

Abstract
To investigate the role of β-catenin in mammary gland development and neoplasia, we expressed a stabilized, transcriptionally active form of β-catenin lacking the NH2-terminal 89 amino acids (ΔN89β-catenin) under the control of the mouse mammary tumor virus long terminal repeat. Our results show that ΔN89β-catenin induces precocious lobuloalveolar development and differentiation in the mammary glands of both male and female mice. Virgin ΔN89β-catenin mammary glands resemble those found in wild-type (wt) pregnant mice and inappropriately express cyclin D1 mRNA. In contrast to wt mammary glands, which resume a virgin appearance after cessation of lactation, transgenic mammary glands involute to a midpregnant status. All transgenic females develop multiple aggressive adenocarcinomas early in life. Surprisingly, the ΔN89β-catenin phenotype differs from those elicited by overexpression of Wnt genes in this gland. In particular, ΔN89β-catenin has no effect on ductal side branching. This suggests that Wnt induction of ductal branching involves additional downstream effectors or modulators.
Keywords: β-catenin, mammary gland, cyclin D1, cadherin, Wnt
Introduction
β-Catenin is a versatile multifunctional protein that is essential for stable cell–cell adhesion and signal transduction (McCrea et al. 1991; Butz et al. 1992; Gumbiner 1995). At the membrane, it facilitates cell–cell adhesion by connecting classical cadherins to the actin cytoskeleton via α-catenin (Cowin and Burke 1996). Several Wnt signaling cascades operate by inhibiting degradation of cytosolic β-catenin and promoting its nuclear entry (Nusse and Varmus 1992; Gumbiner 1995). Once in the nucleus, β-catenin forms a bipartite transcription factor with lymphoid enhancer factor (Lef)/T cell factor (Tcf) proteins (Behrens et al. 1996; Molenaar et al. 1996), and modulates the expression of an array of genes that encode proteins involved in transcription, cell cycle regulation, apoptosis, and matrix remodeling (see http://www.stanford.edu/rnusse/pathways/targets.html for a current list of target genes and references). Perturbation of β-catenin protein levels produces dramatic effects in both embryonic and adult tissues and has been found in many types of tumors (Funayama et al. 1995; Haegel et al. 1995; Gat et al. 1998; Harada et al. 1999; Polakis 1999).
The mammary gland provides an attractive model in which to study β-catenin's in vivo function, because the majority of the patterning and proliferation that constitute the development of this organ take place postnatally (Daniel and Silberstein 1987). Furthermore, mammary differentiation during pregnancy, lactation, and involution can be invoked cyclically. Primary mammary epithelial ducts are established before birth but remain quiescent until puberty (∼3 wk of age in the mouse), at which point ovarian hormones cause secondary branches to grow from club-shaped terminal end buds. The ducts extend, becoming progressively branched, and fill the fat pad at ∼12 wk of age. The gland then remains dormant until the hormonal stimulus of pregnancy causes lobuloalveolar structures to develop. These structures increase dramatically in size and number, synthesizing milk proteins during pregnancy and secreting them after birth. After weaning, the alveolar epithelium undergoes apoptosis, causing the mammary gland to involute and resume an appearance similar to that of the pubertal ductal tree (Daniel and Silberstein 1987).
Mammary development and subsequent mammary cycles are synchronized to the reproductive state of the animal via hormonal induction but are, nevertheless, locally patterned (Henninghausen and Robinson 1998). There is good reason to suspect that β-catenin plays a seminal role in this process, because it is central to the functions of cadherins, Wnts, and Lef-1. Each of these proteins have roles in mammary development and/or neoplasia. The E-cadherin gene is frequently mutated in lobular breast cancer, and P-cadherin null mice show precocious lobuloalveolar development (Berx et al. 1995; Radice et al. 1997). The mammary gland shows differential expression of several Wnt genes (Buhler et al. 1993; Gavin et al. 1990; Weber-Hall et al. 1994), and Wnt-1, -3, and -10b induce tumors when misexpressed in mammary epithelia (Lane and Leder 1997; Tsukamoto et al. 1988). Finally, Lef-1 null mice lack mammary glands, and thus derepression of genes regulated by Lef-1/β-catenin appears to be essential for mammary gland development (van Genderen et al. 1994).
To determine the effects of altering β-catenin cytosolic levels and transcriptional activity during postnatal mammary gland development, we generated transgenic mice expressing a constitutively active form of β-catenin under the control of the mouse mammary tumor virus long terminal repeat. Our results show that ΔN89β-catenin induces precocious lobuloalveolar development and differentiation in both male and female virgin mice. The endpoint for involution differs in transgenics and normals, and both virgin and breeding females develop multiple aggressive adenocarcinomas early in life.
Materials and Methods
Plasmid Construction and Generation of Transgenic Mice
A cDNA encoding a myc-tagged, stabilized form of Xenopus β-catenin, deleted in the first 89 amino acids, was generated by PCR using pCS2-XBC provided to us by Dr. Barry Gumbiner (Memorial Sloan-Kettering, New York, NY), as a template (McCrea et al. 1991). A 250-bp DNA fragment was amplified using forward oligonucleotide 529Fx-myc (ctc ggatcc gaattc gcc gcc ATG GAG CAA AAG CTT ATT TCT GAA GAG GAC TTG AGA GCC CAG CGA GTT CGT GCT), which contains BamHI and EcoRI restriction sites, Kozac's sequence, a sequence encoding the myc epitope (MEQKLISEEDL) followed by amino acids 90–96 of Xenopus β-catenin, and the reverse oligonucleotide 710R (GGG GAT TGCTCGAGT GGC CAA TTC), encompassing an internal XhoI site at base 448 of the coding region. The 727-bp BamHI-XhoI fragment of pCS2-XBC was replaced by the 237-bp BamHI-XhoI PCR fragment to generate pCS2-ΔN89BC. An EcoRI linker was introduced into the SnaBI site in the 3′ end of the pCS2-ΔN89BC polylinker region, permitting the 2,160-bp insert to be excised as an EcoRI fragment, and subcloned into the transgene cassette. The pMMTVEV transgene cassette, containing the mouse mammary tumor virus long terminal repeat and rabbit β-globin (RBG) sequences which provide intron, splicing, and polyadenylation signals, was provided by Dr. Lynn Matrisian (Vanderbilt University, Nashville, TN) (Witty et al. 1995). The 2,704-bp XhoI fragment containing these sequences was excised, linkered with NotI restriction sites, and cloned into pGEM5Z+ (Promega). This cassette permitted cloning of the 2,160-bp EcoRI fragment of pCS2-ΔN89BC into the EcoRI cloning site embedded in the RBG sequence, and excision of the entire 4,864-bp transgene MMTV-ΔN89β-catenin with NotI for injection. The PCR-generated regions and newly generated cloning sites of all clones were fully sequenced. The NotI fragment carrying the transgene was purified by electroelution, followed by dialysis and ethanol precipitation, and microinjected into the fertilized eggs of FVB/N mice (Taconic Farms) by the Skirball Transgenic Facility. MMTV-Wnt-1 transgenic mice on an FVB/N background were purchased from The Jackson Laboratory.
Southern Blot Analysis of Genomic DNAs
Genomic DNA was prepared as described (Charpentier et al. 2000). For Southern blot analysis, 10–20 μg of genomic DNA samples was digested with 80 U of EcoRI, separated by electrophoresis in a 1× TAE-0.8% agarose gel at 3 V/cm, and transferred to Nytran membranes (Schleicher & Schuell). Blots were prehybridized for 1 h at 42 °C in ULTRAhyb (Ambion, Inc.), then hybridized overnight at 42 °C by addition of 50 ng of the 2,160-bp EcoRI fragment of MMTV-ΔN89β-catenin, radiolabeled by random priming to a specific activity of 109 cpm/μg (ICN Biomedicals). Blots were washed twice for 5 min in 2× SSC, 0.1% SDS and twice for 15 min in 0.1× SSC, 0.1% SDS at 42 °C, and exposed to PhosphorImager screens (Molecular Dynamics). The approximate transgene copy number was determined by adding 20 pg (2 copies), 200 pg (20 copies), and 2,000 pg (200 copies) of MMTV–ΔN89β-catenin to genomic DNA from a nontransgenic mouse, and comparing relative intensities of hybridization. Differences in DNA loading were calculated by cohybridization with a gene fragment (a gift of Dr. Alex Joyner, New York University Medical School) specific for the two copy gene engrailed (Wurst et al. 1994).
Northern Analysis
Mammary glands were homogenized in denaturing solution with a polytron, and total RNA was prepared using the ToTally RNA kit (Ambion, Inc.). 10–20 μg RNA was electrophoresed in 1% phosphate-glyoxal agarose gels using the NorthernMax-Gly kit (Ambion, Inc.). The following cDNA probes were gifts: casein and K18 (Caroline Alexander, University of Wisconsin, Madison, WI), cyclin D1 (Richard Pestell, Albert Einstein College of Medicine, Yeshiva University, New York, NY), and c-myc (Anthony Brown, Strang Cancer Research Laboratory, The Rockefeller University, New York, NY).
Immunoprecipitation and Western Blot Analysis
Mammary glands from F1 transgenic and wild-type (wt) sex-matched littermates were cryosectioned, boiled in 200 μl of lysis buffer (25 mM Tris-HCl, pH 7.4, 3 mM EDTA, 150 mM NaCl, 1% SDS, 10 μg/ml aprotinin, 1 mM PMSF), sonicated twice at stop 6, and pelleted for 15 min at 10,000 g. Protein concentration of the supernatant was determined by the method of Bradford, using a bicinchronic acid protein assay kit (Pierce Chemical Co.). For immunoprecipitations, ∼500 μg protein in 200 μl of lysis buffer was processed for immunoprecipitation as described (Witcher et al. 1996), then boiled in Laemmli sample buffer and subjected to 7.5% SDS-PAGE. For Western blotting, 20-μg protein samples were boiled in 40 μl of Laemmli sample buffer, then separated by electrophoresis through 7.5% SDS-PAGE, electrophoretically transferred to Protran membranes (Schleicher & Schuell), and processed for Western blotting as described (Charpentier et al. 2000).
