

Abstract
The localization of various Ca2+ transport and signaling proteins in secretory cells is highly restricted, resulting in polarized agonist-stimulated Ca2+ waves. In the present work, we examined the possible roles of the Sec6/8 complex or the exocyst in polarized Ca2+ signaling in pancreatic acinar cells. Immunolocalization by confocal microscopy showed that the Sec6/8 complex is excluded from tight junctions and secretory granules in these cells. The Sec6/8 complex was found in at least two cellular compartments, part of the complex showed similar, but not identical, localization with the Golgi apparatus and part of the complex associated with Ca2+ signaling proteins next to the plasma membrane at the apical pole. Accordingly, immunoprecipitation (IP) of Sec8 did not coimmunoprecipitate βCOP, Golgi 58K protein, or mannosidase II, all Golgi-resident proteins. By contrast, IP of Sec8 coimmunoprecipitates Sec6, type 3 inositol 1,4,5-trisphosphate receptors (IP3R3), and the Gβγ subunit of G proteins from pancreatic acinar cell extracts. Furthermore, the anti-Sec8 antibodies coimmunoprecipitate actin, Sec6, the plasma membrane Ca2+ pump, the G protein subunits Gαq and Gβγ, the β1 isoform of phospholipase C, and the ER resident IP3R1 from brain microsomal extracts. Antibodies against the various signaling and Ca2+ transport proteins coimmunoprecipitate Sec8 and the other signaling proteins. Dissociation of actin filaments in the immunoprecipitate had no effect on the interaction between Sec6 and Sec8, but released the actin and dissociated the interaction between the Sec6/8 complex and Ca2+ signaling proteins. Hence, the interaction between the Sec6/8 and Ca2+ signaling complexes is likely mediated by the actin cytoskeleton. The anti-Sec6 and anti-Sec8 antibodies inhibited Ca2+ signaling at a step upstream of Ca2+ release by IP3. Disruption of the actin cytoskeleton with latrunculin B in intact cells resulted in partial translocation of Sec6 and Sec8 from membranes to the cytosol and interfered with propagation of agonist-evoked Ca2+ waves. Our results suggest that the Sec6/8 complex has multiple roles in secretory cells including governing the polarized expression of Ca2+ signaling complexes and regulation of their activity.
Keywords: Sec6/8 complex, Ca2+ signaling proteins, assembly, actin cytoskeleton, Ca2+ signaling
Introduction
Ca2+ signaling in secretory cells is highly polarized (Muallem and Wilkie 1999). Agonist stimulation triggers Ca2+ signals in the form of Ca2+ waves that are initiated in the luminal pole and propagate along the cell periphery and the lateral membrane to the basal pole (Kasai et al. 1993; Thorn et al. 1993; Nathanson et al. 1994; Xu et al. 1996; Lee et al. 1997b). The pattern of [Ca2+]i waves is determined, at least in part, by the pattern of expression of multiple Ca2+ signaling and transport proteins. Thus, the inositol 1,4,5-trisphosphate receptors (IP3Rs) Ca2+ release channels (Lee et al. 1997b; Yule et al. 1997) and plasma membrane Ca2+ pumps (PMCA; Lee et al. 1997a) are expressed at high levels in the luminal pole. The sarco/ER Ca2+ pumps (SERCA) are expressed in a cell- and region-specific manner with high levels along the lateral and subapical region (Lee et al. 1997a). The same lateral–subapical region is enriched with several G-protein coupled receptors (Rios et al. 1999). Accordingly, Ca2+ release from the apical pole is the most sensitive to agonist stimulation and to activation by IP3 (Kasai et al. 1993; Thorn et al. 1993).
An important question in Ca2+ and other forms of cell signaling in polarized cells is how the polarized localization of signaling complexes is achieved. The Sec6/8 complex may determine signaling polarity since it is proposed to play a central role in the establishment of cell polarity from yeast to mammals (Finger and Novick 1998; Hsu et al. 1999). Proteins of the Sec6/8 complex were first identified in yeast as proteins involved in exocytosis (Bowser et al. 1992). Later, the proteins were found to form a multisubunit complex, termed the exocyst. Genetic and biochemical analysis showed the exocyst to contain eight subunits, Sec3p, -5p, -6p, -8p, -10p, -15p, Exo70p, and Exo84p (TerBush et al. 1996; Guo et al. 1999). During vegetative growth, the exocyst localizes to the emerging bud tip, and during cytokinesis the exocyst localizes to the site of cytokinesis (TerBush and Novick 1995; Finger and Novick 1998; Finger et al. 1998). The Sec3 subunit serves as a landmark for vesicle delivery since its localization is independent of the secretory pathway and the actin cytoskeleton (Finger et al. 1998). Although the function of the other subunits is not known at present, deletion of individual subunits and/or expression of dominant negatives result in accumulation of secretory vesicles in the cytoplasm and nonpolarized exocytosis and cell growth (Finger and Novick 1998). Hence, in yeast the exocyst mediates the polarized delivery of secretory vesicles to regions of active exocytosis.
The mammalian Sec6/8 complex was originally purified from rat brain and shown to be comprised of eight subunits, analogues to the yeast exocyst (Hsu et al. 1996). Furthermore, strong evidence suggests that the mammalian Sec6/8 complex participates in polarized delivery of vesicles. In the epithelial cell line MDCK cells, the Sec6/8 complex regulates vesicle targeting to the basolateral, but not to the apical membrane (Grindstaff et al. 1998). In developing cultured hippocampal neurons, the Sec6/8 complex localizes to regions of cell growth, including axonal and dendritic growth cones, as well as sites of synaptic formation along axons (Hazuka et al. 1999).
Localization of the Sec6/8 complex is dynamically regulated. In yeast, the exocyst localizes to the bud tip until the bud is about one-half the size of the mother. Then it is disassembled and reformed at the site of cell division (Finger and Novick 1998). In nonpolarized MDCK cells, the complex is soluble in the cytosol. Upon formation of cell–cell contacts, it is found in cell–cell interacting domains. In polarized monolayers, the Sec6/8 complex colocalizes with ZO1 at or near tight junctions (TJ; Grindstaff et al. 1998). After establishment of stable synapses, the Sec6/8 complex is no longer enriched in synapses (Hazuka et al. 1999). This dynamic translocation of the Sec6/8 complex may be mediated by the actin cytoskeleton. In yeast, disruption of the actin cytoskeleton prevents the polarized localization of Sec8 (Ayscough et al. 1997; Finger et al. 1998). The mammalian Sec6/8 complex has been suggested to interact with the cytoskeletal septins (Hsu et al. 1998).
The mammalian Sec6/8 complex is ubiquitously expressed (Hsu et al. 1999), but its function has been examined only in MDCK and neuronal cells. Nothing is known about the localization and function of the Sec6/8 complex in somatic cells. In the present work, we examined the localization and possible roles of the Sec6/8 complex in modulating Ca2+ signaling in pancreatic acinar cells. Immunolocalization by confocal microscopy showed that part of the Sec6/8 complex forms a basket around secretory granules. Another part of the complex associates with Ca2+ signaling complexes to regulate their activity. The Sec6/8 complex can be coimmunoprecipitated with Ca2+ signaling proteins and disruption of this interaction disrupts Ca2+ signaling. Our results suggest that the Sec6/8 complex has several, probably related, functions in polarized epithelial cells, including governing localization and regulating the function of the Ca2+ signaling machinery.
