

Abstract
Willow (Salix babylonica L.) is representative of a large group of plants that have extensive plasmodesmatal connections between minor vein phloem and adjoining cells. Because plasmodesmata provide a diffusion pathway for small molecules, it is unclear how sucrose could be loaded from the mesophyll into the phloem against a concentration gradient. In the studies reported here, the minor vein phloem of willow leaves plasmolyzed in approximately the same concentration of osmoticum as the mesophyll. Sucrose concentrations in mesophyll cells were greater than those reported in the literature for aphid stylet exudate from willow stems. Calculated turgor pressures in the mesophyll and minor vein phloem were greater than turgor reported in the literature for sieve elements in the stems of willow. Images of minor veins were not obtained in autoradiographs when attached leaves, or leaf pieces, were provided with 14CO2 or [14C]sucrose. Therefore, no evidence could be found for accumulation of sucrose against a concentration gradient in the minor vein phloem of willow. In these leaves, the mesophyll apparently acts as the “source” for long distance transport of sugar. The mechanism of translocation in willow, and the evolution of phloem loading, are discussed.
Phloem loading has long been considered an essential component of the mechanism of translocation (1). Although Münch (2) did not include a loading step in his pressure-flow model of long distance transport, by the 1940’s, there was general agreement that a metabolically active pump must be operating. This consensus followed the publication of several studies indicating that the solute content of the phloem is higher than that of surrounding cell types. Especially influential were the papers of Phillis and Mason (3), Huber et al. (4), and Roeckl (5). The term phloem “loading” was coined by Loomis (6), who used it specifically to describe an “endothermic pumping action.” Since that time, the term “loading” has been used to denote an active step that concentrates photoassimilate in the phloem, increasing turgor and thus steepening the pressure gradient between source and sink.
To maintain this steep concentration gradient, the sieve element-companion cell complex (SE-CCC) is isolated symplastically from surrounding cells along the conducting (long distance) pathway (7–9). This is necessary because small molecules diffuse through plasmodesmata. If extensive symplastic leakage occurred, the sugar gradient would dissipate.
However, in the minor veins of mature leaves, where phloem loading occurs, the situation is more complex. The frequency of plasmodesmata joining the SE-CCCs of minor veins to surrounding cells varies greatly in different species. Because the SE-CCC is the site of phloem loading (10) and symplastic connections to sieve elements from outside the complex are very rare, the plasmodesmata leading into the companion cell from adjacent parenchyma or bundle sheath cells are of particular importance. In many species, these plasmodesmata are few in number, but in others, they are extremely abundant. Gamalei (11, 12) has referred to these systems as “closed” (even though some plasmodesmata are present), and “open,” respectively. The degree of symplastic continuity between the SE-CCC and surrounding cells is often a family trait. Thirty percent of the families of dicotyledonous plants studied by Gamalei have members with open phloem in the minor veins (Type 1), 40% have closed phloem (Type 2a-c), and the remaining 30% have plasmodesmatal frequencies between the two groups (Type 1–2a) (11).
The presence of an extensive diffusive pathway between the SE-CCC and surrounding cells—open minor vein phloem—seems paradoxical if the phloem is devoted to establishing and maintaining high solute levels. Nonetheless, it has been shown that sugar accumulates against a steep concentration gradient in the open minor vein phloem of the cucurbits (13, 14) and Coleus blumei (15). These plants apparently load by a “polymer trap” mechanism in which raffinose and stachyose are synthesized in the phloem and are retained there on the basis of size (refs. 1 and 16; see Discussion).
Although this mechanism may explain loading in some plants with open phloem, it does not seem to apply to many others because the only sugar they translocate in quantity is sucrose (17). These are primarily woody plants found in temperate climates. Indeed, most of the tree species in the temperate forest fall into this category. In such plants, the mechanism of phloem loading remains obscure, if it occurs at all. In the study reported here, several approaches were used to analyze sugar accumulation in the minor veins of willow, a species with open phloem (11) that translocates only traces of raffinose and stachyose (18). No evidence for phloem loading was found.
MATERIALS AND METHODS
Plant Material.
Experiments were conducted on mature leaves exposed to full sun and taken from large, open-grown weeping willow trees (Salix babylonica L.) on the bank of Cayuga Lake in a City of Ithaca park. Some experiments were conducted in the field; in others, branches were cut under the surface of water and were brought to the laboratory within 20 min.
Plasmolysis.
