

Abstract
Although activation of natural killer (NK) cytotoxicity is generally inhibited by target major histocompatibility complex (MHC) class I expression, subtle features of NK allorecognition suggest that NK cells possess receptors that are activated by target MHC I. The mouse Ly-49D receptor has been shown to activate NK cytotoxicity, although recognition of MHC class I has not been demonstrated previously. To define Ly-49D–ligand interactions, we transfected the mouse Ly-49D receptor into the rat NK line, RNK-16 (RNK.mLy-49D). As expected, anti– Ly-49D monoclonal antibody 12A8 specifically stimulated redirected lysis of the Fc receptor– bearing rat target YB2/0 by RNK.mLy-49D transfectants. RNK.mLy-49D effectors were tested against YB2/0 targets transfected with the mouse MHC I alleles H-2Dd, Db, Kk, or Kb. RNK.mLy-49D cells lysed YB2/0.Dd targets more efficiently than untransfected YB2/0 or YB2/0 transfected with Db, Kk, or Kb. This augmented lysis of H-2Dd targets was specifically inhibited by F(ab′)2 anti–Ly-49D (12A8) and F(ab′)2 anti–H-2Dd (34-5-8S). RNK.mLy-49D effectors were also able to specifically lyse Concanavalin A blasts isolated from H-2d mice (BALB/c, B10.D2, and DBA/2) but not from H-2b or H-2k mice. These experiments show that the activating receptor Ly-49D specifically interacts with the MHC I antigen, H-2Dd, demonstrating the existence of alloactivating receptors on murine NK cells.
Keywords: natural killer cells, major histocompatibility complex, receptors, cytotoxicity, rodent
Natural killer (NK) lymphocytes have the ability to recognize and to lyse various targets, including neoplastic and hematopoietic cells, although the specific activating receptors involved remain poorly characterized (1–4). In contrast to allogeneic killing by CD8+ T lymphocytes, NK cytotoxicity is generally inhibited, rather than activated, by the expression of class I MHC antigens on targets (5). Inhibition of natural killing is mediated by polymorphic inhibitory NK cell receptors, which are specific for distinct MHC class I alleles (6–8). Each of the known MHC-binding inhibitory NK cell receptors is a member of a family of molecules (killer cell inhibitory receptor [KIR], Ly-49, or CD94/NKG2) with related sequences (8–12). Within each family, receptors that mediate inhibitory function all contain a cytoplasmic immunoreceptor tyrosine based inhibitory motif (ITIM).1 However, each family also includes related receptors that lack ITIMs (4, 8, 13), and recent studies have shown that some of these receptors can activate NK cells (14, 15).
Although the target ligands for these stimulatory receptors have not yet been defined, their extracellular domains share homology with the corresponding domains of MHC-binding inhibitory receptors, suggesting that activating receptors may also bind MHC class I antigens (4, 8, 16–18). Consistent with this hypothesis, functional studies in mice and rats suggest that NK cells, like T cells, may express receptors that activate cytotoxicity in response to MHC class I ligands (19–22).
We have studied the mouse Ly-49D receptor, which was shown by Mason et al. to activate NK cells (14). Initial attempts to define the ligand specificity of Ly-49D in vitro were unsuccessful, although in vivo studies have implicated Ly-49D+ NK cells in the clearance of allogeneic targets (14, 23). It is likely that attempts to define MHC ligands for Ly-49D using Ly-49D–enriched primary mouse NK cells have been complicated by the coexpression of inhibitory MHC-binding NK receptors (14). To define the specificity of mouse Ly-49D, we transfected the Ly-49D receptor into the rat NK cell tumor line, RNK-16, which expresses none of the known mouse inhibitory alloreceptors. We examined the cytotoxicity of RNK.mLy-49D transfectants against a panel of targets expressing polymorphic alleles of murine MHC class I. We demonstrate here that Ly-49D specifically stimulates cytotoxicity of targets expressing H-2Dd, but not of targets expressing H-2b or H-2k MHC antigens. This H-2Dd–augmented killing can be blocked by F(ab′)2 anti–Ly-49D or F(ab′)2 anti–H-2Dd. These experiments identify Ly-49D as an activating receptor for mouse H-2Dd, and therefore demonstrate the existence of alloactivating receptors on murine NK cells.
Materials and Methods
Cells.
RNK-16, a spontaneous NK cell leukemia from F344 rats, was a gift from Craig Reynolds (NCI, Frederick, MD) and was adapted for in vitro growth in cRPMI (RPMI 1640 supplemented with 10% heat-inactivated FCS, 25 μM 2-ME, 2 mM l-glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin) (24). Tumor target cell lines cultured in cRPMI included YAC-1 (mouse lymphoma, H-2a) and YB2/0 (rat myeloma) from the American Type Culture Collection. The mouse Ly-49A transfectant of RNK-16, RNK.mLy-49A (previously described as RNK-mLy-49A.9; reference 25), was grown in cRPMI supplemented with 1 mg/ml of G418 (Boehringer Mannheim). Transfected RNK-16 effectors and transfected YB2/0 targets were maintained in 1 mg/ml G418, but were grown in cRPMI without G418 for at least 2 d before functional assays.
Antibodies and Flow Cytometry.
mAbs to mouse Ly-49D (12A8, rat IgG2a), OVA (2C7, rat IgG2a), or H-2Dd (34-5-8S, mouse IgG2a) were produced from their respective hybridoma lines. Antibodies were partially purified from ascites by ammonium sulfate precipitation. F(ab′)2 fragments were generated by pepsin digestion (26), and completion of digestion was verified by SDS-PAGE and silver staining. For fluorescence analysis, mAbs were used at a concentration of 1 μg/106 cells. Routine analysis was performed using a FACScan®.
Cytotoxicity Assays.
