (C) of 2 mM 3-AT. Cell survival reflects the interaction of the bait with the prey. (D and E) Expression levels of the different baits and B8 in L40-AMR70 mated cells as determined by Western blotting with an antiserum against LexA (D) and HA (E). (F) Coimmunoprecipitation of ICA512 and B8 from cotransfected COS cells. A mouse antibody directed against ICA512cyt or a rabbit antibody directed against the βIV spectrin specific domain (βIV-SD) were used for immunoprecipitation (IP) from Triton X-100 extracts of nontransfected (NT) and ICA512-B8 cotransfected (T) cells. Immunoprecipitates were Western blotted (WB) with affinity purified antibodies directed against the ectodomain of ICA512, βIV-SD, or a mouse antibody directed against HA. The arrows point to the immunoprecipitated ICA512 transmembrane fragment (ICA512 TMF) and B8, while a bracket indicates the position of pro-ICA512.
L40 yeast cells were cotransformed with LexA-ICA512cyt fused to the DNA binding protein LexA and a human brain cDNA library fused to the Gal4 activation domain in pACT2 as a source for preys. Screening of 107 L40 transformants led to the isolation of ∼540 colonies positive for the expression of both the reporter genes HIS3 and β-galactosidase. One of the preys that overcame histidine auxotrophy when expressed with LexA-ICA512cyt (Fig. 1 B) was a novel peptide of 493 amino acids whose last 149 residues were >40% similar to the COOH-terminal region of various β spectrins. This polypeptide, termed B8 (Fig. 2 A), also bound to a PTP active mutant of ICA512cyt (ICA512cyt AD/DA) and to the cytoplasmic domain of the ICA512-related protein, PHOGRIN (PHOGRINcyt) (Wasmeier and Hutton 1996). B8, on the other hand, did not interact with the unrelated control baits lamin and MSS4. When the two-hybrid assay was performed in more stringent conditions (i.e., in the presence of the HIS3 inhibitor 3-AT), B8 still associated with ICA512cyt, but not with ICA512cyt AD/DA or PHOGRINcyt (Fig. 1 C). Notably, cells expressed comparable amounts of B8 (Fig. 1 E), but less ICA512cyt than ICA512cyt AD/DA or PHOGRINcyt (Fig. 1 D). These data indicated that the viability of yeast cells expressing ICA512cyt and B8 in the presence of 3-AT was not accounted for by a higher expression of the bait and suggests that ICA512cyt has a higher affinity for B8 than ICA512cyt AD/DA or PHOGRINcyt. Thus, the replacement of the two residues that prevent ICA512 from acting as a conventional PTP was sufficient to weaken its interaction with B8.
Figure 2.




(A) Schematic diagram of the overlapping human βIV spectrin cDNAs forming the total contig of 8,789 bp. Additional clones that did not affect the contig have been left out of the alignment. (B) Graphical representation of the genomic organization of βIV spectrin. Black boxes represent the 36 exons encoding the full-length βIV spectrin. Exons from alternative spliced isoforms are boxed in grey. The length of the intervening sequences is still undetermined. Translation initiation and stop codons of the four alternative spliced variants are indicated. (C) Schematic representation of the domain structure of βIVΣ1 spectrin. CH, calponin-homology. (D) Primary amino acid sequence of βIVΣ1 spectrin. The four major domains of βIVΣ1 spectrin, numbered I–IV, are boxed. Domain I: the residues in the calponin-homology domain are in bold. Domain II: the arabic numbers on the left correspond to the spectrin repeats. Within this domain, the predicted A, B, and C helices are boxed separately and their conserved charged and aromatic residues are in bold. The glycine-rich region of 15 residues following the predicted A helix of spectrin repeat 4 is shown as an insert. Domain III: the four ERQES repeats in this specific domain are in bold. Domain IV: the residues in the pleckstrin homology domain are in bold. The proline-rich regions in spectrin repeat 15 and in the specific domain are underlined. (E) An alignment of the third ERQES repeat within the specific domain of βIV spectrin with sequences in the spectrin repeat 1 of βII and βIII spectrin. (F) Schematic representation of the different isoforms of βIV spectrin. (Top inset) The amino acids at the boundary between exons 17 and 18 in βIVΣ1 spectrin. (Middle inset, bold) The residues encoded by exon 17B in two distinct reading frames. The top sequence is in frame with the preceding exon 17 and includes a stop codon. The bottom sequence begins with a methionine and is in frame with the following exon 18. (Bottom inset) The amino acid sequence of exon 30B at the COOH terminus of βIVΣ4 spectrin. The roman numbers I–IV indicate the four major domains of βIV spectrin as defined in Fig. 2 D.
The ability of ICA512 to bind B8 was further investigated by coimmunoprecipitations from extracts of COS cells cotransfected with the cDNAs encoding for full-length ICA512 and HA-tagged B8 (Fig. 1 F). In these cells, pro-ICA512 accumulates as a triplet of ∼110 kD, while the ICA512 transmembrane fragment (ICA512 TMF) generated by the cleavage of the protein's ectodomain represents a minor species (Hermel et al. 1999). Immunoprecipitation with the βIV-SD antibody raised against a peptide within B8 followed by Western blotting with a monoclonal anti–HA antibody allowed the detection of a single protein of ∼60 kD in transfected COS cells, but not in mock-transfected COS cells. The size of this protein corresponds to the predicted size of HA-tagged B8. The same protein was also immunoprecipitated from cotransfected COS cells using a monoclonal antibody directed against the cytoplasmic domain of ICA512. These data confirmed the interaction between the two proteins revealed by the two-hybrid assay.
Cloning of Human βIV Spectrin
Screenings of human brain cDNA libraries by filter hybridization and PCR allowed the contig assembly of an 8,789-bp cDNA, including a 5′-untranslated region of 179 bp and a 3′-untranslated region of 933 bp with a polyadenylation signal and a polyA tail (Genbank No. AF082075) (Fig. 2 A). By fluorescence in situ hybridization, this new gene, termed βIV spectrin, was localized on human chromosome 19q13.13 and on the synthenic region on mouse chromosome 7b2 (not shown). Through BLAST analysis, we matched the cDNA of βIV spectrin to a genomic sequence from human chromosome 19 in the High Throughput Genomic Sequence database (Genbank No. AC021625). Comparison of this genomic sequence, which corresponds to a first draft of 34 unordered pieces, with the cDNA of βIV spectrin revealed that the latter is generated from 36 exons (Fig. 2 B). All exons, except for exon 17, are found in this genomic sequence and are flanked by canonical donor/acceptor intron sites. This genomic region still contains many gaps of unknown length, which may account for the apparent absence of the exon 17 (bp's 3819–4021) found in the cDNA sequences.
A high degree of similarity was found between portions of the βIV spectrin cDNA and another genomic sequence of 100 unordered pieces from chromosome 16 (Genbank No. AC009140.4). This genomic draft contains 18 sequences that are bounded by donor/acceptor intron sites and are very conserved with 18 exons of βIV spectrin (exons 1, 8–11, 13, 15–17, 19, 25, 26, 28, 29, 32, 34–36) (not shown). This genomic draft, however, did not appear to include the remaining exons of βIV spectrin. Our cDNA contig for βIV spectrin and the genomic sequence on chromosome 19 only differed in three base pairs, possibly because of polymorphism or sequencing artifacts. In contrast, six of the βIV spectrin-related exons on chromosome 16 contained many base substitutions, deletions, or insertions compared with the related exons in βIV spectrin. These data confirmed that our cDNA contig was entirely derived from the βIV spectrin gene on chromosome 19. They also suggested, on the other hand, that a gene closely related to βIV spectrin resides on chromosome 16. Cloning and characterization of this gene, provisionally termed βVI spectrin, is in progress.
The conceptual translation of the βIV spectrin cDNA corresponds to a protein of 2,559 amino acids with a predicted molecular weight of ∼288 kD and an isoelectric point of ∼5.9. The putative starting methionine of βIV spectrin is not in the context of a Kozak sequence, but it is preceded in the 5′-untranslated region by an in-frame stop codon. The domain structure of βIV spectrin closely resembles that of other β spectrins (Hu et al. 1992; Winkelmann et al. 1990a,Winkelmann et al. 1990b; Ma et al. 1993; Ohara et al. 1998), with the exception of βV spectrin (Stabach and Morrow 2000). It includes two calponin homology domains, followed by 16 complete spectrin repeats, a partial spectrin repeat (repeat 17), and a PH domain (Fig. 2C and Fig. d). Calponin homology domains are found at the NH2 terminus of many proteins, including β spectrins, and mediate the interaction with actin (Karinch et al. 1990; Frappier et al. 1992; Banuelos et al. 1998). Each of the first 16 spectrin repeats of βIV spectrin follows the criteria defined for the repeat 14 of Drosophila β spectrin (Yan et al. 1993; Pascual et al. 1997), and thus is predicted to assemble into three helices termed A, B, and C (Fig. 2 D). The repeat 15 of βIV spectrin, which in other β spectrins binds ankyrins (Kennedy et al. 1991), is 43.8, 46.7, and 31.4% identical to the corresponding repeats of βI, βII, and βIII spectrin, respectively. Unlike typical spectrin repeats, but similar to repeat 15 of other β spectrins, it does not include a tryptophan at position 17 in the helix A (Fig. 2 D). As in all other β spectrins, the repeat 17 of βIV spectrin lacks the segment corresponding to the C helix (Fig. 2 D). This is consistent with the possibility that the A and B helices of repeat 17 of βIV spectrin form a three-helix bundle with the lone C helix at the NH2 terminus of an α spectrin (Kennedy et al. 1994; Deng et al. 1995). The putative loop between the predicted helices A and B of repeat 4 contains a stretch of 15 residues that is not found in other β spectrins (Fig. 2 D). This insert is rich in glycines, consistent with a role as a helix breaker.