Materials and Methods
Materials
Brefeldin A (BFA) and carbachol were purchased from Sigma-Aldrich. Latruculin B, IP3, and 2,4,5-IP3 were from Alexis. The anti-Sec6 and Sec8 mAbs were a generous gift from Dr. Richard Scheller (Stanford University, Stanford, CA) and Shu-Chan Hsu (Rutgers, Piscataway, NJ). Polyclonal antibodies (pAbs) recognizing Gαq and PLC-β1 were a gift from Dr. Paul Sternweis (University of Texas Southwestern, Dallas, TX). Anti-Gβ pAb was a gift by Dr. Susan Mummby (University of Texas Southwestern, Dallas, TX). Dr. Richard Wojcikiewicz (State University of New York, Syracuse, NY) generously provided anti-IP3R1 and IP3R2 pAbs. The NH2-terminal fragment of Gelsolin was a gift from Dr. Helen Yin (University of Texas Southwestern, Dallas, TX). Antiactin pAb, anti-βCOP, and anti-Golgi 58K protein mAb were from Sigma-Aldrich. Antimannosidase II (anti-Man II) pAbs were obtained from the University of Georgia. Anti-PMCA mAbs, anti-IP3R3 mAbs, anti-ZO1 pAbs, and antioccludin pAbs were purchased from ABR, Transduction Laboratories, and Zymed Laboratories, respectively. Antisynaptotagmin pAbs were from Alomane Labs. Antisyncollin pAbs were raised in rabbits against a GST–syncollin construct.
Preparation of Pancreatic Acini and Single Acinar Cells
Acini were prepared from the pancreas of 100–150 g rats by limited collagenase digestion as described previously (Xu et al. 1996). The acini were resuspended in a standard solution A containing (in mM) NaCl 140, KCl 5, MgCl2 1, CaCl2 1, Hepes 10 (pH 7.4 with NaOH), glucose 10, and 0.1% BSA, and kept on ice until use. Single acinar cells were obtained by incubation of a minced pancreas in a 0.025% trypsin, 0.02% EDTA solution for 5 min at 37°C (Zeng et al. 1996). After washing the tissue with solution A supplemented with 0.02% soybean trypsin inhibitor, single cells were liberated by 7-min incubation at 37°C in the same solution that also contained 160 U/ml pure collagenase. The cells were washed with solution A and kept on ice until use.
[Ca2+]i Imaging
Pancreatic acinar cells were loaded with Fura 2 and [Ca2+]i was imaged as detailed before (Xu et al. 1996). Fura 2 fluorescence was measured at a single excitation wavelength of 380 nm, by averaging four consecutive images for each time point. Under these conditions using a frame size of 256 × 240 pixels recording was at a resolution of 90 msec/averaged image. During perfusion with the control solution and just before the first stimulation, the image of resting cells was acquired. This was taken as the fluorescence at time 0 (F0). Pixel values of all subsequent images were divided by this image, and the traces are the calculated F0/Ft, where Ft is the fluorescence at time t.
Electrophysiology
The whole cell configuration of the patch clamp technique was used for measurement of Ca2+-activated Cl− current, which directly correlates with changes in [Ca2+]i (Zeng et al. 1996). The experiments were performed with single acinar cells perfused with solution A. The standard pipette solution contained (in mM): KCl 140, MgCl2 1, EGTA 0.05, Na2ATP 5, and Hepes 10 (pH 7.3 with KOH) as described in previous studies (Zeng et al. 1996). The Sec6 and Sec8 antibodies (Abs) were dialyzed against an ATP-free pipette solution and concentrated to original volume with a centricone. Seals of 6–10 GΩ were produced on the cell membrane, and the whole cell configuration was obtained by gentle suction or voltage pulses of 0.5 V for 0.3–1 ms. The patch clamp output (Axopatch-1B, Axon Instruments) was filtered at 20 Hz. Recording was performed with patch clamp 6 and a Digi-Data 1,200 interface (Axon Instruments). All the traces shown were recorded at a holding potential of −40 mV.
Immunoprecipitation
Pancreatic microsomes were prepared by homogenizing a minced pancreas in a buffer containing (in mM, pH 6.7 with KOH) Mops 20, sucrose 250, EDTA 1, MgCl2 1, benzamidine 10, and PMSF 0.2. The homogenate was centrifuged at 400 g for 10 min. The supernatant was collected and centrifuged at 900 g for 10 min at 4°C. The loose pellet was resuspended in the same buffer while avoiding suspension of the hard, white-colored granular fraction in the bottom of the tube. When needed, the fraction enriched in secretory granules was collected in homogenization buffer into a separate tube. To avoid protein degradation by digestive enzymes, IP was initiated immediately after completion of microsomal preparation. Brain microsomes were prepared by homogenizing rat brain in a buffer containing (in mM, pH 7.6 with KOH) KCl 100, Tris-base 20, EDTA 1, benzamidine 1, and PMSF 1. The homogenate was centrifuged at 1,000 g for 10 min at 4°C. The supernatant was collected and centrifuged at 40,000 g for 30 min. The pellet was resuspended in homogenization buffer and the microsomes were stored at −80°C until use. Pancreatic or brain microsomes were extracted by a 1-h incubation on ice with a buffer containing (in mM) Tris 50 (pH 6.8 with HCl), NaCl 150, EDTA 2, EGTA 2, and 0.5% Triton X-100 supplemented with protease inhibitors (0.2 mM PMSF, 10 μg/ml leupeptin, 15 μg/ml aprotinin, 1 mM benzamidine). The lysate was cleared by centrifugation at 14,000 g for 10 min. About 300 μl of the extract was further incubated with 15 μl of Sepharose A beads for 1 h at 4°C and centrifuged for 2 min at 14,000 g to remove the beads. The cleared supernatant was incubated with 5 μl anti-Sec8, 5 μl anti-PMCA, 10 μl anti-PLC-β1, or 10 μl anti-IP3R1 Abs for 30 min before addition of 30 μl Sepharose A beads and an overnight incubation at 4°C under gentle agitation. The beads were washed five times with 0.8 ml lysis buffer and stripped of proteins by boiling in a 50 μl of SDS sample buffer. To test the effect of the actin cytoskeleton on the binding of the Sec6/8 and Ca2+ signaling complexes, buffer or 20 μg/ml of the NH2-terminal fragment of gelsolin was added to equal portions of beads after the second wash. After 20-min incubation at 4°C, the beads were washed three times with lysis buffer and the proteins remaining attached to the beads were released by boiling in a sample buffer. Released proteins were separated by an SDS-PAGE using 7.5% polyacrylamide gels. The separated proteins were transferred to 0.2 μm polyvinylidene difluoride membranes, and the membranes were blocked by a 1-h incubation at room temperature in 5% nonfat dry milk in a solution containing 20 mM Tris-HCl, pH 7.5, 150 mM NaCl, and 0.05% Tween 20 (TTBS). The Sec6/8 and other proteins were detected by a 1–2-h incubation of individual membranes with the respective Ab diluted in TTBS.
Immunocytochemistry
Cells attached to glass coverslips were fixed and permeabilized with 0.5 ml of cold methanol for 10 min at −20°C, except for the experiments in Fig. 9 where the cells were fixed with 4% formaldehyde for 20 min at room temperature, followed by permeabilization with 0.05% Triton X-100. After removal of methanol or Triton X-100, the cells were washed with PBS and incubated in 0.5 ml of PBS containing 50 mM glycine for 10 min at room temperature. This buffer was aspirated and the nonspecific sites were blocked by 1-h incubation at room temperature with 0.25 ml of PBS containing 5% goat serum, 1% BSA, and 0.1% gelatin (blocking medium). The medium was aspirated and replaced with 50 μl of blocking medium containing control serum or a 1:25 dilution of mAb against Sec6 and Sec8, 1:1,000 dilution of pAb against Man II, 1:100 dilution of pAb recognizing ZO1 and occludin, 1:20 dilution of pAb against IP3R2, and 1:200 dilution of mAb against IP3R3. After incubation with the primary Ab overnight at 4°C and three washes with the incubation buffer (same as blocking buffer, but without serum), the Abs were detected with goat anti–rabbit or anti–mouse IgG tagged with fluorescein or rhodamine. Images were collected with a BioRad MRC 1200 confocal microscope.
Figure 9.