Specific lysis of NK targets was determined by using a standard 4-h 51Cr-release assay as previously described (27). In brief, target cells were harvested and labeled for 1 h at 37°C with 200 μCi of [51Cr]sodium chromate (Amersham) in complete RPMI. Labeled target cells were washed and resuspended at 105 cells/ml, and 0.1 ml of this cell suspension was added to each well of 96-well plates containing 0.1 ml of effector cells at the indicated E/T ratios. Plates were incubated at 37°C for 4 h, then centrifuged for 5 min. 100 μl of supernatant was counted in a gamma counter and the specific cytoxicity was calculated as previously described (27). All assays were performed in triplicate. For antibody inhibition studies, effector cells were preincubated for 15 min at room temperature with F(ab′)2 at a concentration of 25 μg/106 effectors or with intact antibody at a concentration of 10 μg/106 effectors before the addition of targets.
Vectors and cDNA.
The Ly-49D cDNA was obtained by PCR amplification from cDNA prepared from 7-d–IL-2-activated LAK cells from C57BL/6J mice using the 5′ primer 5′-TCACAGAAATCACTCAAGGACAT-3′ and the 3′ primer 5′-TTTACTTTTAACACTCAC-3′. The Ly-49D cDNA obtained is identical to the Ly-49D cDNA previously published in the extracellular and transmembrane domains (17). There is a 9-nucleotide insertion at the 3′ end of the coding sequence for the cytoplasmic domain in the Ly-49D cDNA obtained and used in these studies, which is identical to the alternately spliced long Ly-49D cDNA sequence published by Silver et al. (28). The protein sequence of the 44-amino acid cytoplasmic domain of the Ly-49D cDNA obtained reads MTEQEDTFSAVRFHKSSGLQNEMRLKETRKPEKARLRVCSVPWQ (the initial published sequence is identical except it lacks the underlined 3-amino acid insertion). This Ly-49D cDNA PCR product was subcloned into the TA cloning vector, pCR2.1 (Invitrogen), shuttled through the vector pSP72 (Promega) using the 5′ Hind III/XbaI 3′ sites, and then ligated into the XhoI site in the expression vector BSRαEN (a gift from A. Shaw and M. Olszowy, Washington University, St. Louis, MO). Constructs were confirmed by sequencing in both directions before transfection. Vectors used to transfect YB2/0 target cells with H-2Dd, H-2Db, and chimeric H-2Dd/b class I MHC have been described previously (29). The chimeric molecules were made by exon shuffling between genomic clones of H-2Dd and H-2Db using standard methods (30), and details of the construction have been described previously (29). The chimeric molecule that we have termed Ddα1α2Db encodes the α1 and α2 domains of H-2Dd with the α3 domain of H-2Db. The chimeric molecule that we have termed Dbα1α2Dd encodes the α1 and α2 domains of H-2Db with the α3 domain of H-2Dd.
Transfections.
YB2/0 or RNK-16 cells were transfected by electroporation as previously described, using cesium-purified genomic plasmids or Qiagen-purified (Qiagen) cDNA plasmids using standard methods (25, 29).
Mice.
Strain C57BL/6, BALB/c, BALB.B10 (C.B10-H-2b/ LiMcdJ), BALB.K (C.B10-H-2k/LiMcdJ), B10.D2, B10.BR, B10.S, DBA/2, and C3H mice were obtained from The Jackson Laboratory and were used at 6–8 wk of age.
Con A Blast Preparation.
Con A–stimulated blasts were prepared from the spleens of mice from the strains described above, using methods that have been described previously (31). In brief, spleens were harvested aseptically and separated into single cell suspensions. After lysis of red blood cells, cells were washed in cRPMI and placed in culture at a density of 106 cells/ml in cRPMI with 3 μg/ ml Con A (Sigma Chemical Co.). After 48 h of culture at 37°C, cells were harvested, purified over Ficoll-Hypaque, washed twice in cRPMI, and labeled for use as targets in cytotoxicity assays.
Results
Mouse Ly-49D Functions as an Activating Receptor in RNK-16 Cells.
To examine Ly-49D–ligand interactions in the absence of mouse inhibitory NK cell receptors, we transfected the mouse Ly-49D receptor into RNK-16, a rat tumor cell line with phenotypic and functional characteristics of rat NK cells (24). Cell-surface staining of a representative RNK.mLy-49D clone with the anti–Ly-49D mAb, 12A8, is shown in Fig. 1 B. In murine NK cells, the Ly-49D receptor has previously been demonstrated to mediate “redirected lysis” of FcR-bearing targets in the presence of the anti–Ly-49D mAb, 12A8 (14). As shown in Fig. 1 D, killing of the FcR-bearing target YB2/0 by the RNK. mLy-49D transfectant was similarly stimulated by anti–Ly-49D mAb, but not by isotype-matched control mAb. Neither F(ab′)2 anti–Ly-49D nor control F(ab′)2 2C7 had any effect on cytotoxicity (panel F), indicating that the binding of the intact mAb to target FcR is required for augmentation of killing of YB2/0 (redirected lysis). Lysis of YB2/0 by wild-type RNK-16 cells was unchanged by mAb anti– Ly-49D, by control mAb (panel E), or by F(ab′)2 fragments derived from either antibody (panel G). These studies demonstrate that the mouse Ly-49D receptor can function as an activating receptor in RNK-16.
Figure 1.