The region of 294 residues between the repeat 17 and the PH domain is specific for βIV spectrin as it shows no similarity with regions present in other β spectrins. This domain is rich in prolines (15% of the residues), is very basic (pI: 10.5), and contains a motif of five residues (ERQES) that is repeated four times within a stretch of 90 residues (residues 2220–2309, henceforth defined as ERQES domain). An identical motif was not found in any other protein in public databases. The ERQES domain is highly charged (55.5% charged residues) because of its high content in glutamate (23.3%), glutamine (8.8%), and arginine (26.6%). The presence of only two aspartates (2.2%) and the absence of asparagines and lysines, however, indicates that there is a strong bias toward specific residues in this region. The equal spacing of 13 residues between the ERQES repeats 1–3 further suggests the repetitive nature of this domain. Interestingly, the sequence corresponding to the third ERQES repeat and the four following residues (ERQESAEHE) is 77% identical to the sequence ERLEKAEHE within spectrin repeats 1 of human βII (residues 397–405) and βIII (residues 400–408) spectrin (Fig. 2 E). The motif ERLEKAEHE is part of a domain that associates βII spectrin with membranes, presumably via protein–protein interaction (Davis and Bennett 1994; Lombardo et al. 1994). Prediction of the secondary structure of βIV spectrin suggests that the ERQES domain is exposed at the protein's surface (not shown) and, as such, might also engage in protein–protein interaction.
βIV spectrin contains numerous putative SH3 binding sites (Fig. 2 D). Two of these proline-rich motifs (residues 1859–1864 and 2405–2411) are adjacent to putative membrane association domains. Specifically, the first of these motifs precedes the putative B helix in repeat 15, which may act as an ankyrin-binding site, while the second motif precedes the PH domain. The PH domain of βIV spectrin is very conserved with the corresponding domains in βIΣ2, βII, and βIII spectrins and is therefore likely to bind phosphoinositides, such as phosphatidylinositol 4,5 bisphosphate (Hyvonen et al. 1995).
βIV Spectrin Is Alternatively Spliced
Our data indicated that βIV spectrin undergoes alternative splicing. Specifically, one cDNA clone of ∼2.4 kb contained an insert of 88 bp between exon 17 and 18, which was not present in overlapping cDNA clones (Fig. 2 F). This insert was also found in the βIV spectrin gene, where it is flanked by canonic donor and acceptor splice sites at its 5′ and 3′ ends. Insertion of this exon, termed 17B (Fig. 2B and Fig. f), introduces a stop codon in the open reading frame of βIV spectrin, thereby causing its termination in spectrin repeat 9. The conceptual translation of this cDNA corresponds to a polypeptide of 1,302 amino acids with a predicted molecular weight of ∼145 kD. In a different reading frame, on the other hand, exon 17B encodes for a methionine that is preceded by a stop codon in exon 17 and is in frame with the remaining COOH-terminal portion of βIV spectrin (Fig. 2B and Fig. f). Recognition of this methionine as a novel initiation site for translation would generate an additional polypeptide of 1,307 amino acids with an expected molecular weight of ∼149 kD. It is conceivable that the inclusion of exon 17B gives rise to a bicistronic mRNA, whose translation produces two different βIV spectrin proteins. Based on the established nomenclature for spectrins (Winkelmann and Forget 1993), the originally cloned βIV spectrin has been termed βIVΣ1 spectrin, and hence we will refer to these two variants as βIVΣ2 and βIVΣ3 spectrin (Genbank No. AY004226).
In an additional clone, an insert of 315 bp (exon 30B) followed exon 30 (Fig. 2B and Fig. f). This insert encodes 42 amino acids and a stop codon, contains a polyadenylation site, and was found in the βIV spectrin gene, where it is flanked by canonic 5′ and 3′ splice sites. The isoform including this exon corresponds to a polypeptide of 2,149 amino acids (expected molecular weight: ∼242 kD), which contains the calponin-homology domain and all 17 spectrin repeats, but lacks the specific domain and the PH domain (Fig. 2 F). This isoform has been termed βIVΣ4 spectrin (Genbank No. AY004227). A search in publicly available databases revealed the presence of a partial cDNA clone (Genbank No. AL133093) of 1,444 bp from human testis that is 100% identical to βIV spectrin in the region corresponding to nucleotides 4850–6082, but differs from it in the remaining 211 bp at the 3′ end. This divergent sequence encodes 30 residues, followed by a stop codon and a polyA stretch of 109 bp. On chromosome 19, however, the 102 bp preceding the polyA stretch are found immediately after exon 27 of βIV spectrin, where they are not preceded by a canonic 5′ splice site. Furthermore, this sequence does not contain a polyadenylation signal. Thus, this clone includes a part of the intron following exon 27.
Expression of βIV Spectrin mRNAs
The expression of βIV spectrin was analyzed by Northern blotting on polyA+ RNA from various human tissues using a 324 bp probe within the region encoding for the specific domain common to βIVΣ1 and βIVΣ3 spectrin. This probe hybridized with three transcripts of ∼9.0, ∼5.1, and ∼3.1 kb, all of which were predominantly expressed in brain (Fig. 3 A). The relationship of these transcripts with βIV spectrin was suggested by their detection with a distinct probe derived from the 3′ untranslated region of βIV spectrin (not shown). A mRNA of ∼9.0 kb is in good agreement with the size of the βIVΣ1 spectrin cDNA. Both the ∼9.0 and the ∼5.1 kb transcripts were also detected in various regions of the rat brain, including cerebellum, thalamus, midbrain, medulla, frontal, and posterior cortex and hippocampus (data not shown). Weaker signals for the ∼9.0-kb transcript were detected in human skeletal muscle and kidney, while the ∼5.1-kb transcript was barely detectable in pancreas (Fig. 3 A). Since pancreatic islets account for only ∼3% of the pancreas weight, we directly assessed the expression of βIV spectrin on polyA+ RNA isolated from purified human pancreatic islets. In pancreatic islets, as in brain, the βIV spectrin probe hybridized with three transcripts of ∼8.5, ∼4.2, and ∼2.3 kb (Fig. 3 C). The expression of βIV spectrin in pancreatic islets was confirmed by the amplification by PCR from mouse islets of a product whose cDNA (Genbank No. AF203696) was 98% identical to nucleotides 4223–4665 of human βIV spectrin.
Figure 3.

Northern blots for βIV spectrin on polyA+ RNA from various human tissues (A) and purified human pancreatic islets (C). (B) Autoradiography on the same filter as in A after hybridization with the control probe for β-actin. Heart and skeletal muscle express two β-actin mRNAs of ∼2 and 1.6–1.8 kb.
βIV Spectrin Proteins in Brain and Pancreatic Islets
We further analyzed by Western blotting the expression of βIV spectrin in brain and pancreatic islets, tissues that are enriched in ICA512. To this end, we used two affinity-purified antibodies directed against peptides encoding amino acids 2237–2256 in the specific domain (βIV-SD) and 2542-2559 at the COOH terminus (βIV-CT) of βIVΣ1 spectrin. These peptides were chosen on the basis of their predicted antigenicity and because they share no significant homology with sequences in other α and β spectrins. Both antibodies reacted by Western blotting with the βIV spectrin fragment B8 transiently expressed in COS cells (Fig. 1 F, and not shown). The βIV-CT antibody recognized two proteins of ∼250 and 160 kD in rat brain PNS and total extracts of purified human islets (Fig. 4 A), while it detected an additional protein of ∼140 kD in rat brain only (Fig. 4A and Fig. B). Preincubation of the βIV-CT antibody with the antigenic peptide blocked its binding to these three proteins, supporting their relationship with βIV spectrin (Fig. 4 A). Taking into account the aberrant electrophoretic mobility displayed by all known spectrins (Zagon et al. 1984; Ohara et al. 1998; Stankewich et al. 1998), the ∼250-kD protein might correspond to βIVΣ1 spectrin, which has a predicted molecular weight of ∼288 kD. The identity of the ∼160- and ∼140-kD proteins remains to be established. Accordingly, we will provisionally refer to these proteins as βIV spectrin 160 and 140. The βIV-SD antibody produced a similar immunoblot pattern (Fig. 4 B), consistent with the possibility that the ∼250-, ∼160-, and ∼140-kD proteins originate from the βIV spectrin gene. The βIV-SD antibody, however, recognized much less prominently βIV spectrin 160 in brain, and even less in islets (Fig. 4 B). The peptide used to raise the βIV-SD antibody is located within the ERQES domain and contains several optimal consensus sites for serine phosphorylation. We asked therefore whether the limited reactivity of the βIV-SD antibody toward βIV spectrin 160 could result from the phosphorylation of its epitope within this protein. Evidence that treatment of rat brain PNS with alkaline phosphatase dramatically enhanced the reactivity of the βIV-SD antibody toward βIV spectrin 160 (Fig. 4 B) corroborated this hypothesis. The labeling of βIVΣ1 spectrin and βIV spectrin 140, on the other hand, was unaffected. As expected, the reactivity of the βIV-SD antibody was completely blocked by the incubation with its immunogenic peptide (Fig. 4 B).
Figure 4.