Effect of latrunculin of localization of Sec8 in pancreatic acini. Pancreatic acini were treated with 100 μM LAT for 30 min at 37°C. The cells were homogenized and used to prepare a cytosolic (Sup) and a particulate fraction (pellet) to analyze the translocation of Sec6 and Sec8 from the membranes to the cytosol (left). Control acini and acini treated with LAT were also fixed and stained with phalloidin (red) and Sec8 (green).
Statistics
When appropriate, results are given as the mean ± SEM of the indicated number of experiments. Statistical significance was evaluated by a two way ANOVA. All immunostaining experiments were repeated at least five times with similar results. The effect of anti-Sec6 and Sec8 mAbs on agonist-dependent Ca2+ signaling was tested in at least four experiments at each Ab concentration. The effect of anti-Sec6 and Sec8 mAbs on 2,4,5 IP3-dependent Ca2+ oscillations was tested in three experiments. In all experiments the Abs had no effect on IP3-dependent Ca2+ oscillations.
Results
Localization of the Sec6/8 Complex in Pancreatic Acini
As a first step in understanding the possible role of the mammalian Sec6/8 complex in somatic, polarized epithelial cells, we determined its localization in freshly isolated pancreatic acini. Fig. 1 shows immunofluorescence images obtained with an mAb raised against Sec6. Sec6 was expressed in a confined region distal to the nucleus and proximal to the apical membrane. It appears to form a crown around each cell. Sec8 showed similar localization (see below).
Figure 1.

Localization of Sec6 in pancreatic acini. Isolated pancreatic acini were fixed and stained with mAb specific for Sec6. The right shows a high magnification view of the upper acinus in the left.
The pattern of Sec6 staining resembles that of the Golgi network in pancreatic acinar cells (Komhoff et al. 1994). Therefore, we compared the localization of Sec8 with the Golgi marker Man II by double staining with an mAb raised against Sec8 and a pAb recognizing Man II. Fig. 2 shows that although the localization of Sec8 and Man II is similar, there are also noticeable differences in their localization. First, Man II appearance is elongated and tubular, whereas that of Sec8 is smoother. Second, Man II localization is more perinuclear than that of Sec8, resulting in a rim of Man II around the Sec8 (arrows in Fig. 2, top). Furthermore, treatment of the acini with 0.67 μM BFA significantly disrupted the localization of Man II with little effect on the localization of Sec8 (not shown). Even when BFA concentration was increased tenfold to 6.78 μM (Fig. 2, middle images), localization of Sec8 remained largely unaltered. Only treatment with 19.37 μM BFA considerably reduced staining with the anti-Sec8 Abs (Fig. 2, bottom images). Although Sec6 and Sec8 are predicted to be soluble proteins (Finger and Novick 1998; Hsu et al. 1999), >90% of the cellular pool of the two proteins is associated with the particulate fraction. Treatment with 19.37 μM BFA did not cause the translocation of Sec6 or Sec8 to the cytosol (Fig. 2, top blots). Finally, IP of βCOP effectively IP βCOP, but did not coimmunoprecipitate Sec8 from pancreatic or brain extracts and IP of Sec8 did not coimmunoprecipitate βCOP (Fig. 2, bottom blots). Similarly, IP of the Golgi 58K protein and Man II did not coimmunoprecipitate Sec8, and IP of Sec8 did not coimmunoprecipitate the Golgi 58K protein or Man II (not shown). Hence, although the staining pattern of the Sec6/8 complex appears similar to the Golgi apparatus, the complex does not appear to interact or localize with the Golgi apparatus. Significant portion of the complex must therefore be associated with another cellular compartment.
Figure 2.
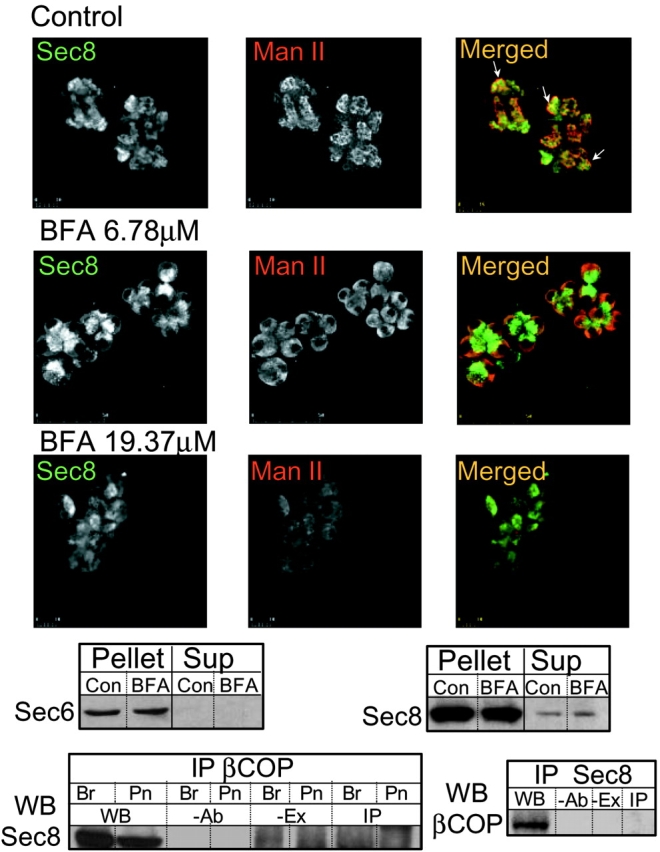
Localization of Sec8 and Man II in control and BFA-treated pancreatic acini. Acini were treated with buffer or the indicated concentration of BFA for 20 min at 37°C before plating on cover slips, fixation, and staining with mAb recognizing Sec8 (left, green) and pAb recognizing Man II (middle, red). The merged images show that Sec8 and Man II localization did not completely coincide in control or BFA-treated cells. In many cells, Man II formed a rim around Sec8 (arrows in top right). Note that localization of Man II was more sensitive to treatment with BFA than that of Sec8. The panels below the images show Western blot (WB) analysis of Sec6 (left) and Sec8 (right) in the particulate (pellet) and cytoplasmic (Sup) fraction in control (con) acini and acini treated with 19.37μm BFA (BFA). The bottom left shows IP of βCOP from brain (Br) or pancreatic (Pn) extracts and probing for Sec8. The bottom right shows IP of Sec8 from brain extract and probing for βCOP. Here and in all subsequent IP experiments, WB donates extract samples used for WB, -Ab donates control IP using extract without Ab in the IP reaction, -Ex donates control IP using Ab without extract in the IP reaction, and IP donates IP using extract and Ab in the IP reaction.
After polarization of MDCK cells and formation of a tight monolayer, the Sec6/8 complex relocates to the TJ. Therefore, it was of interest to determine the localization of the Sec6/8 complex with relation to TJ in the native, polarized pancreatic acini. Staining the acini for the TJ resident proteins ZO1 and occludin was used to follow TJ localization. Fig. 3 clearly shows that in pancreatic acini, the Sec6/8 complex is excluded from the TJ. Furthermore, imaging the cells at the x/z direction reveals that the Sec6/8 complex is expressed in a shape of a basket at the luminal pole. Thus, when following the acinus marked by an arrow, it can be seen that next to the plasma membrane, (first image, marked by B for bottom) the Sec8 is expressed only next to the luminal membrane. ZO1 is not seen since the TJ is at a higher cellular plane. About 2.5 μm up (image M1 for middle 1) Sec8 expression now extends all the way to the perinuclear region and ZO1 begins to appear. At the next 2.5 μm (image M2), Sec8 is expressed only at the perinuclear region and ZO1 is fully observed. At image M3 (2.5 μm up from M2) Sec8 staining is observed beyond the perinuclear region, but not yet at the most luminal region and ZO1 is in a more restricted area. In image M4, which is 7.5 μm up from image M1, Sec8 and ZO1 expression is similar to that in image M1. In the topmost image, Sec8 expression is similar to that in image B.
Figure 3.
