

Abstract
The goal of this study was to determine the mechanisms by which dendritic cells (DCs) in blood could interact with endothelium, a prerequisite to extravasation into tissues. Our results indicate that DCs express both HECA-452–reactive and nonreactive isoforms of P-selectin glycoprotein ligand 1 (PSGL-1) and can tether and roll efficiently on E- and P-selectin under flow conditions in vitro. Freshly isolated blood DCs were further observed to roll continuously along noninflamed murine dermal endothelium in vivo. This interaction is strictly dependent on endothelial selectins, as shown by experiments with blocking antibodies and with E- and P-selectin–deficient mice. We hypothesize that DCs in blood are constitutively poised at the interface of blood and skin, ready to extravasate upon induction of inflammation, and we showed that cutaneous inflammation results in a rapid recruitment of DCs from the blood to tissues. We propose that this is an important and previously unappreciated element of immunosurveillance.
Keywords: inflammation, immunosurveillance, selectins, rolling, extravasation
In the years since their initial description, the central role of dendritic cells (DCs)1 in the development of acquired immune responses has become widely accepted (1, 2). Tissue DCs, particularly those in tissues which constitute epithelial interfaces with the environment, capture antigens and migrate to lymphoid organs, where they present antigens to T cells (1, 2). Another population of DCs also exists in peripheral blood, where they represent ∼0.5% of the circulating PBMCs (3). Extensive study of blood DCs has been limited by their paucity. This population expresses HLA-DR and lacks specific lineage markers or surface activation molecules found on other leukocytes. Although they lack the typical dendritic morphology of mature DCs, they can rapidly acquire these features in culture (3, 4).
A physiological role for blood DCs is unclear, but at least some of these cells are hypothesized to be en route to tissues (3, 4). As such, they must be capable of initiating interactions with vascular endothelium while moving in the blood flow. However, this hypothesis has never been directly tested and the mechanisms regulating this putative process are undetermined (3). The extravasation of leukocytes across endothelia has been described as a multistep cascade of discrete events (5, 6). The initial adhesive step, which involves binding in shear flow, has been associated with a limited subset of surface molecules, including the selectins and the α4 integrins and their respective ligands. In this study, we examine the capacity of blood DCs to complete the initial step involved in leukocyte homing by tethering and rolling under flow, both in vitro and in vivo along the skin vessel surface. We further test the capacity of DCs to extravasate in vivo in the setting of inflammation.
Materials and Methods
Isolation of Fresh Blood DCs.
PBMCs were isolated by density gradient separation from blood cells collected from normal donors during platelet pheresis. Fresh blood DCs were isolated from PBMCs as described previously (7) or using a commercial blood DC isolation kit (Miltenyi Biotec). Both methods resulted in very pure populations of DCs (HLA-DR+ TCR-α/β−, CD14−, CD56−, and CD19−; see Fig. 1). Viability was >95% by trypan blue exclusion. Fresh blood DCs isolated by either method were used within 4 h of purification.
Figure 1.

Blood DCs express HLA-DR but do not express specific lineage markers CD14, CD19, or CD56 or any TCR (reference 3). (a) They represent <1% of the PBMCs (reference 3). After immunomagnetic depletion of T cells, monocytes, and NK cells, using CD3, CD11b, and CD16 antibodies and positive selection of CD4+ leukocytes (blood DC isolation kit from Miltenyi Biotec), a population of >90% pure blood DCs is obtained (b).
Isolation and Culture of Human CD34+ Hematopoietic Progenitor Cells.
When our experiments required quantities of DCs beyond our means to isolate directly from blood (i.e., for the molecular characterization of selectin ligands expressed on DCs), we used immature cultured DCs, derived from cord blood progenitors (8). CD34+ progenitors were isolated from the mononuclear fraction of heparinized umbilical cord blood samples by magnetic separation (CD34+ separation kit; Miltenyi Biotec). Cell preparations were routinely >90% CD34+ by FACS® with viability approaching 100%. Cultures of CD34+ progenitors were established by seeding at 105 cells/ml in RPMI 1640 (GIBCO BRL) supplemented with 10% fetal bovine serum, recombinant human (rh)GM-CSF (1,000 U/ml; Immunex Corp.), IL-4/TNF-α (50 U/ml each, R & D Systems), Flt3 ligand (100 ng/ml; Immunex Corp. and R & D Systems), 10 mM Hepes, 2 mM l-glutamine, 5 × 10−5 M 2-ME, penicillin (100 U/ml), and streptomycin (100 μg/ml). Clusters of nonadherent cells with dendritic morphology appeared after 4–5 d of culture and increased in number and size in the following days. We routinely observed a 10–150-fold increase in total cell number after 2–3 wk of culture. Cells were used on days 10–14 when cultures contained cells with dendritic phenotypic characteristics and surface markers (72–85% HLA-DR+, 30–80% CD1a+, 40–50% CD80+, 30–40% CD83+, >95% CD14−).
Preparation of Murine Bone Marrow–derived DCs.
Bone marrow– derived DCs were prepared as published previously (9). In brief, bone marrow cells from FVB mice were depleted of red cells by lysis in ACK lysing buffer (0.15 M NH4Cl, 1 mM KHCO3, 0.1 mM Na2EDTA, pH 7.3), and the cultures were established in RPMI 1640 (GIBCO BRL) supplemented with 10% fetal bovine serum with recombinant murine (rm)GM-CSF (400 U/ml) and rmIL-4 (150 U/ml; PharMingen). DCs were used between 4 and 6 d after bone marrow harvest. Flow cytometry was performed immediately before the homing experiments and showed immature phenotype DCs expressing class II I-A antigen and moderate amounts of CD40 and B7-2.
FACS® Analysis.
mAbs used included HECA-452 (PharMingen), anti–human P-selectin glycoprotein ligand 1 (PSGL-1) (PSL-275 and 4H10; gifts from the Genetics Institute, Cambridge, MA), anti-CD14, anti-CD19, anti-CD56, and anti–TCR-α/β (Coulter Corp.), anti–mouse I-A, anti-CD40, and anti–B7-2 (PharMingen). Nonbinding isotype-matched antibodies were used as control reagents. Flow cytometry was performed on a FACScan® IV (Becton Dickinson) using CellQuest software (version 1.2). Results are representative of multiple independent experiments.
Immunoblotting.
HECA-immunoreactive DCs were positively selected from CD34+-derived DCs after 9–10 d of culture using magnetic microbeads (Miltenyi Biotec) as published previously (10). Methods for preparing cell lysates from cultured DCs and for immunoblotting under normal and enhanced reducing conditions were as described previously (10) with the following modifications: the anti–PSGL-1 mAb PL1 (mIgG1; Immunotech) was used at a concentration of 2 μg/ml. HECA-452 (a gift from Dr. L. Picker, University of Texas/Southwestern, Dallas, TX [11]) was used at 1.2 μg/ml (rather than 2 μg/ml). Immunoblots were prepared using 300 μg of HECA-immunoreactive DC lysate protein per lane for HECA-452 blots and 600 μg of unselected DCs per lane for PSGL-1 blots. Enhanced reduction of cell lysates was carried out for 5–8 d, rather than 3 d, before SDS-PAGE.
In Vitro Flow Analysis.