Indirect Immunofluorescence
Mammary glands from F1 transgenic and normal sex-matched littermates were removed, snap frozen in liquid nitrogen, and stored at −70°C. 5-μm frozen sections were attached to multiwell slides and processed for indirect immunofluorescence as described (Charpentier et al. 2000). The following antibodies were used: mouse monoclonals 9E10 anti-myc epitope (a gift of Dr. Harold Varmus, National Institutes of Health, Bethesda, MD); anti–β-catenin and anti-smooth muscle actin (Sigma-Aldrich). Rabbit polyclonal antibodies used were specific for β-catenin (Santa Cruz Biotechnology, Inc.), cytokeratin 14 (BAbco), casein (a gift of Dr. Margaret Neville, University of Colorado, Denver, CO), and ribophorin antibody (a gift of Dr. Gert Kreibich, New York University Medical School).
Histology, TUNEL Analysis, and Whole Mounts
Mammary glands were fixed overnight or longer in 10% phosphate-buffered formalin, embedded in paraffin, and sectioned. They were stained with hematoxylin and eosin for histological analysis, or processed for terminal deoxynucleotidyl transferase–mediated dUTP nick end labeling (TUNEL) using the Promega Fluorescein Apoptosis Detection System (Promega). For whole mount tissue preparations, mammary glands were processed as described (http://mammary.nih.gov/tools/histological/Histology/index.html#a1).
Results
MMTV-ΔN89β-Catenin Is Expressed in Luminal Epithelial Cells of the Mammary Gland
We generated a form of β-catenin deleted in the NH2-terminal 89 amino acids (ΔN89β-catenin) that retains cadherin/catenin binding, and has greater stability in the cytoplasmic pool (Munemitsu et al. 1996). ΔN89β-catenin was placed under the control of the mouse mammary tumor virus long terminal repeat (Fig. 1 A), which directs transgene expression to mammary and salivary glands in the very early stages of puberty, and is hormonally upregulated by progesterone during estrus and pregnancy (Witty et al. 1995). 12 mice showed integration of the transgene. F1 mice from founders 1, 3, and 5 expressed the transgene protein product ΔN89β-catenin, as detected by Western blotting of immunoprecipitations of mammary extracts with 9E10 antibody that specifically recognizes the NH2-terminal human c-myc epitope tag present on the transgene product, and does not react with endogenous mouse c-myc (Fig. 1 B). These lines of mice contained ∼50, 1, and 45 copies of the transgene (Fig. 1 C), and varied greatly in their levels of ΔN89β-catenin expression (Fig. 1 D), thus permitting dose responsiveness of the phenotype to be assessed. Expression of ΔN89β-catenin was detectable in 7-wk virgin mammary glands, maximal during pregnancy, and remained elevated after 10 d of involution (Fig. 1 E). ΔN89β-catenin caused proportional downregulation of endogenous β-catenin, and upregulation of endogenous E-cadherin. These effects were most obvious in the high expressing line 5 (Fig. 1 F). Immunofluorescent staining of frozen sections of mammary gland with an anti-myc antibody specifically detected ΔN89β-catenin along the intercellular borders of the epithelium of the mammary ducts, and detected alveoli in both the high expressing lines (Fig. 1 G) and the low expressing line 3 (Fig. 1H and Fig. I). No such staining was found in tissue sections from wt littermates (data not shown). Double immunofluorescence using anti-myc antibody to detect ΔN89β-catenin, and anti-smooth muscle actin to define the myoepithelial cells, confirmed that ΔN89β-catenin expression was restricted to the luminal epithelium (Fig. 1 J). As reported previously for other tissues, ΔN89β-catenin was not detected in the nucleus (Gat et al. 1998; Wong et al. 1998). Nevertheless, the upregulation of putative target genes (see below) suggests that sufficient ΔN89β-catenin is in the nucleus for it to be transcriptionally active in this tissue.
Figure 1.
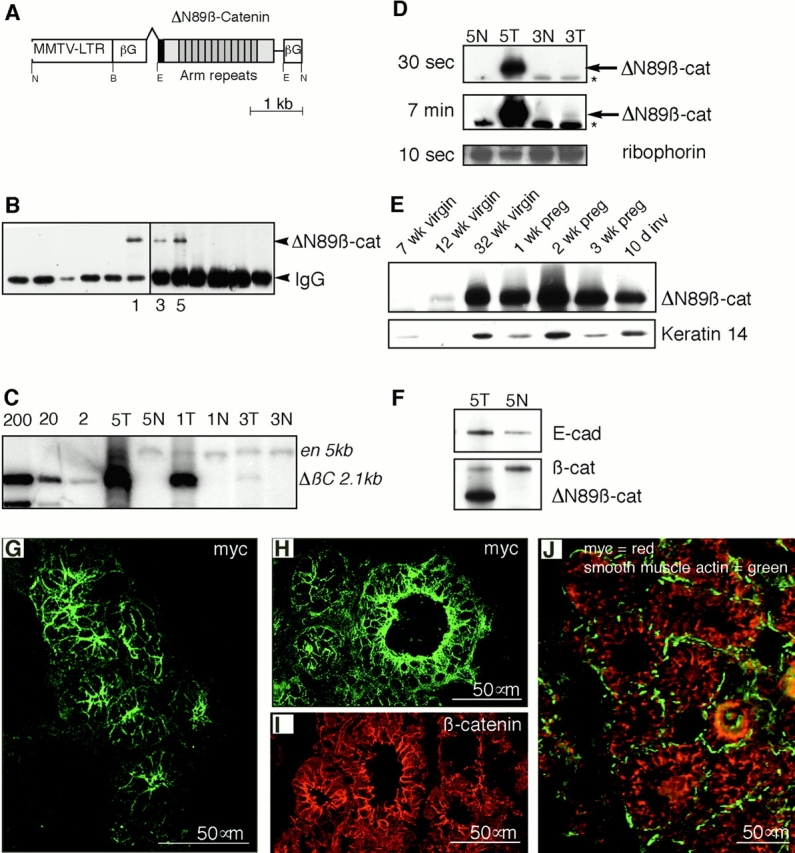
(A) Structure of the transgene construct. DNA sequence encoding a myc epitope tag (black box), followed by sequence encoding the truncated ΔN89-terminal domain, 13 central Armadillo (Arm) repeats, and the COOH-terminal domain of Xenopus β-catenin (shaded boxes) was embedded in β-globin intron/exon/polyadenylation sequences (βG), and cloned downstream of the mouse mammary tumor virus long terminal repeat (MMTV LTR). Pertinent restriction sites are indicated: N, NotI; B, BamHI; E, EcoRI. (B) 9E10 antibody, which specifically recognizes the human myc epitope tag and does not cross-react with murine c-myc, was used to immunoprecipitate myc-tagged ΔN89β-catenin from extracts of the mammary glands from 12 individual transgenic lines. This product was detected in three lines (lines 1, 3, and 5) by Western blotting with the same antibody. Note the lower band, which migrates at M r ∼ 50,000, is the IgG band from the 9E10 immunoprecipitation detected by the HRP-rabbit anti–mouse. (C) Southern blot detection of the ΔN89β-catenin transgene in the tail genomic DNA of transgenic (T) and normal (N) littermates of F1 from lines 1, 3, and 5, as indicated. Membranes were hybridized with the 32P-labeled ΔN89β-catenin cDNA insert (ΔβC), and the mouse engrailed gene (en) fragments, which detect fragments of 2,160 bp and 5 kb, respectively, as indicated. Lanes marked 200, 20, and 2 show respective copy equivalents of ΔN89β-catenin cDNA plasmid. (D) Relative expression levels of ΔN89β-catenin protein in lines 5 and 3 as detected by Western blotting of equal amounts of total mammary proteins with anti-myc antibody. Labels on the left indicate exposure time to film after incubation in ECL reagent. Arrows point to the 75-kD ΔN89β-catenin product; asterisks indicate nonspecific band detected by the anti–mouse IgG antibody. Detection of ribophorin was used as a loading control. (E) Western blot showing expression of ΔN89β-catenin (top) in line 5 transgenic mice during mammary development and differentiation. No reaction was seen on an identical blot of wt littermates (data not shown). Immunodetection of keratin 14 on the same blot (bottom) is shown to control for epithelial content of the sample. Preg, pregnancy; inv, involution. (F) Western blot of equal amounts of total proteins from day 16 pregnant mammary glands from line 5 mice showing expression of endogenous E-cadherin (top) and the ΔN89β-catenin transgenic product and endogenous β-catenin detected with the same anti–β-catenin antibody (bottom). Note densitometry of three similar blots showed that the transgene is expressed to approximately threefold the level of endogenous β-catenin found in wt mice. The transgene downregulates endogenous β-catenin by one third, and upregulates E-cadherin twofold compared with wt levels. (G–J) Indirect immunofluorescence on frozen sections of mammary gland from 6-mo-old virgin line 5 mice (G and J) or day 16 pregnant line 3 mice (H–I). ΔN89β-catenin is detected in green (G and H) using 9E10 anti-myc antibody. Endogenous β-catenin detected with rabbit anti–β-catenin shown in red (I). Double immunolabeling to detect ΔN89β-catenin in red and smooth muscle actin, which demarcates the position of myoepithelial cells in green (J). Note the transgene is located at the intercellular borders of the luminal epithelial cells.
ΔN89β-Catenin Induces Precocious Lobular Development of Virgin Mammary Gland
To determine the effect of MMTV–ΔN89β-catenin on early mammary development, we examined whole mount preparations of mammary glands from female, virgin transgenic and nontransgenic (wt) littermates that were matched for stage of the estrous cycle (Fig. 2). Even at early stages of puberty, marked differences could be seen in both of the high expressing lines between transgenic and wt mice (Fig. 2). At 4 wk, wt mammary epithelial ducts had commenced proliferation from the terminal end buds, and extended to the lymph node (Fig. 2 A). At higher magnification they displayed a smooth surface (Fig. 2 B). In transgenic littermates, the ducts were shorter, wider (Fig. 2 D), and covered with small lobular protuberances (Fig. 2 E). Extensive arrays of lobules were evident even at low magnification in 8-wk-old transgenic mice (Fig. 2C and Fig. F).