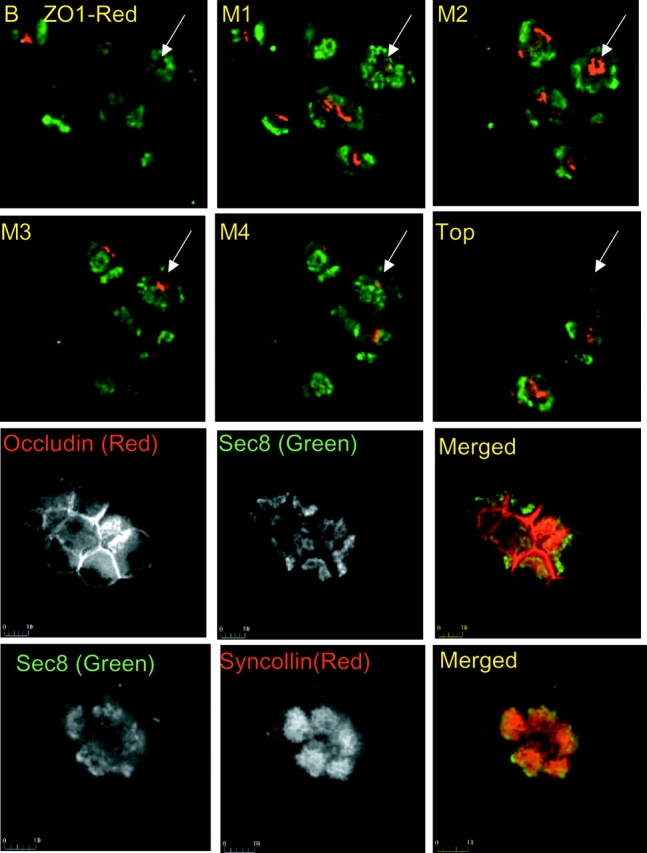
Localization of Sec8, ZO1, occludin, and syncollin in pancreatic acini. The top six panels show an x/z series of acini stained for Sec8 (green) and ZO1 (red). The images were recorded at 2.5-μm intervals in the z direction from bottom (B) to the top. The basket shape of Sec8 expression can be seen by following the acinus marked by an arrow. Acini stained with Sec8 (green) and occludin (red) or syncollin (red) are also shown.
Secretory granules are present in the region where the Sec6/8 complex is expressed. However, the bottom images of Fig. 3 show that expression of the Sec6/8 complex starts at a region proximal to secretory granules. In addition, Western blot analysis of partially purified secretory granules showed that Sec6 and Sec8 did not purify with the granules and were mostly enriched in a heavy microsomal fraction (not shown). Thus, if the Sec6/8 complex participates in polarized zymogen granules budding out of the Golgi apparatus, it does not remain associated with the granules after their maturation.
Ca2+ Signaling Proteins and the Sec6/8 Complex
The restricted region in which the Sec6/8 complex is expressed in pancreatic acini is of particular interest since it overlaps with the region containing the most energized mitochondria (Tinel et al. 1999), IP3R with the highest sensitivity to activation by IP3 (Kasai et al. 1993; Thorn et al. 1993), the sites from which [Ca2+]i waves are launched (Kasai et al. 1993; Thorn et al. 1993; Xu et al. 1996; Lee et al. 1997a) and the site of expression of a high level of several proteins involved in Ca2+ transport and Ca2+ signaling (Lee et al. 1997a,Lee et al. 1997b). Therefore, we tested the possibility that the Sec6/8 complex colocalizes and regulates Ca2+ signaling complexes. Fig. 4 shows localization of Sec8 and the IP3R2. High expression levels of IP3R2 are found in two regions: just underneath the luminal, and along the lateral membrane, and at the apical pole starting at the perinuclear region (best seen in top left). Expression of the Sec6/8 complex closely overlaps with expression of IP3R2 next to the plasma membrane at the apical pole. However, the localization of Sec8 and IP3R2 did not overlap next to the luminal membrane in the middle of the cells (Fig. 4, merged image).
Figure 4.
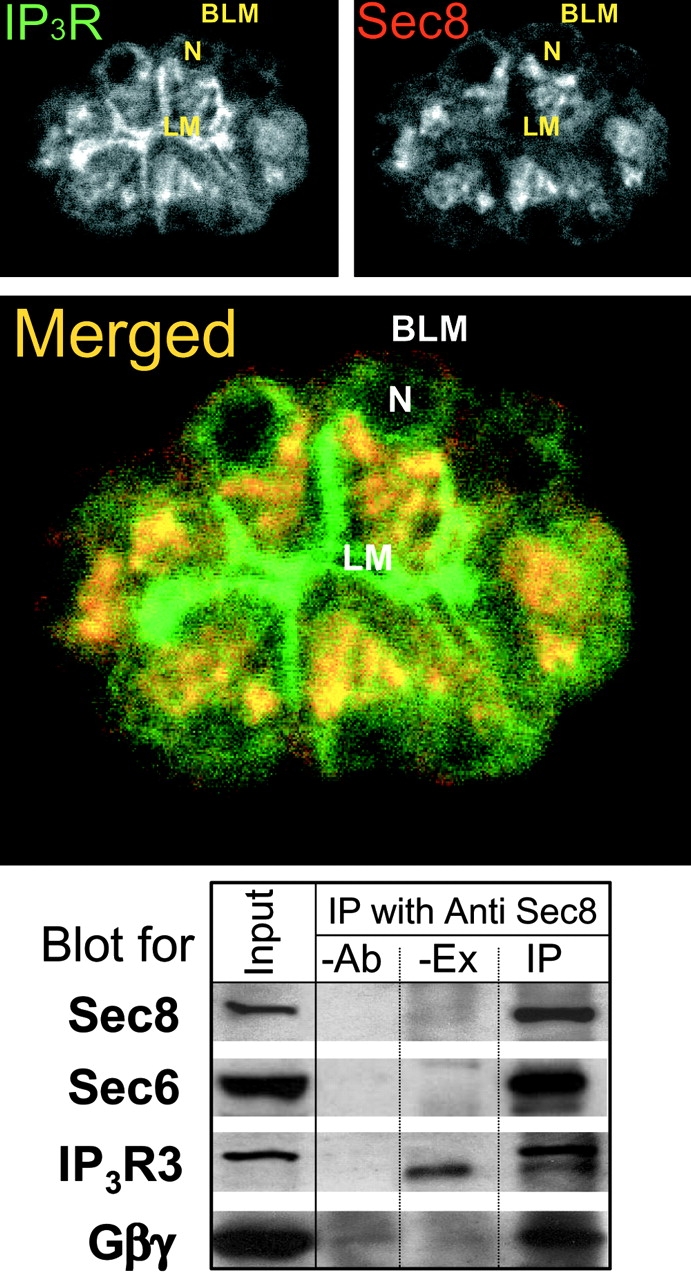
Localization of Sec8 and IP3R2. The images show localization of IP3R2 (top left, green), Sec8 (top right, red), and their colocalization (merged image). BLM donates basolateral membrane, AM donates apical membrane, and N donates the nucleus. In the blot, extract prepared from pancreatic membranes (Input) was used for IP with anti-Sec8 Abs. Controls were either -Ab or -Ex. The immunoprecipitated proteins were blotted for Sec8, Sec6, IP3R3, and Gβγ. The Ab recognizing IP3R3 also detected a nonspecific band in pancreatic acinar cell membrane extracts (lanes -EX and IP). This band was observed in all experiments (n = 4) and was always below the IP3R3 band.