Plasmolysis studies were carried out in each of three growing seasons. The conditions and protocols were essentially the same in all experiments, and the results were virtually identical. The study reported in detail here was conducted on May 29, 1997. The weather had been warm and sunny with intermittent rain for the previous 2 weeks. The experiment was begun at ≈15:00 h. The temperature was 21°C. Lamina pieces (1 mm2) were excised under 40 mM Pipes (1, 4-piperazinediethanesulfonic acid)–NaOH buffer plus 4 mM CaCl2 (pH 7.2). The tissue pieces were randomized, and subsamples were submerged for 20 min in one of a graded series of sorbitol solutions ranging from 200 mM to 1,400 mM, in steps of 100 mM. The subsamples then were fixed in 2% glutaraldehyde in 25 mM Pipes–NaOH buffer, with sufficient added sorbitol to achieve the same total osmolarity as the previous solution (19). After overnight fixation at room temperature, the tissue was washed in buffer, was post-fixed in 1% OsO4 at room temperature for 2 h, was dehydrated in a graded ethanol series, and was embedded in Spurr’s resin for electron microscopy (20). A cell was considered to be plasmolyzed if one quarter or more of the length of the plasmalemma in a section was pulled away from the cell wall (21).
Quantitative Sugar Analysis.
Lamina discs (7.0 mm in diameter) were removed with a cork borer, were frozen in powdered dry ice, were ground in liquid N2, and were extracted at 50°C in methanol/chloroform/water (12/5/3; vol/vol/vol). An aqueous phase was obtained by adding additional water, followed by centrifugation, and this phase was passed through anion and cation exchange resins to obtain the neutral fraction. The neutral fraction was analyzed by high-performance liquid chromatography using a Waters SugarPak column. Details of these procedures have been described elsewhere (22).
Morphometric Analysis.
Volume determinations were made by point-counting methods (23) by using a random array of points (24). Dot density was 1.8 dots⋅cm−2, and electron micrographs were at 2,240× magnification. Each sample consisted of a series of overlapping electron micrographs spanning ≈40 contiguous cells. See ref. 14 for details.
14CO2-Labeling.
A clear plastic bag was placed over an attached branch, enclosing up to eight mature leaves, and the bag was sealed securely. To produce radiolabeled gas, 0.5 MBq 14CO2 was generated in a syringe from Na214CO3 (6.6 × 105 MBq⋅mmol−1) by the addition of excess 80% lactic acid, and the gas then was injected into the bag. The bag was removed 5 min later. Long distance transport was monitored continuously by placing a Geiger–Müller counter against the shoot apex. Leaves were removed at periodic intervals and were frozen in powdered dry ice for autoradiographic analysis (see below).
[14C]Sucrose Uptake into Leaf Discs.
The adaxial surfaces of leaves were abraded or removed, as described in Results. Discs (7.0 mm in diameter) were cut from the leaves under the surface of 2(N-morpholino)ethanesulfonic acid (Mes) buffer (20 mM Mes plus 2 mM CaCl2, pH 5.5, with NaOH). Discs were randomized and transferred, abraded side down, to the surface of fresh buffer in small (3.5 cm diameter) plastic Petri dishes. The buffer was removed and replaced with 3 ml of solution containing 20 mM Mes buffer and [14C]sucrose (1 mM; 15 kBq⋅ml). Discs were incubated at room temperature for 1 h with continuous agitation on a shaker, were washed three times for 10 min each in Mes buffer at room temperature, and were frozen in powdered dry ice.
For autoradiography, leaf discs were lyophilized in a −30°C chamber, were pressed thin and flat between steel plates in a vice, were mounted on cardboard, and were pressed against x-ray film (Hyperfilm-βmax, Amersham). For storage, lyophilized tissue was kept in a desiccator over drying agent. Details of these procedures are given by Weisberg et al. (25).
RESULTS
Plasmolysis.
The mesophyll and SE-CCCs of mature willow leaves plasmolyzed in approximately the same concentration of osmoticum. The plasma membrane was not pulled away from the wall in more than a few cells of either tissue when leaf samples were incubated in 900 mM sorbitol. In 1.0 M sorbitol, 53% of the SE-CCCs and 47% of mesophyll cells were plasmolyzed. In 1.1 M sorbitol, all of the cells of the mesophyll and phloem were plasmolyzed.
Leaf Sugar Concentrations.
Because plasmolysis data indicated high levels of solute in the mesophyll of willow leaves, we measured sugars in leaf samples (Table 1). The amount of sucrose was >10× that found in mature melon leaves, per unit leaf area (14). Much lower amounts of raffinose and stachyose were present, and the amounts of monosaccharides were variable but well below those of sucrose.
Table 1.
Concentrations of sugars in mature willow leaves
| Sugar | Concentration, nmol⋅mm−2 ± SE* |
|---|---|
| Sucrose | 16.0 ± 4.3 |
| Raffinose | 0.18 ± 0.05 |
| Stachyose | trace |
| Glucose | 2.1 ± 1.1 |
| Fructose† | 5.4 ± 2.9 |
SE, Standard error. Leaves were sampled five times over two growing seasons. Each sample consisted of five subsamples, each from a different leaf.