Figure 2.
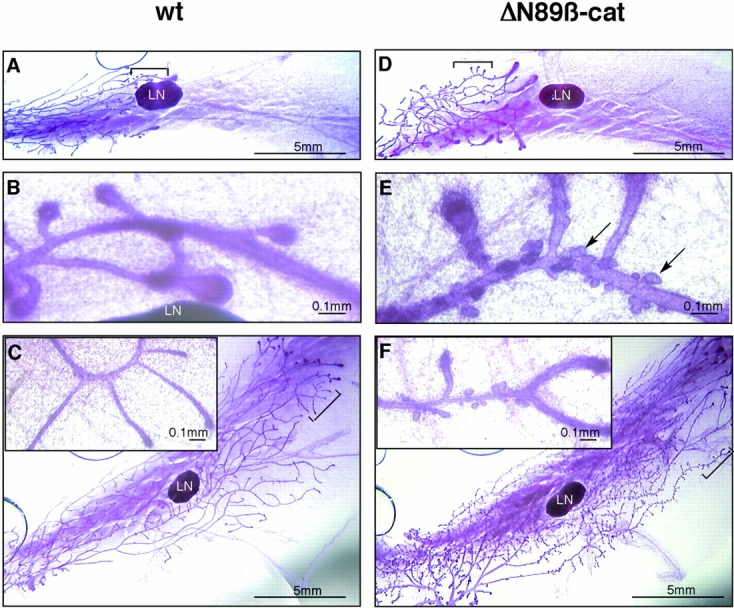
Morphology of ΔN89β-catenin mammary glands shows precocious lobular development during early puberty. Whole mounts of inguinal mammary glands stained with carmine alum from wt (A–C) and transgenic (D–F) 4-wk (A, B, D, and E) and 8-wk (C and F) line 5 littermates matched for stage of the estrous cycle. LN, lymph node. Note the extensive lobuloalveolar development on the transgenic glands (arrows in E). Images in B and E show higher magnifications of the areas bracketed in A and D. Insets in C and F are higher magnifications of bracketed areas.
At the end of puberty (12 wk), wt ducts were smooth and had small, regularly spaced side branches (Fig. 3A and Fig. B). However, virgin ΔN89β-catenin transgenic mammary glands showed extensive lobuloalveolar development of the type that is normally associated with the hormonal stimulation of pregnancy (Fig. 3C and Fig. D). The MMTV–ΔN89β-catenin phenotype contrasts strikingly to the feathery, hyperbranched appearance of mammary glands from MMTV-Wnt-1 mice of the same age and genetic background (Fig. 3E and Fig. F). Virgin MMTV-Wnt-1 mammary glands (Fig. 3 F) resembled those of wt mice in the early stage of pregnancy (8 d postcoitum [dpc]) (Fig. 3 G). In contrast, mammary glands from virgin ΔN89β-catenin mice (Fig. 3 D) resembled those of mid- to late pregnant (13 dpc) wt mice (Fig. 3 H). Mammary glands from pregnant ΔN89β-catenin mice (Fig. 3I and Fig. J) consistently showed lobular hyperplasia compared with those of wt littermates at the same stage of pregnancy (Fig. 3G and Fig. H).
Figure 3.
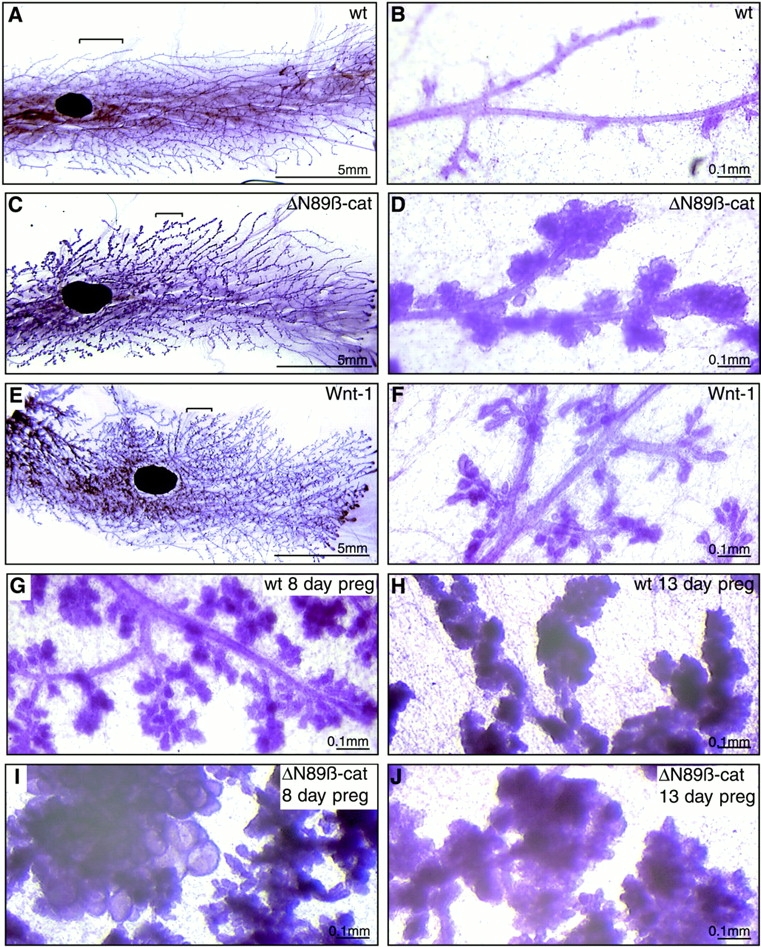
Comparison of the morphology of mature wt, MMTV-ΔN89β-catenin, and MMTV-Wnt-1 transgenic virgin and pregnant mammary glands. Whole mounts of inguinal mammary glands stained with carmine alum from 12-wk virgin wt (A and B) and ΔN89β-catenin transgenic littermates (C and D), and a 12-wk Wnt-1 transgenic mouse (E and F). Mammary glands from 8- and 13-d-old pregnant wt (G and H) and ΔN89β-catenin (I and J) littermates. Note that ΔN89β-catenin virgin mice show a hyperlobular phenotype, whereas Wnt-1 virgin mice show a feathery hyperbranched phenotype.
Differences between wt and transgenic animals from the low expressing line 3 were subtle during early development (data not shown), but became obvious with age (Fig. 4, A–D). Older ΔN89β-catenin line 3 mice again showed lobuloalveolar development in virgins with no indication of hyperbranching (Fig. 4A and Fig. B). Thus, the temporal appearance of the ΔN89β-catenin phenotype correlates with the dose of the transgene. Its later appearance in the low expressing line is likely due to the additive effects of repeated bursts of estrus on expression of the MMTV long terminal repeat.
Figure 4.
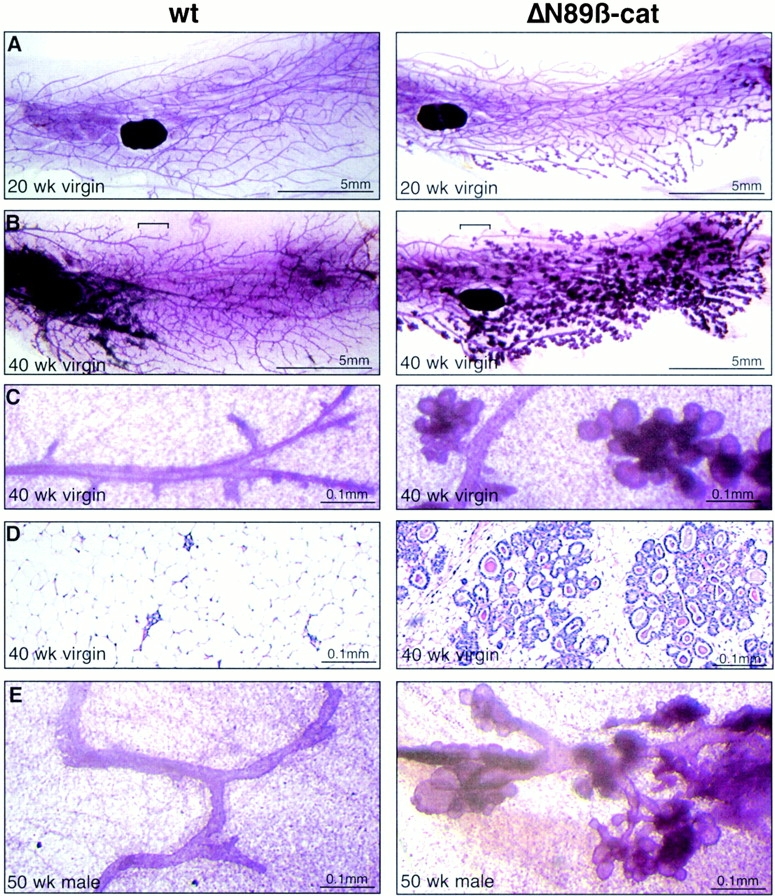
Lobuloalveolar development in older virgin females of the low expressing line and in male mice. Whole mounts of inguinal mammary glands stained with carmine alum (A–C) and hematoxylin and eosin–stained sections of mammary gland (D) from 20-wk-old (A) and 40-wk-old (B and C) wt and ΔN89β-catenin transgenic littermates from the low expressing line 3. Note that low expressing line 3 ΔN89β-catenin virgin mice also show aberrant hyperlobular development with no evidence of ductal hyperbranching. (E) 1-yr-old wt and ΔN89β-catenin transgenic male littermates from line 5.