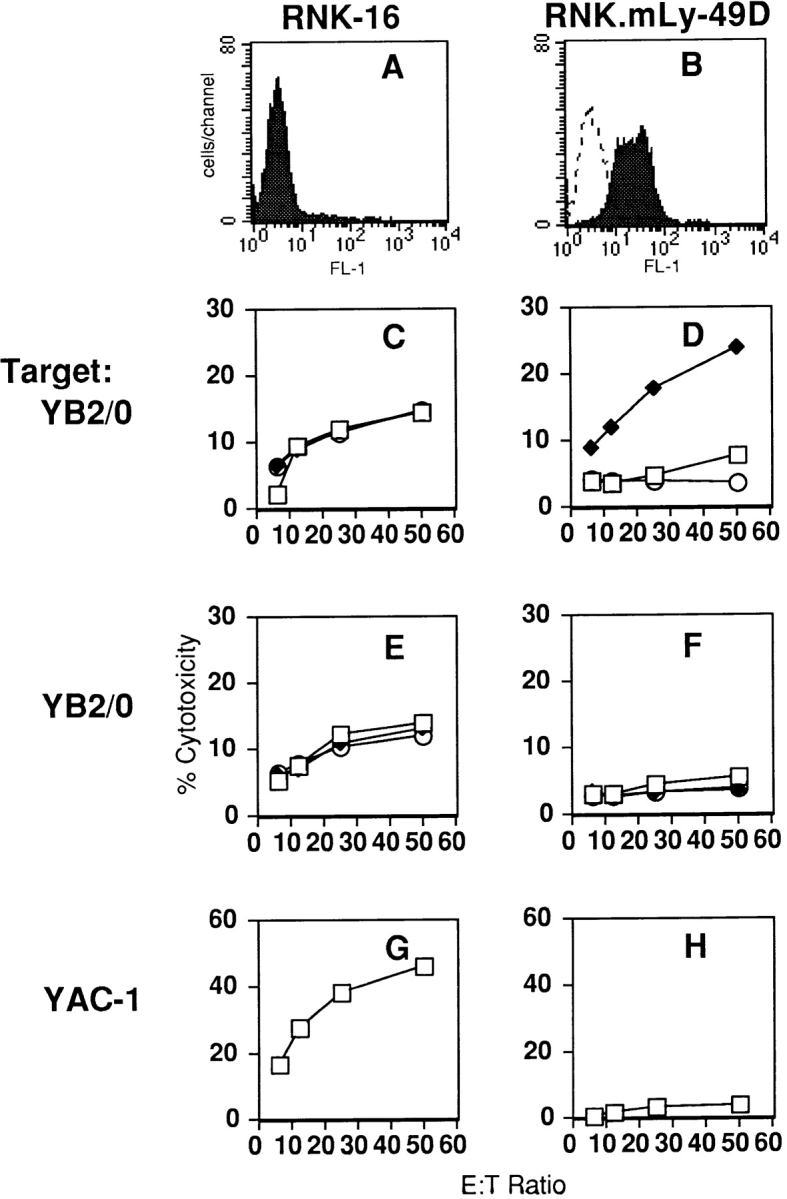
The mouse Ly-49D receptor, expressed on RNK-16 cells, mediates redirected lysis of the FcR-bearing target YB2/0 in the presence of anti–Ly-49D mAb. Staining for Ly-49D on wild-type RNK-16 cells (A) and RNK.mLy-49D transfectants (B) is shown by FACS® analysis of cells exposed to anti–Ly-49D mAb 12A8 (filled histogram) and FITC-conjugated goat anti–rat antibody (FITC-GAR) or FITC-GAR alone (dotted line). Standard 4-h cytotoxicity assays were performed with either FcR-bearing rat myeloma YB2/0 cells or YAC-1 cells as targets. RNK-16 effectors are shown on the left in C, E, and G, and RNK.mLy-49D effectors are shown on the right in D, F, and H. To measure redirected lysis by the Ly-49D receptor, effectors in C and D were preincubated with either media alone (□), intact anti–Ly-49D mAb (12A8) (♦), or intact isotype-matched control mAb (2C7) (○), before addition of YB2/0 targets. In E and F, effectors were preincubated with media alone (□), anti–Ly-49D F(ab′)2 fragments (▴), or control F(ab′)2 fragments (○). G and H show lysis of YAC-1 targets.
RNK.mLy-49D transfectants were also tested against the standard NK cell tumor target, YAC-1. Interestingly, killing of YAC-1 by RNK.mLy-49D was decreased compared with killing by RNK-16 (Fig. 1, G and H). The anti–Ly-49D mAb, 12A8, had no effect on the lysis of YAC-1 by RNK.mLy-49D transfectants (data not shown).
Mouse Ly-49D Is Activated by H-2Dd on YB2/0 Targets.
To examine the MHC restriction of Ly-49D, we tested the RNK.mLy-49D transfectant in cytotoxicity assays against a panel of rat YB2/0 myeloma target cells transfected with one of the murine H-2 alleles Dd, Db, Kb, or Kk. As seen in Fig. 2, RNK-mLy-49D effectors lysed YB2/0.Dd targets (panel A) more efficiently than YB2/0.Db (panel B), YB2/ 0.Kb (panel C), or YB2/0.Kk (panel D). Augmented killing of YB2/0.Dd by RNK.mLy-49D effectors was specifically blocked by F(ab′)2 anti–Ly-49D (12A8), but not by isotype-matched control F(ab′)2 2C7 (panel A). Lysis of the YB2/0.Dd, Db, Kb, and Kk transfectants by wild-type RNK-16 was unchanged in the presence of F(ab′)2 anti– Ly-49D or control F(ab′)2 2C7 (panels E–H). We previously described RNK-16 cells transfected with the mouse NK inhibitory receptor, Ly-49A (RNK.mLy-49A), which also recognizes H-2Dd (25). In contrast to RNK.mLy-49D cells, which were activated by YB2/0.Dd cells, RNK.mLy-49A cells were unable to lyse YB2/0.Dd cells, as they were inhibited through the Ly-49A receptor. Because the 12A8 mAb binds to Ly-49A as well as to Ly-49D (14), we were able examine the effects of 12A8 on lysis by RNK.mLy-49A cells. As shown in Fig. 2 A, F(ab′)2 12A8 blocked H-2Dd– specific activation of cytotoxicity by RNK.mLy-49D cells, but reversed H-2Dd–specific inhibition of lysis by RNK. mLy-49A cells (panel I). Thus, the activating Ly-49D receptor and the inhibitory Ly-49A receptor share ligand specificity for H-2Dd on target cells, and the interaction between ligand H-2Dd and either Ly-49A or Ly-49D can be blocked by F(ab′)2 12A8. Neither F(ab′)2 12A8 nor control F(ab′)2 2C7 had any significant effect on the lysis of YB2/0.Db, Kb, or Kk targets by any effector (panels B–D, F–H, and J–L).