Coimmunoprecipitate studies showed that the Sec6/8 complex not only colocalizes, but also interacts with IP3Rs and other signaling proteins. Fig. 4 shows the results obtained in pancreatic acini. IP of Sec8 coimmunoprecipitate Sec6, IP3R3, and Gβγ. Although the signal/noise ratio made it difficult to demonstrate with consistency the coimmunoprecipitate of Sec8 with other signaling proteins in pancreatic acinar cell membranes, the results in Fig. 4 strongly suggest that the Sec6/8 complex is associated (directly or indirectly) with Ca2+ signaling complexes. To extend this finding, we performed similar coimmunoprecipitate experiments with a brain microsomal extract, a rich source of Ca2+ signaling proteins. The results are illustrated in Fig. 5. IP of Sec8 coimmunoprecipitates Sec6, the PMCA, the β1 isoform of phospholipase C (PLCβ1), IP3R1, which is the isoform enriched in brain ER, Gβγ, and the α subunit Gαq (Fig. 5, left). Furthermore, the right of Fig. 5 shows that IP of PMCA, IP3R1, PLCβ1, or Sec8 coimmunoprecipitate Sec8 and all other signaling proteins. As a negative control we found that IP of Sec8 did not coimmunoprecipitate the Ca2+ binding synaptic vesicle protein synaptotagmin (ST).
Figure 5.

IP of Sec8 and Ca2+ signaling proteins from a brain microsomal extract. An extract was prepared from brain microsomes and used for IP of Sec8 (left), PMCA, IP3R1, PLCβ1, and Sec8 (right). The immunoprecipitates were analyzed by Western blots for the proteins indicated to the left of each. Additional control is provided by showing that IP of Sec8 did not coimmunoprecipitate the Ca2+ sensor synaptotagmin (ST).
The fraction of the Ca2+ signaling proteins in brain extract that was precipitated by Sec8 was estimated as follows. A sample of the extract was used to measure the amount of each protein in the lysate. From the band intensity we calculated that the Sec8 Ab precipitated 19.1 ± 2.2% of the Sec8 present in the extract. This resulted in coimmunoprecipitate of 10.6 ± 0.35% of Sec6. Hence, either not all cellular Sec6 is complexed with Sec8 or cell extraction partially dissociates the Sec6/8 complex. IP of Sec8 coimmunoprecipitate 5.0 ± 0.3% of PMCA, 5.9 ± 1.5% of PLCβ1, 8.1 ± 0.5% of IP3R1, 3.5 ± 0.6% of Gβγ, and 1.2 ± 0.2% of Gαq present in the extract. Thus, the stoichiometry of Sec8 to that of most Ca2+ signaling proteins in the immunoprecipitated complex was ∼3.3:1. That is, as much as 20% of the cellular pool of Ca2+ signaling proteins can be associated with the Sec6/8 complex at a time. This calculation suggests that the Sec6/8 complex interacts with a substantial fraction of the cellular Ca2+ signaling proteins. Therefore, it is clear that part of the Sec6/8 complex interacts with Ca2+ transport and signaling proteins that reside in (PMCA), are attached to (PLCβ1, Gαq, or Gβγ) the plasma membrane, or are resident of the ER (IP3Rs). The Ca2+ transport and signaling proteins associated with the Sec6/8 complex may form the Ca2+ signaling complexes that initiate the Ca2+ signal in the apical pole of pancreatic acini (Muallem and Wilkie 1999). However, considering the large fraction of cellular Ca2+ signaling proteins bound to the complex, the Sec6/8 complex may also participate in delivery of Ca2+ signaling proteins to their final destination.
The Sec6/8 Complex and Ca2+ Signaling
The anti-Sec8 mAbs were shown to inhibit delivery of vesicles to the basolateral membrane in MDCK cells (Grindstaff et al. 1998). Therefore, we reasoned that if the Sec6/8 complex interacts with Ca2+ signaling complexes, the anti-Sec8 mAb should inhibit Ca2+ signaling. This was tested by infusing the Abs into the cells through a patch pipette and measuring the activity of the Ca2+-dependent Cl− channel as a reporter of [Ca2+]i next to the plasma membrane (Zeng et al. 1996). Fig. 6 a shows that stimulation of pancreatic acinar cells infused with 85 μg/ml of a nonrelevant IgG fraction (control) with 0.5 μM carbachol induced typical [Ca2+]i oscillations. The same cell responded with a sustained response when stimulated with 1 mM carbachol or 10 nM cholecystokinin (CCK). Infusing the cell with 13 μg/ml of the anti-Sec8 mAb 17A10 abolished the [Ca2+]i oscillations evoked by weak carbachol stimulation and converted the sustained response induced by intense carbachol stimulation to an oscillatory response (Fig. 6 b). Increasing the concentration of the anti-Sec8 mAb 17A10 to 53 μg/ml further inhibited the responses to intense stimulation by carbachol or CCK (Fig. 6c). Another anti-Sec8 mAb, 5C3, had similar effects when used at 8.4 μg/ml and 28 μg/ml, respectively (Fig. 6d and Fig. e).
Figure 6.

Inhibition of Ca2+ signaling by anti-Sec8 and anti-Sec6 Abs. Pancreatic acinar cells were infused through a patch pipette with nonrelevant IgG (a and f) or the indicated concentrations of anti-Sec8 Abs 17A10 (b and c), 5C3 (d and e), or anti-Sec6 Abs (g–i) for at least 700 s before the first stimulation with 0.5 μM carbachol. As indicated by the bars, the cells were also treated with 1 mM carbachol, 10 μM atropine (Atr), or 10 nM cholecystokinine (CCK). The figure shows the Ca2+-activated Cl− current under the specified conditions.
The effect of the anti-Sec6 mAb is illustrated in Fig. 6f–i. Again, as much as 210 μg/ml of a nonrelevant IgG had no effect on the [Ca2+]i signals observed with weak or intense stimulation (Fig. 6 f). On the other hand, infusion of anti-Sec6 mAb dose-dependently inhibited Ca2+ signaling (Fig. 6, g–i). To identify the step in Ca2+ signaling inhibited by the Abs we then determined the effect of anti-Sec6 and Sec8 mAb on [Ca2+]i oscillations induced by infusion of IP3 into the cells. Fig. 7 a shows that infusion of 10 μM of the nonmetabolized IP3 analogue inositol 2,4,5-trisphosphate, induced Ca2+ oscillations similar to those evoked by weak agonist stimulation. A subsequent intense stimulation with 1 mM carbachol induced a large [Ca2+]i increase. Infusion of 72 μg/ml of anti-Sec6 (Fig. 7 b), 82 μg/ml of anti-Sec8 5C3 (Fig. 7 c), or 53 μg/ml of anti-Sec8 17A10 (Fig. 7 d) had no effect on the oscillations evoked by 2, 4, 5-IP3, but nearly abolished the response of the same cells to intense stimulation by 1 mM carbachol. These results clearly show that the Abs inhibited Ca2+ signaling by acting on a step upstream of activation of the IP3R by IP3. This step is probably the generation of IP3 by the Ca2+ signaling complexes.
Figure 7.

The anti-Sec6 and anti-Sec8 Abs do not inhibit IP3-mediated Ca2+ release. Pancreatic acinar cells were infused with 10 μM of 2-, 4-, or 5-IP3 alone (a), or together with the indicated concentrations of anti-Sec6 (b), anti-Sec8 5C3 (c), or anti-Sec8 17A10 (d). At the time indicated by the bars, the cells were stimulated with 1 mM carbachol.
The Actin Cytoskeleton, the Sec6/8 Complex and Ca2+ Signaling
Interaction of the Sec6/8 complex with Ca2+ signaling proteins can be direct or the complex can be recruited to Ca2+ signaling complexes to regulate their activity. In yeast, the components of the exocyst are recruited to the landmark established by Sec3 (Finger et al. 1998). The actin cytoskeleton is essential for recruitment of the exocyst (Ayscough et al. 1997; Finger et al. 1998). The mammalian Sec6/8 complex strongly interacts and copurifies with the septins (Hsu et al. 1998). These findings suggest a possible role of the actin cytoskeleton in mediating the interaction of the Sec6/8 complex with Ca2+ signaling complexes. In the present work, we used several protocols to show that this is indeed the case.
