

Abstract
Recent evidence suggests that in Escherichia coli, SecA/SecB and signal recognition particle (SRP) are constituents of two different pathways targeting secretory and inner membrane proteins to the SecYEG translocon of the plasma membrane. We now show that a secY mutation, which compromises a functional SecY–SecA interaction, does not impair the SRP-mediated integration of polytopic inner membrane proteins. Furthermore, under conditions in which the translocation of secretory proteins is strictly dependent on SecG for assisting SecA, the absence of SecG still allows polytopic membrane proteins to integrate at the wild-type level. These results indicate that SRP-dependent integration and SecA/SecB-mediated translocation do not only represent two independent protein delivery systems, but also remain mechanistically distinct processes even at the level of the membrane where they engage different domains of SecY and different components of the translocon. In addition, the experimental setup used here enabled us to demonstrate that SRP-dependent integration of a multispanning protein into membrane vesicles leads to a biologically active enzyme.
Keywords: membrane proteins, protein transport signal recognition particle, SecA, SecYEG complex
Introduction
In Escherichia coli, secretory proteins are targeted by means of the chaperone SecB and the ATPase SecA to the translocon of the plasma membrane, whose minimal pore consists of SecY, SecE, and SecG. In addition, E. coli contains Ffh (P48), which (together with 4.5S RNA) represents the bacterial homologue of the eukaryotic signal recognition particle (SRP), and FtsY, which is structurally related to the eukaryotic SRP receptor (SR) α subunit. Studies with conditional mutants of the SRP/SR components first suggested an involvement of the E. coli SRP and SR in the integration of proteins into the plasma membrane (MacFarlane and Müller 1995; de Gier et al. 1996; Seluanov and Bibi 1997; Ulbrandt et al. 1997). Recently, the first homologous assay system for SRP/SR of E. coli was described (Koch et al. 1999). Mannitol permease (MtlA) and SecY, two polytopic membrane proteins that lack large periplasmic domains, thus, were shown to require all three SRP/SR components for integration into inner membrane vesicles (INV) of E. coli. Most strikingly, integration of both proteins did not require SecA and SecB, whereas the translocation of a secretory protein (pOmpA) into INV was exclusively dependent on these Sec proteins. Both integration of MtlA and translocation of pOmpA, however, required an active SecY, suggesting that in E. coli, SRP/SR and SecA/SecB constitute two independent pathways that target specific substrates to the SecY-containing translocon of the plasma membrane. The specificity of SRP for polytopic inner membrane proteins is achieved by the ability of the ribosome-associated chaperone trigger factor to prevent binding of the SRP to the signal sequence of a secretory protein (Beck et al. 2000).
The concept of an exclusively SRP-dependent integration of E. coli inner membrane proteins, however, is not consistent with recent reports demonstrating the involvement of SecA in the membrane assembly of FtsQ and of AcrB-PhoA fusions (Valent et al. 1998; Qi and Bernstein 1999). To obtain independent evidence that, in E. coli, SRP/SR are sufficient to mediate proper membrane integration of multispanning proteins harboring short periplasmic loops, we have now challenged the involvement in integration of those parts of the E. coli translocon that are known to mediate the SecA function (i.e., SecG and the COOH terminus of SecY).
Materials and Methods
Strains and Plasmids
The following E. coli strains were used: MRE 600 (Müller and Blobel 1984), N4156 pARA14-FtsY′ (Luirink et al. 1994), CJ107 (secY24ts) (Wolfe et al. 1985), TY0 (ompT::kan), TY1 (ompT::kan, secY205) (Matsumoto et al. 1997), CU164 (secY39) (Baba et al. 1990), PS163 (secEcs15) (Schatz et al. 1991), CM124 (secEΔ19-111, pCM22) (Traxler and Murphy 1996), and KN553 ΔuncB-C::Tn10 ΔsecG::kan) (Nishiyama et al. 1996). For in vitro protein synthesis, the following plasmids were used: pDMB (OmpA) (Behrmann et al. 1998), p717MtlA-B (mannitol permease; Beck et al. 2000), pJM8CS7 (SecY) (Koch et al. 1999), pLacY-Bla#3 (Prinz and Beckwith 1994), and p717LamB-B (λ receptor [LamB]). To create p717LamB-B, the 1.5-kb HindIII-EcoRI fragment of pLB8000 (Swidersky et al. 1992) was truncated at the 5′ end and subcloned as a 1.4-kb BamHI-EcoRI fragment into pKSM717 (Maneewannakul et al. 1994). For in vitro expression of the secY205 allele, the plasmid pJM8CS7 was mutagenized using the QuikChange Kit (Stratagene) and the mutagenic primers secY205-1 (5′-ATGTCCAGTCAGGATGAGTCTGCATTG-3′) and secY205-2 (5′-TACAGGTCAGTCCTACTCAGACGTAAC-3′). Thus, the plasmid pETSecY205 created carries a T to G point mutation within secY replacing Y429 with D. The presence of this mutation was confirmed by DNA sequencing.