Figure 2.

The Ly-49D receptor activates lysis of H-2Dd–expressing YB2/0 target cells. Standard 4-h cytotoxicity assays were performed using either RNK. mLy-49D transfectants (A–D), wild-type RNK-16 (E–H), or RNK.mLy-49A transfectants (I–L) as effectors. Each effector was tested against a panel of YB2/0 cells transfected with one of the MHC class I alleles as targets: panels A, E, and I, H-2Dd; panels B, F, and J, Db; panels C, G, and K, Kb; panels D, H, and L, Kk. Effector cells were preincubated with media alone (□), anti–Ly-49D (12A8) F(ab′)2 fragments (♦), or control (2C7) F(ab′)2 fragments (○).
RNK.mLy-49D Cells Are Activated by H-2d Con A Blasts.
We next extended our observations regarding the lysis of transfected cells by examining the lysis of Con A blasts derived from mice of various H-2 haplotypes. For these studies we tested RNK.mLy-49D effectors in cytotoxicity assays against Con A blasts prepared from MHC congenic resistant strains of BALB/c and C57BL/10 mice.
As shown in Fig. 3, RNK.mLy-49D cells effectively lysed Con A blasts prepared from BALB/c (H-2d; panel A) and B10.D2 (H-2d; panel B) mice, but not Con A blasts prepared from BALB.B10 (H-2b; panel C), C57BL/6 (H-2b; panel D), BALB.K (H-2k; panel E), or B10.BR (H-2k; panel F) mice. In experiments not shown, RNK.mLy-49D effectors also lysed Con A blasts from DBA/2 (H-2d) mice but not blasts from C3H (H-2k) or B10.S (H-2s) mice. Lysis of all H-2d blasts by RNK.mLy-49D effectors was inhibited by anti–Ly-49D mAb 12A8, but not by isotype-matched control mAb 2C7 (panels A and B).
Figure 3.
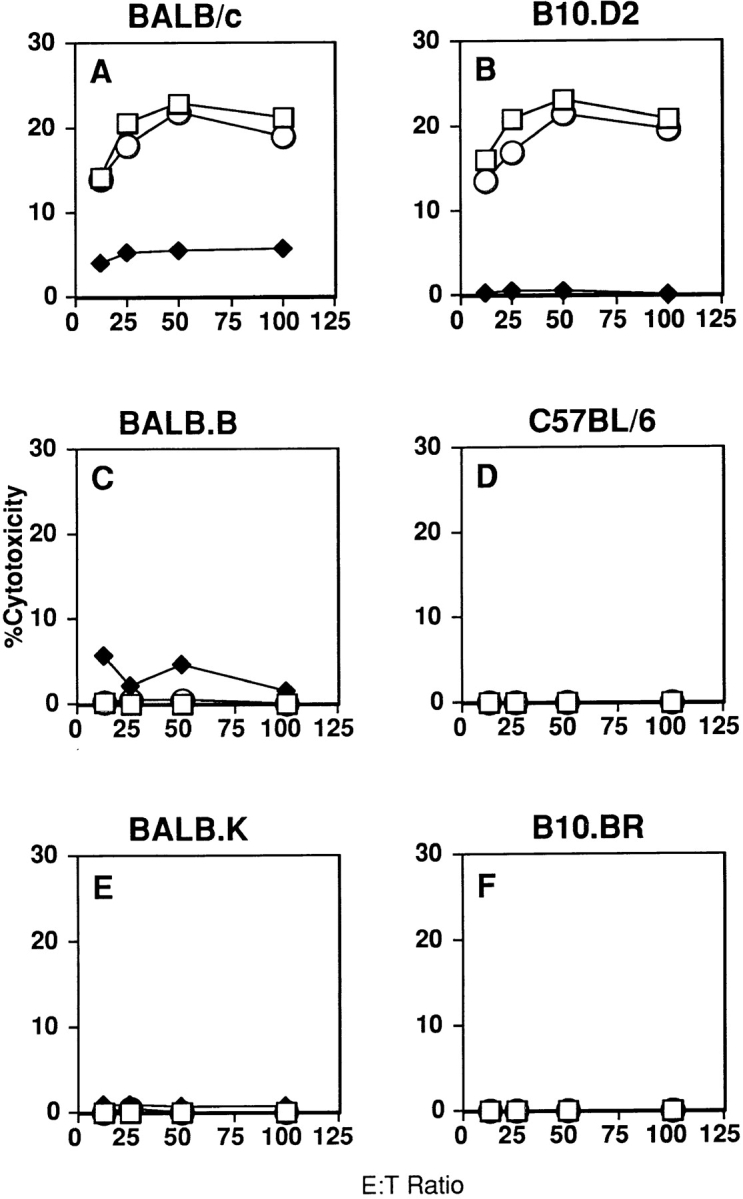
The Ly-49D receptor activates lysis of H-2d–expressing Con A blasts. RNK.mLy-49D transfectants were tested as effector cells for lysis of Con A blast targets prepared from mice that were congenic resistant at the H-2 locus. Con A blasts were prepared from splenocytes isolated from mice expressing either H-2d (BALB/c, B10.D2; panels A and B), H-2b (BALB.B, C57BL/6; panels C and D), or H-2k (BALB.K, B10.BR; panels E and F). Assays were performed after preincubation of effector cells with media alone (□), anti–Ly-49D mAb (12A8) (♦), or control mAb (2C7) (○) before addition of targets.