ΔN89β-Catenin Induces Lobuloalveolar Development along the Primary Branch of the Male Rudiment in the Absence of Hormonal Stimuli
As ΔN89β-catenin female mice show lobular development in the absence of the hormonal stimulation associated with pregnancy, we asked if this phenotype could also be induced in males, in the complete absence of ovarian hormonal function. Mammary glands develop as epidermal invaginations during embryogenesis in a process that is similar in both males and females up to day 14. At this point they are pinched off in males by androgen-induced mesenchymal constriction (Sakakura 1987). In four pairs of whole mounts, the mammary rudiment of wt FVB/N mice formed a small blind duct at one end of the fat pad, occupying <2% of the gland (Fig. 4 E). In their ΔN89β-catenin littermates from the high expressing lines, this duct was covered in lobuloalveolar structures (Fig. 4 E). Significantly, the ducts did not branch or elongate, as reported for MMTV-Wnt-1 mice, nor did they expaand into the male fat pad, as occurs in MMTV-Wnt-10b mice (Tsukamoto et al. 1988; Lane and Leder 1997). No phenotype was observed in males of the low expressing line 3. Thus, ΔN89β-catenin specifically induces lobuloalveolar formation rather than hyperbranching in a dose-dependent manner in males, and can do so in the absence of female hormonal stimuli.
ΔN89β-Catenin Induces Functional Differentiation of Mature Virgin Mammary Gland
Histological examination of sections of the mammary glands of 6–8-mo-old virgin female wt and transgenic mice from low expressing line 3 (Fig. 4 D) and high expressing line 5 (Fig. 5, A–D) confirmed that ΔN89β-catenin induces extensive alveolar development. Large intracellular vesicles could be seen in the cuboidal cells lining the alveolar lumina of transgenic glands (Fig. 5 D), demonstrating that ΔN89β-catenin induces a secretory phenotype in the epithelium. To investigate if the precocious alveolar development was accompanied by functional differentiation, we examined expression levels of the milk protein β-casein in virgin and pregnant transgenic and wt littermates. As expected, β-casein expression was not detectable in wt virgin mice; its expression began around day 8 of pregnancy, became robust around day 13 of pregnancy, and was downregulated significantly by day 10 of involution (Fig. 5E and Fig. F). In contrast, young virgin transgenic mice produced low but detectable levels of β-casein, and old virgins produced amounts surpassing those seen during midpregnancy in wt mice (Fig. 5E and Fig. F). Taken together, these data show that ΔN89β-catenin induces lobular development and functional differentiation of the virgin mammary gland to that of midpregnant status.
Figure 5.
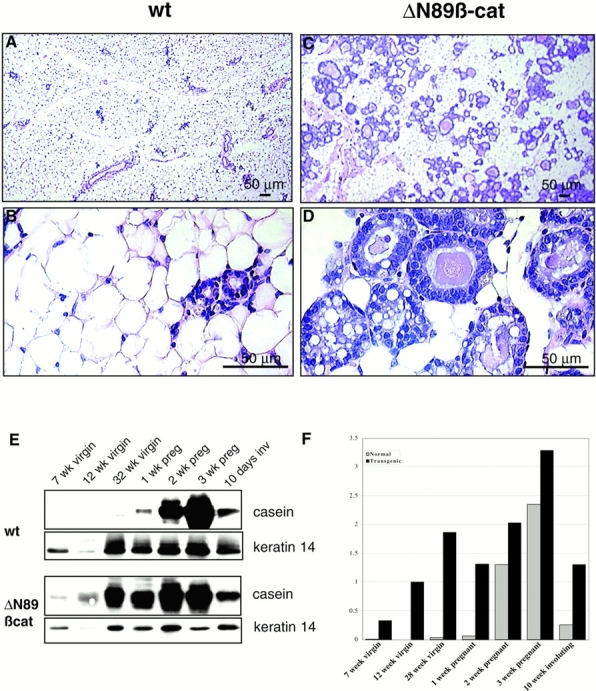
Histological and biochemical evaluation shows evidence of precocious epithelial differentiation in ΔN89β-catenin virgin mice. (A–D) Hematoxylin and eosin–stained sections of mammary gland from 6-mo-old virgin wt (A and B) and ΔN89β-catenin (C and D) littermates. Note the presence of vesicles in the transgenic samples. (E) Western blot of total proteins from the inguinal mammary glands at various stages of development, pregnancy (preg), and involution (inv) with anti–β-casein antibody and anti-keratin 14 antibody on the same blot, which serves as a control of epithelial content of the sample. (F) Graph of densitometry figures derived from the casein blots shown in E after normalization for keratin 14 levels. Note β-casein is expressed in ΔN89β-catenin but not wt virgin mice, and in all pregnant mice.
ΔN89β-Catenin Upregulates Cyclin D1 and c-myc mRNA in Virgin Mammary Gland
The phenotypes of cyclin D1−/− mice support a role for cyclin D1 expression in the epithelial cell autonomous induction of lobuloalveolar structures (Sicinski et al. 1995; Fantl et al. 1999). Cyclin D1 and c-myc induce precocious mammary development and transformation when overexpressed, and genes are targets of β-catenin transcriptional activity (Sinn et al. 1987; Wang et al. 1994; He et al. 1998; Tetsu and McCormick 1999). Therefore, we examined the effect of MMTV–ΔN89β-catenin on cyclin D1 and c-myc mRNA levels during mammary development. As expected, wt mice do not express significant levels of cyclin D1 mRNA, until midpregnancy (Fig. 6) (Stepanova et al. 1996, Stepanova et al. 2000). In contrast, virgin ΔN89β-catenin mice express abnormally high levels of cyclin D1 mRNA as well as elevated levels of c-myc mRNA (Fig. 6). Expression levels of these mRNAs were similar in wt and transgenic glands throughout pregnancy.
Figure 6.

Northern analysis of cyclin D1 and c-myc mRNA levels in wt and ΔN89β-catenin mammary glands. mRNA was extracted from virgin (10 wk [10w Vir]), midpregnant (8 d [8d Preg]), and late pregnant (18 d [18d Preg]) mammary glands. Northern blots were prepared and probed with cDNAs encoding cyclin D1, c-myc, and K18 as a control to permit normalization for epithelial content of the mRNA samples. Note cyclin D1 and c-myc levels are abnormally elevated in transgenic (TG) virgin mice compared with their wt littermates.
ΔN89β-Catenin Effects on Involution of the Mammary Gland
We next asked whether MMTV–ΔN89β-catenin mammary glands were able to involute correctly after cessation of lactation. Transgenic and wt littermates were allowed to give birth and nurse seven pups for 7 d. We killed the mothers at various time points up to 21 d later, and processed their mammary glands as whole mounts and histological sections. Involution is a two-step process. Early after the removal of pups, programmed cell death of the secretory epithelium occurs with loss of differentiated product (Lund et al. 1996). Epithelial cell death continues during the second stage, at which time proteinases remodel the stroma and a biosynthetic wave of adipogenesis restores the fat pad (Lund et al. 1996). No significant histological difference was seen between the wt (Fig. 7 A, top) and transgenic samples (Fig. 7 A, bottom), up to day 7 of involution. Both wt and transgenic glands undergo an early apoptotic process that rapidly peaks in a scheduled fashion in both samples at day 3 of involution, as assessed by TUNEL staining of tissue sections (Fig. 7 B and Fig. 8 A). By day 10, however, wt mammary glands show greater alveolar degeneration and overall reduction in the relative epithelial mass than the transgenic alveolar epithelium, which remains hyperplastic (Fig. 8 B). This discrepancy between the wt and transgenic glands persists even up to day 21, the time at which involution is considered complete in wt mice. The transgenic epithelial cells contain many large intracellular vesicles, indicating continued secretory activity (data not shown). Moreover, casein and cyclin D1 mRNA expression, which are lost in wt mice, continue to be detected at day 21 in transgenic mice (Fig. 8 C). Thus, the mammary gland reverts to the virgin status in both wt and transgenic animals. However, as virgin ΔN89β-catenin mice already have a midpregnancy phenotype (Fig. 5E and Fig. F), their involution does not result in full loss of alveolar structure or differentiation.
Figure 7.
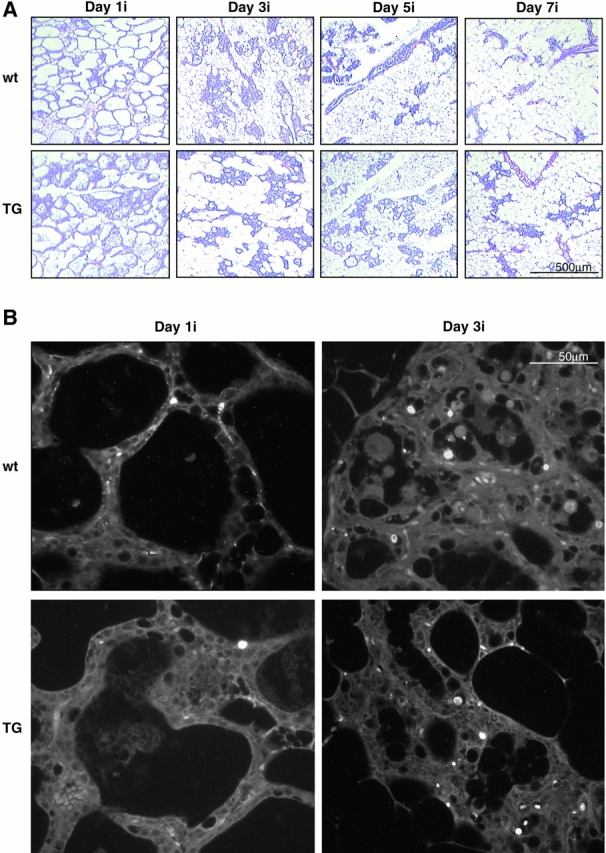
A comparison of early involution of wt and ΔN89β-catenin mammary glands. (A) Hematoxylin and eosin–stained sections of wt (top) and ΔN89β-catenin (bottom) inguinal mammary glands from littermates 1, 3, 5, and 7 d of involution (i) after removal of pups, as indicated. (B) Apoptotic changes detected by TUNEL staining of sections from wt (top) and ΔN89β-catenin (TG, bottom) inguinal mammary glands 1 and 3 d after removing pups, as indicated (1i, 3i). Note no significant differences were observed between wt and transgenic samples during this early stage of involution.