Combined fructose and myo-inositol, which were not resolved on the column.
To calculate sugar concentrations (see Discussion), we used morphometric procedures to determine the volume of leaf compartments. The total volume of the leaf was 0.198 mm3⋅mm−2 leaf area. Because sucrose is most likely to be compartmentalized in cytosol and vacuoles, the volume fractions of these two compartments was determined. Pooling data from all cell types, 7.0 ± 0.2% (standard error; n = 4) of leaf volume was cytosol (not including chloroplasts) and 22.7 ± 0.7% (standard error; n = 4) was vacuole.
Autoradiography.
To visualize photoassimilate accumulation in veins, branches that were still attached to the tree were provided with 14CO2 for 5 min, and leaves were removed 0, 25, and 55 min after the end of the presentation period. Radiolabel was detected in the shoot apex within 45 min after exposure to 14CO2, indicating that translocation was taking place. The leaves were flash-frozen, were lyophilized, were compressed to improve resolution, and were pressed against x-ray film. No images of minor veins were detected (Fig. 1A; white indicates the presence of radiolabel). In similar experiments, accumulated radiolabel in the minor veins of C. blumei was visualized readily (Fig. 1B).
Figure 1.

Autoradiographs of (A) willow and (B) C. blumei leaves exposed to 14CO2 for 5 min and allowed to transport for a further 55 min. Leaves were flash frozen and lyophilized. Autoradiographs were used as negatives; therefore, white indicates the presence of radiolabel. (Bar = 2 mm.)
Greater contrast between veins and mesophyll in vein loading images is sometimes achieved by starving leaves for carbohydrate (26). Therefore, several attached willow branches were sheathed in aluminum foil for 3 days. These branches then were exposed to light for 20 min to allow the stomata to open, and 14CO2 experiments were performed. Again, no autoradiographic vein images were seen (not shown).
In many studies of phloem loading, it has been shown, by the autoradiographic technique, that minor veins accumulate exogenous [14C]sucrose (1). However, when excised discs from willow leaves were floated on [14C]sucrose solution, no vein images were seen in the autoradiographs; labeling was uniform across the discs (Fig. 2). Because the epidermis of willow is thick, three methods were used to eliminate this barrier to the entry of [14C]sucrose: The adaxial surface was abraded with 320-grit carborundum powder to varying degrees, was scraped to different degrees with either 320- or 600-grit sandpaper, or was removed entirely with a razor blade. The results were the same in all cases: No vein images were seen in the autoradiographs (Fig. 2A, C, and D). When 2 mM p-chloromercuribenzenesulfonic acid (PCMBS) was added to the incubation solution, the density of labeling in the tissue noticeably decreased (Fig. 2B, compare with Fig. 2A).
Figure 2.
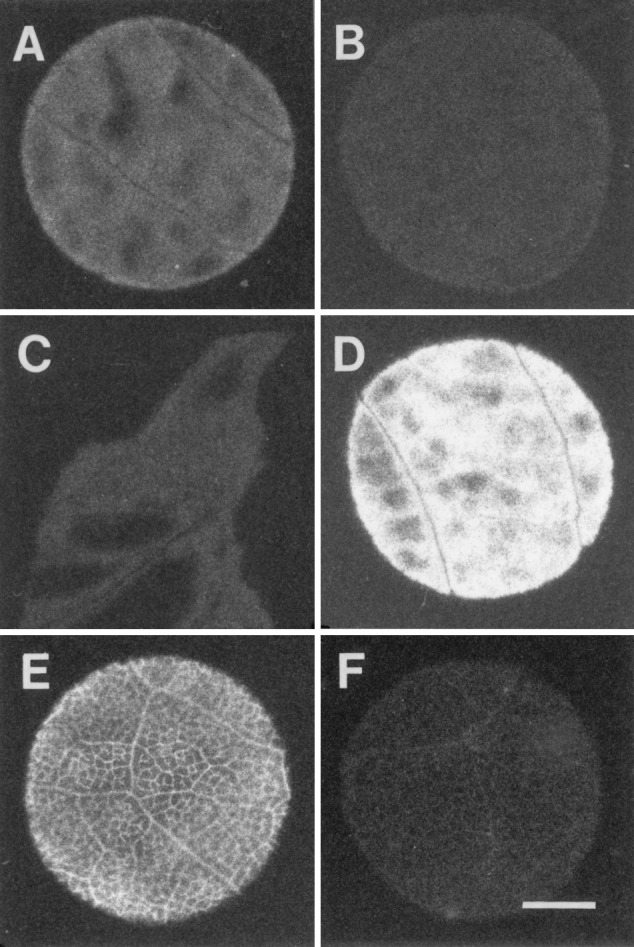
Autoradiographs of willow (A–D) and Nicotiana tabacum (E and F) leaf tissue floated for periods of time, as indicated, on [14C]sucrose solution ± PCMBS, washed in buffer, flash frozen, and lyophilized. All tissue was abraded with carborundum except C, in which the lower epidermis was removed with a razor blade. (A) 20 min, minus PCMBS. (B) 20 min, plus PCMBS. (C) 20 min, minus PCMBS. (D) 1 h, minus PCMBS. (E) 20 min, minus PCMBS. (F) 20 min, plus PCMBS. Autoradiographs were used as negatives; therefore, white indicates the presence of radiolabel. (Bar = 2 mm.)