In Vitro Reactions
In vitro protein synthesis and the preparation of its components was performed as previously described (Hoffschulte et al. 1994; Koch et al. 1999). INV of the temperature-sensitive secY mutant CJ107 were obtained from cells grown either at 30 or 42°C. For the preparation of INV from the secE mutant PS163, cells were incubated for 2 h at 37°C, and then shifted to 25°C for additional 4 h before harvesting. INV from strain CM124 were prepared from cells grown either in the presence or absence of arabinose. Urea treatment of INV (generating U-INV) was performed as previously described (Helde et al. 1997).
SecY Integration Assay
In vitro synthesis of wild-type SecY and SecY205 was performed in the presence of INV or U-INV prepared from the secY205 mutant strain TY1. Mock-treated secY205-INV were generated by performing the in vitro reaction in the absence of DNA. When indicated, 8 ng/μl Ffh (P48) and 20 ng/μl FtsY were added. After 30 min incubation at 37°C, the entire reaction mixture was applied on a two-step sucrose gradient and separated into soluble material, membrane fraction, and pelleted material as described previously (Koch et al. 1999). The membranes were collected by centrifugation through a 750-mM sucrose cushion prepared in 50 mM triethanolamine acetate, pH 7.5, and employed for a translocation assay with in vitro synthesized OmpA as previously described (Koch et al. 1999). For U-INV, the translocation assay was performed in the presence of 80 ng/μl SecA, 80 ng/μl SecB, and 40 ng/μl F1-ATPase.
Sample Analysis and Quantification
All samples were analyzed on 13% SDS–polyacrylamide gels. Radiolabeled proteins were visualized by phosphorimaging using a Molecular Dynamics PhosphorImager and quantified using ImageQuant software from Molecular Dynamics.
Results
A secY Mutation Preventing a Productive Interaction with SecA Does Not Interfere with the Integration of Polytopic Membrane Proteins
A number of secY mutants have been isolated that display a diminished capability to support translocation of secretory proteins and to stimulate the ATPase activity of SecA (Baba et al. 1990; Matsumoto et al. 1997; Taura et al. 1997). To analyze whether these mutants are equally defective in the integrase activity of SecY, we prepared inside-out inner membrane vesicles (INV) from the three secY mutants, secY24ts, secY39cs, secY205cs, and tested them for their ability to support the integration of MtlA. As shown in Fig. 1 a, INV from the secY24ts mutant cells, grown at the nonpermissive temperature (42°C), were significantly impaired in both translocation of pOmpA and integration of MtlA when compared with wild-type vesicles. Likewise, INV from the sec39cs mutant were defective in both translocation and integration activity, although the effect on integration was less severe than that on translocation when compared with the secY24ts mutant. In contrast, secY205 INV, while possessing a strongly reduced translocation activity, did not exhibit any defect in the integration of MtlA.
Figure 1.