Figure 8.
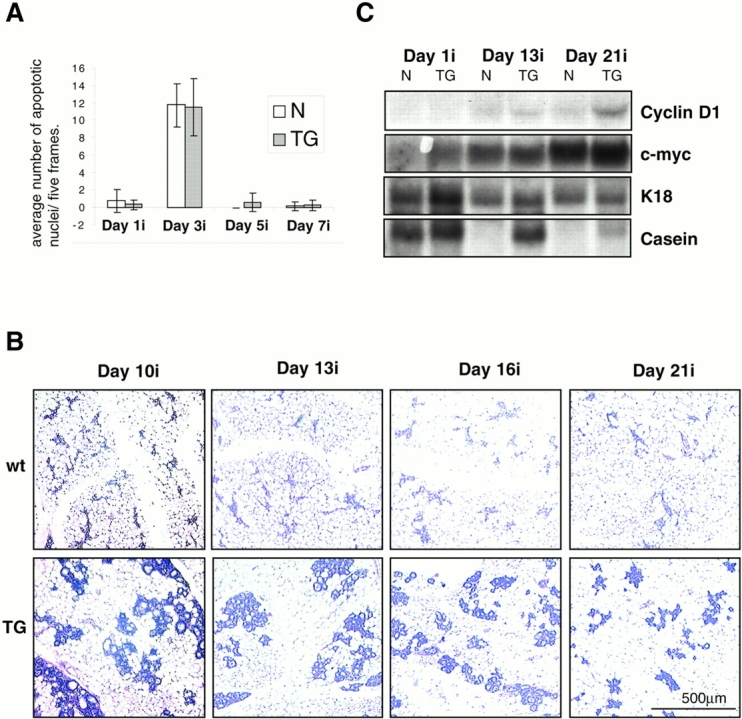
A comparison of late involution in wt and ΔN89β-catenin mammary glands. (A) Graph of the average number of TUNEL-positive nuclei in five random frames taken from sections of wt and ΔN89β-catenin (TG) inguinal mammary glands from 1, 3, 5, and 7 d after removing pups as indicated. (B) Hematoxylin and eosin–stained sections of wt (top) and ΔN89β-catenin (bottom) inguinal mammary glands from littermates 10, 13, 16, and 21 d after removing pups. Note that in contrast to wt glands, which show continued signs of deterioration and reduction of the epithelial component, ΔN89β-catenin mice show continued lobular hyperplasia. (C) mRNA was extracted from mammary glands having undergone involution for 1, 13, and 21 d, as indicated. Northern blots were prepared and probed with cDNAs encoding cyclin D1, c-myc, casein, and K18 as a control to permit normalization for epithelial content of the mRNA samples. Note cyclin D1 becomes elevated and casein mRNA persists at day 21 of involution in transgenic mice compared with their wt littermates.
ΔN89β-Catenin Induces Adenocarcinomas in 100% of Female Mice
In both transgenic lines, breeding females were unable to feed the third and subsequent litters of pups, and developed large mammary growths (Fig. 9 A). This did not occur in wt littermates. The mammary growths began as large, fluid-filled cysts, and at later times formed highly vascularized encapsulated tumors with necrotic centers (Fig. 9 B). Metastases were not observed, but the tumors sometimes showed invasion of local muscular tissues. The tumors displayed both a glandular (Fig. 9C and Fig. D) and an undifferentiated pattern (Fig. 9C and Fig. F) consisting of tightly packed epithelial cells (Fig. 9 F). A secretory phenotype was variably evident in both components (Fig. 9 E). Cellular atypia and mitotic activity were most prominent in the undifferentiated regions (Fig. 9 F). These features are consistent with adenocarcinoma with glandular, secretory, and undifferentiated elements. Northern analysis of tumors that developed in virgin animals revealed that both cyclin D1 and c-myc mRNA levels were elevated in tumor samples, compared with mammary tissue from wt littermates after normalization to K18 levels to correct for the expansion of the epithelial compartment seen in the tumor samples (Fig. 9 G). To quantify the tumor incidence and rate of progression, we established wt and transgenic pairs of males, virgin females, and breeding females from both high and low expressing lines 5 and 3, respectively (Fig. 9 H). 100% of transgenic breeding females developed multiple (average three per mouse) large tumors, which became visible at ∼4 mo of age in the high expressing line 5, and at 6.5 mo of age in the low expressing line 3. In both lines the tumors grew rapidly, requiring killing of the animals ∼4 wk later. Tumors developed later in life (∼7 mo) in virgin females of the high expressing line 5, and grew with similar frequency and rate of progression. 30% of virgin females of the low expressing line 3, which are currently 13 mo old, have developed mammary tumors. Only one male (line 5) developed a tumor.
Figure 9.
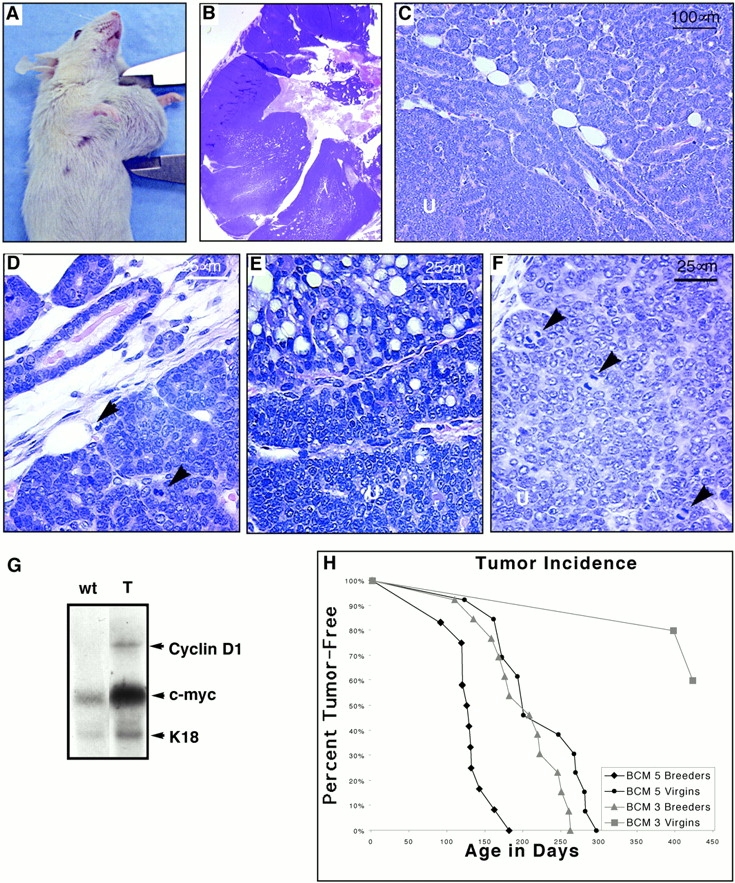
ΔN89β-catenin mice develop mammary adenocarcinomas. (A) Founder mouse line 5 with single large tumor on a thoracic mammary gland. (B) Hematoxylin and eosin–stained section of similar tumor from an F1 daughter. Tumors comprise regions of glandular hyperplasia (C and D) with central masses of disorganized, undifferentiated (U) poorly adherent cells (C, E, and F). Some showed evidence of secretory activity (E) but most showed undifferentiated cells with a high cytoplasmic to nuclear ratio and many mitotic figures (arrowheads in D and F). (G) mRNA was extracted from mammary tumors (T) and mammary glands of wt littermates. Northern blots were prepared and probed with cDNAs encoding cyclin D1, c-myc, and K18 as a control to permit normalization for epithelial content of the mRNA samples. (H) Graph of tumor incidence in ΔN89β-catenin mice. The percentage of animals in each cohort remaining free of palpable tumors was plotted as a function of age for ΔN89β-catenin line 5 breeding females (n = 12), line 3 breeding females (n = 13), and line 5 virgins (n = 13). No female wt littermates (n = 25) or wt littermate males (n = 20) developed tumors. To date, among mice that are currently > 1 yr old, three line 3 virgin females (n = 10), no line 3 males (n = 10), and only one line 5 male (n = 10) have developed tumors.
Discussion
Our goal in this study was to determine the effect of increasing the pool of transcriptionally active β-catenin on mammary gland development and pathology. Our first observation was that expression of MMTV–ΔN89β-catenin during early mammary development produced a mild retardation in ductal extension. This effect is reminiscent of the inhibition of tubulogenesis in MDCK cysts produced by a similar mutant, ΔN90β-catenin, which was hypothesized to interfere with the ability of adenomatous polyposis coli to bundle microtubules at the tips of cell extension (Nathke et al. 1996; Pollack et al. 1997). However, the most striking change elicited by the early expression of MMTV–ΔN89β-catenin is the inappropriate lobuloalveolar development of the virgin mammary gland. Stem cells that generate the alveoli are dispersed throughout the mammary gland and cyclically activated during pregnancy (Kordon and Smith 1998). ΔN89β-catenin's specific effect on these mammary stem cells parallels its actions in mouse skin and intestine. ΔN89β-catenin reprograms both epidermal and follicle stem cells to generate supernumerary hair follicles and induces stem cells in the intestinal crypts to form adenomatous polyps, but has no effect on the differentiated cells of the villi (Gat et al. 1998; Wong et al. 1998; Harada et al. 1999). As mice lacking Lef-1 or Tcf-4, the transcriptional partners of β-catenin, fail to form mammary glands and intestinal crypt cells, respectively, it has been proposed that β-catenin's ability to modify Lef/Tcf function may maintain stem-like multipotency (van Genderen et al. 1994; Korinek et al. 1998).