In case phloem loading occurs only during a restricted period of the day or night, a series of [14C]sucrose incubations was conducted on leaf tissue collected at 4-h intervals over a 24-h period by using all abrasion methods. Again, no vein images were detected (not shown).
Tobacco leaf discs were used as a control. Veins were readily apparent in tobacco leaf discs floated on [14C]sucrose (Fig. 2E). As expected, PCMBS reduced uptake into tobacco leaf discs and prevented vein loading (Fig. 2F).
DISCUSSION
Phloem Loading Defined.
As used here, the term “phloem loading” refers only to energy-dependent accumulation of solutes in the SE-CCC, without regard to mechanism and exclusive of other transport steps within the mesophyll or phloem. This usage is a common one and is consistent with the initial intent of Loomis, who postulated an “endothermic pumping action” that elevates phloem sugar concentration (6). It also avoids the confusion that arises when other transport steps in the mesophyll or phloem, which might have entirely different mechanisms, are included. Defined in this simple way, polymer-trapping via the symplast, and carrier-mediated transport from the apoplast, are both mechanisms of phloem loading, even though the former does not involve a true membrane-based active transport component. By this definition, the minor veins of willow do not appear to engage in phloem loading, at least under the conditions in which the experiments reported here were performed.
Solute Levels in the Willow Leaf.
Several methods have been used to demonstrate phloem loading in minor veins. Of these methods, plasmolysis was the first (10), and it continues to be valuable because it estimates directly the osmotic potential of individual cells. In all reports to date (Fig. 3), with the exception of one (ref. 21; Fig. 3), the SE-CCCs of minor veins have been found to be especially resistant to plasmolysis; they remain, for the most part, unplasmolyzed in osmoticum >1.2 M (Fig. 3). Much lower concentrations of osmoticum severely shrink the protoplasts of the surrounding mesophyll, indicating that total solute levels in SE-CCCs are much higher than in the mesophyll.
Figure 3.

Plasmolysis in source leaves. Data are from the literature except for willow (this study). The molar concentration of osmoticum needed to induce incipient plasmolysis is plotted for both mesophyll cells and the SE-CCC of minor veins. ∗, Intermediary cells. #, ordinary companion cells. Reference citations are as follows: Moricandia (27), Coleus (28), Amaranthus (29), Squash (30), sugar beet (10), and Populus (21).
This difference in plasmolytic response between phloem and mesophyll is not found in leaves of willow (this study; Fig. 3), or Populus deltoides (ref, 21 and Fig. 3). In both species, the two tissue types plasmolyze in approximately the same concentration of osmoticum, below 1.2 M. Indeed, Russin and Evert (21) reported that the SE-CCCs of P. deltoides leaves plasmolyzed in slightly lower concentrations of osmoticum than required to plasmolyze the mesophyll (Fig. 3). Both willow and Populus are in the Salicaceae. They are Type 1 plants, according to Gamalei (11), and they translocate only small amounts of raffinose and stachyose (18). Consistent with the latter finding, we found little raffinose or stachyose in willow leaves (Table 1).
Willow and P. deltoides are the only two species examined to date by the plasmolysis technique that have open phloem and transport small amounts of raffinose and stachyose. They are also the only two species examined to date that do not demonstrate a characteristic difference in solute potential between the mesophyll and minor vein phloem.
It should be noted that Russin and Evert (21) found evidence for phloem loading within the minor veins of P. deltoides by comparing the osmotic behavior of the sieve elements and companion cells. The concentration of mannitol required to induce 50% plasmolysis in the sieve elements was calculated to be 0.57 M, higher than that needed to reach the same frequency of plasmolysis in the companion cells (0.50 M). However, this difference was less than the increments in osmoticum used in their experiments (0.1 M). Considering the possibility that there may be subtle uncertainties in the plasmolysis technique, it is not clear that this small difference is a reliable measure of active solute accumulation.
Autoradiographic Analysis of Sugar Accumulation.
Individual solute species are not identified in plasmolysis studies. Therefore, even though the solute potentials of the mesophyll and minor vein phloem are approximately the same in willow leaves, it is still possible that sucrose accumulates to higher concentrations in the latter tissue. In other words, it is still possible that phloem loading of sucrose takes place. If it does so, radiolabeled photoassimilate should accumulate in the venation after a leaf is presented with 14CO2 (1). No such vein labeling pattern was seen in autoradiographs at any time point, even though veins were clearly apparent in autoradiographs of C. blumei leaves. Prior starvation of attached leaves, by shading, did not alter this result.