For E- and P-selectin binding analysis, cells were analyzed in a parallel plate flow chamber using protein A–bound E- or P-selectin IgG chimeras as described previously (12). Washed cells were resuspended in medium with 2 mM Ca2+ immediately before use, perfused into the chamber, and allowed to interact under static conditions for 3 min. Flow was initiated at a wall shear stress of 1 dyn/cm2, and interactions were observed for an additional 1 min. Shear was subsequently increased in a stepwise fashion every 10–30 s, and the percentage of rolling cells remaining bound was determined at each step. Nonspecific binding was defined as cells remaining bound in 5 mM EDTA perfused at 50 dyn/cm2. The percentage of cells able to form adhesions was calculated as the number of cells remaining bound at 1 dyn/cm2 after a 3-min static incubation, divided by the number of cells present just before reinitiating flow. The number of cells attached/μm2/min was determined by counting the number of new tethers observed in the field during 1 min of flow at 1 dyn/cm2.
Intravital Microscopy.
Normal adult Swiss Webster mice, mice deficient in both E- and P-selectin (E/P-selectin−/−; reference 13), and control wild-type mice from the same strain as the deficient mice (C57BL/129SV) were anesthetized by intraperitoneal injection of saline (10 ml/kg) containing 5 mg/ml ketamine and 1 mg/ml xylazine. The hair on the left ear as well as on the submandibular area of the neck was removed using hair removal lotion. A PE-10 polyethylene catheter was inserted into the right common carotid artery of a thermo-controlled mouse whose left ear was covered with glycerol and gently positioned between a microscope slide and a coverslip under an intravital microscope (model IV-500; Mikron Instruments). DCs were fluorescently labeled (2.5 μg calcein/ml/107 cells; Molecular Probes, Inc.) and introduced by retrograde injection into the right carotid artery. Fluorescent DCs in ear microvessels were visualized by stroboscopic fluorescent epi-illumination using infinity-corrected water-immersion optics (Carl Zeiss) and a silicon-intensified target camera (Dage). Rolling fractions were determined as the percentage of interacting cells in the total flux of fluorescent DCs that passed through each venule during the same period. The velocities of individual rolling cells were determined by off-line analysis of videotapes using a PC-based interactive image analysis system (14). Hemodynamic parameters were determined in 3 animals, 4 venules, and 82 noninteracting cells (12, 12, 28, and 30 fast cells per venule) for cultured DCs in wild-type mice; in 3 mice, 3 venules, and 96 cells (31, 32, and 33 per venule) for cultured DCs in E/P-selectin−/− mice; and in 2 wild-type mice, 3 venules, and 55 cells (18, 18, and 19 per venule) for freshly isolated DCs. Anti–mouse E- and P-selectin blocking antibodies, 9A9 and 5H1, respectively (from B. Wolitzky, Hoffman-La Roche, Inc., Nutley, NJ), were used at 100 μg/mouse and injected into the blood circulation 5 min before the infusion of the cells.
Homing of DCs during a Hypersensitivity Contact Reaction in Mice.
FVB mice were sensitized with a classical contact sensitizer, oxazolone (100 μl of 2% oxazolone in acetone on abdominal skin) 6 d before challenge with 10 μl of 0.8% oxazolone on both sides of the right ear. The left ear was not treated and is referred to as the control ear. Immature bone marrow–derived DCs were labeled with 51Cr (NEN) as follows: 107 DCs/ml were resuspended in RPMI containing 20% FCS and incubated for 1 h with 200–400 μl of [51Cr] sodium chromate (1 mCi/ml) at 37°C in 5% CO2. The cells were washed twice in RPMI, and 5 × 106 DCs/mouse were infused into the tail vein 48 h after the challenge was applied. The mean specific radioactivity of injected DCs was 288 cpm/103 cells (range 88–620). 6 h after DC infusion, the mice were killed, and the radioactivities of both ears were counted in a gamma counter.
Calcein-labeled DCs were also used in similar experiments. 100 μg of PE-conjugated CD31 mAb (PharMingen) was infused into the tail veins of mice 5 min before they were killed in order to visualize the vessels by confocal microscopy. Ears were split in two halves parallel to their broad surfaces using fine forceps, put between a microscope slide and a coverslip, and examined using a BioRad MRC-1024 confocal imaging system.
Results
DCs Uniformly Express the Selectin Ligand PSGL-1/Cutaneous Lymphocyte-associated Antigen.
Magnetic separation of blood DCs allowed the recovery of >85% pure DCs, as defined by the expression of HLA-DR and the lack of specific lineage markers CD14, CD56, CD19, and TCR-α/β (Fig. 1). We examined DC expression of three leukocyte cell surface molecules known to mediate binding under physiologic flow conditions: PSGL-1, L-selectin, and the integrin α4β1. Roughly 50% of freshly isolated DCs expressed L-selectin (not shown), whereas α4β1 (not shown) and PSGL-1 (Fig. 2 a) were expressed by 100% of the cells. We had previously shown that PSGL-1 on T cells could be expressed in an uniquely glycosylated form that reacts with an oligosaccharide-specific antibody called HECA-452 and binds to both E- and P-selectin under flow (10, 11, 15). When expressed on memory T cells, this isoform of PSGL-1 is known as cutaneous lymphocyte-associated antigen (CLA [10]) and is thought to direct the extravasation of T cells into inflamed skin.
Figure 2.