(A) Western blotting with the affinity-purified antibody against a COOH-terminal peptide of βIV spectrin (βIV-CT) on 50 μg protein from rat brain PNS (brain, lanes 1 and 3) and homogenates of human pancreatic islets (islets, lanes 2 and 4). In the case of islets, the βIV-CT antibody was incubated with an excess of the immunogenic peptide before its incubation with the nitrocellulose strips. (B) Western blotting with the affinity-purified antibody against a peptide in the specific domain of βIV spectrin (βIV-SD) on 50 μg protein from rat brain PNS (brain, lanes 1, 3, and 4) and homogenates of human pancreatic islets (islets, lanes 2 and 5). (brain + alk. phos., lane 3) Rat brain PNS was treated with calf intestine alkaline phosphatase for 1 h at 37°C. (lanes 4 and 5) Western blotting with the βIV-SD antibody after preincubation with its antigenic peptide.
βIV Spectrins Have Different Biochemical Properties
Upon subcellular fractionation of brain homogenates, both βIVΣ1 spectrin and βIV spectrin 140 were recovered in the HSP, while βIV spectrin 160 was distributed between the HSS and the HSP (Fig. 5 A). Virtually all βIVΣ1 spectrin partitioned in the HSP detergent-insoluble material (Fig. 5A and Fig. B). βIV spectrin 160 and 140, on the other hand, were found in both the HSP detergent soluble and insoluble fractions (Fig. 5A and Fig. B). Notably, the pools of βIV spectrin 160 in the HSS and HSP detergent soluble fraction did not react with the βIV-SD antibody (Fig. 5 B), unless these fractions were preincubated with alkaline phosphatase (Fig. 5 C). These data suggest that phosphorylation of the ERQES domain is associated with the solubilization of βIV spectrin 160 and its dissociation from membranes. Treatment of the PNS with alkaline phosphatase did not affect the subcellular distribution of βIVΣ1 spectrin, but allowed an increased recovery of βIV spectrin 140 in the HSP-soluble fraction (Fig. 5 D). This treatment also resulted in a shift of most of βIV spectrin 160 from the HSP to the HSS (Fig. 5 D). Taken together, these results suggest that the interaction of βIV spectrins with cytoskeletal and/or membrane proteins is modulated by phosphorylation.
Figure 5.

Equal volumes of rat brain subcellular fractions immunoblotted with βIV-CT (A and D) or βIV-SD (B and C) antibodies. (C) Fractions were treated with alkaline phosphatase before immunoblotting. (D) The PNS was treated with alkaline phosphatase before fractionation. HSP-S, HSP-detergent soluble material. HSP-IS, HSP detergent-insoluble material.
Localization of βIV Spectrin in Pancreatic Islets
To rule out the possibility that our anti-βIV spectrin antibodies would recognize other β spectrins by immunocytochemistry, both antibodies were employed for the staining of CHO cells transiently transfected with V5-tagged COOH-terminal domains of βIΣ2 (amino acids 2117–2328), βIIΣ1 (2140–2365), βIII (amino acids 2154–2390), or βIVΣ1 spectrin (2222–2559). All four spectrin fragments were recognized by the anti-V5 mouse monoclonal antibody and partitioned, albeit to a different extent, between the cytosol and the nucleus of transfected cells (Fig. 6 A). Conversely, the βIV-CT (Fig. 6 A) and βIV-SD (not shown) antibodies only stained CHO cells expressing the COOH-terminal region of βIVΣ1 spectrin.
Figure 6.
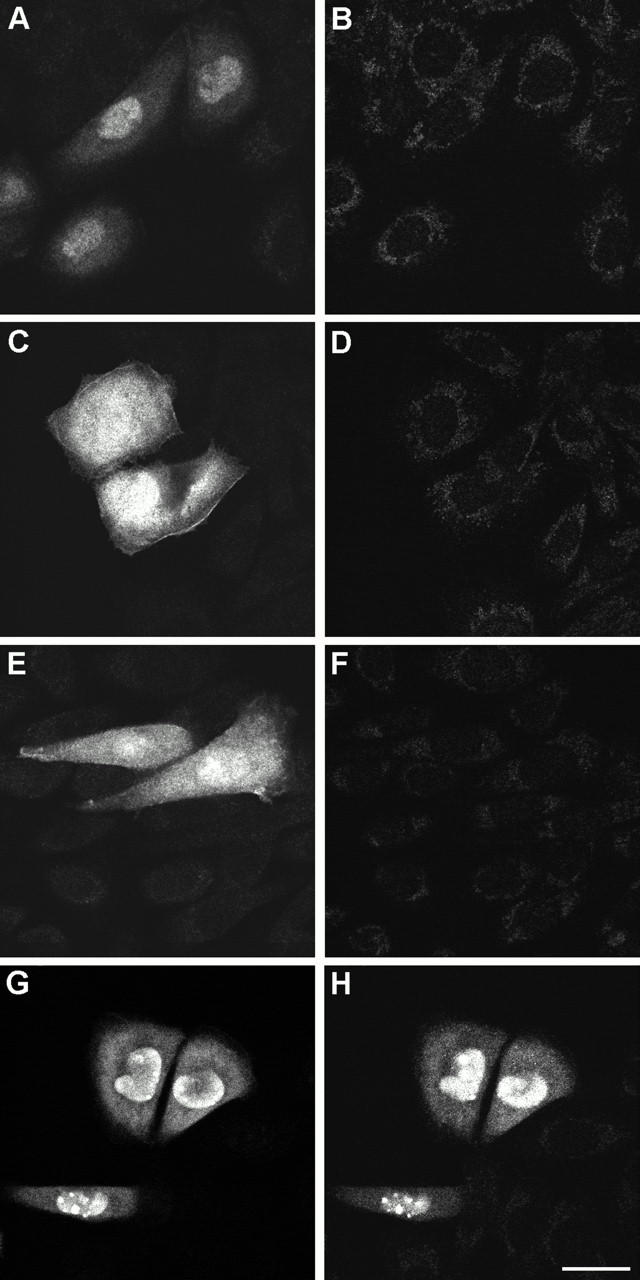
Double immunolabeling of V5-tagged COOH-terminal domains of human βIΣ2 (A and B), βIIΣ1 (C and D), βIII (E and F), and βIVΣ1 (G and H) spectrins with the V5 (left) and βIV-CT (right) antibodies in transiently transfected CHO cells. Bar, 25 μm.
The tissue and intracellular distribution of βIV spectrin was then investigated by confocal microscopy using the βIV-CT antibody. In rat pancreas, βIV spectrin immunoreactivity was restricted to pancreatic islets. Like ICA512, βIV spectrin was detected in both the insulin-secreting β cells as well as in the surrounding α cells that secrete glucagon (Fig. 7, A–F). In β cells, there was a fine punctate staining throughout the cytoplasm (not shown). A more intense cytoplasmic labeling was observed in the α cells (Fig. 7, A–F), often in correspondence of rod-like perinuclear structures of several micrometers in length (Fig. 7 G). These organelles did not significantly overlap with secretory granules, as shown by double labeling for glucagon (Fig. 7 H). The specificity of this staining was suggested by its complete disappearance when the βIV-CT antibody was preincubated with its antigenic peptide (Fig. 7. I). β Spectrins are known to associate with various intracellular compartments, including the Golgi complex, endosomes, and lysosomes (Devarajan et al. 1997; Hoock et al. 1997; Beck and Nelson 1998). Thus, we compared the distribution of βIV spectrin with that of these various organelles in α cells. There was no significant colocalization of βIV spectrin with γ-adaptin and TGN38, two markers of the trans-Golgi network, with the endosomal markers transferrin receptor and early endosomal antigen-1 or with the lysosome marker lgp120 (not shown). Thus, the identity of the βIV spectrin-immunoreactive structures in pancreatic islets remains to be established.
Figure 7.

Confocal microscopy for βIV spectrin (pseudo green), and insulin or glucagon (pseudo red) in rat pancreas. (A and B) Staining of a pancreatic islet for βIV spectrin (A) and insulin (B). (C) Merge of A and B. (D and E) Staining of a pancreatic islet for βIV spectrin (D) and glucagon (E). (F) Merge of D and E. (G and H) High magnifications of pancreatic islet α cells stained either for βIV spectrin alone (G) or both βIV spectrin and glucagon (H). (I) The βIV-CT antibody was incubated with an excess of the antigenic peptide before the incubation with the tissue sections. Bars, 100 μm (A–F), 10 μm (G–I).
βIV Spectrin Is Present at the Axon Initial Segments and Nodes of Ranvier in the Central and Peripheral Nervous System
In brain, expression of βIV spectrin was first examined by in situ hybridization on adult rat brain sections using an antisense riboprobe corresponding to the partial βIV spectrin clone B8. The highest levels of expression were detected in the cell bodies of large myelinated neurons, such as pyramidal neurons of the hippocampus, granule cells of the dentate gyrus, and Purkinje cells of the cerebellar cortex (Fig. 8). No signal was detected in nonneuronal cells, including glial and endothelial cells. Likewise, no signal was detected on sections incubated with the corresponding control sense riboprobe (not shown).
Figure 8.