When excised leaf tissue was presented with [14C]sucrose, radiolabel did not accumulate in the veins to a greater degree than in the mesophyll, as would be expected if the veins take up sucrose from the apoplast against a concentration gradient (31). [14C]sucrose uptake was diminished in the presence of PCMBS, a sulfhydryl-modifying compound that inhibits sucrose–proton cotransport (1, 32). Therefore, the cells of willow leaves are capable of active sucrose uptake, but this mechanism is not used to increase the amount of sucrose in the minor vein phloem. Eschrich and Fromm (33) were unable to detect vein loading of [14C]sucrose in Salix viminalis or in a number of other woody species.
Sucrose in the Willow Leaf.
If there is no active accumulation of sucrose in minor veins, sucrose may be entering the phloem by diffusion. To assess this possibility, it is necessary to determine whether there is sufficient sucrose in the mesophyll to create a diffusion gradient. The concentration of sucrose in mesophyll cells can be approximated simply by measuring the amount of this sugar in the leaf and assuming that it is evenly distributed in the mesophyll and veins. This assumption is reasonable because only a small fraction of the sucrose in the leaf could be contained in the SE-CCCs, which constitute only 1% or less of total leaf volume (14). For example, even if sucrose were loaded into the phloem to a concentration of 600 mM (60% of total molarity as measured by plasmolysis), the SE-CCCs would contain only 7.4% of the leaf sugar, assuming that they occupy 1% of total leaf volume. This is less than the statistical variance between samples.
Assuming even distribution throughout the mesophyll and veins, the concentration of sucrose in the mesophyll can be determined. Because the combined fraction of the mesophyll cytosol (7.0%) and vacuoles (22.7%) totals 29.7% of the entire leaf volume, and the amount of sucrose is 16.0 nmol⋅mm−2 leaf area, the concentration of sucrose in these two compartments is, by calculation, 271 mM. However, this value does not take subcellular compartmentation into account. In all plants examined by Heldt and his colleagues, sucrose in mesophyll cells is more concentrated in the cytosol than in the vacuoles. The percentage of mesophyll sucrose in the cytosol ranges from 26% in spinach (34) to 83% in tobacco (35). Although subcompartmentation of sucrose has not been analyzed in willow leaves, it is clear that an asymmetry in distribution between the cytosol and the vacuole on the order seen in other species would create higher, and potentially very high, concentrations of sugar in the mesophyll cytosol. Theoretically, if the fraction of sucrose in the mesophyll cytosol (13.9 nl⋅mm−2 leaf area) were 26%, 50%, or 83%, the concentration of sucrose would be 299 mM, 577 mM, or 923 mM, respectively. Using a similar approach, Moing et al. (36) concluded that sorbitol potentially could enter the phloem of peach leaves by diffusion, although studies with PCMBS suggested a carrier-mediated apoplastic loading pathway.
If sucrose diffuses through the mesophyll and into the phloem, there should be a concentration gradient along this path. Unless this gradient is counterbalanced by a reverse gradient of other solute species, it might be detected in plasmolysis studies. However, the concentration drop across plasmodesmata required to maintain reasonable diffusional flux along the path from mesophyll to phloem is only in the micromolar to millimolar range (37) and probably would be too small to be detected by the plasmolysis technique.
Turgor in the Phloem of Willow Stems.
Münch (2) did not include a loading step in his model of long distance phloem transport. He considered the mesophyll to be the “source” of the pressure gradient. If this is true in willow, one would expect the turgor pressure in the phloem of the stems, the “path phloem,” to be lower than in the mesophyll (and minor vein phloem). Assuming the same water potential measured for willow leaves by Wright and Fisher (−0.19 MPa; ref. 48), the turgor pressure of the mesophyll and minor vein phloem can be calculated from the value for osmotic potential measured in the present study by the plasmolysis technique. Because plasmolysis occurred in ≈1.0 M sorbitol, this corresponds to an osmotic pressure of 2.78 MPa (converting molarity to molality) and a turgor pressure of 2.59 MPa. This value is well above the turgor pressure in the path phloem of willow measured by Wright and Fisher (0.79 MPa; ref. 48) by using capillary manometers glued over exuding aphid stylets.
The low turgor value reported by Wright and Fisher implies that the sucrose concentration in the path phloem of willow is low. Indeed, a comparison of literature values for sucrose concentrations in aphid stylet exudate from the path phloem of various species indicates that this is the case (Fig. 4). Note that a wide range of values has been reported. The highest is 1.4 M in corn. Values for mature willow trees under normal physiological conditions are at the low end of the scale. The mean value for willow, from three laboratories, is 235 mM.