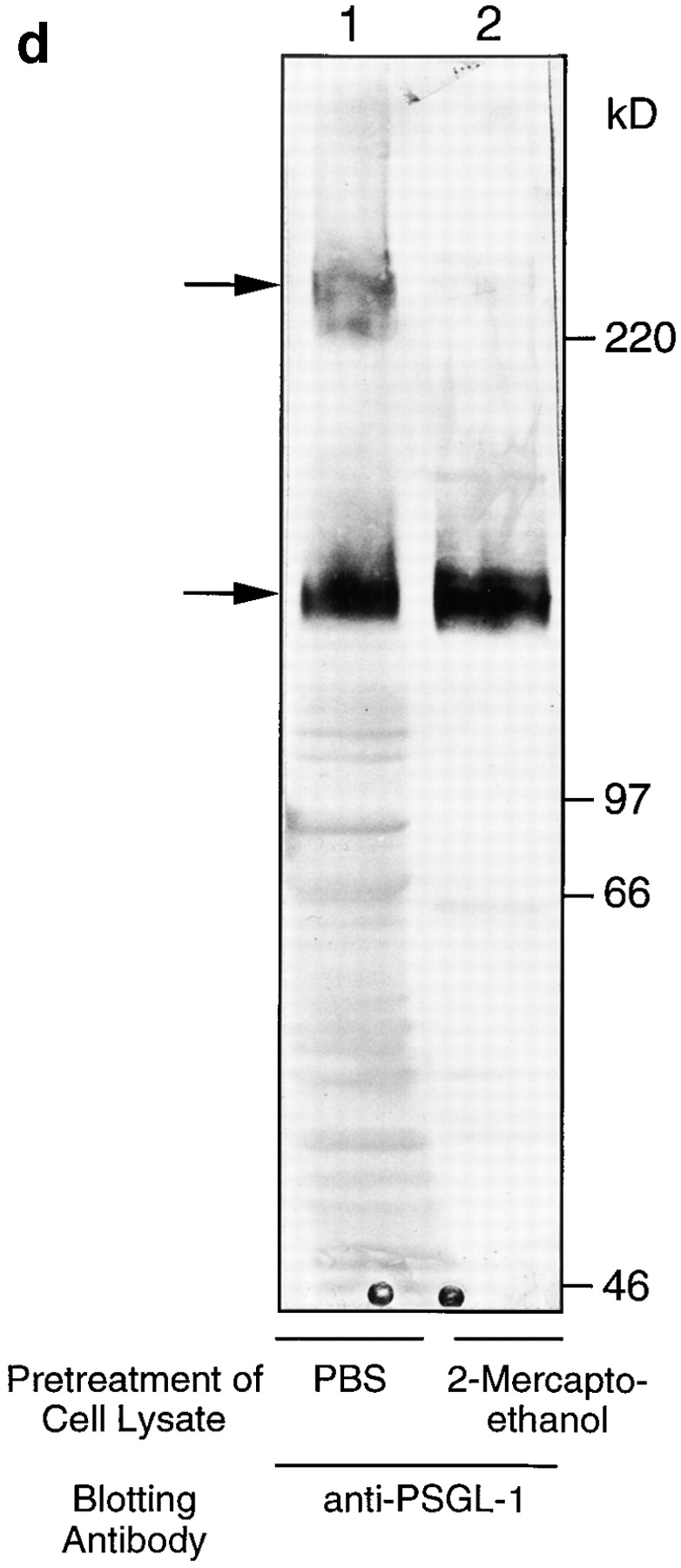
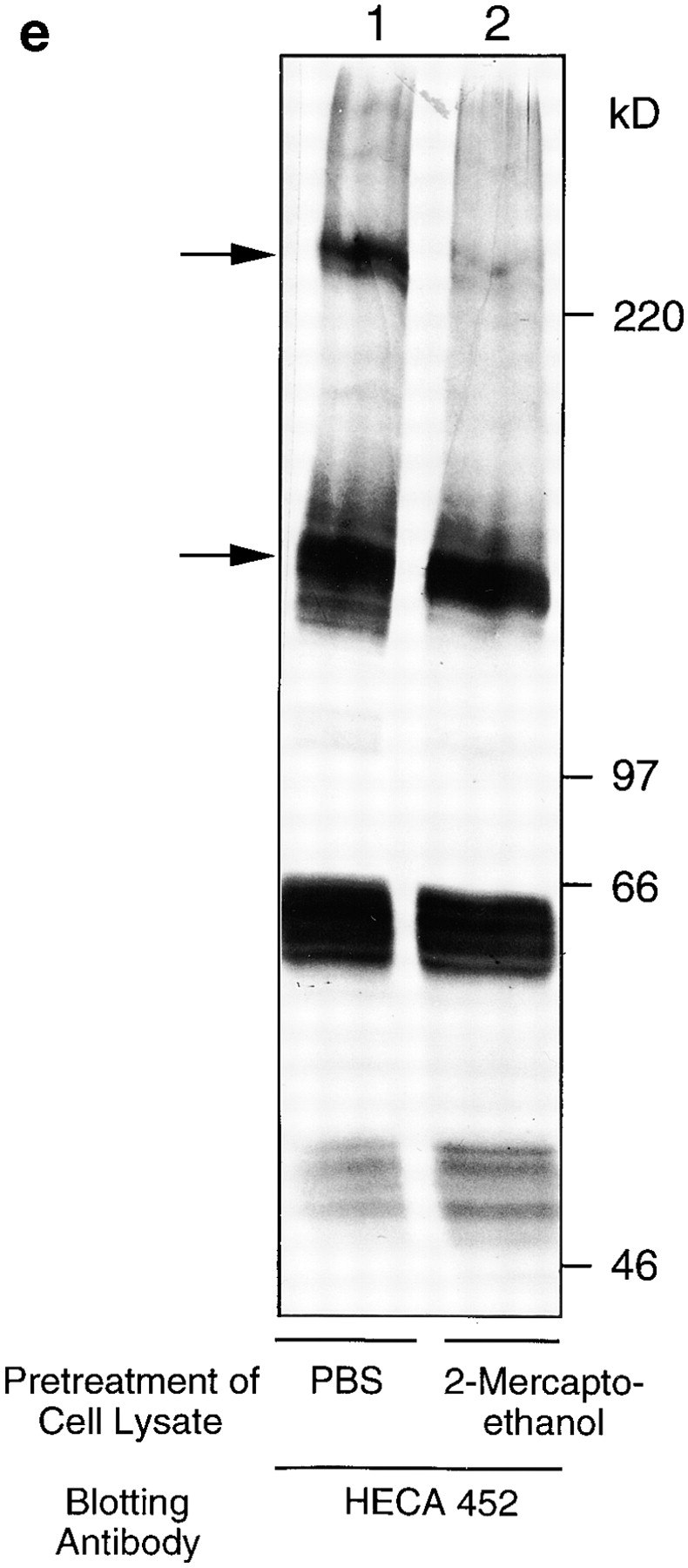
(a and b) Freshly isolated DCs homogeneously express P- and E-selectin ligand, PSGL-1/CLA. FACS® analysis of cultured DCs stained with mAb to PSGL-1 (PSL-275) and to CLA (HECA-452). (c) Cultured DCs express PSGL-1 (4H10), and 30% (30–50%) of them also express CLA. (d and e) Immunoblotting of cultured DC lysates with anti–PSGL-1 (d) and HECA-452 (e) shows that the CLA epitope is expressed on a single glycoprotein. Analysis under standard reducing conditions (lane 1) revealed specific immunoreactive bands at 240 kD (dimer) and 140 kD (monomer) with each specific antibody; and under enhanced reducing conditions (lane 2), only one specific band at 140 kD (monomer) was seen for both HECA-452 and anti– PSGL-1 immunoreactivities. The bands at 50 and 60 kD in the HECA-452 blot represent the HECA antibody used for immunomagnetic separation, as shown previously (reference 10).
Although HECA-452 immunoreactivity has been shown on Langerhans cells, a subset of DCs (16, 17), neither the structure of the molecules carrying this oligosaccharide epitope nor its functional capability as a selectin ligand on DCs has been reported. FACS® analysis was performed using 4H10 or PSL-275, two mAbs that recognize peptide determinants in the P-selectin binding domain of PSGL-1, and HECA-452, an IgM mAb that recognizes a carbohydrate epitope similar to sialyl Lewis X (18) and defines CLA expression on T cells (11). PSGL-1 and CLA were expressed by virtually all freshly isolated blood DCs (Fig. 2, a and b). PSGL-1 was also homogeneously expressed on ex vivo–derived DCs (cultured for 10–14 d), whereas HECA-452 was expressed on only a subset of these cells (25–40%; Fig. 2 c). To further characterize the structure of these molecules on DCs, immunoblotting of cell lysates with HECA-452 and anti–PSGL-1 antibodies was performed (Fig. 2, d and e). Both antibodies recognized bands of ∼240 and 140 kD under normal reducing conditions, representing the dimeric and monomeric forms of CLA/ PSGL-1 as described previously (10). Enhanced reduction resulted in conversion of all immunoreactivity to a single band of 140 kD, indicating that in DCs, as in T cells, the epitope recognized by HECA-452 is expressed on only a single major surface glycoprotein, PSGL-1.