These studies fully supported our experiments with transfected tumor targets, identifying an H-2d–encoded structure or structures as ligands for Ly-49D. Notably, Con A blasts expressing alleles of H-2k failed to activate Ly-49D transfectants. Thus, in constrast to the Ly-49A receptor, Ly-49D does not recognize H-2Dk.
The α1/α2 Domains of H-2D d Are Required for Recognition by mLy-49D.
We next examined the domains of H-2Dd that are recognized by Ly-49D. To this end, we examined the lysis by RNK.mLy-49D effectors of YB2/0 cells transfected with genes in which exons for the α1, α2, and α3 domains of H-2Dd and H-2Db were recombined to create chimeric class I molecules. As shown in Fig. 4 C, the chimeric molecule Dbα1,2Dd, containing only the α3 domain of Dd, failed to activate cytotoxicity by RNK.mLy-49D cells, whereas the chimeric H-2 molecule Ddα1,2Db, which contains the α1,2 domains of Dd and the α3 domain of Db, activated cytotoxicity by these cells (panel F). Augmentation of RNK.mLy-49D lysis of Ddα1,2Db was inhibited by F(ab′)2 anti–Ly-49D, but not by control F(ab′)2 (panel F). As controls for the specific effect of the mAb on the Ly-49D receptor, lysis of targets expressing Dbα1,2Dd by either RNK-16 or RNK.mLy-49A cells was unchanged by F(ab′)2 12A8 or control F(ab′)2 (panels A and B). As we have previously shown, RNK.mLy-49A effectors were unable to lyse YB2/0 cells expressing Ddα1,2Db (29). However, lysis of this target by RNK.mLy-49A could be restored by the addition of blocking F(ab′)2 12A8 (panel E). Thus, both Ly-49D and Ly-49A bind to the α1 and α2 domains, but not the α3 domain, of H-2Dd.
Figure 4.

The α1/α2 region of H-2Dd is required to activate mLy-49D. RNK-16 cells (A and D), RNK.mLy-49A transfectants (B and E), and RNK.mLy-49D transfectants (C and F) were used as effectors against YB2/0 cells expressing chimeric class I molecules Dbα1,2Dd (A–C) or Ddα1,2Db (D–F) as target cells in standard 4-h cytotoxicity assays. Effector cells were preincubated with media alone (□), anti– Ly-49D (12A8) F(ab′)2 fragments (♦), or control (2C7) F(ab′)2 fragments (○).
Previous studies have shown that the mLy-49A/H-2Dd interaction can be blocked by one anti–H-2Dd antibody (34-5-8S), which is specific for a conformationally dependent epitope in the H-2Dd α1/α2 domains. Other mAbs to H-2Dd do not interrupt its interaction with Ly-49A (32). We, therefore, determined whether F(ab′)2 34-5-8S could inhibit the Ly-49D/H-2Dd interaction as well. As shown in Fig. 5, activation of RNK.mLy-49D effectors by YB2/ 0.Dd was inhibited by F(ab′)2 34-5-8S (panel C), whereas lysis of YB2/0.Dd by RNK.mLy-49A effectors was restored in the presence of F(ab′)2 34-5-8S. RNK-16 lysis of YB2/0.Dd was unaffected by F(ab′)2 34-5-8S.
Figure 5.

Activation of Ly-49D by H-2Dd is inhibited by an antibody specific for the α1/α2 domains of H-2Dd. Effector cells RNK-16 (A), RNK.mLy-49A transfectants (B), and RNK.mLy-49D transfectants (C) were tested in a standard 4-h cytotoxicity assay against YB2/0 targets expressing Dd (YB2/0.Dd) in the presence of media alone (□) or anti–H-2Dd F(ab′)2 fragments (34-5-8S) (♦).
These data show that the activating Ly-49D receptor shares some aspects of ligand recognition with the inhibitory Ly-49A receptor. Both receptors recognize the α1/α2 domains of H-2Dd, and F(ab′)2 anti–H-2Dd 34-5-8S can block the interaction of H-2Dd with either Ly-49A or Ly-49D. However, the ligand specificity of Ly-49D does not completely mirror that of Ly-49A, as Ly-49D–mediated cytotoxicity is not activated by H-2k blasts, which express the Ly-49A ligand H-2Dk (33–35). These data suggest that the structural features that restrict the allelic specificities of Ly-49D and Ly-49A are similar, but not identical.
Discussion
Cytotoxicity by NK cells is generally inhibited by the expression of class I MHC antigens on target cells. The “missing self ” hypothesis proposes that loss of target cell MHC class I may lead to the unopposed activation of natural killing and eventually to target cell lysis (5). However, several previous lines of evidence suggest that rodent NK cells may possess activating receptors in addition to inhibitory receptors for MHC-encoded target structures. First, irradiated C57BL/6 (H-2b) mice rapidly reject bone marrow from H-2Dd transgenic C57BL/6 mice, unless recipient mice are depleted of NK cells before transplantation (19, 20). Thus, the transgenic expression of H-2Dd on C57BL/6 blasts is associated with the selective acquisition of susceptibility to lysis by mouse NK cells. Second, genetic studies in rats have shown that NK cells are activated by structures encoded within the MHC locus, and NK cells from the PVG (RT1c) rat strain efficiently lyse blast targets from LEW (RT1l) rats, but not from mutant LEW.LM1 (RT1lm1) rats, which have a homozygous 100-kb deletion in the nonclassical RT1.Cl region (21). These data suggest that the ability of NK cells to lyse allogeneic blast targets may directly involve activating receptors for target MHC. A gene controlling NK alloactivating responses in rats has been linked to the Ly-49 gene family in NK alloresponder PVG rats backcrossed to DA rats, which are selectively deficient in NK allorecognition. These studies implicate Ly-49–like molecules in the activation of cytotoxicity by target MHC antigens (22).