In the first protocol shown in Fig. 8, we determined whether actin filaments are needed for interaction of the Sec6/8 complex with Ca2+ signaling proteins. In these experiments, we first coimmunoprecipitate the Sec8 and the Ca2+ signaling proteins. The immunoprecipitate was then treated with buffer (control) or 20 μg/ml of an NH2-terminal fragment of gelsolin. This fragment rapidly severs actin filaments in a Ca2+-independent manner (Sun et al. 1999). After washing, the proteins remained attached to the beads were analyzed by Western blot. Fig. 8 shows that severing actin filaments dissociated most of the actin from the complex, but had no effect on the binding of Sec6 to Sec8 in the immunoprecipitate. By contrast, severing actin filaments dissociated between the Sec6/8 complex and all the Ca2+ signaling proteins, PMCA, PLCβ1, IP3R1, and Gβγ.
Figure 8.

Disassembly of actin filaments dissociates between the Sec6/8 complex and Ca2+ signaling proteins. Brain extract was used to immunoprecipitate proteins with anti-Sec8 Abs. After IP, the Sepharose A beads were divided into two portions. One portion was treated with buffer (con) and one portion with 20 μg/ml of the NH2-terminal fragment of gelsolin (Gelsol) for 20 min at 0°C. The beads were then washed before analysis of proteins remaining attached to the beads by Western blots. Note that Sec6 remained bound to Sec8, but actin and all Ca2+ signaling proteins were released as a consequence of actin disassembly.
The findings in Fig. 8 indicate that the Sec6/8 complex is recruited to Ca2+ signaling complexes and their interaction is mediated by actin filaments. To examine whether actin filaments mediate the interaction between the complexes in intact pancreatic acinar cells, and the possible significance of this interaction, we tested the effect of latrunculin B (LAT) on the localization of the Sec6/8 complex and the details of Ca2+ signaling. LAT dissociates actin filaments in intact cells by binding and scavenging monomeric actin (Coué et al. 1987). In yeast, LAT disrupted actin filaments and consequently the localization of the exocyst to the bud tip (Ayscough et al. 1997; Finger et al. 1998). Fig. 9 shows that LAT had similar effect in pancreatic acini. Western blot analysis reveals that treatment of pancreatic acini with 100 μM LAT resulted in partial translocation of Sec6 and Sec8 from the particulate fraction to the cytosol. The amount of Sec6 and Sec8 associated with the particulate fraction was reduced by an average of 51 ± 11% (n = 4) and 47 ± 7% (n = 4), respectively. Consequently, localization of the complex in intact cells appeared more cytosolic and less localized (Fig. 9, bottom).
The effects of LAT treatment on the properties of agonist-evoked Ca2+ signals are shown in Fig. 10 and Fig. 11. Fig. 10 shows an example of the images of individual experiments and Fig. 11 is the summary of multiple experiments. In pancreatic acinar (Kasai et al. 1993; Thorn et al. 1993; Nathanson et al. 1994; Xu et al. 1996; Lee et al. 1997b) and other secretory cells (Elliott et al. 1992; Lee et al. 1997b), agonist stimulation triggers a polarized [Ca2+]i wave that is initiated in the luminal pole, propagates along the cell periphery to the basal pole, and then collapses into the cytosol and the nucleus. The top of Fig. 10 shows such a [Ca2+]i wave induced by carbachol stimulation. Treating the cells with LAT had a profound effect on all properties of the [Ca2+]i wave. The images in the bottom of Fig. 10 and the summary in Fig. 11 show that disruption of actin filaments with LAT had minimal effect on the extent of Ca2+ release and [Ca2+]i increase in the luminal pole, but markedly reduced the rate of Ca2+ release. LAT treatment also reduced the [Ca2+]i wave speed by a factor of three and propagation of the wave from the luminal to the basal poles. In many LAT-treated cells, a stable luminal to basal Ca2+ gradient was attained before the reduction of [Ca2+]i by PMCA.
Figure 10.
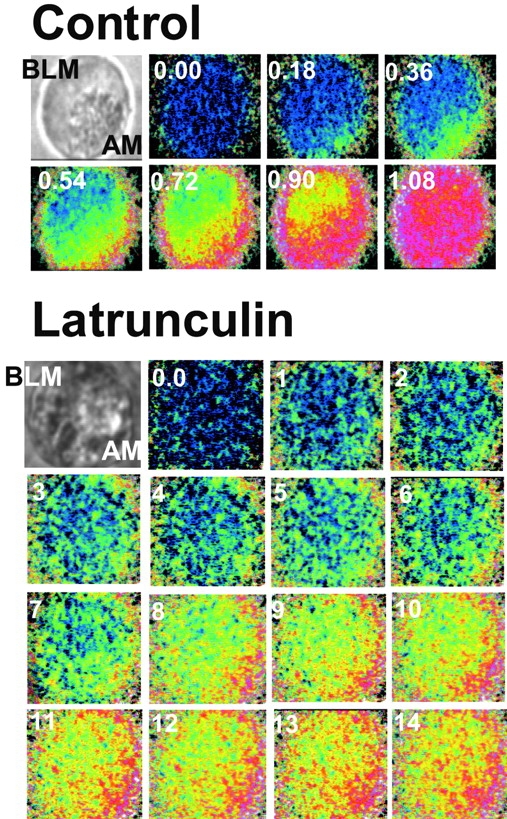
Effect of LAT on agonist-evoked [Ca2+]i waves. Fura2-loaded pancreatic acinar cells were treated with buffer (control) or 100 μM LAT for 30 min at 37°C before stimulation with 1 mM carbachol. Images were recorded at a resolution of 90 msec/image. Selective images recorded at the indicated times (in sec) are shown in pseudocolor. The images in top left of each series are the bright-field images. The secretory granules can be seen in the apical pole. BLM, Basolateral membrane; AM, apical membrane.
Figure 11.

Summary of effect of LAT on agonist-evoked [Ca2+]i waves. A shows the time-dependent change in Fura2 fluorescence ratio in control cells and cells treated with LAT. B shows the effect of LAT on the extent of [Ca2+]i increase in the apical (AP) and basolateral (BLP) poles (left), wave speed (middle), and rate of Ca2+ release (right). The results are the average of 19 control experiments and 16 experiments with cells treated with LAT.
Discussion
The role of the yeast exocyst in establishment of cell polarity and vesicles delivery to site of cell growth is well established (Finger and Novick 1998). The mammalian homologue of the exocyst, the Sec6/8 complex, also plays a role in establishment of cell polarity in developing neurons and epithelial cells in culture (Hsu et al. 1999). In the present work, we examined the possible role of the Sec6/8 complex in pancreatic acinar cells, differentiated, polarized epithelial cells. Our results suggest that the Sec6/8 complex may have more than one function in pancreatic acinar cells. It may control the polarized delivery of cargo, which includes Ca2+ signaling proteins to the plasma membrane, and it interacts with and regulates the activity of [Ca2+]i signaling complexes.
The localization of the Sec6/8 complex in pancreatic acini is different from that reported in neurons and MDCK cells (Grindstaff et al. 1998; Hazuka et al. 1999). The complex was expressed in the apical pole, but was excluded from the TJ and did not interact with secretory granules. It showed similar, but not identical, localization with the Golgi apparatus. A higher concentration of BFA was needed to disrupt the localization of the Sec6/8 complex than the localization of the Golgi marker Man II. The Golgi resident proteins Man II, βCOP, and Golgi 58K protein, and Sec8 did not coimmunoprecipitate. Hence, despite similar apparent colocalization, the Sec6/8 in the luminal pole does not reside in the Golgi apparatus. The basket-shape expression of the Sec6/8 complex in this region (Fig. 3) suggests that it is associated with part of the ER and the lateral membrane close to the TJ. This portion of the Sec6/8 complex is not likely to regulate Ca2+ signaling. Thus, treatment with BFA did not result in translocation of Sec6 or Sec8 from cellular membranes to the cytosol and had no apparent effect on the Ca2+ signal evoked by any of the agonists acting on pancreatic acini. Considering the role of the exocyst and the Sec6/8 complex in polarized vesicle delivery (Finger and Novick 1998; Hsu et al. 1999) and the interaction of the Sec6/8 complex with a large portion of the cellular Ca2+ signaling proteins pool, it is conceivable that the Sec6/8 complex in the luminal pole is involved in polarized delivery of Ca2+ signaling proteins. The finding that dissociation of actin filaments in intact cells not only disrupted Ca2+ signaling, but also the localization of Sec8 in the luminal pole supports this interpretation.