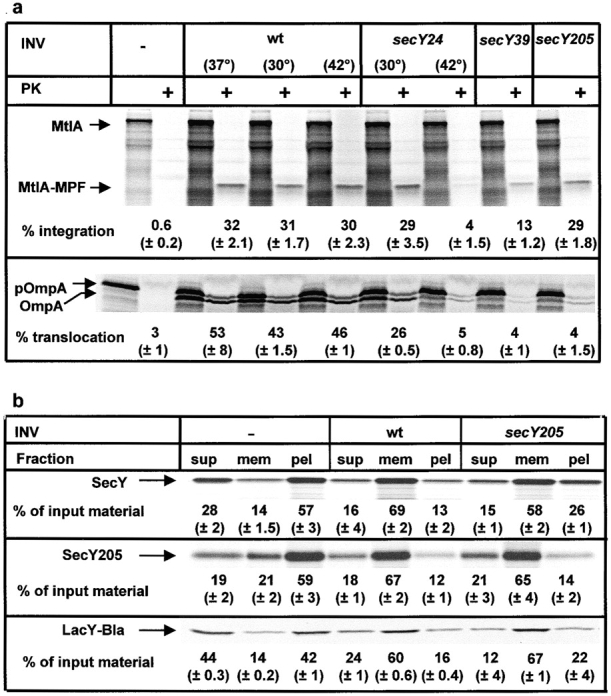
The secY205 mutant is exclusively impaired in the translocation of secretory proteins but not in the integration of polytopic membrane proteins. (a) The polytopic membrane protein mannitol permease (MtlA) and the secretory protein OmpA were synthesized in vitro in a cell-free translation system in the presence of inside-out inner membrane vesicles (INV) that were prepared from wild-type and different secY mutant strains. [35S]methionine-labeled translation products were either directly precipitated with TCA or only after incubation with 0.5 mg/ml proteinase K (PK) for 20 min at 25°C. Indicated are the positions of full-length MtlA, the membrane-protected fragment of MtlA resistant toward proteinase K (MtlA-MPF), the precursor (pOmpA), and the signal sequence–free form of OmpA. The percentage of integration was calculated after quantification of the radioactivity of individual protein bands using a PhosphorImager and calculating the ratio between MtlA-MPF and MtlA. The values obtained were corrected for the loss of Met residues occurring during cleavage by proteinase K. The percentage of translocation equals the ratio of radioactivity present in the proteinase K–resistant bands of pOmpA and OmpA and that recovered from the corresponding bands before proteolytic digestion. The values shown represent the means of at least three independent experiments with the SDs given in parenthesis. (b) Three polytopic inner membrane proteins, SecY, SecY205, a mutated version of SecY, and a lactose permease fusion protein (LacY-Bla) were synthesized in vitro in the presence of wild-type or secY205 INV. Membrane association was analyzed by subfractionation of the reaction mixture on a two-step sucrose gradient. Three fractions were sequentially withdrawn and analyzed on SDS-PAGE: the supernatant (sup), the membrane fraction (mem), and the pellet fraction (pel). Radioactivity present in the individual bands was quantified, and the sum of the three subfractions was each set at 100%.
Unimpaired integration into secY205 INV was confirmed for two additional polytopic membrane proteins. Using a two-step sucrose gradient, we have recently shown that SecY itself, like MtlA, integrates into INV in a strictly SRP-dependent manner (Koch et al. 1999). As illustrated in Fig. 1 b, cosedimentation of SecY with the membranes (69% compared with 14% in the INV-free control) did not significantly change by using INV from the secY205 mutant. Identical results were obtained with a LacY-Bla fusion protein, in which the first two transmembrane helices of lactose permease are fused to β-lactamase (Prinz and Beckwith 1994). As for SecY, cosedimentation of LacY-Bla with INV occurs independently of SecA, but it requires both Ffh and FtsY (data not shown). Using the same sedimentation analysis, only a minor portion of OmpA could be recovered from the secY205 membranes (data not shown). In summary, the secY205 mutation, which specifically impairs the SecA–SecY interaction (Matsumoto et al. 1997), does not interfere with the integration of the three membrane proteins tested here.
In Contrast to SecE, SecG Acting in Concert with SecA Is Not Required for the Integration of Polytopic Membrane Proteins
There is substantial evidence that SecY, together with SecE and SecG, constitutes the central core of the membrane-embedded protein translocase for secretory proteins (for review see Duong et al. 1997). To test whether SRP-dependent membrane proteins also require SecE for integration, INV from two conditional secE mutants grown under permissive and nonpermissive conditions were prepared. As shown in Fig. 2 a, the integration of MtlA into INV obtained after depletion of SecE was impaired to the same drastic extent as the translocation of OmpA. A lack of SecE also interfered with the integration of SecY and LacY-Bla as demonstrated by cosedimentation analysis (Fig. 2 b, compare wild-type and secE INV). The diminished integration observed in these conditions was in fact due to a depletion of SecE. This was verified by Western blot analyses (Fig. 2 c), which also revealed, for both secE mutant vesicles, the previously reported destabilization of SecY in the absence of SecE (Akiyama et al. 1996).
Figure 2.




In contrast to SecE, SecG is not required for the integration of polytopic membrane proteins. (a) MtlA and OmpA were synthesized in vitro as detailed in Fig. 1 in the presence of inner membrane vesicles prepared either from the wild-type, the cold-sensitive secE mutant PS163, the SecE-depletion mutant CM124, or the secG mutant KN553. Integration and translocation were quantified as described in Fig. 1. (b) Integration of SecY and LacY-Bla into INV, derived from the secG KN533 mutant, or the secE mutant CM124, grown in the absence of arabinose, was analyzed as specified in Fig. 1. (c) Immunoblot of wild-type and secE mutant vesicles using rabbit antisera directed against SecY and SecE and an HRP-based enhanced chemiluminescence detection system. (d) Translocation of pLamB was analyzed as specified for pOmpA translocation in Fig. 1.