DCs Tether and Roll on Both E- and P-selectin In Vitro.
Having identified a candidate ligand for E- and P-selectin, the ability of these cells to tether and roll on both E- and P-selectin was assessed in vitro using a parallel plate flow chamber (19). HL60 cells, a promyelocytic cell line known to bind to E- and P-selectin (20), were included as a control. DCs bound to both E-selectin and P-selectin with high affinity and rolled in shear flow (Fig. 3, a and b). The percentage of cells interacting with E- or P-selectin after 3 min of static incubation varied between 77 and 94%. Resistance to detachment from E- and P-selectin during stepwise increases in shear flow was extremely high (Fig. 3, c and d). Even at a wall shear stress as high as 100 dyn/cm2, >50% of the tethered cells remained bound and rolling.
Figure 3.

DCs tether and roll on E- and P-selectin in vitro. Interaction of DCs with E- and P-selectin was assessed in a parallel plate flow chamber. HL60 cells are included as a positive binding control. Both fresh and cultured DCs formed tethers on E-selectin (a) and P-selectin (b) at 1 dyn/ cm2 and rolled in a calcium-dependent fashion. DCs bound to either E- (c) or P-selectin (d) showed strong resistance to detachment by increasing wall shear stress with >80–100% of cells remaining bound at 25 dyn/cm2. E- and P-selectin binding was abolished by perfusion of medium containing 5 mM EDTA. The data presented are the mean and range of two experiments and are representative of several independent observations.
No significant differences were observed between fresh and cultured DCs within the physiologic shear range of postcapillary venular blood flow (1–25 dyn/cm2). However, at high shear stresses (50–100 dyn/cm2) fresh DCs showed an increased shear resistance compared with cultured DCs (Fig. 3, c and d).
DCs Tether and Roll on Endothelium of Noninflamed Skin.
While the parallel plate flow chamber assay is a useful model for assessment of distinct adhesion pathways under defined shear flow conditions, the advent of intravital microscopy has made possible the direct observation of leukocyte interaction with postcapillary venular endothelium in vivo (14). The ability of human E- and P-selectin ligands to interact effectively with murine selectins allows human leukocytes to be studied in mouse microvessels (Stein, J., and U.H. von Andrian, unpublished data). Selectin-dependent rolling of leukocytes has been observed in mouse skin microcirculation without surgical manipulation of the tissue, thus allowing observation of basal interactions between leukocytes and noninflamed endothelium (21; and Ulfman, L.H., and U.H. von Andrian, manuscript submitted for publication). We tested the hypothesis that the high level of CLA/PSGL-1 expression on blood DCs would permit tethering and rolling of these cells in uninflamed ear postcapillary venules. Fluorescently labeled human DCs were injected through the right common carotid artery into the aortic arch of anesthetized mice and observed directly in the microcirculation of the left ear. In the absence of any overt skin inflammation, a significant fraction of injected blood DCs were observed to tether and roll in cutaneous microvessels in vivo (Table I). Injected cells interacted exclusively with postcapillary venules, where the extravasation of other leukocytes is known to take place (22), and not with arterioles or capillaries (Fig. 4 a). A significant proportion of cultured DCs also rolled in murine ear postcapillary venules, and displayed comparable rolling properties (Table I).
Table I.
DC Rolling Parameters in Ear Venules
| Fresh DCs | Cultured DCs | |||||||
|---|---|---|---|---|---|---|---|---|
| No Antibody | Plus antibodies* | Wild-type mice | E/P-selectin−/− mice | |||||
| No. of animals/venules/cells | 3/31/534 | 2/14/241 | 5/27/627 | 3/31/96 | ||||
| Rolling fraction (%) | 26.5 ± 7.5 | 0.01 ± 0 | 50 ± 12.4 | 0.3 ± 0 | ||||
| Mean rolling velocity (μm/s) | 59.8 ± 35.4 | — | 57.8 ± 4 | — | ||||
| Mean blood flow velocity (μm/s) | 1,549 ± 633 | ND | 1,294 ± 264 | 1,434 ± 563 | ||||
| Wall shear rate (s−1) | 406 ± 228 | ND | 339 ± 78 | 253 ± 39 | ||||
| Shear stress (dyn/cm2) | 10.1 ± 5.7 | ND | 8.5 ± 2 | 6.3 ± 1 | ||||
Rolling fractions were obtained from the numbers of cells and venules indicated in the table. Hemodynamic parameters were determined in 2 animals, 3 venules, and 55 noninteracting cells for freshly isolated DCs; 4 animals, 4 venules, and 82 noninteracting cells for cultured DCs in wild-type mice; and in 2 animals, 2 venules, and 96 noninteracting cells for the E/P-selectin−/− mice and were calculated as described previously (reference 22). Centerline blood flow velocity (V CL) was calculated from the velocity of the fastest cell traversing the field during the period of observation. The mean blood flow velocity was estimated assuming a parabolic flow profile as V blood = V CL/1.6. The wall shear stress was approximated from the wall shear rate assuming the viscosity of blood to be 0.025 poise.
100 μg of anti–mouse E- and P-selectin blocking antibodies (9A9 and 5H1, respectively; provided by B. Wolitzky) was injected per mouse.
Figure 4.