A novel function of the Sec6/8 complex suggested by our work is the regulation of Ca2+ signaling complexes activity. The fraction of the Sec6/8 complex adjacent to the plasma membrane likely interact with signaling complexes since IP of Sec8 coimmunoprecipitate proteins embedded (PMCA) or tightly attached (PLCβ1, G protein subunits) to the plasma membrane. It is interesting that IP of Sec8 also coimmunoprecipitates part of the ER resident IP3Rs. Recent work showed that IP3Rs in the ER gate the activity of store-operated Ca2+ channels in the plasma membrane (Kiselyov et al. 1998; Kiselyov et al. 1999). This gating is probably mediated by IP3Rs expressed in a subcompartment of the ER that contains the Ca2+ pool responsible for gating of store-operated channels (Broad et al. 1999; Krause et al. 1999). It is possible that IP of Sec8 coimmunoprecipitates this population of IP3Rs that is part of the Ca2+ signaling complex in the plasma membrane (Muallem and Wilkie 1999).
The Sec6/8 complex proteins are predicted to be soluble proteins, yet they mostly associate with cellular membranes in neurons (Hazuka et al. 1999), differentiated MDCK cells (Grindstaff et al. 1998), and pancreatic acini (present work). How this is achieved is not clear at present. However, our work suggests that the actin cytoskeleton plays a critical role in this association. In yeast, disruption of the actin cytoskeleton inhibited the polarized accumulation of exocyst proteins (Ayscough et al. 1997; Finger et al. 1998). In pancreatic acini, dissociation of the actin cytoskeleton in intact cells disrupted the localization of the Sec6/8 complex and resulted in translocation of ∼50% of the complexes from the membrane to the cytosol. Notably, dissociation of actin filaments also caused the dissociation between the Sec6/8 complex and Ca2+ signaling proteins. This finding implies that the Sec6/8 and Ca2+ signaling complexes do not interact directly, but rather the Sec6/8 complex is recruited to Ca2+ signaling complexes with the aid of the actin cytoskeleton. This may occur while the Sec6/8 complex delivers the Ca2+ signaling proteins from the ER and the Golgi apparatus to their destination in plasma membrane microdomains.
The interaction of the Sec6/8 complex with Ca2+ signaling complexes appears to modulate Ca2+ signaling in polarized cells. Abs against both Sec6 and Sec8 inhibited Ca2+ signaling by acting at a step upstream of Ca2+ release by IP3. In addition, disruption of the actin cytoskeleton in intact cells reduced the rate of Ca2+ release and interfered with propagation of [Ca2+]i waves. Disassembly of the actin cytoskeleton can affect the localization of many cytosolic and membrane-associated proteins and thus disrupt Ca2+ signaling independent of its effect on the localization of the Sec6/8 complex. Nonetheless, the effect of actin filament disassembly on the interaction of the Sec6/8 and Ca2+ signaling complexes in vitro and in vivo is consistent with its effect on Ca2+ signaling. Another important implication of these experiments is that interaction of the Sec6/8 complex with Ca2+ signaling proteins is not obligatory for initiation of Ca2+ release. The Sec6/8 complex is required for efficient Ca2+ signaling to allow maximal Ca2+ release and propagation of Ca2+ waves. The anti-Sec6 and anti-Sec8 Abs completely inhibited Ca2+ signaling, probably since their presence in the Sec6/8-Ca2+ signaling complexes prevented proper functioning of Ca2+ signaling.
Regulation of cell signaling by the Sec6/8 complex is probably not restricted to mammalian cells. The potential for such regulation also exists in yeast. During yeast mating, the nucleotide exchange factor Cdc24 and Far1 are exported together out of the nucleus and targeted to polarized sites of cell growth (Nern and Arkowitz 2000; O'Shea and Herskowitz 2000; Shimada et al. 2000). At these sites, Far1 binds Gβγ and a signaling complex is assembled with the aid of the scaffolding protein, Bem1 (O'Shea and Herskowitz 2000; Shimada et al. 2000). The exocyst is likely to deliver vesicles to these sites of cell growth. It might then interact with the pheromone signaling complex to regulate its activity.
In summary, our results suggest that in polarized cells, the Sec6/8 complex is present at least in two sites, at the luminal pole and next to the plasma membrane, in association with Ca2+ signaling complexes at specific plasma membrane domains. The two pools of the Sec6/8 complex are likely to participate in at least two cellular activities: protein delivery and regulation of Ca2+ signaling. The activities may be related in that the same Sec6/8 complex that established the polarized expression of Ca2+ signaling proteins in secretory cells may also regulate their activity. Testing of such a model requires a better understanding of the function of individual subunits of the Sec6/8 complex.
Footnotes
Abbreviations used in this paper: Ab, antibody; BFA, brefeldin A; IP, immunoprecipitation; IP3, inositol 1,4,5-trisphosphate; IP3R, IP3 receptor; LAT, latrunculin B; Man II, mannosidase II; pAb, polyclonal antibody; PMCA, plasma membrane Ca2+; TJ, tight junctions.
References
- Ayscough K.R., Stryker J., Pokala N., Sanders M., Crews P., Drubin D.G. High rates of actin filament turnover in budding yeast and roles for actin in establishment and maintenance of cell polarity revealed using the actin inhibitor latrunculin-A. J. Cell Biol. 1997;137:399–416. doi: 10.1083/jcb.137.2.399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowser R., Müller H., Govindan B., Novick P. Sec8p and Sec15p are components of a plasma membrane-associated 19.5S particle that may function downstream of Sec4p to control exocytosis. J. Cell Biol. 1992;118:1041–1056. doi: 10.1083/jcb.118.5.1041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broad L.M., Armstrong D.L., Putney J.W., Jr. Role of the inositol 1,4,5-trisphosphate receptor in Ca2+ feedback inhibition of calcium release-activated calcium current Icrac. J. Biol. Chem. 1999;274:32881–32888. doi: 10.1074/jbc.274.46.32881. [DOI] [PubMed] [Google Scholar]
- Coué M., Brenner S., Spector I., Korn E. Inhibition of actin polymerization by Latrunculin A. FEBS Lett. 1987;213:316–318. doi: 10.1016/0014-5793(87)81513-2. [DOI] [PubMed] [Google Scholar]
- Elliott A.C., Cairns S.P., Allen D.G. Subcellular gradients of intracellular free calcium concentration in isolated lacrimal acinar cells. Pflugers Arch. 1992;422:245–252. doi: 10.1007/BF00376209. [DOI] [PubMed] [Google Scholar]
- Finger F.P., Hughes T.E., Novick P. Sec3p is a spatial landmark for polarized secretion in budding yeast. Cell. 1998;92:559–571. doi: 10.1016/s0092-8674(00)80948-4. [DOI] [PubMed] [Google Scholar]