Figure 4.

Sucrose concentrations in exudate taken from severed aphid stylets. Data are from the literature. Reference citations are as follows: corn (38, 39), barley (40), spinach (41), peach (42), rice (43), wheat (44, 45), lettuce (46), clover (47), and willow (48–52). Data for willow from the Fisher laboratory (48–50) are shown as the mean and range of three studies.
The Mechanism of Phloem Transport in Willow.
Transport of photoassimilate can be broken down into short and long distance components. The most parsimonious explanation of the data presented here is that short distance movement of photoassimilate from mesophyll cells into the companion cells and sieve elements of the minor veins is entirely diffusional, without an active step. If this is correct, sucrose moves into the minor vein phloem as long as its concentration in the cytosol of mesophyll cells is sufficiently high to provide a gradient for diffusion. This condition is apparently met, given the high concentration of sucrose in mesophyll cells. Even if sucrose is uniformly distributed throughout the cytosol and vacuoles of mesophyll cells, the concentration would be as high as in aphid stylet exudate taken from stem phloem. It is unlikely that the cytosolic concentration of sucrose in mesophyll cells is this low, however, because in other species there is more sucrose in the cytosol than in the vacuoles.
At present, we do not have an accurate estimate of cytosolic sucrose in willow leaves. However, it is worth noting that the osmotic potential of mesophyll cells in willow leaves is quite high [but not as high as in mangroves (53) or other plants that tolerate salt stress]. Osmotic potential in the mesophyll of willow leaves indicates that the solute concentration in these cells is ≈400 mM higher than in the mesophyll of other plants that have been analyzed by the same plasmolysis technique (Fig. 3). This excess in the cytosol may be sucrose, in part or in whole, which would mean that the sucrose concentration in the cytosol of mesophyll cells could be considerably higher than in the sieve elements of the minor veins or the path phloem.
Long distance transport of photoassimilate from leaves to distant parts of the willow tree cannot occur by diffusion—the distances involved are far too great. Once the sugar has entered the sieve elements of the minor veins, it presumably is carried away in a mass flow of solution as described by Münch (2), the pressure being lower in the stem phloem than in the leaf phloem and water being provided by the xylem.
How is phloem transport controlled in willow? Because export rate is governed by the pressure differential between source and sink, the capacity of sink tissues to remove solute from the phloem must be an important governing factor, just as it is in the presence of phloem loading. However, source tissue in willow also may be able to influence the pressure difference. Because sucrose apparently enters the phloem by diffusion, it is possible that control is exerted at the mesophyll tonoplast: The cytosolic concentration of sucrose, and, therefore, the rate of diffusion into the phloem, may be governed by adjusting the distribution of sucrose between cytosol and vacuole.
The Evolution of Phloem Loading.
A taxonomic analysis of Gamalei’s data set led to the hypothesis that symplastic phloem loading in Type 1 plants is the ancestral condition and that specialization over time, in some groups but not others, led to the evolution of closed phloem and the apoplastic loading mechanism (12, 54). However, Type 1 plants are composed of at least two subgroups (17). Of, these, the smaller has received by far the most attention from physiologists. This subgroup is characterized by the translocation of large amounts of raffinose and stachyose, especially the latter, in addition to sucrose (22), and by a distinct anatomical specialization of minor vein companion cells. These specialized companion cells are called “intermediary cells” and are remarkably similar in appearance in the various families in which they have been described; they have dense cytoplasm, numerous small vacuoles, rudimentary plastids, and fields of highly branched plasmodesmata leading into them from the bundle sheath. It is from the study of these species that the polymer trap model arose (15, 16).
The only families that are known to have members with true intermediary cells are the Cucurbitaceae (55, 56), Lamiaceae (12, 28), Verbenaceae (12), Scrophulariaceae (22), and Oleaceae (11, 57), although there are several other candidate families that translocate high levels of raffinose oligosaccharides that have not been studied adequately. All of these known families are in derived subclasses of the dicotyledons; none are in the ancestral Magnoliidae (58).
Plants of the other subgroup have open minor vein phloem in the sense of extensive symplastic connections to surrounding cells, but they typically translocate only small amounts of raffinose and stachyose (17). Relatively few micrographs have been published on minor vein ultrastructure in these species; however, those that have been studied do not appear to have typical intermediary cells (21).