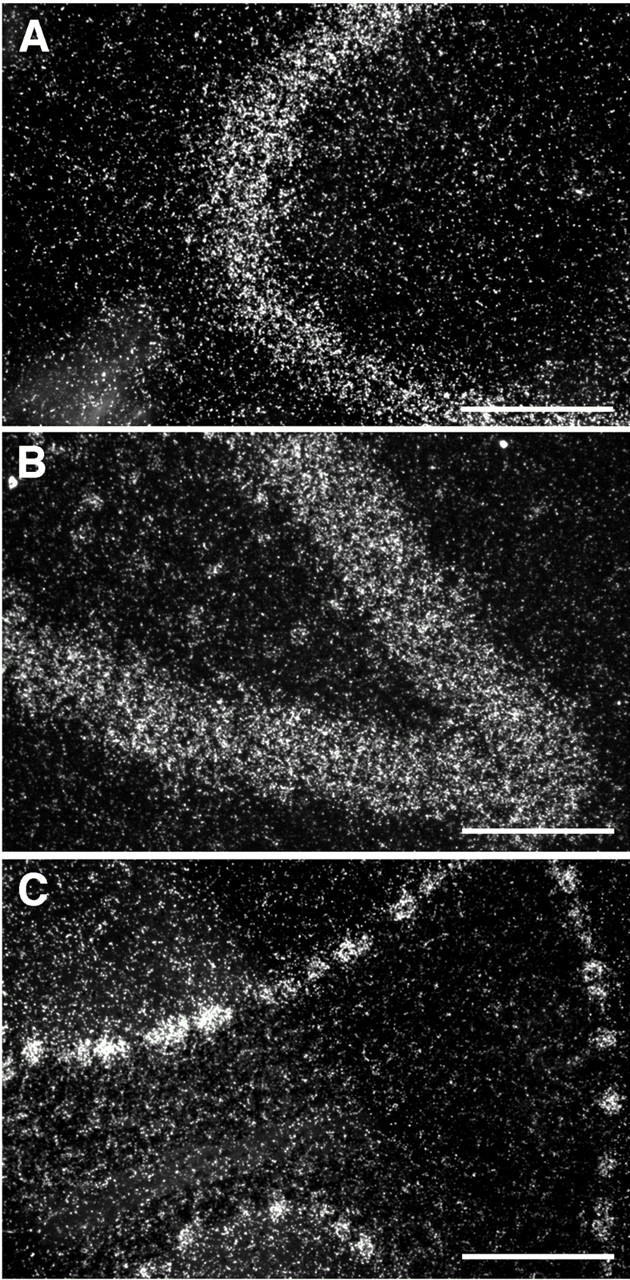
In situ hybridization for βIV spectrin on rat brain (dark field images). (A) CA3 region of the hippocampus, (B) dentate gyrus, (C) cerebellar cortex. Bars, 100 μm (A and B), 50 μm (C).
By immunocytochemistry on rat brain sections, the βIV-CT antibody produced a strong labeling of pyramidal neurons in the cerebral cortex and the hippocampus, granule cells of the dentate gyrus, and Purkinje cells in the cerebellum (Fig. 9, and data not shown). This immunolabeling was most prominent in the axon initial segments (Fig. 9A, Fig. D, and Fig. G) and nodes of Ranvier (D and H), while no staining was observed in the myelinated internodal segments, as visualized with an antibody against myelin basic protein (Fig. 9B and Fig. C). A fine granular staining reminiscent of the labeling observed in pancreatic islets was appreciable in the perikarya of Purkinje cells (Fig. 9 A). No immunoreactivity was detected in dendrites, axon terminals, glial cells, or blood vessels. The specificity of this labeling was supported by the absence of staining in sections incubated with the corresponding rabbit preimmune serum (not shown) and by the ability of the antigenic peptide to compete with endogenous βIV spectrin for the binding of the βIV-CT antibody (not shown). Immunocytochemistry on dissociated myelinated fibers from the rat sciatic nerve showed that, like in the central nervous system, βIV spectrin is concentrated at the nodes of Ranvier in the peripheral nervous system (Fig. 9 I).
Figure 9.
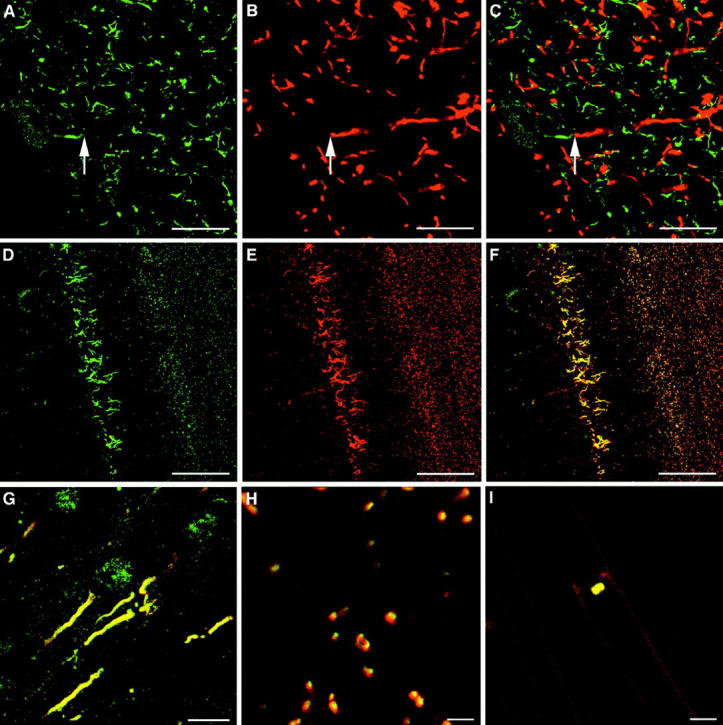
Confocal microscopy for βIV spectrin (pseudo green) and myelin basic protein or ankyrin G480/270-kD (pseudo red) in rat brain. (A and B) Staining of the cerebellar cortex for βIV spectrin (A) and myelin basic protein (B). (C) Merge of A and B. The arrow points to the boundary between the initial segment and the internodal segment of a Purkinje cell myelinated axon. (D and E) Staining of the hippocampus for βIV spectrin (D) and ankyrinG 480/270-kD (E). (F) Merge of D and E. (G) High power magnification of axon initial segments in the cerebral cortex costained for βIV spectrin and ankyrinG. (H) High power magnification of nodes of Ranvier in the cerebellum costained for βIV spectrin and ankyrinG. (I) Staining of teased nerve preparation with βIV spectrin and ankyrinG 480/270. Bars, 40 μm (A–C), 100 μm (D–F), 5 μm (H–I).
Previous studies demonstrated that, in neurons, the ankyrinG 480/270-kD isoform (also known as ankyrin3) is selectively concentrated at initial segments and nodes of Ranvier (Kordeli et al. 1990, Kordeli et al. 1995; Peters et al. 1995). Here, it is involved in the clustering of the voltage-gated Na+ channels that generate the saltatory action potentials (Zhou et al. 1998). Double immunostaining for βIV spectrin and ankyrinG 480/270 showed a perfect overlap of the two proteins at initial segments (Fig. 9, D–F) and central (H) and peripheral (I) nodes of Ranvier. These data raise the possibility that βIV spectrin and ankyrinG 480/270 bind each other in the brain as well as the peripheral nervous system.
The βIV Spectrin Isoform Localized at the Axon Initial Segments Is βIVΣ1 Spectrin
Next, we investigated the expression of βIV spectrin during brain development. By Western blotting, βIVΣ1 spectrin was first detected at embryonic day 19 and its expression level progressively increased until adult age (Fig. 10). βIV spectrin 140 appeared only after birth and, similar to βIVΣ1 spectrin, it was maximally expressed in the adult. βIV spectrin 160, instead, was already present at embryonic day 10, and its level appeared constant throughout development. These data further suggested that βIV spectrin 160 is not a proteolytic fragment of βIVΣ1 spectrin.
Figure 10.

Temporal expression of βIV spectrin during brain development. Western blotting with the βIV-CT antibody on PNS (40 μg protein) from rat brain at embryonic day 10 (E10), 15 (E15), 19 (E19), postnatal day 1 (P1), 10 (P10), and adult age.
The expression of βIV spectrin during brain development was also investigated by immunocytochemistry (Fig. 11). A weak immunoreactivity for βIV spectrin was detectable in the neurons of the developing hippocampus at embryonic day 15, but there was no staining reminiscent of axon initial segments (Fig. 11 A). By embryonic day 19, however, the initial segments were clearly labeled both in the cortex and the hippocampus (Fig. 11 B). Many more initial segments positive for βIV spectrin were present by postnatal day 10 in both regions (Fig. 11 C, and data not shown), with maximum immunoreactivity in the adult brain (D). Taken together, these data suggested that βIVΣ1 spectrin is the specific isoform localized at axon initial segments.
Figure 11.

Appearance of βIV spectrin immunoreactivity at axon initial segments during brain development. Confocal microscopy with the βIV-CT antibody (pseudo green) on sections from rat brain at embryonic day 15 (A), 19 (B), postnatal day 10 (C), and adult age (D). Bars, 50 μm.
Discussion
In this study, we have identified a novel spectrin gene in human, termed βIV spectrin, that is localized on chromosome 19q13.13. The longest product of this gene, termed βIVΣ1 spectrin, includes 36 exons and corresponds to a protein of 2,559 amino acids, whose domain structure closely resembles that of other β spectrins (for review, see Bennett and Gilligan 1993; Goodman et al. 1995; Viel and Branton 1996). Specifically, βIVΣ1 spectrin contains two calponin-homology domains at its NH2 terminus and may therefore associate with F-actin and protein 4.1. Similar to other β spectrins, it may also interact with ankyrin through spectrin repeat 15 and with an α spectrin via its partial spectrin repeat 17, whereas its COOH-terminal PH domain could bind phospholipids. βIVΣ1 spectrin also contains a specific domain between its partial spectrin repeat 17 and the PH domain. This very basic domain includes several putative SH3 binding sites and a unique ERQES domain in which there is a sequence of significant similarity with a motif within the membrane association domain of βII and βIII spectrin (Davis and Bennett 1994; Lombardo et al. 1994). Thus, this specific domain could mediate the interaction of βIVΣ1 with other proteins.