β-Catenin and Wnt Signaling in Mammary Gland
ΔN89β-catenin induces lobuloalveolar development in males as well as females, and therefore, joins several proteins whose overexpression or loss of function bypasses the requirement for hormonal stimuli to induce lobuloalveolar development (Henninghausen and Robinson 1998; Robinson et al. 2000). Among this list of proteins, there is a considerable body of data to link β-catenin, Wnts, cadherins, matrix metalloproteinases, and cyclin D1 into a putative pathway. Wnts comprise a large family of secreted, short-range signaling proteins. Mammary gland expresses Wnt-2, -5a, -7b, and -10b during puberty, expresses Wnt-4, -5b, and -6 during pregnancy, loses Wnt expression during lactation, and resumes it again during involution (Gavin et al. 1990; Buhler et al. 1993; Weber-Hall et al. 1994). The phenotype induced by MMTV–ΔN89β-catenin most closely resembles the changes associated with late pregnancy, suggesting that endogenous β-catenin may normally convey signals from the pregnancy-associated Wnts. Wnt-4 has recently been demonstrated to be regulated by progesterone (Brisken et al. 2000). To date, there is no evidence to connect Wnt-4, -5b, or -6 to β-catenin. Indeed, these particular Wnts do not upregulate β-catenin in C57MG cells, although it is not clear if these cells express appropriate receptors (Shimizu et al. 1997). In contrast, Wnt-2, which is expressed in stromal cells during ductal proliferation and is maintained during pregnancy, does upregulate β-catenin in C57MG cells, and is, therefore, a potential paracrine regulator of β-catenin function in vivo (Shimizu et al. 1997). Significantly, Wnt-2 amplification has been found in mouse mammary tumors, and its upregulation has been reported in human breast carcinomas (Roelink et al. 1992; Dale et al. 1996).
Wnt-10b, a gene that is expressed in mammary gland, and Wnt-1 and Wnt-3, which are normally silent in this tissue, share with ΔN89β-catenin the ability to induce lobular hyperplasia and adenocarcinomas when expressed from the MMTV promoter (Tsukamoto et al. 1988; Roelink et al. 1990; Lane and Leder 1997). This strongly suggests that β-catenin mediates the proliferative and oncogenic actions of these Wnts. However, the early morphogenic phenotypes of MMTV-Wnt-1, Wnt-3, Wnt-10b, and ΔN89β-catenin mice do not fully superimpose (Tsukamoto et al. 1988; Lane and Leder 1997). ΔN89β-catenin specifically induces lobuloalveolar development in females and males, whereas Wnt-1 and Wnt-10b additionally induce feathery, ductal hyperbranching in females and significant male ductal extension. There are several possible explanations for the partial incongruence among the phenotypes. First, it can be argued that different Wnts may sustain distinct levels of β-catenin that critically determine the phenotype. This argument can be dismissed, as both high and low expressing lines of ΔN89β-catenin mice ultimately produce the same phenotype. A second possible reason for the disparity could involve the use, in our study, of an NH2-terminally deleted ΔN89β-catenin. This mutant lacks sites essential for its degradation, and is therefore commonly used to mimic Wnt-induced stabilization of β-catenin. Deletion of the NH2-terminus, however, also removes sites that can bind additional regulatory factors, such as nemo-like kinase (NLK), a protein that modulates the function and location of downstream elements of Wnt cascades (Ishitani et al. 1999). A third important difference involves the possibility that Wnts may operate through additional or alternative downstream effectors than β-catenin. While some Wnts have been shown to stimulate G protein–coupled pathways, Wnt-1, -3, and -10b are commonly thought to signal through β-catenin. However, Wnt-1 expression in cultured cells also upregulates plakoglobin, a close relative of β-catenin, and plakoglobin rather than β-catenin has been shown to reproduce a Wnt-3 overexpression phenotype in mouse epidermis (Bradley et al. 1993; Hinck et al. 1994; Charpentier et al. 2000). In some cell types plakoglobin appears to act as an oncogene, and in others it suppresses proliferation and tumorigenicity (Simcha et al. 1996; Charpentier et al. 2000; Hakimelahi et al. 2000; Kolligs et al. 2000). It has variously been reported to augment, inhibit, or have no effect on cytosolic levels and signaling function of β-catenin (Merriam et al. 1997; Miller and Moon 1997; Salomon et al. 1997; Charpentier et al. 2000; Williams et al. 2000). Currently, the role of plakoglobin in mammary gland is not known, and its potential contribution to Wnt signaling in this gland needs to be tested.
Our data indicate that ΔN89β-catenin recapitulates specific elements of MMTV-Wnt-1, -3, and -10b phenotypes. However, the endogenous Wnt pathways activated by expression of these Wnts appear to involve one additional downstream effector or modulator. Our work is consistent with the possibility that endogenous β-catenin functions to trigger lobuloalveolar differentiation. Proof of this hypothesis, however, requires an analysis of the effects of blocking β-catenin signaling on the different stages of mammary development. Overexpression of axin and dominant negative forms of Tcf has been used to achieve this effect in other systems, but has the disadvantage of potentially affecting plakoglobin, which also binds to these proteins (Kodama et al. 1999; Williams et al. 2000). The use of dominant negative β-catenin appears to avoid this potential problem, and we are currently exploring this approach (Montross et al. 2000).
ΔN89β-Catenin and Potential Downstream Targets
Our results demonstrate upregulation of E-cadherin, cyclin D1, c-myc, and casein in ΔN89β-catenin mammary gland. An increase in E-cadherin protein levels in response to expression of β-catenin or its relative plakoglobin has been observed in other in vitro and in vivo systems (Bradley et al. 1993; Hinck et al. 1994; Wong et al. 1998). Indeed, the E-cadherin gene may be a transcriptional target of Wnt (Huber et al. 1996; Yanagawa et al. 1997). However, the early ΔN89β-catenin phenotype resembles that of P-cadherin null and E-cadherin dominant negative mice, suggesting that cadherins antagonize β-catenin in the induction of lobuloalveolar development (Radice et al. 1997; Delmas et al. 1999).
At the present time, the most significant observation with respect to potential downstream effectors of β-catenin signaling involves the ΔN89β-catenin–induced upregulation of cyclin D1 and c-myc mRNA in virgin mice. β-Catenin was first demonstrated to promote expression of the cell cycle regulators cyclin D1 and c-myc in colonic cells, and appears to be operating in mammary gland in a similar fashion (He et al. 1998; Tetsu and McCormick 1999). Mice expressing MMTV–c-myc, MMTV–cyclin D1, and MMTV-CDC37, the chaperone for the cyclin D1 cdk4 partner, develop lobuloalveolar hyperplasia and are prone to adenocarcinoma at ∼1–2 yr of age (Sinn et al. 1987; Wang et al. 1994; Stepanova et al. 2000). There is complete congruency between the early phenotype of the MMTV–ΔN89β-catenin mice, and that of mice lacking cyclin D1. Analyses of the cyclin D1 null mice show that like ΔN89β-catenin, the cyclin D1 gene specifically affects lobuloalveolar development and has no effect on ductal branching (Sicinski et al. 1995; Fantl et al. 1999). Cyclin D1−/− mice show normal side branching but marked reduction of alveoli and a lactation defect, a phenotype that fits exactly that predicted for loss of a β-catenin–regulated gene.
ΔN89β-Catenin Induces Poorly Differentiated Adenocarcinomas
ΔN89β-catenin induces hyperplasia and benign tumors in other tissues; however, only in mammary gland does it produce such aggressive tumors. Several molecular mechanisms have been invoked to explain the tumor-promoting action of β-catenin. Endogenous cytoplasmic levels of β-catenin oscillate with the cell cycle, and increasing β-catenin expedites the G1-S phase transition (Orford et al. 1999). This can be explained in the ΔN89β-catenin mammary gland by the upregulation of both cyclin D1 and c-myc. Cyclin D1–cdk4 complexes phosphorylate and functionally inactivate RB, leading to release of E2F, which activates transcription of genes necessary for the completion of the cell cycle (Sutherland et al. 1998). Cyclin D1–cdk4 complexes also promote cyclin E/cdk2 activity by sequestering inhibitory p21 and p27 subunits. C-myc elevates expression of cyclin E, cdc25A, and factors that prevent formation of the p27– and p21–cdk inhibitory complexes, and lowers expression of the inhibitory p27 subunit (Sutherland et al. 1998). Thus, the concerted actions of cyclin D1 and c-myc result in rapidly cycling cells which produce a preneoplastic alveolar hyperplasia. Further genetic insults to this increased pool of cells escape G1-S checkpoint control and lead to tumor progression. Overexpression of cyclin D1 or c-myc predisposes mice to mammary tumors, but with much slower kinetics and penetrance than is seen even with the low expressing ΔN89β-catenin line (Stepanova et al. 2000). It is likely that the ability of ΔN89β-catenin to concurrently elevate both c-myc and cyclin D1 accounts for the robust growth of the ΔN89β-catenin mammary tumors. Both cyclin D1 and c-myc are highly expressed in 40% of human breast tumors. Significantly, more than half of these tumors show no amplification or genetic changes in the c-myc or cyclin D1 locus (Leder et al. 1986; Buckley and Middleton 1987; Wang et al. 1994). By analogy to human colon cancer, the possibility must now be investigated that mutations in tissue-specific, negative regulators of β-catenin may be involved in such a subset of human breast cancer.
Acknowledgments
We thank Elizabeth Acquista for excellent technical assistance with the immunofluorescence and photography, and Drs. Peggy Neville and Caroline Alexander of the Universities of Colorado and Wisconsin, respectively, for helpful comments.
This work was supported by National Institutes of Health grant GM47429 and a New York State Health Research Science Board Empire award to P. Cowin, and a New York State Health Research Science Board postdoctoral fellowship to A. Imbert.
Footnotes
Alexandra Imbert and Rachel Eelkema contributed equally to this work and should be considered co-first authors.
Abbreviations used in this paper: Lef, lymphoid enhancer factor; Tcf, T cell factor; TUNEL, terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling; wt, wild-type.