SecG, the third component of the translocase is involved in the translocation of secretory proteins by assisting the insertion of the SecA–preprotein complex into the translocation channel (Nishiyama et al. 1994, Nishiyama et al. 1996; Matsumoto et al. 1997). Thus, it seemed conceivable that SecA-independent proteins do not require SecG for their membrane insertion. To examine this, INV were prepared from the secG mutant KN553 and tested for the integration of MtlA. As shown in Fig. 2 a, these INV did not exhibit any negative effect on MtlA integration, whereas translocation of OmpA was almost completely diminished. Strikingly, processing of pOmpA was not considerably altered in the secG mutant INV, albeit the obvious translocation defect. However, this effect was rather substrate specific since processing and translocation of pLamB, a second secretory protein tested, were both severely reduced by the same secG mutant INV (Fig. 2 d).
Membrane integration of SRP-dependent proteins in a SecG-independent manner was further substantiated for the two other membrane proteins SecY and LacY-Bla. Their cosedimentation with INV was not affected by the secG mutation, which clearly contrasts the results obtained with secE vesicles (Fig. 2 b). The finding that SecG is dispensable for the integration of membrane proteins is completely consistent with the observed SecA independence of this process.
SRP-dependent Integration of SecY into Membrane Vesicles Results in a Biologically Active Protein
The results described above rely on the premise that the integration of membrane proteins into INV, followed by the detection of protease resistance or cosedimentation with the vesicles, reflects the authentic in vivo events. The finding that the secY205 mutation exclusively impairs the translocation of secretory proteins but not the integration of membrane proteins, enabled us to design an integration assay monitoring the biological activity of a membrane protein as a criterion for its correct assembly. The concept is that if in vitro synthesized wild-type SecY correctly inserts into INV derived from the secY205 mutant, these INV would contain two versions of the SecY molecule, i.e., the inactive SecY205 version and the in vitro synthesized and integrated wild-type form. The latter should functionally reconstitute the mutant vesicles by restoring their translocation activity towards OmpA.
In a first step, SecY was synthesized in vitro in the presence of secY205 INV as described in Material and Methods. These INV were used for a translocation assay with OmpA as a substrate. As shown above, secY205 INV were not able to support translocation of OmpA (Fig. 3 a, lanes 5 and 6) in contrast to wild-type INV (Fig. 3, lanes 3 and 4). However, if in vitro synthesized wild-type SecY had been allowed to integrate into secY205 INV before the OmpA translocation assay, then the translocation efficiency increased almost 10-fold in comparison to mock-treated secY205 INV (Fig. 3, compare lanes 5 and 6 with 9 and 10). To confirm the ability of in vitro synthesized SecY to reconstitute secY mutant INV, despite being made in radiochemical amounts only, an inactive form of SecY was employed. When SecY205 instead of wild-type SecY was synthesized in vitro and allowed to integrate into mutant INV, the translocation activity of these vesicles did not increase beyond the level of mock-treated controls (Fig. 3, compare lanes 5 and 6 with 7 and 8). The failure of in vitro synthesized SecY205 to reconstitute the mutant INV was, however, not due to reduced expression levels or impaired membrane association of the SecY205. As shown in Fig. 1 b, the SecY205 mutant protein was expressed in vitro and integrated into INV at a level indistinguishable from wild-type SecY.
Figure 3.


SecY synthesized in vitro and integrated into INV is biologically active. (a) To reconstitute the secY205 INV in a first step, wild-type SecY and, as a negative control, SecY205 were in vitro synthesized in the presence of secY205 INV (pretreatment). The secY205 INV were isolated and subsequently tested for their ability to support the translocation of in vitro synthesized OmpA. No pretreatment was performed in the control reactions shown in lanes 1–4. Translocation was quantified as specified in Fig. 1. (b) To demonstrate the SRP dependency of the reconstitution process, vesicles were extracted with urea (U-INV) to remove Ffh and FtsY before reconstitution with in vitro synthesized SecY. Readdition of Ffh (SRP) and FtsY (SR) is indicated. After the individual pretreatments, vesicles were isolated and subsequently tested for translocation activity towards OmpA.