Mason et al. previously demonstrated that Ly-49D can activate NK cell cytotoxicity, but the specific activation of Ly-49D+ NK cells by target MHC class I antigens was not demonstrated in vitro (14). Subsequent in vivo studies revealed that depletion of Ly-49D+ cells from C57BL/6 mice prevented their ability to reject H-2Dd+ bone marrow grafts (23), consistent with the hypothesis that these cells are functionally activated by MHC-encoded structures. We examined target-induced Ly-49D activation using RNK.mLy-49D transfectants, which can activate NK cell cytotoxicity through the Ly-49D receptor. Interestingly, when compared with wild-type RNK-16, RNK. mLy-49D effectors demonstrated diminished lysis of YAC-1 and YB2/0, as well as diminished antibody-dependent cellular cytotoxicity and diminished redirected lysis through the rat NKR-P1A receptor (data not shown). This change was not associated with changes in expression of NKR-P1A on the RNK-16 transfectants (data not shown). Because these changes in cytolytic specificity were seen in three different RNK.mLy-49D clones, it seems unlikely that they were unique to Ly-49D integration sites in stable transfectants, although this possibility cannot be ruled out completely. It is also possible that the overexpression of the Ly-49D activating receptor leads to sequestration of signaling intermediates required for activation, which, in turn, leads to a decrease in lysis through other activating receptors. However, we were easily able to observe specific activation through Ly-49D, demonstrating that the cytolytic capacity of RNK.mLy-49D cells was intact.
Our in vitro studies demonstrate that Ly-49D is an activating NK cell receptor specific for H-2Dd. The acquisition of enhanced cytotoxicity against H-2Dd–transfected YB2/0 targets was specifically blocked by F(ab′)2 anti–Ly-49D or by F(ab′)2 anti–H-2Dd. Activation of NK cells by Ly-49D was not unique to transfected targets, as Ly-49D also stimulated lysis against blasts from H-2d mice, but not against blasts from H-2b or H-2k mice. We considered the possibility that these results might be obtained if Ly-49D were not the activation receptor itself but instead, through its interaction with H-2Dd, facilitated activation through a separate activating receptor. However, it is unlikely that adhesion alone between Ly-49D and H-2Dd accounts for the observed activation of cytotoxicity. First, stimulation of the Ly-49D receptor with antibody is known to induce activation of cytotoxicity by NK cells, as shown here and by others (14). Second, we have previously demonstrated that adhesion between another Ly-49 receptor and H-2Dd is not sufficient to activate NK cell lysis. Specifically, we have previously demonstrated that interaction of an inactive Ly-49A receptor with H-2Dd does not activate NK cell lysis through other receptors on RNK-16 (25). The Ly-49A receptor normally binds to H-2Dd and thereby leads to inhibition of NK cell lysis. Our previous studies showed inhibition of lysis of H-2Dd expressing targets by RNK-16 cells transfected with Ly-49A. We also studied a mutated Ly-49A receptor, containing a point mutation in the cytoplasmic domain of the receptor which disrupts the ITIM motif required for inhibitory function. This mutated Ly-49A receptor expressed on RNK-16 cells failed to inhibit or to augment lysis of H-2Dd–expressing targets. This mutated Ly-49A receptor is unaltered in the extracellular or transmembrane domains so its binding to H-2Dd is presumably unaltered. Thus, engagement of H-2Dd by the mutant Ly-49A receptor does not facilitate the activation of lytic pathways through other receptors on RNK-16.
It is of considerable interest whether the inhibitory Ly-49A receptor and the activating Ly-49D receptor recognize the same or distinct domains of the H-2Dd antigen. As with Ly-49A, Ly-49D–dependent effects required the α1/α2 domains of H-2Dd and could be blocked by an α1/α2-specific F(ab′)2 anti–H-2Dd mAb (34-5-8S) (references 32–34, 36). The extracellular domains of Ly-49A and Ly-49D may recognize similar regions of H-2Dd, but only Ly-49A recognizes H-2Dk (33–35). Thus, Ly-49A and Ly-49D appear to exhibit overlapping but distinct allelic specificities for murine class I MHC antigens. Studies by several groups have indicated that Ly-49A does not discriminate between different peptides presented by H-2Dd (32, 37). The role of antigenic peptides in the specificity of Ly-49D–H-2Dd interaction remains to be examined.
It has been speculated that activating Ly-49–like receptors may participate in the recognition of abnormal or virally-encoded MHC-like molecules by NK cells, although this has not been proven (6, 38). However, the physiologic significance of alloactivating NK receptors is unknown. Although their effects may be partially masked by inhibitory NK cell receptors, the identification of in vivo NK alloactivating functions indicates that alloactivation is not completely abrogated by inhibitory MHC receptors in vivo (19–22). A delicate balance between activating and inhibitory alloreceptors may be influenced by different affinities or structural requirements for ligand binding. Alternatively, expression of opposing alloreceptors on different NK cell subsets may permit NK alloresponses in vivo. Alloactivating NK cell receptors, in addition to allospecific T cell receptors, likely play a role in the physiologic rejection of foreign cells (6, 7, 23, 39).
In conclusion, our experiments using gene transfer provide the first direct evidence that an activating receptor on murine NK cells specifically directs cytotoxicity against a classical class I MHC ligand. They also provide a model for dissecting the molecular pathways involved in this interaction. Other candidate alloactivating receptors on human NK cells include the Ig-like short domain KIR (killer cell inhibitory receptor) receptors, which may recognize alleles of HLA-C (8, 15, 16), and the activating CD94/NKG2C receptor, which can bind to ligand HLA-E (11). Future studies to define the distribution and activation requirements for these receptors will help us to understand the physiologic functions of alloactivating receptors in NK cell biology.
Acknowledgments
This work was supported by the Veterans Administration and by National Institutes of Health (NIH) grant RO1 CA69299 (to W.E. Seaman). J.C. Ryan is the recipient of NIH grant R29 CA60944 and is supported by the International Human Frontiers in Science Program. M.C. Nakamura is supported by NIH grant K11 AR01927, the Rosalind Russell Arthritis Foundation, and NIH Multipurpose Arthritis Center grant P60 AR20684.