The apparent size of the βIVΣ1 spectrin transcript was ∼9.0 kb in brain and ∼8.5 kb in pancreatic islets. In both tissues, however, βIVΣ1 spectrin was detected as a protein of 250 kD. The small discrepancy in the size of the transcripts may result therefore from tissue-specific differences in the untranslated region or from variations in the laboratory procedures, as the blot of human islets was prepared in our laboratory, while the blot including the brain sample was acquired from a commercial source.
Similar to other β spectrins, the βIV spectrin gene undergoes alternative splicing. Specifically, we have identified three βIV spectrin splice variants in addition to βIVΣ1 spectrin. Two of these isoforms, βIVΣ2 and βIVΣ3 spectrin, originate from one messenger that contains an additional exon (exon 17B) between exons 17 and 18 of βIVΣ1 spectrin. The predicted amino acid sequences of βIVΣ2 and βIVΣ1 spectrin are identical up to the middle of the C helix in repeat 9. The sequence corresponding to the last 23 residues of βIVΣ2 spectrin is unique and does not follow the consensus for spectrin repeats, as it is very rich in prolines (6/23 amino acids). Accordingly, the last repeat of βIVΣ2 spectrin (repeat 9) is not complete, and contains only helices A and B. This observation raises the intriguing possibility that this partial repeat, similar to the partial repeat 17 of other β spectrins, including βIV spectrin, interacts with the lone C helix at the NH2 terminus of an α spectrin. An alternative possibility is that this partial repeat interacts in a head-to-tail fashion with the NH2-terminal domain of βIVΣ3 spectrin. Specifically, the first 19 amino acids of βIVΣ3 encoded by exon 17 cannot be part of an α helix because they include six prolines. The next 10 amino acids together with the following 15 residues encoded by exon 18, however, are predicted to form a coiled-coil domain (Lupas et al. 1991). Similar to the partial NH2-terminal repeat of α spectrin, this domain contains an arginine at position 8, a residue that is thought to interact with residues 7 and 29 in helices A and B, respectively (Yan et al. 1993). This domain of βIVΣ3 spectrin could therefore be equivalent to the single C helix at the NH2 terminus of an α spectrin and pair with the predicted A and B helices at the COOH terminus of βIVΣ2 spectrin. The fourth βIV spectrin isoform, termed βIVΣ4 spectrin, extends only 42 residues beyond the partial repeat 17, because of the insertion of an additional exon (exon 30B) that contains an in-frame stop codon. Thus, βIVΣ4 spectrin, similar to βIΣ1 (Winkelmann et al. 1990b) and βIIΣ2 (Hayes et al. 2000) spectrin, does not include a PH domain.
Both the βIV-SD and βIV-CT antibodies are directed against peptides that are not found in βIVΣ2 and βIVΣ4 spectrin. Thus, the biochemical properties and localization of these isoforms remain to be determined. Both antibodies, on the other hand, recognize two proteins of 160 and 140 kD. βIV spectrin 160 and 140 show significantly different properties than βIVΣ1 spectrin. For instance, treatment of the PNS with alkaline phosphatase increased the pool of βIV spectrin 160 recovered in the HSS, and enhanced the detergent extractability of βIV spectrin 140. Conversely, βIVΣ1 spectrin partitioned in all conditions in the HSP detergent insoluble material. Furthermore, βIV spectrin 140 was detected in brain, but not in islets. Finally, the temporal expression of βIV spectrins 160 and 140 during brain development was different from that of βIVΣ1 spectrin. These data suggest that βIV spectrin 160 and 140 are not proteolytic fragments of βIVΣ1 spectrin.
We have shown that the reactivity of the βIV-SD antibody with βIV spectrin 160 is significantly enhanced upon alkaline phosphatase treatment. Specifically, upon subcellular fractionation, this antibody reacts with the pool of βIV spectrin 160 recovered in the HSP detergent-insoluble material, but not with the pool present in the HSS and HSP detergent-soluble material, unless these fractions are incubated with alkaline phosphatase before immunoblotting. Since the antigenic peptide of the βIV-SD antibody contains an optimal consensus sequence for phosphorylation, these data suggest that the antigenic epitope within the ERQES domain is phosphorylated in vivo and that phosphorylation at this site affects the association of βIV spectrin 160 with membranes. This possibility is consistent with previous studies indicating that an increased phosphorylation of β spectrin during mitosis is associated with its redistribution from the detergent-insoluble to the detergent-soluble fraction and its dissociation from membranes (Fowler and Adam 1992).
Like ICA512, βIV spectrin is enriched in pancreatic islets and brain. In the α cells, βIV spectrin's immunoreactivity is concentrated in the cytosol, most often in perinuclear, rod-like structures. This compartment did not significantly overlap with the Golgi, endosomes, or lysosomes, three organelles along the secretory pathway where β spectrins have been found (Beck and Nelson 1998; De Matteis and Morrow 1998). Thus, its identity remains to be determined. In rat brain and teased fibers of the sciatic nerve, βIV spectrin is concentrated at the axon initial segments and the nodes of Ranvier. βIV spectrin is the first spectrin that is selectively enriched in these specialized domains, in both the central and the peripheral nervous tissue. βII spectrin is present both in axons and synaptic terminals (Riederer et al. 1986; Zagon et al. 1986; Trapp et al. 1989), while βIΣ2 spectrin is found in neuronal cell bodies, dendrites (Riederer et al. 1986; Zagon et al. 1986; Malchiodi-Albedi et al. 1993), and post-synaptic densities (Malchiodi-Albedi et al. 1993). βIII spectrin is also expressed in brain, where it is enriched in dendrites (Ohara et al. 1998; Stankewich et al. 1998). Taken together, these data suggest a role for β spectrins as potential organizers of different neuronal microdomains.
The axon initial segments and the nodes of Ranvier share a common molecular organization and may have evolved from a common precursor (Davis et al. 1996). Both compartments are characterized by an electron-dense cytoskeletal matrix beneath the axolemma (Matsumoto and Rosenbluth 1985) and by a 40-fold enrichment of voltage-gated sodium channels (Ritchie and Rogart 1977), whose opening triggers the generation of action potentials and their conduction from one node to the next. Other proteins enriched at these sites are the cell adhesion molecules neurofascin and NrCAM (Davis et al. 1996), ankyrinG 480/270-kD (Kordeli et al. 1995; Lambert et al. 1997), and an amphiphysin II–like protein (Butler et al. 1997). The repeat 15 of βIV spectrin is similar to the corresponding ankyrin-binding repeat in βI and βII spectrins (Kennedy et al. 1991). In view of their colocalization at initial segments and nodes of Ranvier, it seems plausible that βIV spectrin and ankyrinG are associated and together mediate the anchoring of nodal membrane proteins to the actin cytoskeleton.
In the developing rat hippocampus, the immunoreactivity for βIV spectrin at the initial segments first appears at embryonic day 19 and progressively increases thereafter. This pattern closely resembles the developmental expression profile of βIVΣ1 spectrin in rat brain. A similar correlation may exist between the appearance of βIV spectrin 140 at postnatal day 10 and the progressive development of nodes of Ranvier during myelination after birth. The initial segments and the nodes of Ranvier play a similar role in the initiation and propagation of action potentials. Initial segments, in addition, act as barriers that prevent lateral diffusion of membrane proteins and entry of various organelles, including rough endoplasmic reticulum, Golgi complex, and lysosomes into axons, while allowing the progression of others, such as mitochondria and secretory vesicles. Although the molecular foundation of this selective diffusion barrier remains unknown, there is evidence that the actin cytoskeleton is essential in its establishment and maintenance (Winckler et al. 1999). Thus, the localization of βIVΣ1 spectrin and βIV spectrin 140 at initial segments and nodes of Ranvier, respectively, could correlate with the related and yet distinct functions of these axonal compartments. It would be interesting to determine whether a similar different localization occurs in the case of the ankyrinG isoforms 480 and 270 kD.
We have identified βIV spectrin as an interactor of the RPTP-like protein ICA512 in a two-hybrid screening in yeast. The binding of ICA512 to the COOH-terminal domain of βIV spectrin has been confirmed by coimmunoprecipitation from transfected fibroblasts. βIV spectrin and ICA512 are both enriched in neurons and pancreatic islets, but their intracellular localization is different. This observation, however, does not preclude the possibility that ICA512, at one stage of its intracellular route, interacts with βIV spectrin. In steady state conditions, virtually all ICA512 (Lee et al. 1998; Hermel et al. 1999; our unpublished observations) is found on secretory granules (Solimena et al. 1996), while it is not detectable in other compartments associated with the secretory and endocytic pathways. Taking into account the fact that ICA512 has a very short half life (Lee et al. 1998; Hermel et al. 1999; our unpublished observations), it would be very difficult to detect by immunocytochemistry a transient colocalization of the two proteins in a compartment other than the secretory granules. More sensitive assays for protein–protein interaction in vivo, such as coimmunoprecipitation, were precluded by the fact that virtually all βIVΣ1 spectrin and most of βIV spectrin 140 and 160 partitioned in the HSP detergent insoluble fraction. We have recently shown that, in insulinoma cells, ICA512 is associated with β2 syntrophin (Ort et al. 2000). β2 syntrophin, in turn, interacts with the spectrin-related protein utrophin. These data, consistent with our previous studies (Hermel et al. 1999), suggest that ICA512 is associated with the actin cytoskeleton.