We have previously shown that integration of membrane proteins into urea-treated INV (U-INV) is strictly dependent on exogenously added Ffh and FtsY (Koch et al. 1999). This finding was applied to the functional reconstitution of secY205 INV with in vitro synthesized SecY. In contrast to untreated secY205 INV, the OmpA translocation activity of secY205 U-INV could not be restored by in vitro synthesized SecY alone (Fig. 3 b, compare lanes 9 and 10 with 11 and 12). However, if during reconstitution of secY205 U-INV, Ffh and FtsY were provided, the reisolated vesicles exhibited a significantly increased translocation efficiency for OmpA (lanes 13 and 14). Restoration required SecY and was not due solely to the presence of Ffh and FtsY, neither during preincubation of the vesicles (lanes 7 and 8) nor during the translocation reaction (lanes 15 and 16). In summary, these results clearly demonstrate that in vitro synthesized SecY has acquired biological activity after its SRP-dependent integration into membrane vesicles.
Discussion
Whether or not the integration of membrane proteins into the inner membrane of E. coli requires SecA in addition to SRP/SR, is still under debate. SecA can be cross-linked to nascent chains of the single spanning membrane protein FtsQ (Valent et al. 1998). Other membrane proteins that, like FtsQ, harbor long translocated domains were shown to depend on SecA for proper assembly (Wolfe et al. 1985; Traxler and Murphy 1996; Qi and Bernstein 1999). On the other hand, two polytopic membrane proteins recently were found not to require SecA, but only SRP/SR for integration. In this in vitro analysis, the exclusively SRP-dependent integration of MtlA and SecY into INV was assessed by protease resistance and cosedimentation, respectively (Koch et al. 1999). We have now designed a functional assay for correct integration into INV, which is compatible with the low amounts of membrane proteins usually synthesized in vitro. The finding that in vitro synthesized SecY acquires its biological activity after being integrated into membrane vesicles is a clear indication that this SRP-dependent integration faithfully reproduces the in vivo events.
These functional studies in conjunction with a more recent analysis, probing the molecular environment of nascent secretory and membrane E. coli proteins by cross-linking (Beck et al. 2000), altogether did not reveal any SecA dependence of the integration of multispanning membrane proteins devoid of major periplasmic loops. This is now further corroborated by two additional lines of evidence: (1) the integration of MtlA, LacY-Bla, and SecY is not affected by a mutation within the COOH-terminal domain of SecY that abolishes functional interaction with SecA. The theoretical possibility that SecA might still be involved in the integration of membrane proteins, yet in a manner not requiring interaction with the COOH terminus of SecY, is ruled out by the fact that the reconstitution of secY205 vesicles with wild-type SecY does not require the addition of SecA. (2) SecG, whose topology inversion is tightly coupled to the membrane insertion of SecA (Suzuki et al. 1998), is not required for the integration of MtlA, SecY, and LacY-Bla under conditions in which a lack of SecG almost completely abolishes the translocation of OmpA and LamB. Incidentally, the secG mutant INV had been prepared from an E. coli ΔuncB-C strain (Nishiyama et al. 1996) that lacks all components of the F1F0-ATPase. Under our experimental conditions, these vesicles cannot generate a H+ gradient. Nevertheless, integration of MtlA was unimpaired, which confirms earlier reports that the proton motive force is not required for the integration of such membrane proteins (Ahrem et al. 1989; Werner et al. 1992).
The SecA/SecB- and SRP/SR-mediated targeting pathways converge at the SecYEG translocon, as evidenced by cross-linking studies (Valent et al. 1998; Beck et al. 2000). This has been confirmed by the use of translocation intermediates that block the SecYEG channel and equally inhibit the translocation of OmpA as well as the integration MtlA (Koch et al. 1999). In addition, depletion of SecE not only affects the translocation of secretory proteins, but also the integration of membrane proteins (Schatz et al. 1991; Traxler and Murphy 1996; de Gier et al. 1998; and this study). In SecE-depleted membranes, SecY is rapidly degraded by the FtsH protease (Akiyama et al. 1996). The cytoplasmic domain C4 of SecY seems to be important for the SecY–SecE interaction since the secY24ts mutation positioned in the C4 domain of SecY abolishes copurification of SecE and SecY (Baba et al. 1994; Homma et al. 1997). Therefore, it is conceivable that the defects of the secY24ts mutant in both translocation and integration are primarily the result of an impaired SecY–SecE interaction. Interestingly, whereas the translocation defect of secY24 INV is partially suppressed by the addition of SecA (Fandl et al. 1988), no such improvement was observed for the integrase function of secY24ts INV (our unpublished results) which lends further support to the SecA independence of this process. Thus, in addition to SecE, the C4 domain of SecY might also interact with SecA.