It is not possible to make convincing generalizations about the evolution of phloem loading based on such meager data. However, it is clear that those species for which we have the best evidence for phloem loading, either symplastic or apoplastic, are in derived rather than ancestral families. Therefore, it is possible that the ancestral condition in the angiosperms is the absence of phloem loading. If this is true, symplastic loading may have evolved by the up-regulation of raffinose and stachyose synthesis in minor vein companion cells, with the concomitant narrowing of plasmodesmata joining them to the bundle sheath, thus effecting a polymer trap. Apoplastic loading may have evolved by the recruitment of the ubiquitous sucrose–proton cotransport system for the purpose of elevating sucrose levels in the phloem. This type of loading would have been made more efficient by a reduction in plasmodesmatal numbers to reduce leakage. Because this is a highly selective system for sucrose transport, raffinose and stachyose synthesis may have become obsolete in plants with an apoplastic loading mechanism.
Whether these speculations on the evolution of phloem loading do or do not prove to have merit, more detailed attention should be paid to studies of short distance sugar movement in the leaves of woody plants. It should not be assumed that phloem loading is a universal phenomenon.
Acknowledgments
This work was supported by National Science Foundation Grant IBN-9603152 to R.T.
ABBREVIATIONS
- SE-CCC
sieve element-companion cell complex
- PCMBS
p-chloromercuribenzenesulfonic acid
References
- 1. Grusak M A, Beebe D U, Turgeon R. In: Photoassimilate Distribution in Plants and Crops: Source-Sink Relationships. Zamski E, Schaffer A A, editors. New York: Marcel Dekker; 1996. pp. 209–227. [Google Scholar]
- 2.Münch E. Die Stoffbewegungen in der Pflanze. Jena, Germany: Gustav Fischer; 1930. [Google Scholar]
- 3.Phillis E, Mason T G. Ann Bot (London) 1933;47:585–634. [Google Scholar]
- 4.Huber B, Schmidt E, Jahnel H. Tharandter Forstl Jahrb. 1937;88:1017–1050. [Google Scholar]
- 5.Roeckl B. Planta. 1949;36:530–550. [Google Scholar]
- 6.Loomis W E. In: Origin of Resistance to Toxic Agents. Sevag M G, Reid R D, Reynolds O E, editors. New York: Academic; 1955. pp. 99–121. [Google Scholar]
- 7.Ammerlaan A, Kempers R, van Bel A J E. J Exp Bot. 1996;47:1300. [Google Scholar]
- 8.Oparka K J, Prior D A M, Wright K M. J Exp Bot. 1995;46:187–197. [Google Scholar]
- 9.van Bel A J E, Kempers R. Planta. 1991;183:69–76. doi: 10.1007/BF00197569. [DOI] [PubMed] [Google Scholar]
- 10.Geiger D R, Giaquinta R T, Sovonick S A, Fellows R J. Plant Physiol. 1973;52:585–589. doi: 10.1104/pp.52.6.585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gamalei Y. Trees. 1989;3:96–110. [Google Scholar]
- 12.Gamalei Y. Trees. 1991;5:50–64. [Google Scholar]
- 13.Pristupa N A. Sov Plant Physiol. 1979;26:467–474. [Google Scholar]
- 14.Haritatos E, Keller F, Turgeon R. Planta. 1996;198:614–622. doi: 10.1007/BF00262649. [DOI] [PubMed] [Google Scholar]
- 15.Turgeon R, Gowan E. Plant Physiol. 1990;94:1244–1249. doi: 10.1104/pp.94.3.1244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Turgeon R. In: Recent Advances in Phloem Transport and Assimilate Compartmentation. Bonnemain J-L, Delrot S, Lucas W J, Dainty J, editors. Nantes, France: Ouest Editions; 1991. pp. 18–22. [Google Scholar]
- 17.Turgeon R. In: Carbon Partitioning and Source-Sink Interactions in Plants. Madore M A, Lucas W J, editors. Rockville, Maryland: Am. Soc. Plant Physiol.; 1995. pp. 195–203. [Google Scholar]
- 18.Zimmermann M H, Ziegler H. In: Encyclopedia of Plant Physiology, New Series: Transport in Plants 1. Phloem Transport. Zimmermann M H, Milburn J A, editors. New York: Springer; 1975. pp. 480–503. [Google Scholar]
- 19.Lackney V K, Sjolund R D. Plant Cell Environ. 1991;14:213–220. [Google Scholar]
- 20.Volk G M, Turgeon R, Beebe D U. Planta. 1996;199:425–432. [Google Scholar]
- 21.Russin W A, Evert R F. Am J Bot. 1985;72:1232–1247. [Google Scholar]