References
- Behrens J., von Kries J.P., Kuhl M., Bruhn L., Wedlich D., Grosschedl R., Birchmeier W. Functional interaction of β-catenin with the transcription factor LEF-1. Nature. 1996;382:638–642. doi: 10.1038/382638a0. [DOI] [PubMed] [Google Scholar]
- Berx G., Cleton-Jansen A.M., Nollet F., de Leeuw W.J., van de Vijver M., Cornelisse C., van Roy F. E-cadherin is a tumour/invasion suppressor gene mutated in human lobular breast cancers. EMBO (Eur. Mol. Biol. Organ.) J. 1995;14:6107–6115. doi: 10.1002/j.1460-2075.1995.tb00301.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradley R.S., Cowin P., Brown A.M.C. Expression of Wnt-1 in PC12 cells results in modulation of plakoglobin and E-cadherin and increased cellular adhesion. J. Cell Biol. 1993;123:1857–1865. doi: 10.1083/jcb.123.6.1857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brisken C., Heineman A., Chavarra T., Elenbaas B., Tan J., Dey S.K., McMahon A.P., Weinberg R. Essential function of Wnt-4 in mammary gland development downstream of progesterone signaling. Genes Dev. 2000;14:650–654. [PMC free article] [PubMed] [Google Scholar]
- Buckley A., Middleton M.C. Retinoic acid alters the keratinization of cultured rat sublingual keratinocytes in vitro. Arch. Dermatol. Res. 1987;279:257–265. doi: 10.1007/BF00417325. [DOI] [PubMed] [Google Scholar]
- Buhler T.A., Dale T.C., Kieback C., Humphreys R.C., Rosen J.M. Localization and quantification of Wnt-2 gene expression in mouse mammary development. Dev. Biol. 1993;155:87–96. doi: 10.1006/dbio.1993.1009. [DOI] [PubMed] [Google Scholar]
- Butz S., Stappert J., Weissig H., Kemler R. Plakoglobin and β-catenindistinct but closely related. Science. 1992;257:1142–1143. doi: 10.1126/science.257.5073.1142-a. [DOI] [PubMed] [Google Scholar]
- Charpentier E., Lavker R., Acquista E., Cowin P. Plakoglobin regulates cell proliferation and hair growth in vivo. J. Cell Biol. 2000;149:503–520. doi: 10.1083/jcb.149.2.503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cowin P., Burke B. Cytoskeleton-membrane interactions. Curr. Opin. Cell Biol. 1996;8:56–65. doi: 10.1016/s0955-0674(96)80049-4. [DOI] [PubMed] [Google Scholar]
- Dale T.C., Weber-Hall S.J., Smith K., Huguet E.L., Jayatilake H., Gusterson B.A., Shuttleworth G., O'Hare M., Harris A.L. Compartment switching of Wnt-2 expression in human breast tumors. Cancer Res. 1996;56:4320–4323. [PubMed] [Google Scholar]
- Daniel C.W., Silberstein G.B. Postnatal development of the rodent mammary gland. In: Neville M.C., Daniel C.W., editors. The Mammary Gland. Plenum; New York: 1987. pp. 3–31. [Google Scholar]
- Delmas V., Pla P., Feracci H., Thiery J.P., Kemler R., Larue L. Expression of the cytoplasmic domain of E-cadherin induces precocious mammary epithelial alveolar formation and affects cell polarity and cell-matrix integrity. Dev. Biol. 1999;216:491–506. doi: 10.1006/dbio.1999.9517. [DOI] [PubMed] [Google Scholar]
- Fantl V., Edwards A.W., Steel J.H., Vonderhaar B.K., Dickson D. Impaired mammary gland development in Cyc−/− mice during pregnancy and lactation is epithelial cell autonomous. Development. 1999;212:1–11. doi: 10.1006/dbio.1999.9329. [DOI] [PubMed] [Google Scholar]
- Funayama N., Fagotto F., McCrea P., Gumbiner B.M. Embryonic axis induction by the Armadillo repeat domain of β-cateninevidence for intracellular signaling. J. Cell Biol. 1995;128:959–968. doi: 10.1083/jcb.128.5.959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gat U., Dasgupta R., Degenstein L., Fuchs E. De novo hair follicle morphogenesis and hair tumors in mice expressing a truncated β-catenin in skin. Cell. 1998;95:605–614. doi: 10.1016/s0092-8674(00)81631-1. [DOI] [PubMed] [Google Scholar]
- Gavin B.J., McMahon J.A., McMahon A.P. Expression of multiple novel Wnt-1/int-1 related genes during fetal and adult mouse development. Genes Dev. 1990;4:2319–2332. doi: 10.1101/gad.4.12b.2319. [DOI] [PubMed] [Google Scholar]
- Gumbiner B. Signal transduction by β-catenin. Curr. Opin. Cell Biol. 1995;7:634–640. doi: 10.1016/0955-0674(95)80104-9. [DOI] [PubMed] [Google Scholar]
- Haegel H., Larue L., Oshugi M., Fedorov L., Herrenknecht K., Kemler R. Lack of β-catenin affects mouse development at gastrulation. Development. 1995;121:3529–3537. doi: 10.1242/dev.121.11.3529. [DOI] [PubMed] [Google Scholar]
- Hakimelahi S., Parker H.R., Gilchrist A.J., Barry M., Li Z., Bleackley R.C., Pasdar M. Plakoglobin regulates the expression of the anti-apoptotic protein BCL-2. J. Biol. Chem. 2000;275:10905–10911. doi: 10.1074/jbc.275.15.10905. [DOI] [PubMed] [Google Scholar]
- Harada N., Tamai Y., Ishikawa T., Sauer B., Takaku K., Oshima M., Taketo M. Intestinal polyposis in mice with a dominant stable mutation of the β-catenin gene. EMBO (Eur. Mol. Biol. Organ.) J. 1999;18:5931–5942. doi: 10.1093/emboj/18.21.5931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He T.-C., Sparks A.B., Rago C., Hermeking H., Zawel L., da Costa L.T., Morin P.J., Vogelstein B., Kinzler K.W. Identification of c-myc as a target of the APC pathway. Science. 1998;281:1509–1512. doi: 10.1126/science.281.5382.1509. [DOI] [PubMed] [Google Scholar]
- Henninghausen L., Robinson G.W. Think globally, act locallythe making of a mouse mammary gland. Genes Dev. 1998;12:449–455. doi: 10.1101/gad.12.4.449. [DOI] [PubMed] [Google Scholar]
- Hinck L., Nelson W.J., Papkoff J. Wnt-1 modulates cell–cell adhesion in mammalian cells by stabilizing β-catenin binding to the cell adhesion protein cadherin. J. Cell Biol. 1994;124:729–741. doi: 10.1083/jcb.124.5.729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huber O., Korn R., McLaughlin J., Ohsugi M., Herrmann B.G., Kemler R. Nuclear localization of β-catenin by interaction with transcription factor LEF-1. Mech. Dev. 1996;59:3–10. doi: 10.1016/0925-4773(96)00597-7. [DOI] [PubMed] [Google Scholar]
- Ishitani T., Ninomiya-Tsuji J., Nagai S., Nishita M., Meneghini M., Barker N., Waterman M., Bowerman B., Clevers H., Shibuya H., Matsumoto K. The Tak1-NLK-MAPK-related pathway antagonizes signalling between β-catenin and transcription factor TCF. Nature. 1999;399:798–802. doi: 10.1038/21674. [DOI] [PubMed] [Google Scholar]
- Kodama S., Ikeda S., Asahara T., Kishida M., Kikuchi A. Axin directly interacts with plakoglobin and regulates its stability. J. Biol. Chem. 1999;274:27682–27688. doi: 10.1074/jbc.274.39.27682. [DOI] [PubMed] [Google Scholar]
- Kolligs F.T., Kolligs B., Hajra K.M., Hu G., Tani M., Cho K.R., Fearon E.R. Gamma-catenin is regulated by the APC tumor suppressor and its oncogenic activity is distinct from that of beta-catenin. Genes Dev. 2000;14:1319–1331. [PMC free article] [PubMed] [Google Scholar]
- Kordon E.C., Smith G.H. An entire functional mammary gland may comprise the progeny from a single cell. Development. 1998;125:1921–1930. doi: 10.1242/dev.125.10.1921. [DOI] [PubMed] [Google Scholar]
- Korinek V., Barker N., Moerer P., van Donselaar E., Huls G., Peters P., Clevers H. Depletion of epithelial stem-cell compartments in the small intestine of mice lacking Tcf-4. Nat. Genet. 1998;19:379–383. doi: 10.1038/1270. [DOI] [PubMed] [Google Scholar]
- Lane T.F., Leder P. Wnt10B directs hypermorphic development and transformation in mammary glands of male and female mice. Oncogene. 1997;15:2133–2144. doi: 10.1038/sj.onc.1201593. [DOI] [PubMed] [Google Scholar]
- Leder A., Pattengale P.K., Kuo A., Stewart T.A., Leder P. Consequences of widespread deregulation of the c-myc gene in transgenic micemultiple neoplasms and normal development. Cell. 1986;45:485–495. doi: 10.1016/0092-8674(86)90280-1. [DOI] [PubMed] [Google Scholar]
- Lund L.R., Romer J., Thomasset N., Solberg H., Pyke C., Bissell M.J., Dano K., Werb Z. Two independent phases of apoptosis in mammary gland involutionproteinase-independent and -dependent pathways. Development. 1996;122:181–193. doi: 10.1242/dev.122.1.181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCrea P.D., Turck C.W., Gumbiner B. A homolog of the Armadillo protein in Drosophila (plakoglobin) associated with E-cadherin. Science. 1991;254:1359–1361. doi: 10.1126/science.1962194. [DOI] [PubMed] [Google Scholar]
- Merriam J.M., Rubenstein A.B., Klymkowsky M.W. Cytoplasmically anchored plakoglobin induces a WNT-like phenotype in Xenopus . Dev. Biol. 1997;185:67–81. doi: 10.1006/dbio.1997.8550. [DOI] [PubMed] [Google Scholar]