Mutations within the C5 and C6 cytoplasmic domains of SecY interfere with the translocase activity of SecY (Baba et al. 1990; Taura et al. 1997). We show here that the secY39cs mutation, located within domain C5, also negatively affects the integration activity of SecY, yet to a lesser degree than translocation. On the other hand, the secY40 mutation, which is located in close proximity to the secY39 mutation selectively impairs the integration of fusions between membrane proteins and alkaline phosphatase, but has only minor effects on the translocation of secretory proteins (Newitt and Bernstein 1998). The secY205 mutant, which carries a mutation at the very COOH terminus of SecY, is specifically defective in the ATP-dependent SecA insertion into the plasma membrane (Matsumoto et al. 1997). This defect causes a significant translocation deficiency for SecA-dependent proteins (Taura et al. 1997; and this study), but leaves the integration of membrane proteins like MtlA, LacY-Bla, or SecY unaffected. In conclusion, while the SecY translocon is used for translocation of secretory proteins across, and integration of membrane proteins into the plasma membrane of E. coli, both activities can be dissected because they depend on different domains of SecY and require different components of the translocon.
Acknowledgments
We gratefully acknowledge Dr. Koreaki Ito (Institute for Virus Research Kyoto University, Kyoto, Japan) for providing the secY205 mutant, Drs. Ken-Ichi Nishiyama and Haijme Tokuda (Institute of Molecular and Cellular Biosciences, Tokyo University, Tokyo, Japan) for strain KN553, Dr. Andreas Kuhn (Institute for Microbiology, University Hohenheim, Stuttgart, Germany) for strain CM124, Dr. Michael Ehrmann (School of Biosciences, Cardiff University, Cardiff, UK) for strain PS163, and Dorothea Trescher (University of Freiburg) for the preparation of anti-SecE IgG. The SecE peptide was a gift of Dr. John M. Tomich (Kansas State University, Manhatten, Kansas).
This work was supported by a grant from the Sonderforschungsbereich 388 and the Fonds der Chemischen Industrie. M. Müller was also supported by a Quality of Life and Management of Living Resources grant (QLK3-CT-1999-00917) from the European Union.
Footnotes
Abbreviations used in this paper: INV, inside-out inner membrane vesicles; LamB, λ receptor; MtlA, mannitol permease; OmpA, outer membrane protein A; SR, SRP receptor, SRP, signal recognition particle.
References
- Ahrem B., Hoffschulte H.K., Müller M. In vitro membrane assembly of a polytopic, transmembrane protein results in an enzymatically active conformation. J. Cell Biol. 1989;108:1637–1646. doi: 10.1083/jcb.108.5.1637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akiyama Y., Kihara A., Tokuda H., Ito K. FtsH (HflB) is an ATP-dependent protease selectively acting on SecY and some other membrane proteins. J. Biol. Chem. 1996;271:31196–31201. doi: 10.1074/jbc.271.49.31196. [DOI] [PubMed] [Google Scholar]
- Baba T., Jacq A., Brickmann E., Beckwith J., Taura T., Ueguchi C., Akiyama Y., Ito K. Characterization of cold sensitive secY mutants of Escherichia coli . J. Bacteriol. 1990;172:7005–7010. doi: 10.1128/jb.172.12.7005-7010.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baba T., Taura T., Shimoike T., Akiyama Y., Yoshihisa T., Ito K. A cytoplasmic domain is important for the formation of a SecY-SecE translocator complex. Proc. Natl. Acad. Sci. USA. 1994;91:4539–4543. doi: 10.1073/pnas.91.10.4539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beck K., Wu L.F., Brunner J., Müller M. Discrimination between SRP- and SecA/SecB-dependent substrates involves selective recognition of nascent chains by SRP and trigger factor. EMBO (Eur. Mol. Biol. Organ.) J. 2000;19:134–143. doi: 10.1093/emboj/19.1.134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrmann M., Koch H.-G., Hengelage T., Wieseler B., Hoffschulte H.K., Müller M. Requirements for the translocation of elongation-arrested, ribosome-associated OmpA across the plasma membrane of Escherichia coli . J. Biol. Chem. 1998;273:13898–13904. doi: 10.1074/jbc.273.22.13898. [DOI] [PubMed] [Google Scholar]