- 22.Turgeon R, Beebe D U, Gowan E. Planta. 1993;191:446–456. [Google Scholar]
- 23.Weibel E R. Stereological Methods: Practical Methods for Biological Morphometry. Vol. 1. New York: Academic; 1980. [Google Scholar]
- 24.Parkhurst D F. Am J Bot. 1982;69:31–39. [Google Scholar]
- 25.Weisberg L A, Wimmers L E, Turgeon R. Planta. 1988;175:1–8. doi: 10.1007/BF00402875. [DOI] [PubMed] [Google Scholar]
- 26.van Bel A J E, Gamalei Y V, Ammerlaan A, Bik L P M. Planta. 1992;186:518–525. doi: 10.1007/BF00198031. [DOI] [PubMed] [Google Scholar]
- 27.Beebe D U, Evert R F. Int J Plant Sci. 1992;153:61–77. [Google Scholar]
- 28.Fisher D G. Planta. 1986;169:141–152. doi: 10.1007/BF00392308. [DOI] [PubMed] [Google Scholar]
- 29.Fisher D G, Evert R F. Planta. 1982;155:377–389. doi: 10.1007/BF00394465. [DOI] [PubMed] [Google Scholar]
- 30.Turgeon R, Hepler P K. Planta. 1989;179:24–31. doi: 10.1007/BF00395767. [DOI] [PubMed] [Google Scholar]
- 31.Wimmers L E, Turgeon R. Planta. 1991;186:2–12. doi: 10.1007/BF00201491. [DOI] [PubMed] [Google Scholar]
- 32.Giaquinta R. Plant Physiol. 1976;57:872–875. doi: 10.1104/pp.57.6.872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Eschrich W, Fromm J. Physiol Plant. 1994;90:699–707. [Google Scholar]
- 34.Winter H, Robinson D G, Heldt H W. Planta. 1994;193:530–535. [Google Scholar]
- 35.Heineke D, Wildenberger K, Sonnewald U, Willmitzer L, Heldt H W. Planta. 1994;194:29–33. [Google Scholar]
- 36.Moing A, Carbonne F, Zipperlin B, Svanella L, Gaudillère J P. Physiol Plant. 1997;101:489–496. [Google Scholar]
- 37.Gunning B E S. In: Intercellular Communication in Plants: Studies on Plasmodesmata. Gunning B E S, Robards A W, editors. New York: Springer; 1976. pp. 203–227. [Google Scholar]
- 38.Weiner H, Blechschmidt-Schneider S, Mohme H, Eschrich W, Heldt H W. Plant Physiol Biochem (Paris) 1991;29:19–23. [Google Scholar]
- 39.Ohshima T, Hayashi H, Chino M. Plant Cell Physiol. 1990;31:735–737. [Google Scholar]
- 40.Winter H, Lohaus G, Heldt H W. Plant Physiol. 1992;99:996–1004. doi: 10.1104/pp.99.3.996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Riens B, Lohaus G, Heineke D, Heldt H W. Plant Physiol. 1991;97:227–233. doi: 10.1104/pp.97.1.227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Moing A, Carbonne F, Rashad M H, Gaudillère J P. Plant Physiol. 1992;100:1878–1884. doi: 10.1104/pp.100.4.1878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Fukumorita T, Chino M. Plant Cell Physiol. 1982;23:273–283. [Google Scholar]
- 44.Fisher D B, Gifford R M. Plant Physiol. 1986;82:1024–1030. doi: 10.1104/pp.82.4.1024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hayashi H, Chino M. Plant Cell Physiol. 1986;27:1387–1393. [Google Scholar]
- 46.Helden M V, Tjallingii W F, van Beck T A. J Chem Ecol. 1994;20:3191–3206. doi: 10.1007/BF02033720. [DOI] [PubMed] [Google Scholar]
- 47.Girousse C, Bonnemain J-L, Delrot S, Bournoville R. Plant Physiol Biochem (Paris) 1991;29:41–48. [Google Scholar]
- 48.Wright J P, Fisher D B. Plant Physiol. 1980;65:1133–1135. doi: 10.1104/pp.65.6.1133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wright J P, Fisher D B. Plant Physiol. 1983;73:1042–1047. doi: 10.1104/pp.73.4.1042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Fisher D B. Planta. 1983;159:529–533. doi: 10.1007/BF00409142. [DOI] [PubMed] [Google Scholar]
- 51.Canny M J. Ann Bot (New Series) 1961;25:152–167. [Google Scholar]
- 52.Mittler T E. J Exp Biol. 1958;35:74–84. [Google Scholar]
- 53.Medina E, Francisco M. Estuarine, Costal Shelf Sci. 1997;45:337–344. [Google Scholar]
- 54.van Bel A J E. Acta Bot Neerl. 1992;41:121–141. [Google Scholar]
- 55.Turgeon R, Webb J A, Evert R F. Protoplasma. 1975;83:217–232. [Google Scholar]
- 56.Schmitz K, Cuypers B, Moll M. Planta. 1987;171:19–29. doi: 10.1007/BF00395064. [DOI] [PubMed] [Google Scholar]
- 57.Flora L L, Madore M A. Planta. 1993;189:484–490. [Google Scholar]
- 58.Cronquist A. An Integrated System of Classification of Flowering Plants. New York: Columbia Univ. Press; 1981. [Google Scholar]