Besides ICA512, the COOH-terminal domain of βIV spectrin bound the cytoplasmic domain of the RPTP-like molecule PHOGRIN (Wasmeier and Hutton 1996). PHOGRIN (also known as IA-2β, ICAAR, IAR, PTP-NP, and PTPX) has many properties in common with ICA512. Its cytoplasmic domain is 74% identical to the corresponding region of ICA512, and also contains an aspartate instead of an alanine in its inactive PTP “signature motif.” Moreover, PHOGRIN is also enriched in the secretory granules of neurons and peptide-secreting endocrine cells (Wasmeier and Hutton 1996) and is a major target of autoimmunity in type I diabetes (Lu et al. 1996). ICA512 and PHOGRIN are not the only members of the RPTP family that interact with spectrin-related molecules. LAR, an RPTP expressed in numerous cell types including neurons, interacts through its distal, “non-catalytic” PTP domain with Trio, a protein that includes numerous spectrin repeats and two PH domains (Debant et al. 1996). CD45, an RPTP involved in T and B cell signaling, binds βII spectrin through its “inactive,” distal domain. Notably, binding of spectrin increases the catalytic activity of its proximal PTP domain (Lokeshwar and Bourguignon 1992; Iida et al. 1994). Like the “inactive” PTP domains of ICA512 and PHOGRIN, the distal PTP domain of CD45 includes an aspartate (D) rather than the obligatory alanine (A) at position +2 from the catalytic cysteine of canonic PTP domains. Overall, these data point to spectrin-related proteins as common binding partners of noncatalytic PTP domains of RPTPs.
Acknowledgments
We thank M. Altieri for technical assistance, Drs. P. De Camilli and E. Ullu for helpful discussion and encouragement, Dr. J. Morrow for discussion, Dr. C. Ricordi and the JDF Human Islet Distribution Centers at the University of Miami and Washington University for providing purified human islets, Dr. N. El-Sayed at TIGR Institute for foot-print analysis of human BAC clone, Drs. I. Mellman and G. Warren for the kind gifts of antibodies, Drs. J.A. Black and S.G. Waxman for the preparation of sciatic nerves, Dr. B.G. Forget for the βIΣ2 spectrin clone, and Drs. D.U. Rabin, S. Gleason, and D. Michaels at Bayer Corp. for their generous support.
This work was supported by grants to M. Solimena from the National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK), the American Diabetes Association (ADA), the Donaghue Foundation, and Bayer Diagnostic. S. Berghs and D. Aggujaro were the recipients of an ADA Mentor-based Post-Doctoral Fellowship. S. Berghs is currently supported by a postdoctoral fellowship of the Juvenile Diabetes Foundation. Immunomicroscopy and in-situ hybridization studies were supported by an NIDDK grant to the Yale Diabetes Endocrinology Research Center.
Note added in proof. While this manuscript was in revision, a partial amino acid sequence of our human βIV spectrin clone was reported in the manuscript by Stabach and Morrow (Stabach and Morrow 2000) on the cloning of human βV spectrin.
Footnotes
Portions of this work were previously published in abstract form (Berghs, S., D. Aggujaro, R. Dirkx, J.-P. Zhang, and M. Solimena. 1998. Soc. Neurosci. 24:204–210).
Abbreviations used in this paper: 3-AT, 3-amino-1,2,4-triazole; CT, COOH-terminal domain antiserum; HA, hemagglutinin; HB, homogenization buffer; HSP, high-speed pellet; HSS, high-speed supernatant; PH, pleckstrin homology; PNS, post-nuclear supernatant; PTP, protein tyrosine phosphatase; SD, specific domain antiserum.
References
- Altschul S.F., Gish W., Miller W., Myers E.W., Lipman D.J. Basic local alignment search tool. J. Mol. Biol. 1990;215:403–410. doi: 10.1016/S0022-2836(05)80360-2. [DOI] [PubMed] [Google Scholar]
- Banuelos S., Saraste M., Carugo K.D. Structural comparisons of calponin homology domainsimplications for actin binding. Structure. 1998;6:1419–1431. doi: 10.1016/s0969-2126(98)00141-5. [DOI] [PubMed] [Google Scholar]
- Beck K.A., Nelson W.J. A spectrin membrane skeleton of the Golgi complex. Biochim. Biophys. Acta. 1998;1404:153–160. doi: 10.1016/s0167-4889(98)00054-8. [DOI] [PubMed] [Google Scholar]
- Beck K.A., Buchanan J.A., Malhorta V., Nelson W.J. Golgi spectrinidentification of an erythroid β-spectrin homolog associated with the Golgi complex. J. Cell Biol. 1994;127:707–723. doi: 10.1083/jcb.127.3.707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett V., Gilligan D.M. The spectrin-based membrane skeleton and micron-scale organization of the plasma membrane. Annu. Rev. Cell Biol. 1993;9:27–66. doi: 10.1146/annurev.cb.09.110193.000331. [DOI] [PubMed] [Google Scholar]
- Burridge K., Kelly T., Mangeat P. Nonerythrocyte spectrinsactin-membrane attachment proteins occurring in many cell types. J. Cell Biol. 1982;95:478–486. doi: 10.1083/jcb.95.2.478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Butler M.H., David C., Ochoa G.C., Freyberg Z., Daniell L., Grabs D., Cremona O., De Camilli P. Amphiphysin II (SH3P9; BIN1), a member of the amphiphysin/Rvs family, is concentrated in the cortical cytomatrix of axon initial segments and nodes of Ranvier in brain and around T tubules in skeletal muscle. J. Cell Biol. 1997;137:1355–1367. doi: 10.1083/jcb.137.6.1355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Byers T.J., Branton D. Visualization of the protein associations in the erythrocyte membrane skeleton. Proc. Natl. Acad. Sci. USA. 1985;82:6153–6157. doi: 10.1073/pnas.82.18.6153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cameron P.L., Südhof T.C., Jahn R., De Camilli P. Colocalization of synaptophysin with transferrin receptorsimplications for synaptic vesicle biogenesis. J. Cell Biol. 1991;115:151–164. doi: 10.1083/jcb.115.1.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conboy J.G. Structure, function, and molecular genetics of erythroid membrane skeletal protein 4.1 in normal and abnormal red blood cells. Semin. Hematol. 1993;30:58–73. [PubMed] [Google Scholar]
- Davis J.Q., Lambert S., Bennett V. Molecular composition of the node of Ranvieridentification of ankyrin-binding cell adhesion molecules neurofascin (mucin+/third FNIII domain-) and NrCAM at nodal axon segments. J. Cell Biol. 1996;135:1355–1367. doi: 10.1083/jcb.135.5.1355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis L.H., Bennett V. Identification of two regions of βG spectrin that bind to distinct sites in brain membranes. J. Biol. Chem. 1994;269:4409–4416. [PubMed] [Google Scholar]
- De Camilli P., Cameron R., Greengard P. Synapsin I (protein I), a nerve terminal-specific phosphoprotein. I. Its general distribution in synapses of the central and peripheral nervous system demonstrated by immuno-fluorescence in frozen and plastic sections. J. Cell Biol. 1983;96:1337–1354. doi: 10.1083/jcb.96.5.1337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Matteis M.A., Morrow J.S. The role of ankyrin and spectrin in membrane transport and domain formation. Curr. Opin. Cell Biol. 1998;10:542–549. doi: 10.1016/s0955-0674(98)80071-9. [DOI] [PubMed] [Google Scholar]
- Debant A., Serra-Pages C., Seipel K., O'Brien S., Tang M., Park S.H., Streuli M. The multidomain protein Trio binds the LAR transmembrane tyrosine phosphatase, contains a protein kinase domain, and has separate ras-specific and rho-specific guanine nucleotide exchange factor domains. Proc. Natl. Acad. Sci. USA. 1996;93:5466–5471. doi: 10.1073/pnas.93.11.5466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng H., Lee J.K., Goldstein L.S., Branton D. Drosophila development requires spectrin network formation. J. Cell. Biol. 1995;128:71–79. doi: 10.1083/jcb.128.1.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devarajan P., Stabach P.R., De Matteis M.A., Morrow J.S. Na,K-ATPase transport from endoplasmic reticulum to Golgi requires the Golgi spectrin-ankyrin G119 in Madin Darby canine kidney cells. Proc. Natl. Acad. Sci. USA. 1997;94:10711–10716. doi: 10.1073/pnas.94.20.10711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dirkx R., Jr., Thomas A., Li L., Lernmark A., Sherwin R.S., De Camilli P., Solimena M. Targeting of the 67-kDa isoform of glutamic acid decarboxylase to intracellular organelles is mediated by its interaction with the NH2-terminal region of the 65-kDa isoform of glutamic acid decarboxylase. J. Biol. Chem. 1995;270:2146–2241. doi: 10.1074/jbc.270.5.2241. [DOI] [PubMed] [Google Scholar]