- de Gier J.W.L., Mansournia P., Valent Q.A., Phillips G.J., Luirink J., von Heijne G. Assembly of a cytoplasmic membrane protein in Escherichia coli is dependent on the signal recognition particle. FEBS (Fed. Eur. Biochem. Soc.) Lett. 1996;399:307–309. doi: 10.1016/s0014-5793(96)01354-3. [DOI] [PubMed] [Google Scholar]
- de Gier J.W.L., Scotti P., Sääf A., Valent Q.A., Kuhn A., Luirink J., von Heijne G. Differential use of the signal recognition particle translocase targeting pathway for inner membrane protein assembly in Escherichia coli . Proc. Natl. Acad. Sci. USA. 1998;95:14646–14651. doi: 10.1073/pnas.95.25.14646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duong F., Eichler J., Price A., Leonard M.R., Wickner W. Biogenesis of the gram negative bacterial envelope. Cell. 1997;91:567–573. doi: 10.1016/s0092-8674(00)80444-4. [DOI] [PubMed] [Google Scholar]
- Fandl J.P., Cabelli R., Oliver D., Tai P.C. SecA suppresses the temperature-sensitive secY24 defect in protein translocation in Escherichia coli membrane vesicles. Proc. Natl. Acad. Sci. USA. 1988;85:8953–8957. doi: 10.1073/pnas.85.23.8953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helde R., Wieseler R., Wachter E., Neubüser A., Hoffschulte H.K., Hengelage T., Schimz K.-L., Stuart R.A., Müller M. Comparative characterization of secA from the α subclass purple bacterium Rhodobacter capsulatus and Escherichia coli reveals differences in membrane and precursor specificity. J. Bacteriol. 1997;179:4003–4012. doi: 10.1128/jb.179.12.4003-4012.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffschulte H.K., Drees B., Müller M. Identification of a soluble SecA/SecB-complex by means of a subfractionated cell-free export system. J. Biol. Chem. 1994;269:12833–12839. [PubMed] [Google Scholar]
- Homma T., Yoshihisa T., Ito K. Subunit interactions in the Escherichia coli protein translocaseSecE and SecG associate independently with SecY. FEBS (Fed. Eur. Biochem. Soc.) Lett. 1997;408:11–15. doi: 10.1016/s0014-5793(97)00376-1. [DOI] [PubMed] [Google Scholar]
- Koch H.-G., Hengelage T., Neumann-Haefelin C., Macfarlane J., Hoffschulte H.K., Schimz K.-L., Mechler B., Müller M. In vitro studies with purified components reveal signal recognition particle (SRP) and SecA/SecB as constituents of two independent protein-targeting pathways of Escherichia coli . Mol. Biol. Cell. 1999;10:2163–2173. doi: 10.1091/mbc.10.7.2163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luirink J., ten Hagen-Jongmann C.M., van der Weijden C.C., Oudega B., High S., Dobberstein B., Kusters R. An alternative protein targeting pathway in Escherichia colistudies on the role of FtsY. EMBO (Eur. Mol. Biol. Organ.) J. 1994;13:2289–2296. doi: 10.1002/j.1460-2075.1994.tb06511.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacFarlane J., Müller M. The functional integration of a polytopic membrane protein of Escherichia coli is dependent on the bacterial signal recognition particle. Eur. J. Biochem. 1995;233:766–771. doi: 10.1111/j.1432-1033.1995.766_3.x. [DOI] [PubMed] [Google Scholar]
- Maneewannakul S., Maneewannakul K., Ippen-Ihler K. The pKSM710 vector cassette provides tightly regulated lac and T7lac promoters for manipulating N-terminal protein sequences. Plasmid. 1994;31:300–307. doi: 10.1006/plas.1994.1032. [DOI] [PubMed] [Google Scholar]
- Matsumoto G., Yoshihisa T., Ito K. SecY and SecA interact to allow SecA insertion and protein translocation across the Escherichia coli plasma membrane. EMBO (Eur. Mol. Biol. Organ.) J. 1997;16:6384–6393. doi: 10.1093/emboj/16.21.6384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müller M., Blobel G. In vitro translocation of bacterial proteins across the plasma membrane of Escherichia coli . Proc. Natl. Acad. Sci. USA. 1984;81:7421–7425. doi: 10.1073/pnas.81.23.7421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newitt J.A., Bernstein H.D. A mutation in the Escherichia coli secY gene that produces distinct effects on inner membrane protein insertion and protein export. J. Biol. Chem. 1998;273:12451–12456. doi: 10.1074/jbc.273.20.12451. [DOI] [PubMed] [Google Scholar]