Abbreviation used in this paper
- ITIM
immunoreceptor tyrosine based inhibitory motif
Footnotes
The first two authors contributed equally to this work.
References
- 1.Trinchieri G. Biology of natural killer cells. Adv Immunol. 1989;47:187–376. doi: 10.1016/S0065-2776(08)60664-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lanier LL. NK cell receptors. Adv Immunol. 1998;16:359–393. doi: 10.1146/annurev.immunol.16.1.359. [DOI] [PubMed] [Google Scholar]
- 3.Leibson PJ. Signal transduction during natural killer cell activation: inside the mind of a killer. Immunity. 1997;6:655–661. doi: 10.1016/s1074-7613(00)80441-0. [DOI] [PubMed] [Google Scholar]
- 4.Ryan JC, Seaman WE. Divergent functions of lectin-like receptors on NK cells. Immunol Rev. 1997;155:79–89. doi: 10.1111/j.1600-065x.1997.tb00941.x. [DOI] [PubMed] [Google Scholar]
- 5.Ljunggren H-G, Kärre K. In search of the “missing self ” MHC molecules and NK cell recognition. Immunol Today. 1990;11:237–244. doi: 10.1016/0167-5699(90)90097-s. [DOI] [PubMed] [Google Scholar]
- 6.Rolstad B, Vaage JT, Naper C, Lambracht D, Wonigeit K, Joly E, Butcher GW. Positive and negative MHC class I recognition by rat NK cells. Immunol Rev. 1997;155:91–104. doi: 10.1111/j.1600-065x.1997.tb00942.x. [DOI] [PubMed] [Google Scholar]
- 7.Höglund P, Sundbäck J, Olsson-Alheim MY, Johansson M, Salcedo M, Öhlén C, Ljunggren H-G, Sentman CL, Kärre K. Host MHC class I gene control of NK-cell specificity in the mouse. Immunol Rev. 1997;155:11–28. doi: 10.1111/j.1600-065x.1997.tb00936.x. [DOI] [PubMed] [Google Scholar]
- 8.Long EO, Burshtyn DN, Clark WP, Peruzzi M, Rajagopalan S, Rojo S, Wagtmann N, Winter CC. Killer cell inhibitory receptors: diversity, specificity and function. Immunol Rev. 1997;155:135–144. doi: 10.1111/j.1600-065x.1997.tb00946.x. [DOI] [PubMed] [Google Scholar]
- 9.Takei F, Brennan J, Mager DL. The Ly-49 family: genes, proteins and recognition of class I MHC. Immunol Rev. 1997;155:67–77. doi: 10.1111/j.1600-065x.1997.tb00940.x. [DOI] [PubMed] [Google Scholar]
- 10.Lazetic S, Chang C, Houchins JP, Lanier LL, Phillips JH. Human NK cell receptors involved in MHC class I recognition are disulfide linked heterodimers of CD94 and NKG2 subunits. J Immunol. 1996;157:4741–4745. [PubMed] [Google Scholar]
- 11.Braud VM, Allan DSJ, O'Callaghan CA, Soderstrom K, D'Andrea A, Ogg GS, Lazetic S, Young NT, Bell JI, Phillips HH, et al. HLA-E binds to natural killer cell receptors CD94/NKG2A, B, and C. Nature. 1998;391:795–798. doi: 10.1038/35869. [DOI] [PubMed] [Google Scholar]
- 12.Borrego F, Ulbrecht M, Weiss EH, Coligan JE, Brooks AG. Recognition of human histocompatibility leukocyte antigen (HLA)-E complexed with HLA class I signal sequence–derived peptides by CD94/NKG2 confers protection from natural killer cell–mediated lysis. J Exp Med. 1998;187:813–818. doi: 10.1084/jem.187.5.813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Houchins JP, Yabe T, McSherry C, Bach FH. DNA sequence analysis of NKG2, a family of related cDNA clones encoding type II integral membrane proteins on natural killer cells. J Exp Med. 1991;173:1017–1020. doi: 10.1084/jem.173.4.1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Mason LH, Anderson SK, Yokoyama WM, Smith HRC, Winkler-Pickett R, Ortaldo JR. The Ly-49D receptor activates murine natural killer cells. J Exp Med. 1996;184:2119–2128. doi: 10.1084/jem.184.6.2119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Moretta A, Sivori S, Vitale M, Pende D, Morelli L, Augugliaro R, Bottino C, Moretta L. Existence of both inhibitory (p58) and activatory (p50) receptors for HLA-C molecules in human natural killer cells. J Exp Med. 1995;182:875–884. doi: 10.1084/jem.182.3.875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Biassoni R, Cantoni C, Falco M, Verdiani S, Bottino C, Vitale M, Conte R, Poggi A, Moretta A, Moretta L. The human leukocyte antigen (HLA)-C–specific “activatory” or “inhibitory” natural killer cell receptors display highly homologous extracellular domains but differ in their transmembrane and intracytoplasmic portions. J Exp Med. 1996;183:645–650. doi: 10.1084/jem.183.2.645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Smith HRC, Karlhofer FM, Yokoyama WM. Ly-49 multigene family expressed by IL-2-activated NK cells. J Immunol. 1994;153:1068–1079. [PubMed] [Google Scholar]
- 18.Wong S, Freeman JD, Kelleher C, Mager D, Takei F. Ly-49 multigene family: new members of a supergene family of type II membrane proteins with lectin-like domains. J Immunol. 1991;147:1417–1423. [PubMed] [Google Scholar]
- 19.Öhlén C, Kling G, Höglund P, Hansson M, Scangos G, Bieberich C, Jay G, Kärre K. Prevention of allogeneic bone marrow graft rejection by H-2 transgene in donor mice. Science. 1989;246:666–668. doi: 10.1126/science.2814488. [DOI] [PubMed] [Google Scholar]