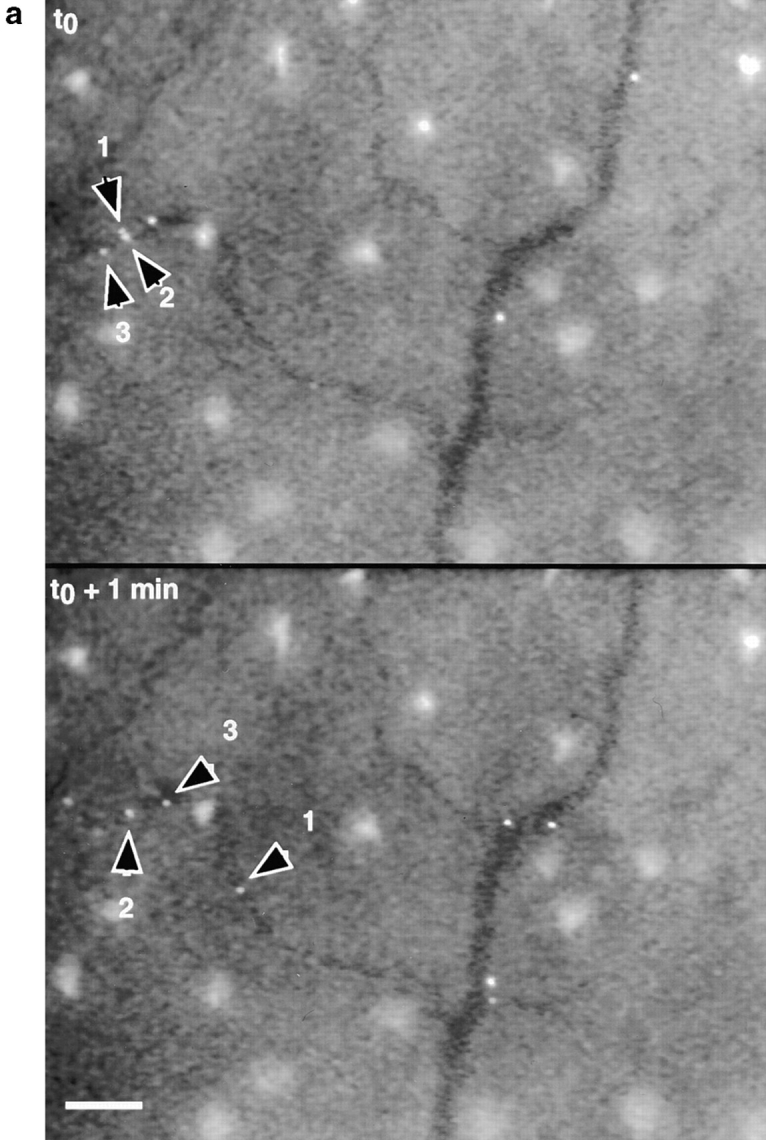

(a) DCs tether and roll in murine ear postcapillary venules in vivo. Calcein-labeled DCs were observed in ear postcapillary venules (top panel) after carotid injection. Continuous observation of bound cells (representative examples identified by arrowheads) reveals considerable heterogeneity of individual velocities. Three cells identified in the top panel are observed to move significantly different distances over the 1-min period of observation. All other cells shown have rolled out of the field of view during the 1-min interval. During that same period, several additional rolling cells have appeared (bottom panel). Unbound cells moved through the field of view within <1 s (not shown). The large bright areas seen between vessels are autofluorescent hair follicles. Bar, 100 μm. (b) Attachment and rolling of cultured DCs in wild-type mice (51 cells in a 28-μm-diameter venule; mean blood flow velocity = 890.6 μm/s, V crit = 388.2 μm/s) and E/P-selectin double-deficient mice (64 cells in a 34-μm-diameter venule; mean blood flow velocity = 1,281 μm/s, V crit = 471.7 μm/ s). V crit, the velocity of an idealized noninteracting cell (i.e., moving freely with the bloodstream) traveling at the vessel wall, was calculated as described (reference 22). DCs were not observed to form productive attachments in E/P-selectin−/− mice, where all cells (solid line) traveled at velocities greater than V crit, whereas ∼53% of cells observed in wild-type mice (dashed line) rolled at velocities less than V crit.
Interaction of DCs with Endothelium In Vivo Is Mediated by Endothelial Selectins.
To assess the role of endothelial selectins in DC binding to postcapillary venules, we performed experiments in the presence of blocking antibodies as well as in E/P-selectin−/− animals (13). As shown in Table I, pretreatment with anti–E- and P-selectin mAbs effectively blocked interaction of DCs with skin vessels. Objective assessment of leukocyte endothelial interaction was obtained by comparing the velocity distribution of cells observed in comparable size vessels in E/P-selectin−/− and control mice (Fig. 4 b). Cells traveling below V crit, the calculated velocity of noninteracting cells at the endothelial surface of a blood vessel with a parabolic flow profile (22), are defined as interacting with (i.e., rolling on) the vessel walls while the cells moving above V crit are traveling free in the bloodstream. Although ∼50% of labeled DCs observed in the vessels of wild-type mice formed rolling interactions, no significant interactions were observed in endothelial selectin–deficient animals (<0.5% labeled cells traveling below V crit). Besides E- and P-selectin ligands, the only known leukocyte adhesion molecules capable of mediating tethering and rolling on vascular endothelium are L-selectin, very late antigen 4 (VLA-4) and α4β7. The vascular ligands for L-selectin and integrin α4β7, peripheral lymph node addressin (PNAd) and mucosal addressin cell adhesion molecule 1 (MAdCAM-1), respectively, are not expressed in normal skin endothelium (23, 24) and therefore would not be expected to participate in these interactions. However, murine vascular cell adhesion molecule 1 (VCAM-1) may be present, and can readily bind human VLA-4 (integrin α4β1 [25]). The lack of interactions observed in anti-selectin antibody–blocked or selectin-deficient animals unambiguously indicates that the spontaneous tethering and rolling of DCs observed in wild-type animals is mediated by one or both of the endothelial selectins constitutively expressed on skin postcapillary venules, and that a major role for other known ligand pairs can be excluded, at least in the dermal vascular bed studied here. We cannot rule out unknown or uncharacterized ligand pairs that do not bind across species. However, these data are consistent with observations in rats, mice, and rabbits that the constitutive rolling of endogenous leukocytes in skin venules is mediated primarily via interaction with vascular selectins (26, 27; and Ulfman, L.H., and U.H. von Andrian, manuscript submitted for publication), and not via α4 integrins, or L-selectin, in contrast to other anatomic sites such as bone marrow (28) or peripheral lymph nodes (29).
DCs Are Recruited into Inflamed Skin.
Endothelial selectins and other adhesion molecules involved in leukocyte recruitment are upregulated during cutaneous inflammation (30). Since blood DCs express functional selectin ligands, chemokine receptors, and cell surface integrins (2, 3, 7), they should have all the requisite molecules necessary for recruitment into inflamed tissues. To test whether DCs could be recruited to inflamed skin, we measured accumulation of radioactively labeled DCs in ear skin of oxazolone-sensitized mice challenged with antigen on one ear 48 h before the infusion of the cells. To avoid potential incompatibility between human and mouse elements in the downstream portions of the adhesion/extravasation cascade, we used immature murine bone marrow–derived DCs, grown for 4–6 d with GM-CSF and IL-4 (9). 51Cr-labeled DCs (mean specific activity = 288 cpm/103) were injected intravenously, and the mice were killed 6 h later. Radioactivity counts were significantly higher in inflamed ears compared with control ears (cpm ratio in challenged versus control ears ranged from 1.7 to 9; mean ratio = 3.5, P < 0.01; Fig. 5, a and b). Cells recovered in inflamed ears ranged from 0.02 to 0.25% of injected cells or 103 to 1.25 × 104 cells/ear. This is consistent with previous reports of cells recovered from significantly larger areas of inflamed skin samples after intravenous injection of radiolabeled T cells (31, 32). A trivial explanation would be that this increase in cpm resulted from an increase of the volume of blood in the inflamed ear, and not from extravasation of DCs. One argument against this explanation is that this phenomenon required living cells, since no specific label was found in the skin after injection of dead DCs (not shown). To confirm that this increase in radioactivity actually represented extravasated DCs, we performed similar experiments with calcein-labeled DCs and injected PE-conjugated anti–mouse CD31 mAb 5 min before the animals were killed, to stain the vessels in red. Confocal microscopy of inflamed ears showed extravascular DCs (green) clearly outside of the skin vessels and in the extravascular tissue (Fig. 5, c–e), confirming the actual extravasation of the DCs, whereas no extravascular cells were observed in noninflamed ears. It should be emphasized that this experiment was designed to be purely qualitative and to address the anatomical location rather than absolute number of extravasated cells.
Figure 5.