- Dubreuil R.R., Byers T.J., Stewart C.T., Kiehart D.P. A beta-spectrin isoform from Drosophila (beta H) is similar in size to vertebrate dystrophin. J. Cell Biol. 1990;111:1849–1858. doi: 10.1083/jcb.111.5.1849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunbar M.E., Young P., Zhang J.-P., McCaughern-Carucci J., Lanske B., Orloff J.J., Karaplis A., Cunha G., Wysolmerski J.J. Stromal cells are critical targets in the regulation of mammary ductal morphogenesis by parathyroid hormone-related protein. Dev. Biol. 1998;203:75–89. doi: 10.1006/dbio.1998.9029. [DOI] [PubMed] [Google Scholar]
- Fjell J., Hjelmstrom P., Hormuzdiar W., Milenkovic M., Aglieco F., Tyrrell L., Dib-Hajj S., Waxman S.G., Black J.A. Localization of the tetrodotoxin-resistant sodium channel NaN in nociceptors. Neuroreport. 2000;11:199–202. doi: 10.1097/00001756-200001170-00039. [DOI] [PubMed] [Google Scholar]
- Fowler V.M., Adam E.J. Spectrin redistributes to the cytosol and is phosphorylated during mitosis in cultured cells. J. Cell Biol. 1992;119:1559–1572. doi: 10.1083/jcb.119.6.1559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frappier T., Derancourt J., Pradel L.A. Actin and neurofilament binding domain of brain spectrin beta subunit. Eur. J. Biochem. 1992;205:85–91. doi: 10.1111/j.1432-1033.1992.tb16754.x. [DOI] [PubMed] [Google Scholar]
- Gallagher P.G., Forget B.G. Hematologically important mutationsspectrin and ankyrin variants in hereditary spherocytosis. Blood Cells Mol. Dis. 1998;24:539–543. doi: 10.1006/bcmd.1998.0217. [DOI] [PubMed] [Google Scholar]
- Glenney J.R., Jr., Glenney P., Weber K. F-actin-binding and cross-linking properties of porcine brain fodrin, a spectrin-related molecule. J. Biol. Chem. 1982;25:9781–9787. [PubMed] [Google Scholar]
- Godi A., Santone I., Pertile P., Devarajan P., Stabach P.R., Morrow J.S., Di Tullio G., Polishuck R., Petrucci T.C., Luini A., De Matteis M.A. ADP ribosylation factor regulates spectrin binding to the Golgi complex. Proc. Natl. Acad. Sci. USA. 1998;95:8607–8612. doi: 10.1073/pnas.95.15.8607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodman S.R., Zimmer W.E., Clark M.B., Zagon I.S., Barker J.E., Bloom M.L. Brain spectrinof mice and men. Brain Res. Bull. 1995;36:593–606. doi: 10.1016/0361-9230(94)00264-2. [DOI] [PubMed] [Google Scholar]
- Goodman S.R., Zagon I.S., Kulikowski R.R. Identification of a spectrin-like protein in nonerythroid cell. Proc. Natl. Acad. Sci. USA. 1981;12:7570–7574. doi: 10.1073/pnas.78.12.7570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayes N.V., Scott C., Heerkens E., Ohanian V., Maggs A.M., Pinder J.C., Kordeli E., Baines A.J. Identification of a novel C-terminal variant of (&bgr;)II spectrintwo isoforms of (&bgr;) II spectrin have distinct intracellular locations and activities. J. Cell Sci. 2000;113:2023–2034. doi: 10.1242/jcs.113.11.2023. [DOI] [PubMed] [Google Scholar]
- Hermel J.-M., Dirkx R., Jr., Solimena M. Post-translational modifications of ICA512, a receptor tyrosine phosphatase-like protein of secretory granules. Eur. J. Neurosci. 1999;11:2609–2620. doi: 10.1046/j.1460-9568.1999.00677.x. [DOI] [PubMed] [Google Scholar]
- Hoock T.C., Peters L.L., Lux S.E. Isoforms of ankyrin-3 that lack the NH2-terminal repeats associate with mouse macrophage lysosomes. J. Cell Biol. 1997;136:1059–1070. doi: 10.1083/jcb.136.5.1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu R.-J., Watanabe M., Bennett V. Characterization of human brain cDNA encoding the general isoform of beta-spectrin. J. Biol. Chem. 1992;267:18715–18722. [PubMed] [Google Scholar]
- Hyvonen M., Macias M.J., Nilges M., Oschkinat H., Saraste M., Wilmanns M. Structure of the binding site for inositol phosphates in a PH domain. EMBO (Eur. Mol. Biol. Organ.) J. 1995;14:4676–4685. doi: 10.1002/j.1460-2075.1995.tb00149.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iida N., Lokeshwar V.B., Bourguignon L.Y. Mapping the fodrin binding domain in CD45, a leukocyte membrane-associated tyrosine phosphatase. J. Biol. Chem. 1994;269:28576–28583. [PubMed] [Google Scholar]
- Karinch A.M., Zimmer W.E., Goodman S.R. The identification and sequence of the actin-binding domain of human red blood cell beta-spectrin. J. Biol. Chem. 1990;265:11833–11840. [PubMed] [Google Scholar]
- Kennedy S.P., Weed S.A., Forget B.G., Morrow J.S. A partial structural repeat forms the heterodimer self-association site of all beta-spectrins. J. Biol. Chem. 1994;269:11400–11408. [PubMed] [Google Scholar]
- Kennedy S.P., Warren S.L., Forget B.G., Morrow J.S. Ankyrin binds to the 15th repetitive unit of erythroid and nonerythroid β-spectrin. J. Cell Biol. 1991;115:267–277. doi: 10.1083/jcb.115.1.267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kordeli E., Davis J., Trapp B., Bennett V. An isoform of ankyrin is localized at nodes of Ranvier in myelinated axons of central and peripheral nerves. J. Cell Biol. 1990;110:1341–1352. doi: 10.1083/jcb.110.4.1341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kordeli E., Lambert S., Bennett V. Ankyrin Ga new ankyrin gene with neural-specific isoforms localized at the axon initial segment and nodes of Ranvier. J. Biol. Chem. 1995;270:2352–2359. doi: 10.1074/jbc.270.5.2352. [DOI] [PubMed] [Google Scholar]
- Lambert S., Davis J.Q., Bennett V. Morphogenesis of the node of Ranvierco-clusters of ankyrin and ankyrin-binding integral proteins define early developmental intermediates. J. Neurosci. 1997;17:7025–7036. doi: 10.1523/JNEUROSCI.17-18-07025.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lan M.S., Lu J., Goto Y., Notkins A.L. Molecular cloning and identification of a receptor-type protein tyrosine phosphatase, IA-2, from human insulinoma. DNA Cell Biol. 1994;13:505–514. doi: 10.1089/dna.1994.13.505. [DOI] [PubMed] [Google Scholar]
- Lee M.S., Dirkx R., Jr., Solimena M., Dannies P.S. Stabilization of the receptor protein tyrosine phosphatase-like protein ICA512 in GH4C1 cells upon treatment with estradiol, insulin, and epidermal growth factor. Endocrinology. 1998;139:2727–2733. doi: 10.1210/endo.139.6.6039. [DOI] [PubMed] [Google Scholar]
- Lokeshwar V.B., Bourguignon L.Y. Tyrosine phosphatase activity of lymphoma CD45 (GP180) is regulated by a direct interaction with the cytoskeleton. J. Biol. Chem. 1992;267:21551–21557. [PubMed] [Google Scholar]
- Lombardo R.C., Weed S.A., Kennedy S.P., Forget B.G., Morrow J.S. βII-spectrin (fodrin) and βIΣ2-spectrin (muscle) contain NH2- and COOH-terminal membrane association domains (MAD1 and MAD2) J. Biol. Chem. 1994;269:29212–29219. [PubMed] [Google Scholar]
- Lu J. Q., Li H., Xie Z.J., Chen A.E., Borovitskaya N.K., Maclaren A.L., Notkins, Lan M.S. Identification of a second transmembrane protein tyrosine phosphatase, IA-2beta, as an autoantigen in insulin dependent diabetes mellitusprecursur of the 37-kDa tryptic fragment. Proc. Natl. Acad. Sci. USA. 1996;93:2307–2311. doi: 10.1073/pnas.93.6.2307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lupas A., Van Dyke M., Stock J. Predicting coiled coils from protein sequences. Science. 1991;252:1162–1164. doi: 10.1126/science.252.5009.1162. [DOI] [PubMed] [Google Scholar]
- Ma Y., Zimmer W.E., Riederer B.M., Bloom M.L., Barker J.E., Goodman S.R. The complete amino acid sequence for brain beta spectrin (beta fodrin)relationship to globin sequences. Brain Res. Mol. Brain Res. 1993;18:87–99. doi: 10.1016/0169-328x(93)90176-p. [DOI] [PubMed] [Google Scholar]
- Magistrelli G., Toma S., Isacchi A. Substitution of two variant residues in the protein tyrosine phosphatase-like PTP35/IA-2 sequence reconstitutes catalytic activity. Biochem. Biophys. Res. Commun. 1996;227:581–588. doi: 10.1006/bbrc.1996.1549. [DOI] [PubMed] [Google Scholar]
- Malchiodi-Albedi F., Ceccarini M., Winkelmann J.C., Morrow J.S., Petrucci T.C. The 270 kDa splice variant of erythrocyte β-spectrin (βIΣ2) segregates in vivo and in vitro to specific domains of cerebellar neurons. J. Cell Sci. 1993;106:67–78. doi: 10.1242/jcs.106.1.67. [DOI] [PubMed] [Google Scholar]
- Marchesi S.L. The erythrocyte cytoskeleton in hereditary elliptocytosis and spherocytosis. In: Agre P., Parker J.C., editors. Red Blood Cell Membranes. Marcel Dekker Inc; New York, NY: 1988. pp. 77–110. [Google Scholar]
- Marchesi V.T., Steers E., Jr. Selective solubilization of a protein component of the red cell membrane. Science. 1968;159:203–204. doi: 10.1126/science.159.3811.203. [DOI] [PubMed] [Google Scholar]