- 20.Yu YYL, Forman J, Aldrich C, Blazar B, Flaherty L, Kumar V, Bennett M. Natural killer cells recognize common antigenic motifs shared by H-2Dd, H-2Ld and possibly H-2Drmolecules expressed on bone marrow cells. Int Immunol. 1994;6:1297–1306. doi: 10.1093/intimm/6.9.1297. [DOI] [PubMed] [Google Scholar]
- 21.Vaage JT, Naper C, Lovik G, Lambracht D, Rehm A, Hedrich HJ, Wonigeit K, Rolstad B. Control of rat natural killer cell-mediated allorecognition by a major histocompatibility complex region encoding nonclassical class I antigens. J Exp Med. 1994;180:641–651. doi: 10.1084/jem.180.2.641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Dissen E, Ryan JC, Seaman WE, Fossum S. An autosomal dominant locus, Nka, mapping to the Ly-49 region of a rat natural killer (NK) gene complex, controls NK cell lysis of allogeneic lymphocytes. J Exp Med. 1996;183:2197–2207. doi: 10.1084/jem.183.5.2197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Raziuddin A, Longo DL, Mason L, Ortaldo JR, Bennett M, Murphy WJ. Differential effects of the rejection of bone marrow allografts by the depletion of activating versus inhibiting Ly-49 NK cell subset. J Immunol. 1998;160:87–94. [PubMed] [Google Scholar]
- 24.Reynolds CW, Bere EW, Jr, Ward JM. Natural killer activity in the rat. III. Characterization of transplantable large granular lymphocyte (LGL) leukemias in the F344 rat. J Immunol. 1984;132:534–540. [PubMed] [Google Scholar]
- 25.Nakamura MC, Niemi EC, Fisher MJ, Shultz LD, Seaman WE, Ryan JC. Mouse Ly-49A interrupts early signaling events in NK cell cytotoxicity and functionally associates with the SHP-1 tyrosine phosphatase. J Exp Med. 1997;185:673–684. doi: 10.1084/jem.185.4.673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Coligan JE, Kruisbeck AM, Margulies DH, Strober W. Large scale fragmentation of IgG to F(ab′)2using pepsin. Curr Protocols Immunol. 1994;1:2.8.6. [Google Scholar]
- 27.Ryan JC, Niemi EC, Goldfien RD, Hiserodt JC, Seaman WE. NKR-P1, an activating molecule on rat natural killer cells, stimulates phosphoinositide turnover and a rise in intracellular calcium. J Immunol. 1991;147:3244–3250. [PubMed] [Google Scholar]
- 28.Silver ET, Elliott JF, Kane KP. Alternatively spliced Ly-49D and H transcripts are found in IL-2 activated NK cells. Immunogenetics. 1996;44:478–482. doi: 10.1007/BF02602811. [DOI] [PubMed] [Google Scholar]
- 29.Sundbäck J, Nakamura MC, Waldenström M, Niemi EC, Seaman WE, Ryan JC, Kärre K. The α2 domain of H-2Ddrestricts the allelic specificity of the murine NK cell inhibitory receptor Ly-49A. J Immunol. 1998;160:5971–5978. [PubMed] [Google Scholar]
- 30.Sambrook, J., E.F. Fritsch, T. Maniatis. 1989. Molecular Cloning: A Laboratory Manual, 2nd Edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- 31.Yu YYL, George T, Dorfman JR, Roland J, Kumar V, Bennett M. The role of Ly49A and 5E6(Ly49C) molecules in hybrid resistance mediated by murine natural killer cells against normal T cell blasts. Immunity. 1996;4:67–76. doi: 10.1016/s1074-7613(00)80299-x. [DOI] [PubMed] [Google Scholar]
- 32.Orihuela M, Margulies DH, Yokoyama WM. The natural killer cell receptor Ly-49A recognizes a peptide-induced conformational determinant on its major histocompatibility complex class I ligand. Proc Natl Acad Sci USA. 1996;93:11792–11797. doi: 10.1073/pnas.93.21.11792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Karlhofer FM, Ribaudo RK, Yokoyama WM. MHC class I alloantigen specificity of Ly-49+IL-2- activated natural killer cells. Nature. 1992;358:66–70. doi: 10.1038/358066a0. [DOI] [PubMed] [Google Scholar]
- 34.Kane KP. Ly-49 mediates EL-4 lymphoma adhesion to isolated class I major histocompatibility complex molecules. J Exp Med. 1994;179:1011–1015. doi: 10.1084/jem.179.3.1011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Brennan J, Mager D, Jefferies W, Takei F. Expression of different members of the Ly-49 gene family defines distinct natural killer cell subsets and cell adhesion properties. J Exp Med. 1994;180:2287–2295. doi: 10.1084/jem.180.6.2287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Daniels BF, Karlhofer FM, Seaman WE, Yokoyama WM. A natural killer cell receptor specific for a major histocompatibility complex class I molecule. J Exp Med. 1994;180:687–692. doi: 10.1084/jem.180.2.687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Correa I, Raulet DH. Binding of diverse peptides to MHC class I molecules inhibits target cell lysis by activated natural killer cells. Immunity. 1995;2:61–71. doi: 10.1016/1074-7613(95)90079-9. [DOI] [PubMed] [Google Scholar]
- 38.Scalzo AA, Lyons PA, Fitzgerald NA, Forbes CA, Yokoyama WM, Shellam GR. Genetic mapping of Cmv1in the region of mouse chromosome 6 encoding the NK gene complex-associated loci Ly-49 and musNKR-P1. Genomics. 1995;27:435–441. doi: 10.1006/geno.1995.1074. [DOI] [PubMed] [Google Scholar]
- 39.Loke YW, King A. Immunology of human placental implantation: clinical implications of our current understanding. Mol Med Today. 1997;3:153–159. doi: 10.1016/s1357-4310(97)01011-3. [DOI] [PubMed] [Google Scholar]