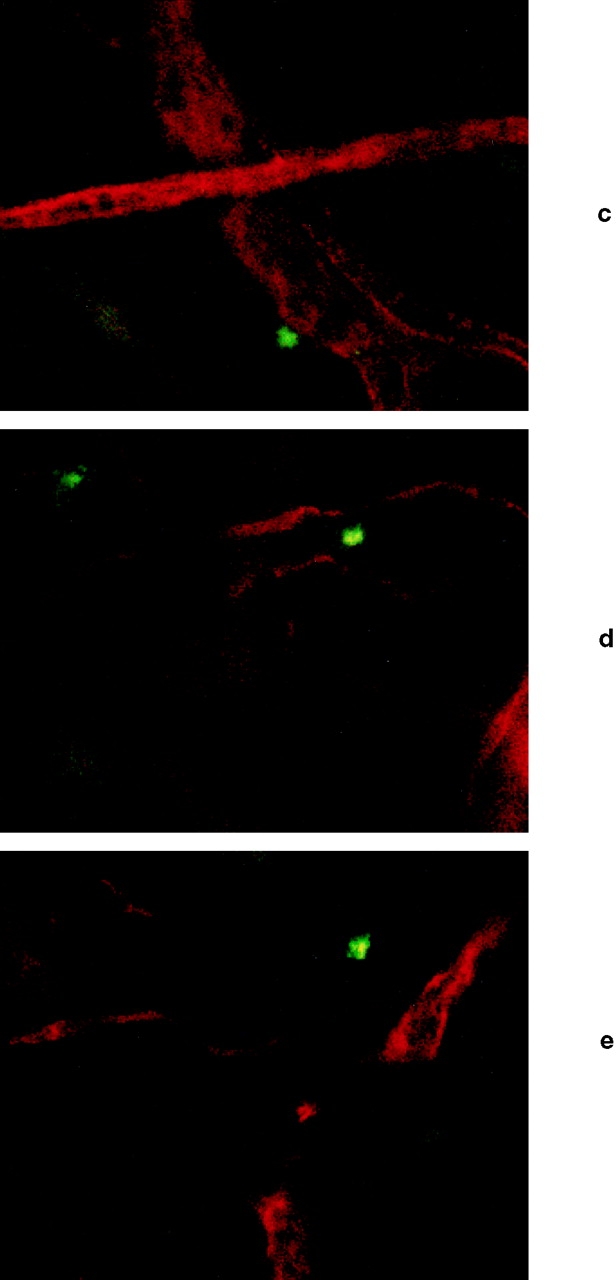
Immature murine bone marrow–derived DCs are acutely recruited into inflamed skin. Mice sensitized to oxazolone were challenged on the right ear 48 h before intravenous infusion of 5 × 106 51Cr-labeled DCs. 6 h after DC infusion, the mice were killed and the cpm were compared in both ears. (a) Results obtained in nine mice are shown; cpm values are corrected by subtraction of the background radioactivity. There is a significant difference between the challenged and control ear (P < 0.01, Student's t test). (b) Shows the fold increase in cpm measured in inflamed ears versus contralateral control ears for nine independent determinations. The area from 0 to 1 is shaded. Values above the shaded area reflect an increased number of labeled cells in the inflamed ear, and values within the shaded area would reflect a decreased number of cells in the inflamed ear relative to control. The solid line and hatched field represent the mean and 95% confidence interval for fold increase of inflamed ear counts versus control (P < 0.01, Student's t test). (c–e) Confocal microscopy images of an inflamed ear 6 h after infusion of 20 × 106 calcein-labeled murine bone marrow–derived DCs. 100 μg of PE-conjugated mAb anti–mouse CD31 (PharMingen) was infused 5 min before the animal was killed, in order to visualize the vessels. Several fields of the same ear (photographed through a 40× objective) are shown: (c) a calcein-labeled DC is seen just outside of the vessel wall; (d) another DC is seen between two small vessels; and (e) a calcein- labeled DC has clearly extravasated and is shown deeper in the surrounding tissues. No extravascular cells were observed in contralateral, noninflamed ear.
Discussion
In this study, we have shown that DCs efficiently tether and roll on E- and P-selectin in vitro. Taking advantage of previous studies showing that relevant adhesion molecules which mediate leukocyte rolling (i.e., selectins/selectin ligands and VLA-4/VCAM-1) are functional across human and murine species (25; and Stein, J., and U.H. von Andrian, unpublished data), we have demonstrated a direct interaction between human DCs and murine skin postcapillary venules in vivo. This confirms the hypothesis that DCs can participate in the essential first step of the adhesion/ extravasation pathway on vessels expressing E- and/or P-selectin. This interaction takes place in the absence of overt inflammation and may also facilitate the extravasation of DCs into injured skin. To undergo extravasation, rolling cells have to be activated (e.g., by chemoattractants) and to bind via surface integrins to vascular cell adhesion molecules (e.g., intercellular adhesion molecule 1 [ICAM-1] and VCAM-1). DCs express multiple chemokine receptors (7, 33, 34). As described previously for different types of lymphocytes (35), it was recently shown that DCs at different maturational stages express patterns of chemokine receptors that seem to correlate with recruitment to distinct anatomic sites (36–38). DCs also express the cell surface integrins LFA-1 and VLA-4 (2, 3), the ligands for ICAM-1 and VCAM-1, respectively. Thus, DCs appear to have all the surface molecules required to participate in the adhesion/extravasation cascade. We have confirmed this potential by demonstrating active recruitment of immature cultured DCs into inflamed skin after intravenous injection and by direct visualization of extravasated DCs using confocal microscopy. These experiments were highly reproducible despite the low absolute number of cells found in ear skin. The association between skin homing potential and in vivo rolling on murine skin venules is strengthened by the observation that naive lymphocytes, which do not express PSGL-1/CLA and do not normally migrate to skin (15), do not show significant interaction with murine skin endothelium in vivo. In contrast, T cells isolated from a cutaneous T cell lymphoma patient, which express PSGL-1/ CLA at high levels and do target to skin, roll spontaneously on uninflamed mouse ear postcapillary venules (Robert, C., unpublished data).
There are at least two non-mutually exclusive reasons why blood DCs should be able to migrate to peripheral tissues. First, they may represent a pool of tissue DC precursors that continuously extravasate to repopulate tissues with resident DCs (e.g., for skin, the dermal DCs and/or the Langerhans cells). Second, blood DCs may also represent a circulating pool of APCs that are acutely recruited to sites of inflammation. Consistent with this hypothesis, it has been reported that the number of DCs is increased in tissues in different models of inflammation (39–41), though DCs have never been shown to be actively recruited from the blood.
We showed that blood DCs constitutively interact with normal murine skin endothelium in vivo via selectins. This spontaneous rolling of CLA+ DCs in skin microvessels, in the absence of an inflammatory stimulus, suggests that a large number of blood DCs may be interacting with cutaneous postcapillary venules at any given time. This continuous interaction is likely to play a role in the seeding of peripheral tissues with DCs to provide the tissue-resident pool of APCs. We also showed that DCs can be acutely recruited into inflamed skin. We propose that constitutive selectin-mediated rolling represents surveillance of the luminal aspect of skin endothelium by blood DCs for activating signals (e.g., chemokines). This enhances the ability of these potent APCs to rapidly extravasate when they encounter inflamed endothelium, as in the setting of skin injury or infection. Rapid recruitment of blood DCs, facilitated by these constitutive reversible interactions between blood DCs and endothelial surfaces, could enhance local antigen capture and antigen presentation activity both at the site of inflammation and in the draining lymph nodes. Teleologically, this may represent a previously unrecognized element of skin immunosurveillance and a highly adaptive interface between innate and acquired immunity.