- Finger F.P., Novick P. Spatial regulation of exocytosislessons from yeast. J. Cell Biol. 1998;142:609–612. doi: 10.1083/jcb.142.3.609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grindstaff K.K., Yeaman C., Anandasabapathy N., Hsu S.C., Rodriguez-Boulan E., Scheller R.H., Nelson W.J. Sec6/8 complex is recruited to cell–cell contacts and specifies transport vesicle delivery to the basal–lateral membrane in epithelial cells. Cell. 1998;93:731–740. doi: 10.1016/s0092-8674(00)81435-x. [DOI] [PubMed] [Google Scholar]
- Guo W., Grant A., Novick P. Exo84p is an exocyst protein essential for secretion. J. Biol. Chem. 1999;274:23558–23564. doi: 10.1074/jbc.274.33.23558. [DOI] [PubMed] [Google Scholar]
- Hazuka C.D., Foletti D.L., Hsu S.C., Kee Y., Hopf F.W., Scheller R.H. The sec6/8 complex is located at neurite outgrowth and axonal synapse-assembly domains. J. Neurosci. 1999;19:1324–1334. doi: 10.1523/JNEUROSCI.19-04-01324.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu S.C., Ting A.E., Hazuka C.D., Davanger S., Kenny J.W., Kee Y., Scheller R.H. The mammalian brain rsec6/8 complex. Neuron. 1996;17:1209–1219. doi: 10.1016/s0896-6273(00)80251-2. [DOI] [PubMed] [Google Scholar]
- Hsu S.C., Hazuka C.D., Roth R., Foletti D.L., Heuser J., Scheller R.H. Subunit composition, protein interactions, and structures of the mammalian brain sec6/8 complex and septin filaments. Neuron. 1998;20:1111–1122. doi: 10.1016/s0896-6273(00)80493-6. [DOI] [PubMed] [Google Scholar]
- Hsu S.C., Hazuka C.D., Foletti D.L., Scheller R.H. Targeting vesicles to specific sites on the plasma membranethe role of the sec6/8 complex. Trends Cell Biol. 1999;9:150–153. doi: 10.1016/s0962-8924(99)01516-0. [DOI] [PubMed] [Google Scholar]
- Kasai H., Li Y.X., Miyashita Y. Subcellular distribution of Ca2+ release channels underlying Ca2+ waves and oscillations in exocrine pancreas. Cell. 1993;74:669–677. doi: 10.1016/0092-8674(93)90514-q. [DOI] [PubMed] [Google Scholar]
- Kiselyov K., Xu X., Mozhayeva G., Kuo T., Pessah I., Mignery G., Zhu X., Birnbaumer L., Muallem S. Functional interaction between InsP3 receptors and store-operated Htrp3 channels. Nature. 1998;396:478–482. doi: 10.1038/24890. [DOI] [PubMed] [Google Scholar]
- Kiselyov K., Mignery G.A., Zhu M.X., Muallem S. The N-terminal domain of the IP3 receptor gates store-operated hTrp3 channels. Mol. Cell. 1999;4:423–429. doi: 10.1016/s1097-2765(00)80344-5. [DOI] [PubMed] [Google Scholar]
- Komhoff M., Hollinshead M., Tooze J., Kern H.F. Brefeldin A induced dose-dependent changes to Golgi structure and function in the rat exocrine pancreas. Eur. J. Cell. Biol. 1994;63:192–207. [PubMed] [Google Scholar]
- Krause E., Schmid A., Gonzalez A., Schulz I. Low cytoplasmic Ca2+ activates ICRAC independently of global Ca2+ store depletion in RBL-1 cells. J. Biol. Chem. 1999;274:36957–36962. doi: 10.1074/jbc.274.52.36957. [DOI] [PubMed] [Google Scholar]
- Lee M.G., Xu X., Zeng W., Diaz J., Kuo T.H., Wuytack F., Racymaekers L., Muallem S. Polarized expression of Ca2+ pumps in pancreatic and salivary gland cells. Role in initiation and propagation of [Ca2+]i waves J. Biol. Chem. 272 1997. 15771 15776a [DOI] [PubMed] [Google Scholar]
- Lee M.G., Xu X., Zeng W., Diaz J., Wojcikiewicz R.J., Kuo T.H., Wuytack F., Racymaekers L., Muallem S. Polarized expression of Ca2+ channels in pancreatic and salivary gland cells. Correlation with initiation and propagation of [Ca2+]i waves J. Biol. Chem. 272 1997. 15765 15770b [DOI] [PubMed] [Google Scholar]
- Muallem S., Wilkie T.M. G protein-dependent Ca2+ signaling complexes in polarized cells. Cell Calcium. 1999;26:173–180. doi: 10.1054/ceca.1999.0077. [DOI] [PubMed] [Google Scholar]
- Nathanson M.H., Fallon M.B., Padfield P.J., Maranto A.R. Localization of the type 3 inositol 1,4,5-trisphosphate receptor in the Ca2+ wave trigger zone of pancreatic acinar cells. J. Biol. Chem. 1994;269:4693–4696. [PubMed] [Google Scholar]
- Nern A., Arkowitz R.A. Nucleocytoplasmic shuttling of the Cdc42p exchange factor Cdc24p. J. Cell Biol. 2000;148:1115–1122. doi: 10.1083/jcb.148.6.1115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Shea E.K., Herskowitz I. The ins and outs of cell-polarity decisions. Nat. Cell Biol. 2000;2:E39–E41. doi: 10.1038/35004065. [DOI] [PubMed] [Google Scholar]
- Rios J.D., Zoukhri D., Rawe I.M., Hodges R.R., Zieske J.D., Dartt D.A. Immunolocalization of muscarinic and VIP receptor subtypes and their role in stimulating goblet cell secretion. Invest. Ophthalmol. Vis. Sci. 1999;40:1102–1111. [PubMed] [Google Scholar]
- Shimada Y., Gulli M.-P., Peter M. Nuclear sequestration of the exchange factor Cdc24 by Far1 regulates cell polarity during yeast mating. Nat. Cell Biol. 2000;2:117–124. doi: 10.1038/35000073. [DOI] [PubMed] [Google Scholar]
- Sun H.Q., Yamamoto M., Mejillano M., Yin H.L. Gelsolin, a multifunctional actin regulatory protein. J. Biol. Chem. 1999;274:33179–33182. doi: 10.1074/jbc.274.47.33179. [DOI] [PubMed] [Google Scholar]
- TerBush D.R., Novick P. Sec6, Sec8, and Sec15 are components of a multisubunit complex which localizes to small bud tips in Saccharomyces cerevisiae . J. Cell Biol. 1995;130:299–312. doi: 10.1083/jcb.130.2.299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- TerBush D.R., Maurice T., Roth D., Novick P. The exocyst is a multiprotein complex required for exocytosis in Saccharomyces cerevisiae . J. Cell Biol. 1996;15:6483–6494. [PMC free article] [PubMed] [Google Scholar]
- Thorn P., Lawrie A.M., Smith P.M., Gallacher D.V., Petersen O.H. Local and global cytosolic Ca2+ oscillations in exocrine pancreas. Cell. 1993;74:661–668. doi: 10.1016/0092-8674(93)90513-p. [DOI] [PubMed] [Google Scholar]
- Tinel H., Cancela J.M., Mogami H., Gerasimenko J.V., Gerasimenko O.V., Tepikin A.V., Petersen O.H. Active mitochondria surrounding the pancreatic acinar granule region prevent spreading of inositol trisphosphate-evoked local cytosolic Ca2+ signals. EMBO (Eur. Mol. Biol. Organ.) J. 1999;18:4999–5008. doi: 10.1093/emboj/18.18.4999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu X., Zeng W., Diaz J., Muallem S. Spacial compartmentalization of Ca2+ signaling complexes in pancreatic acini. J. Biol. Chem. 1996;271:24684–24690. doi: 10.1074/jbc.271.40.24684. [DOI] [PubMed] [Google Scholar]
- Yule D.I., Ernst S.A., Ohnishi H., Wojcikiewicz R.J. Evidence that zymogen granules are not a physiologically relevant calcium pool. Defining the distribution of inositol 1,4,5-trisphosphate receptors in pancreatic acinar cells. J. Biol. Chem. 1997;272:9093–9098. doi: 10.1074/jbc.272.14.9093. [DOI] [PubMed] [Google Scholar]
- Zeng W., Xu X., Muallem S. Gβγ transduces [Ca2+]i oscillations and Galphaq a sustained response during stimulation of pancreatic acinar cells with [Ca2+]i-mobilizing agonists. J. Biol. Chem. 1996;271:18520–18526. doi: 10.1074/jbc.271.31.18520. [DOI] [PubMed] [Google Scholar]