- Matsumoto E., Rosenbluth J. Plasma membrane structure at the axon hillock, initial segment and cell body of frog dorsal root ganglion cells. J. Neurocytol. 1985;14:731–747. doi: 10.1007/BF01170825. [DOI] [PubMed] [Google Scholar]
- Ohara O., Ohara R., Yamakawa H., Nakajima D., Nakayama M. Characterization of a new beta-spectrin gene which is predominantly expressed in brain. Brain Res. Mol. Brain Res. 1998;57:181–192. doi: 10.1016/s0169-328x(98)00068-0. [DOI] [PubMed] [Google Scholar]
- Ort T., Maksimova E., Dirkx R., Kachinsky A.M., Berghs S., Froehner S.C., Solimena M. The receptor tyrosine phosphatase like-protein ICA512 binds the PDZ domains of β2-syntrophin and nNOS in pancreatic β-cells. Eur. J. Cell Biol. 2000;79:621–630. doi: 10.1078/0171-9335-00095. [DOI] [PubMed] [Google Scholar]
- Pascual J., Casteresana J., Saraste M. Evolution of the spectrin repeat. Bioessays. 1997;19:811–817. doi: 10.1002/bies.950190911. [DOI] [PubMed] [Google Scholar]
- Peters L.L., John K.M., Lu F.M., Eicher E.M., Higgins A., Yialamas M., Turtzo L.C., Otsuka A.J., Lux S.E. Ank3 (epithelial ankyrin), a widely distributed new member of the ankyrin gene family and the major ankyrin in kidney, is expressed in alternatively spliced forms, including forms that lack the repeat domain. J. Cell Biol. 1995;130:313–330. doi: 10.1083/jcb.130.2.313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rabin D.U., Pleasic S.M., Shapiro J.A., Yoo-Warren H., Oles J., Hicks J.M., Goldstein D.E., Rae P.M. Islet cell autoantigen 512 is a diabetes-specific islet autoantigen related to protein tyrosine phosphatases. J. Immunol. 1994;152:3183–3188. [PubMed] [Google Scholar]
- Rabin D.U., Pleasic S.M., Palmer-Crocker R., Shapiro A. Cloning and expression of IDDM-specific human autoantigens. Diabetes. 1992;41:183–186. doi: 10.2337/diab.41.2.183. [DOI] [PubMed] [Google Scholar]
- Riederer B.M., Zagon I.S., Goodman S.R. Brain spectrin (240/235) and brain spectrin (240/235E)two distinct spectrin subtypes with different locations within mammalian neural cells. J. Cell Biol. 1986;102:2088–2097. doi: 10.1083/jcb.102.6.2088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ritchie J.M., Rogart R.B. Density of sodium channels in mammalian myelinated nerve fibers and nature of the axonal membrane under the myelin sheath. Proc. Natl. Acad. Sci. 1977;USA. 74:211–215. doi: 10.1073/pnas.74.1.211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sambrook J., Fritsch E.F., Maniatis T.M. Molecular CloningA Laboratory Manual. 2nd ed. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, New York: 1989. [Google Scholar]
- Shizuya H., Birren B., Kim U., Mancino V., Slepak T., Tachiiri Y., Simon M. Cloning and stable maintenance of 300-kilobase-pair fragments of human DNA in Escherichia coli using an F-factor–based vector. Proc. Natl. Acad. Sci. 1992;USA. 89:8794–8797. doi: 10.1073/pnas.89.18.8794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sikorski A.F., Sangerman J., Goodman S.R., Critz S.D. Spectrin (βSpIIΣ1) is an essential component of synaptic transmission. Brain Res. 2000;852:161–166. doi: 10.1016/s0006-8993(99)02253-2. [DOI] [PubMed] [Google Scholar]
- Sikorski A.F., Terlecki G., Zagon I.S., Goodman S.R. Synapsin I–mediated interaction of brain spectrin with synaptic vesicles. J. Cell Biol. 1991;114:313–318. doi: 10.1083/jcb.114.2.313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solimena M. Vesicular autoantigens of type 1 diabetes. Diabetes Metab. Rev. 1998;14:227–240. doi: 10.1002/(sici)1099-0895(1998090)14:3<227::aid-dmr215>3.0.co;2-e. [DOI] [PubMed] [Google Scholar]
- Solimena M., Aggujaro D., Muntzel C., Dirkx R., Butler M., De Camilli P., Hayday A. Association of GAD-65, but not of GAD-67, with the Golgi complex of transfected Chinese hamster ovary cells mediated by the NH2-terminal region. Proc. Natl. Acad. Sci. USA. 1993;90:3073–3077. doi: 10.1073/pnas.90.7.3073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solimena M., Dirkx R., Jr., Hermel J.-M., Pleasic-Williams S., Shapiro J.A., Caron L., Rabin D.U. ICA512, an autoantigen of type I diabetes, is an intrinsic membrane protein of neurosecretory granules. EMBO (Eur. Mol. Biol. Organ.) J. 1996;15:2102–2114. [PMC free article] [PubMed] [Google Scholar]
- Stankewich M.C., Tse W.T., Peters L.L., Ch'ng Y., John K.M., Stabach P.R., Devarajan P., Morrow J.S., Lux S.E. A widely expressed betaIII spectrin associated with Golgi and cytoplasmic vesicles. Proc. Natl. Acad. Sci. USA. 1998;95:14158–14163. doi: 10.1073/pnas.95.24.14158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Speicher D.W., Marchesi V.T. Erythrocyte spectrin is comprised of many homologous triple helical segments. Nature. 1984;311:177–180. doi: 10.1038/311177a0. [DOI] [PubMed] [Google Scholar]
- Stabach P.R., Morrow J.S. Identification and characterization of βV spectrin, a mammalian ortholog of Drosophila betaH spectrin. J. Biol. Chem. 2000;275:21385–21395. doi: 10.1074/jbc.C000159200. [DOI] [PubMed] [Google Scholar]
- Stokke T., Collins C., Kuo W., Kowbel D., Shadravan F., Tanner M., Kallioniemi A., Kalloniemi O., Pinkel D., Deaven L., Gray J. A physical map of chromosome 20 established using fluorescent in situ hybridization (FISH) and digital image analysis. Genomics. 1995;26:134–137. doi: 10.1016/0888-7543(95)80092-z. [DOI] [PubMed] [Google Scholar]
- Trapp B.D., Andrews S.B., Wong A., O'Connell M., Griffin J.W. Co-localization of the myelin-associated glycoprotein and the microfilament components, F-actin and spectrin, in Schwann cells of myelinated nerve fibres. J. Neurocytol. 1989;18:47–60. doi: 10.1007/BF01188423. [DOI] [PubMed] [Google Scholar]
- Viel A., Branton D. Spectrinon the path from structure to function. Curr. Opin. Cell Biol. 1996;8:49–55. doi: 10.1016/s0955-0674(96)80048-2. [DOI] [PubMed] [Google Scholar]
- Vojtek A.B., Hollenberg S.M., Cooper J.A. Mammalian Ras interacts directly with the serine/threonine kinase Raf. Cell. 1993;74:205–214. doi: 10.1016/0092-8674(93)90307-c. [DOI] [PubMed] [Google Scholar]
- Wasmeier C., Hutton J.C. Molecular cloning of phogrin, a protein-tyrosine phosphatase homologue localized to insulin secretory granule membranes. J. Biol. Chem. 1996;271:18161–18170. doi: 10.1074/jbc.271.30.18161. [DOI] [PubMed] [Google Scholar]
- Winckler B., Forscher P., Mellman I. A diffusion barrier maintains distribution of membrane proteins in polarized neurons. Nature. 1999;397:698–701. doi: 10.1038/17806. [DOI] [PubMed] [Google Scholar]
- Winkelmann J.C., Forget B.G. Erythroid and nonerythroid spectrins. Blood. 1993;81:3173–3185. [PubMed] [Google Scholar]
- Winkelmann J.C., Chang J.-G, Tse W.T., Scarpa A.L., Marchesi V.T., Forget B.G. Full-length sequence of the cDNA for human erythroid β-spectrin J. Biol. Chem 265 1990. 11827 11832a [PubMed] [Google Scholar]
- Winkelmann J.C., Costa F.F., Linzie B.L., Forget B.G. β Spectrin in human skeletal muscle. Tissue specific differential processing of 3′ β spectrin pre-mRNA generates a β spectrin isoform with a unique carboxyl terminus J. Biol. Chem 265 1990. 20449 20454b [PubMed] [Google Scholar]
- Yan Y., Winograd E., Viel A., Cronin T., Harrison S.C., Branton D. Crystal structure of the repetitive segments of spectrin. Science. 1993;262:2027–2030. doi: 10.1126/science.8266097. [DOI] [PubMed] [Google Scholar]
- Yao L., Janmey P., Frigeri L.G., Han W., Fujita J., Kawakami Y., Apgar J.R., Kawakami T. Pleckstrin homology domains interact with filamentous actin. J. Biol. Chem. 1999;274:19752–29761. doi: 10.1074/jbc.274.28.19752. [DOI] [PubMed] [Google Scholar]
- Zagon I.S., Higbee R., Riederer B.M., Goodman S.R. Spectrin subtypes in mammalian brainan immunoelectron microscopic study. J. Neurosci. 1986;6:2977–2986. doi: 10.1523/JNEUROSCI.06-10-02977.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zagon I.S., McLaughlin P.J., Goodman S.R. Localization of spectrin in the mammalian brain. J. Neurosci. 1984;4:3089–3100. doi: 10.1523/JNEUROSCI.04-12-03089.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou D., Lambert S., Malen P.L., Carpenter S., Boland L.M., Bennett V. Ankyrin G is required for clustering of voltage-gated Na channels at axon initial segments and for normal action potential firing. J. Cell Biol. 1998;143:1295–1304. doi: 10.1083/jcb.143.5.1295. [DOI] [PMC free article] [PubMed] [Google Scholar]