- Nishiyama K.-I., Hanada M., Tokuda H. Disruption of the gene encoding p12 (SecG) reveals the direct involvement and important function of SecG in the protein translocation of Escherichia coli at low temperature. EMBO (Eur. Mol. Biol. Organ.) J. 1994;13:3272–3277. doi: 10.1002/j.1460-2075.1994.tb06628.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishiyama K.-I., Suzuki T., Tokuda H. Inversion of the membrane topology of SecG coupled with SecA-dependent preprotein translocation. Cell. 1996;85:71–81. doi: 10.1016/s0092-8674(00)81083-1. [DOI] [PubMed] [Google Scholar]
- Prinz W.A., Beckwith J. Gene fusion analysis of membrane protein topologya direct comparison of alkaline phosphatase and β-lactamase fusions. J. Bacteriol. 1994;176:6410–6413. doi: 10.1128/jb.176.20.6410-6413.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi H.-Y., Bernstein H.D. SecA is required for the insertion of inner membrane proteins targeted by the Escherichia coli signal recognition particle. J. Biol. Chem. 1999;274:8993–8997. doi: 10.1074/jbc.274.13.8993. [DOI] [PubMed] [Google Scholar]
- Schatz P.J., Bieker K.L., Ottemann K.M., Silhavy T.J., Beckwith J. One of the three transmembrane stretches is sufficient for the functioning of the SecE protein, a membrane component of the E. coli secretion machinery. EMBO (Eur. Mol. Biol. Organ.) J. 1991;10:1749–1757. doi: 10.1002/j.1460-2075.1991.tb07699.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seluanov A., Bibi E. FtsY, the prokaryotic signal recognition particle receptor homologue, is essential for biogenesis of membrane proteins. J. Biol. Chem. 1997;272:2053–2055. doi: 10.1074/jbc.272.4.2053. [DOI] [PubMed] [Google Scholar]
- Suzuki H., Nishiyama K.-I., Tokuda H. Coupled structure changes of SecA and SecG revealed by the synthetic lethalitiy of the secAcsR11 and ΔsecG::kan double mutant. Mol. Microbiol. 1998;29:331–341. doi: 10.1046/j.1365-2958.1998.00937.x. [DOI] [PubMed] [Google Scholar]
- Swidersky U.E., Rienhöfer-Schweer A., Werner P.K., Ernst F., Benson S.A., Hoffschulte H.K., Müller M. Biochemical analysis of the biogenesis and function of the Escherichia coli export factor SecY. Eur. J. Biochem. 1992;207:803–811. doi: 10.1111/j.1432-1033.1992.tb17111.x. [DOI] [PubMed] [Google Scholar]
- Taura T., Yoshihisa T., Ito K. Protein translocation functions of Escherichia coli SecYin vitro characterization of cold-sensitive secY mutants. Biochimie. 1997;79:517–521. doi: 10.1016/s0300-9084(97)82744-7. [DOI] [PubMed] [Google Scholar]
- Traxler B., Murphy C. Insertion of the polytopic membrane protein MalF is dependent on the bacterial secretion machinery. J. Biol. Chem. 1996;271:12394–12400. doi: 10.1074/jbc.271.21.12394. [DOI] [PubMed] [Google Scholar]
- Ulbrandt N.D., Newitt J.A., Bernstein H.D. The E. coli signal recognition particle is required for the insertion of a subset of inner membrane proteins. Cell. 1997;88:187–196. doi: 10.1016/s0092-8674(00)81839-5. [DOI] [PubMed] [Google Scholar]
- Valent Q.A., Scotti P.A., High S., de Gier J.W.L., von Heijne G., Lentzen G., Wintermeyer W., Oudega B., Luirink J. The Escherichia coli SRP and SecB targeting pathways converge at the translocon. EMBO (Eur. Mol. Biol. Organ.) J. 1998;17:2504–2512. doi: 10.1093/emboj/17.9.2504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werner P.K., Saier M.H., Müller M. Membrane insertion of the mannitol permease of Escherichia coli occurs under conditions of impaired SecA function. J. Biol. Chem. 1992;267:24523–24532. [PubMed] [Google Scholar]
- Wolfe P.B., Rice M., Wickner W. Effects of two sec genes on protein assembly into the plasma membrane of Escherichia coli . J. Biol. Chem. 1985;260:1836–1841. [PubMed] [Google Scholar]