It is interesting to note that DCs are often referred to as “nature's adjuvant” (1). The rapid recruitment of APCs to sites of inflammation may be an important element in determining the efficiency of primary immune responses, as seen, for example, in the enhancing effect of adjuvant- induced inflammation on the response to immunization. The relative importance of rapid recruitment of DCs and monocytes in this process is at present unknown. We favor the hypothesis that DCs provide an immediate bolus of APCs that can present antigens to CLA+ memory T cells that have extravasated under similar conditions in inflamed skin. Monocytes also express PSGL-1/CLA and can tether and roll on E- and P-selectin in vitro (Kieffer, J.D., unpublished data). They can differentiate into DCs in vitro (42) but require at least 48 h to do so, and therefore may be more important in the amplification and perpetuation of the immune response. These hypotheses are currently being tested experimentally.
In summary, we show that immature blood DCs can participate in the leukocyte adhesion/extravasation pathway and be acutely recruited to inflamed skin. Constitutive interactions between DCs and normal skin endothelium may represent an important immunosurveillance mechanism by which DCs could sample local activating signals and rapidly extravasate into inflamed tissues.
Acknowledgments
The authors thank B. Wolitzky for generously providing the anti E- and P-selectin antibodies.
This work was supported by the Dermatology Foundation, the Association pour la Recherche contre le Cancer, the National Institutes of Health, the LED (Laboratoires d'Evolution Dermatologique), the René Touraine Foundation, the Philippe Foundation, and the Howard Hughes Medical Institute.
Abbreviations used in this paper
- CLA
cutaneous lymphocyte-associated antigen
- DC
dendritic cell
- PSGL-1
P-selectin glycoprotein ligand 1
- VCAM-1
vascular cell adhesion molecule 1
- VLA-4
very late antigen 4
References
- 1.Steinman RM. The dendritic cell system and its role in immunogenicity. Annu Rev Immunol. 1991;9:271–296. doi: 10.1146/annurev.iy.09.040191.001415. [DOI] [PubMed] [Google Scholar]
- 2.Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. doi: 10.1038/32588. [DOI] [PubMed] [Google Scholar]
- 3.Hart DN. Dendritic cells: unique leukocyte populations which control the primary immune response. Blood. 1997;90:3245–3287. [PubMed] [Google Scholar]
- 4.O'Doherty U, Peng M, Gezelter S, Swiggard WJ, Betjes M, Bhardwaj N, Steinman RM. Human blood contains two subsets of dendritic cells, one immunologically mature and the other immature. Immunology. 1994;82:487–493. [PMC free article] [PubMed] [Google Scholar]
- 5.Springer TA. Traffic signals on endothelium for lymphocyte recirculation and leukocyte emigration. Annu Rev Physiol. 1995;57:827–872. doi: 10.1146/annurev.ph.57.030195.004143. [DOI] [PubMed] [Google Scholar]
- 6.Butcher EC, Picker LJ. Lymphocyte homing and homeostasis. Science. 1996;272:60–66. doi: 10.1126/science.272.5258.60. [DOI] [PubMed] [Google Scholar]
- 7.Ayehunie S, Garcia-Zepeda EA, Hoxie JA, Horuk R, Kupper TS, Luster AD, Ruprecht RM. Human immunodeficiency virus-1 entry into purified blood dendritic cells through CC and CXC chemokine coreceptors. Blood. 1997;90:1379–1386. [PubMed] [Google Scholar]
- 8.Caux C, Dezutter-Dambuyant C, Schmitt D, Banchereau J. GM-CSF and TNF-alpha cooperate in the generation of dendritic Langerhans cells. Nature. 1992;360:258–261. doi: 10.1038/360258a0. [DOI] [PubMed] [Google Scholar]
- 9.Mayordomo JI, Zorina T, Storkus WJ, Zitvogel L, Celluzzi C, Falo LD, Melief CJ, Ildstad ST, Kast WM, Deleo AB, et al. Bone marrow-derived dendritic cells pulsed with synthetic tumour peptides elicit protective and therapeutic antitumour immunity. Nat Med. 1995;1:1297–1302. doi: 10.1038/nm1295-1297. [DOI] [PubMed] [Google Scholar]
- 10.Fuhlbrigge RC, Kieffer JD, Armerding D, Kupper TS. Cutaneous lymphocyte antigen is a specialized form of PSGL-1 expressed on skin-homing T cells. Nature. 1997;389:978–981. doi: 10.1038/40166. [DOI] [PubMed] [Google Scholar]
- 11.Picker LJ, Terstappen LW, Rott LS, Streeter PR, Stein H, Butcher EC. Differential expression of homing-associated adhesion molecules by T cell subsets in man. J Immunol. 1990;145:3247–3255. [PubMed] [Google Scholar]
- 12.Fuhlbrigge RC, Alon R, Puri KD, Lowe JB, Springer TA. Sialylated, fucosylated ligands for L-selectin expressed on leukocytes mediate tethering and rolling adhesions in physiologic flow conditions. J Cell Biol. 1996;135:837–848. doi: 10.1083/jcb.135.3.837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Frenette PS, Mayadas TN, Rayburn H, Hynes RO, Wagner DD. Susceptibility to infection and altered hematopoiesis in mice deficient in both P- and E-selectins. Cell. 1996;84:563–574. doi: 10.1016/s0092-8674(00)81032-6. [DOI] [PubMed] [Google Scholar]
- 14.Atherton A, Born GV. Quantitative investigations of the adhesiveness of circulating polymorphonuclear leucocytes to blood vessel walls. J Physiol (Lond) 1972;222:447–474. doi: 10.1113/jphysiol.1972.sp009808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Picker LJ, Treer JR, Ferguson-Darnell B, Collins PA, Bergstresser PR, Terstappen LW. Control of lymphocyte recirculation in man. II. Differential regulation of the cutaneous lymphocyte-associated antigen, a tissue- selective homing receptor for skin-homing T cells. J Immunol. 1993;150:1122–1136. [PubMed] [Google Scholar]
- 16.Strunk D, Egger C, Leitner G, Hanau D, Stingl G. A skin homing molecule defines the Langerhans cell progenitor in human peripheral blood. J Exp Med. 1997;185:1131–1136. doi: 10.1084/jem.185.6.1131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ebner S, Lenz A, Reider D, Fritsch P, Schuler G, Romani N. Expression of maturation-/migration-related molecules on human dendritic cells from blood and skin. Immunobiology. 1998;198:568–587. doi: 10.1016/S0171-2985(98)80079-X. [DOI] [PubMed] [Google Scholar]
- 18.Berg EL, Yoshino T, Rott LS, Robinson MK, Warnock RA, Kishimoto TK, Picker LJ, Butcher EC. The cutaneous lymphocyte antigen is a skin lymphocyte homing receptor for the vascular lectin endothelial cell– leukocyte adhesion molecule 1. J Exp Med. 1991;174:1461–1466. doi: 10.1084/jem.174.6.1461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lawrence MB, Springer TA. Leukocytes roll on a selectin at physiologic flow rates: distinction from and prerequisite for adhesion through integrins. Cell. 1991;65:859–873. doi: 10.1016/0092-8674(91)90393-d. [DOI] [PubMed] [Google Scholar]