- Miller J.R., Moon R.T. Analysis of the signaling activities of localization mutants of β-catenin during axis specification in Xenopus . J. Cell Biol. 1997;139:229–243. doi: 10.1083/jcb.139.1.229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molenaar M., van de Wetering M., Oosterwegel M., Peterson-Maduro J., Godsave S., Korinek V., Roose J., Destree O., Clevers H. XTcf-3 transcription factor mediates β-catenin-induced axis formation in Xenopus embryos. Cell. 1996;86:391–399. doi: 10.1016/s0092-8674(00)80112-9. [DOI] [PubMed] [Google Scholar]
- Montross W.T., Ji H., McCrea P.D. A beta-catenin/engrailed chimera selectively suppresses Wnt signaling. J. Cell Sci. 2000;113:1759–1770. doi: 10.1242/jcs.113.10.1759. [DOI] [PubMed] [Google Scholar]
- Munemitsu S., Albert I., Rubinfeld B., Polakis P. Deletion of an amino-terminal sequence stabilizes β-catenin in vivo and promotes hyperphosphorylation of the adenomatous polyposis coli tumor suppressor protein. Mol. Cell. Biol. 1996;16:4088–4094. doi: 10.1128/mcb.16.8.4088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nathke I.S., Adams C.L., Polakis P., Sellin J.H., Nelson W.J. The adenomatous polyposis coli tumor suppressor protein localizes to plasma membrane sites involved in active cell migration. J. Cell Biol. 1996;134:165–179. doi: 10.1083/jcb.134.1.165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nusse R., Varmus H. Wnt genes. Cell. 1992;69:1073–1087. doi: 10.1016/0092-8674(92)90630-u. [DOI] [PubMed] [Google Scholar]
- Orford K., Orford C.C., Byers S. Exogenous expression of β-catenin regulates contact inhibition, anchorage-independent growth, anoikis, and radiation-induced cell cycle arrest. J. Cell Biol. 1999;146:855–868. doi: 10.1083/jcb.146.4.855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polakis P. The oncogenic activation of β-catenin. Curr. Opin. Genet. Dev. 1999;9:15–21. doi: 10.1016/s0959-437x(99)80003-3. [DOI] [PubMed] [Google Scholar]
- Pollack A.L., Barth A., Altshuler Y., Nelson W.J., Mostov K.E. Dynamics of β-catenin interactions with APC protein regulate epithelial tubulogenesis. J. Cell Biol. 1997;137:1651–1662. doi: 10.1083/jcb.137.7.1651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radice G., Ferreira-Cornwall C., Robinson S.D., Rayburn H., Chodosh L.A., Takeichi M., Hynes R.O. Precocious mammary gland development in P-cadherin–deficient mice. J. Cell Biol. 1997;139:1025–1032. doi: 10.1083/jcb.139.4.1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson G.W., Henninghausen L., Johnson P.E. Side-branching in the mammary glandthe progesterone-Wnt connection. Genes Dev. 2000;14:889–894. [PubMed] [Google Scholar]
- Roelink H., Wagenaar E., Lopes da Silva S., Nusse R. Wnt-3, a gene activated by proviral insertion in mouse mammary tumors, is homologous to int-1/Wnt-1 and is normally expressed in mouse embryos and adult brain. Proc. Natl. Acad. Sci. USA. 1990;87:4519–4523. doi: 10.1073/pnas.87.12.4519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roelink H., Wagenaar E., Nusse R. Amplification and proviral activation of several Wnt genes during progression and clonal variation of mouse mammary tumors. Oncogene. 1992;7:487–492. [PubMed] [Google Scholar]
- Sakakura T. Mammary embryogenesis. In: Neville M.C., Daniel C.W., editors. The Mammary Gland. Plenum; New York: 1987. pp. 37–63. [Google Scholar]
- Salomon D., Sacco P., Roy S., Simcha I., Johnson K.R., Wheelock M.J., Ben-Ze'ev A. Regulation of β-catenin levels and localization by overexpression of plakoglobin and inhibition of the ubiquitin-proteosome system. J. Cell Biol. 1997;139:1325–1335. doi: 10.1083/jcb.139.5.1325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimizu H., Julius M.A., Giarre M., Zheng Z., Brown A.M.C., Kitajewsky J. Transformation by Wnt family proteins correlates with regulation of β-catenin. Cell Growth Differ. 1997;8:1349–1358. [PubMed] [Google Scholar]
- Sicinski P., Donaher J.L., Parker S.B., Li T., Fazeli A., Gardner H., Haslam S.Z., Bronson R.T., Elledge S.J., Weinberg R.A. Cyclin D1 provides a link between development and oncogenesis in the retina and breast. Cell. 1995;82:621–630. doi: 10.1016/0092-8674(95)90034-9. [DOI] [PubMed] [Google Scholar]
- Simcha I., Geiger B., Yehuda-Levenberg S., Salomon D., Ben-Ze'ev A. Suppression of tumorigenicity by plakoglobinan augmenting effect of N-cadherin. J. Cell Biol. 1996;133:199–209. doi: 10.1083/jcb.133.1.199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinn E., Muller W., Pattengale P., Tepler I., Wallace R., Leder P. Coexpression of MMTV/v-Ha-ras and MMTV/c-myc genes in transgenic micesynergistic action of oncogenes in vivo. Cell. 1987;49:465–475. doi: 10.1016/0092-8674(87)90449-1. [DOI] [PubMed] [Google Scholar]
- Stepanova L., Leng X., Parker S.B., Harper J.W. Mammalian p50cdc37 is a protein kinase-targeting subunit of Hsp90 that binds and stabilizes Cdk4. Genes Dev. 1996;10:1491–1502. doi: 10.1101/gad.10.12.1491. [DOI] [PubMed] [Google Scholar]
- Stepanova L., Finegold M., DeMayo F., Schmidt E.V., Harper J.W. The oncoprotein kinase chaperone CDC37 functions as an oncogene in mice and collaborates with both c-myc and cyclin D1 in transformation of multiple tissues. Mol. Cell. Biol. 2000;20:4462–4473. doi: 10.1128/mcb.20.12.4462-4473.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutherland R., Prall O., Watts C., Musgrove E.A. Estrogen and progestin regulation of cell cycle progression. J. Mammary Gland Biol. Neoplasia. 1998;3:63–72. doi: 10.1023/a:1018774302092. [DOI] [PubMed] [Google Scholar]
- Tetsu O., McCormick F. Beta-catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature. 1999;398:422–426. doi: 10.1038/18884. [DOI] [PubMed] [Google Scholar]
- Tsukamoto A., Grosschedl R., Guzman R., Parslow T., Varmus H. Expression of the int-1 gene in transgenic mice is associated with mammary gland hyperplasia and adenocarcinomas in male and female mice. Cell. 1988;55:619–625. doi: 10.1016/0092-8674(88)90220-6. [DOI] [PubMed] [Google Scholar]
- van Genderen C., Okamura R.M., Farinas I., Quo R.-G., Parslow T.G., Bruhn L., Grosschedl R. Development of several organs that require inductive epithelial-mesenchymal interactions is impaired in Lef-1 deficient mice. Genes Dev. 1994;8:2691–2704. doi: 10.1101/gad.8.22.2691. [DOI] [PubMed] [Google Scholar]
- Wang T.C., Cardiff R.D., Zukerberg L., Lees E., Arnols A., Schmidt E.V. Mammary hyperplasia and carcinoma in MMTV-cyclin D1 transgenic mice. Nature. 1994;369:669–671. doi: 10.1038/369669a0. [DOI] [PubMed] [Google Scholar]
- Weber-Hall S.J., Phippard D.J., Niemeyer C.C., Dale T.C. Developmental and hormonal regulation of Wnt gene expression in the mouse mammary gland. Differentiation. 1994;57:205–214. doi: 10.1046/j.1432-0436.1994.5730205.x. [DOI] [PubMed] [Google Scholar]
- Williams B.O., Barish G.D., Klymkowsky M.W., Varmus H.E. A comparative evaluation of beta-catenin and plakoglobin signaling activity. Oncogene. 2000;19:5720–5728. doi: 10.1038/sj.onc.1203921. [DOI] [PubMed] [Google Scholar]
- Witcher L., Collins R., Puttagunta S., Mechanic S., Munson M., Gumbiner B., Cowin P. Desmosomal cadherin binding domains of plakoglobin. J. Biol. Chem. 1996;271:10904–10909. doi: 10.1074/jbc.271.18.10904. [DOI] [PubMed] [Google Scholar]
- Witty J.P., Wright J.H., Matrisian L.M. Matrix metalloproteinases are expressed during ductal and alveolar mammary morphogenesis, and misregulation of stromelysin-1 in transgenic mice induces unscheduled alveolar development. Mol. Biol. Cell. 1995;6:1287–1303. doi: 10.1091/mbc.6.10.1287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wong M., Rubinfeld B., Gordon J.L. Effects of forced expression of an NH2-terminal truncated β-catenin on mouse intestinal epithelial homeostasis. J. Cell Biol. 1998;141:765–777. doi: 10.1083/jcb.141.3.765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wurst W., Auerbach A.B., Joyner A.L. Engrailed-1 mutant micean early mid-hindbrain deletion and patterning defects in forelimbs and sternum. Development. 1994;120:2065–2075. doi: 10.1242/dev.120.7.2065. [DOI] [PubMed] [Google Scholar]
- Yanagawa S., Lee J., Haruna T., Oda H., Uemura T., Takeichi M., Ishimoto A. Accumulation of armadillo induced by wingless, dishevelled and dominant negative zeste-white 3 leads to elevated DE-cadherin in Drosophila clone 8 wing disc cells. J. Biol. Chem. 1997;272:25243–25251. doi: 10.1074/jbc.272.40.25243. [DOI] [PubMed] [Google Scholar]