- 20.Asa D, Raycroft L, Ma L, Aeed PA, Kaytes PS, Elhammer AP, Geng JG. The P-selectin glycoprotein ligand functions as a common human leukocyte ligand for P- and E-selectins. J Biol Chem. 1995;270:11662–11670. doi: 10.1074/jbc.270.19.11662. [DOI] [PubMed] [Google Scholar]
- 21.Maly P, Thall A, Petryniak B, Rogers CE, Smith PL, Marks RM, Kelly RJ, Gersten KM, Cheng G, Saunders TL, et al. The alpha(1,3)fucosyltransferase Fuc-TVII controls leukocyte trafficking through an essential role in L-, E-, and P-selectin ligand biosynthesis. Cell. 1996;86:643–653. doi: 10.1016/s0092-8674(00)80137-3. [DOI] [PubMed] [Google Scholar]
- 22.Ley K, Gaehtgens P. Endothelial, not hemodynamic, differences are responsible for preferential leukocyte rolling in rat mesenteric venules. Circ Res. 1991;69:1034–1041. doi: 10.1161/01.res.69.4.1034. [DOI] [PubMed] [Google Scholar]
- 23.Berg EL, Mullowney AT, Andrew DP, Goldberg JE, Butcher EC. Complexity and differential expression of carbohydrate epitopes associated with L-selectin recognition of high endothelial venules. Am J Pathol. 1998;152:469–477. [PMC free article] [PubMed] [Google Scholar]
- 24.Briskin M, Winsor-Hines D, Shyjan A, Cochran N, Bloom S, Wilson J, McEvoy LM, Butcher EC, Kassam N, Mackay CR, et al. Human mucosal addressin cell adhesion molecule-1 is preferentially expressed in intestinal tract and associated lymphoid tissue. Am J Pathol. 1997;151:97–110. [PMC free article] [PubMed] [Google Scholar]
- 25.Miyake K, Hasunuma Y, Yagita H, Kimoto M. Requirement for VLA-4 and VLA-5 integrins in lymphoma cells binding to and migration beneath stromal cells in culture. J Cell Biol. 1992;119:653–662. doi: 10.1083/jcb.119.3.653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Fiebig E, Ley K, Arfors KE. Rapid leukocyte accumulation by “spontaneous” rolling and adhesion in the exteriorized rabbit mesentery. Int J Microcirc Clin Exp. 1991;10:127–144. [PubMed] [Google Scholar]
- 27.Nolte D, Schmid P, Jager U, Botzlar A, Roesken F, Hecht R, Uhl E, Messmer K, Vestweber D. Leukocyte rolling in venules of striated muscle and skin is mediated by P-selectin, not by L-selectin. Am J Physiol. 1994;267:H1637–H1642. doi: 10.1152/ajpheart.1994.267.4.H1637. [DOI] [PubMed] [Google Scholar]
- 28.Mazo IB, Gutierrez-Ramos JC, Frenette PS, Hynes RO, Wagner DD, von Andrian UH. Hematopoietic progenitor cell rolling in bone marrow microvessels: parallel contributions by endothelial selectins and vascular cell adhesion molecule 1. J Exp Med. 1998;188:465–474. doi: 10.1084/jem.188.3.465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Warnock RA, Askari S, Butcher EC, von Andrian UH. Molecular mechanisms of lymphocyte homing to peripheral lymph nodes. J Exp Med. 1998;187:205–216. doi: 10.1084/jem.187.2.205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Groves RW, Ross E, Barker JN, Ross JS, Camp RD, MacDonald DM. Effect of in vivo interleukin-1 on adhesion molecule expression in normal human skin. J Investig Dermatol. 1992;98:384–387. doi: 10.1111/1523-1747.ep12499816. [DOI] [PubMed] [Google Scholar]
- 31.Austrup F, Vestweber D, Borges E, Lohning M, Brauer R, Herz U, Renz H, Hallmann R, Scheffold A, Radbruch A, Hamann A. P- and E-selectin mediate recruitment of T-helper-1 but not T-helper-2 cells into inflamed tissues. Nature. 1997;385:81–83. doi: 10.1038/385081a0. [DOI] [PubMed] [Google Scholar]
- 32.Tietz W, Hamann A. The migratory behavior of murine CD4+ cells of memory phenotype. Eur J Immunol. 1997;27:2225–2232. doi: 10.1002/eji.1830270916. [DOI] [PubMed] [Google Scholar]
- 33.Sozzani S, Sallusto F, Luini W, Zhou D, Piemonti L, Allavena P, Van Damme J, Valitutti S, Lanzavecchia A, Mantovani A. Migration of dendritic cells in response to formyl peptides, C5a, and a distinct set of chemokines. J Immunol. 1995;155:3292–3295. [PubMed] [Google Scholar]
- 34.Luster AD. Chemokines—chemotactic cytokines that mediate inflammation. N Engl J Med. 1998;338:436–445. doi: 10.1056/NEJM199802123380706. [DOI] [PubMed] [Google Scholar]
- 35.Sallusto F, Lenig D, Mackay CR, Lanzavecchia A. Flexible programs of chemokine receptor expression on human polarized T helper 1 and 2 lymphocytes. J Exp Med. 1998;187:875–883. doi: 10.1084/jem.187.6.875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Dieu MC, Vanbervliet B, Vicari A, Bridon JM, Oldham E, Ait-Yahia S, Briere F, Zlotnik A, Lebecque S, Caux C. Selective recruitment of immature and mature dendritic cells by distinct chemokines expressed in different anatomic sites. J Exp Med. 1998;188:373–386. doi: 10.1084/jem.188.2.373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Sozzani S, Allavena P, D'Amico G, Luini W, Bianchi G, Kataura M, Imai T, Yoshie O, Bonecchi R, Mantovani A. Differential regulation of chemokine receptors during dendritic cell maturation: a model for their trafficking properties. J Immunol. 1998;161:1083–1086. [PubMed] [Google Scholar]
- 38.Sallusto F, Schaerli P, Loetscher P, Schaniel C, Lenig D, Mackay CR, Qin S, Lanzavecchia A. Rapid and coordinated switch in chemokine receptor expression during dendritic cell maturation. Eur J Immunol. 1998;28:2760–2769. doi: 10.1002/(SICI)1521-4141(199809)28:09<2760::AID-IMMU2760>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- 39.Christensen OB, Wall LM. Long term effect on epidermal dendritic cells of four different types of exogenous inflammation. Acta Derm Venereol. 1987;67:305–309. [PubMed] [Google Scholar]
- 40.McWilliam AS, Nelson D, Thomas JA, Holt PG. Rapid dendritic cell recruitment is a hallmark of the acute inflammatory response at mucosal surfaces. J Exp Med. 1994;179:1331–1336. doi: 10.1084/jem.179.4.1331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.McWilliam AS, Napoli S, Marsh AM, Pemper FL, Nelson DJ, Pimm CL, Stumbles PA, Wells TN, Holt PG. Dendritic cells are recruited into the airway epithelium during the inflammatory response to a broad spectrum of stimuli. J Exp Med. 1996;184:2429–2432. doi: 10.1084/jem.184.6.2429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Pickl WF, Majdic O, Kohl P, Stockl J, Riedl E, Scheinecker C, Bello-Fernandez C, Knapp W. Molecular and functional characteristics of dendritic cells generated from highly purified CD14+ peripheral blood monocytes. J Immunol. 1996;157:3850–3859. [PubMed] [Google Scholar]