

Abstract
To determine the relative contributions of respiratory burst–derived reactive oxygen intermediates (ROI) versus reactive nitrogen intermediates (RNI) to macrophage-mediated intracellular host defense, mice genetically deficient in these mechanisms were challenged with Leishmania donovani, a protozoan that selectively parasitizes visceral tissue macrophages. During the early stage of liver infection at wk 2, both respiratory burst–deficient gp91phox −/− (X-linked chronic granulomatous disease [X-CGD]) mice and inducible nitric oxide synthase (iNOS) knockout (KO) mice displayed comparably increased susceptibility. Thereafter, infection was unrestrained in mice lacking iNOS but was fully controlled in X-CGD mice. Mononuclear cell influx into infected liver foci in X-CGD and iNOS KO mice was also overtly impaired at wk 2. However, granuloma assembly in parasitized tissue eventually developed in both hosts but with divergent effects: mature granulomas were functionally active (leishmanicidal) in X-CGD mice but inert in iNOS-deficient animals. These results suggest that (a) ROI and RNI probably act together in the early stage of intracellular infection to regulate both tissue recruitment of mononuclear inflammatory cells and the initial extent of microbial replication, (b) RNI alone are necessary and sufficient for eventual control of visceral infection, and (c) although mature granulomas have traditionally been associated with control of such infections, these structures fail to limit intracellular parasite replication in the absence of iNOS.
Keywords: visceral leishmaniasis, protozoan, parasite, granuloma, macrophage
L eishmania donovani, a protozoan that causes visceral leishmaniasis, selectively parasitizes and initially replicates within resident macrophages in the liver, spleen and bone marrow. The host capable of controlling this disseminated intracellular infection develops an effective T cell– dependent immune response mediated by proinflammatory, largely Th1 cell–associated cytokines including IL-12 and IFN-γ (1–3; for review see reference 4). The net effect of this complex response, which involves granuloma formation (4, 5), is sufficient activation of both tissue macrophages and influxing blood monocytes to kill most intracellular L. donovani and induce quiescence in residual infection (4).
In vitro studies suggest that mononuclear phagocytes possess multiple leishmanicidal mechanisms. Cytokine-activated mouse peritoneal macrophages kill ingested L. donovani amastigotes by secreting either reactive oxygen intermediates (ROI; primarily H2O2 [6, 7]) or reactive nitrogen intermediates (RNI; derived from nitric oxide [8]). Inhibit ing either pathway prevents mouse macrophage killing of L. donovani in vitro (6–8); thus, these mechanisms may also act in concert (9, 10). In cytokine-activated human monocyte-derived macrophages, H2O2 also readily induces leishmanicidal effects (11); however, a similar role for RNI has been difficult to detect in cells from normal donors (12). In vitro–activated human macrophages can also limit L. donovani replication by degrading extracellular tryptophan (13).
Studies carried out to define the macrophage mechanisms of intracellular Leishmania killing in vivo have been more limited. Thorough observations in a separate mouse model of L. major cutaneous infection largely (14–17) but not entirely (18, 19) support a critical role for inducible nitric oxide synthase (iNOS)-generated RNI. RNI are not only leishmanicidal (8, 14–17) but also regulate immunologic pathways, including endogenous secretion of IL-12 and IFN-γ (15, 17). In L. donovani–infected BALB/c mice, iNOS mRNA is induced in parasitized tissues (2, 20) and presumed inhibition of iNOS by aminoguanidine (AG) treatment impairs host defense (2). However, iNOS mRNA induction (20) and generation of iNOS-derived products in serum and infected organs (21) do not necessarily correlate with control over L. donovani (20, 21). In the one published study of parasitized tissue in human visceral leishmaniasis, splenic mononuclear phagocytes from each of 22 untreated Indian patients showed iNOS immunoreactivity (22) (36 ± 3% of cells were iNOS positive; Murray, H., A. Ding, S. Sundar, and C. Nathan, unpublished data).
To directly compare RNI and ROI as macrophage- derived antimicrobial mediators in vivo, we used L. donovani to challenge gene-disrupted iNOS knockout (KO) (23) and respiratory burst–deficient gp91phox −/− (X-linked chronic granulomatous disease [X-CGD]) mice (24).
Materials and Methods
Animals.
Mice with a targeted disruption of the gp91phox subunit of the NADPH-oxidase complex (phox), derived from a C57BL/6 × 129/Sv background and backcrossed six times with C57BL/6 mice (24), were provided as breeders by Dr. M. Dinauer (Indiana University Medical Center, Indianapolis, IN). C57BL/6 mice (Charles Rivers Labs.) were used as controls. iNOS−/− KO mice (C57BL/6 × 129/Sv) were maintained as originally described (23); wild-type+/+ littermates served as controls. Mice were 8–15 wk old when infected; both males and females were used randomly except for control C57BL/6 mice (all female).
Visceral Infection and Treatment.
Groups of four to five mice were injected via the tail vein with 1.5 × 107 hamster spleen– derived L. donovani amastigotes (1 Sudan strain, provided by Dr. D. Sacks, National Institutes of Health, Bethesda, MD) (2, 3). Visceral infection was followed microscopically using Giemsa-stained liver imprints by counting the number of amastigotes per 500 cell nuclei × liver weight (g) (Leishman-Donovan units, or LDU) (2, 3). Granuloma formation was scored using formalin-fixed tissue sections stained with hematoxylin and eosin (1–3). In some experiments, starting 1 d after infection AG (Sigma Chemical Co.) was added at 1% (wt/vol) to acidified drinking water changed twice weekly; controls received acidified water alone (2). Differences between mean values were analyzed by a two-tailed Student's t test.
Results and Discussion
Initial Kinetics of Parasite Replication.
L. donovani replicated in the livers of both strains of control mice during the first 2 wk after challenge (Fig. 1); thereafter, parasite burdens declined consistent with killing and a self-healing phenotype (1). At wk 2, liver burdens in both X-CGD and iNOS KO mice were significantly higher (by 1.6- and 2.3-fold, respectively) than in control animals. Since macrophages from X-CGD mice produce RNI normally (24) and macrophages from iNOS KO mice show intact respiratory burst activity (23), the results at wk 2 suggested that (a) neither mechanism by itself was sufficient to help limit early L. donovani replication and (b) phox and iNOS may act in concert to achieve optimal initial activity.
Figure 1.

Course of L. donovani liver infection in (A) X-CGD (○) and control C57BL/6 mice (•), and in (B) iNOS KO mice (○) and wild-type controls (•). Results are mean ± SEM values for 8–15 mice at each time point from two to four experiments. Asterisk, significantly higher (P < 0.05) than value in control mice.
Acquired Resistance and Resolution of Infection.
After wk 2, C57BL/6 and wild-type control mice acquired resistance and showed near-resolution of infection by wk 8 (Fig. 1). Although initially more susceptible, X-CGD mice also controlled infection after wk 2 (Fig. 1 A), indicating that the phox-derived ROI mechanism was dispensable. In contrast, liver infection was unrestrained in iNOS KO mice and increased to high levels by wk 8 (Fig. 1 B and see Fig. 3). Thus, although an intact respiratory burst was neither required (Fig. 1 A) nor sufficient (Fig. 1 B) for acquired resistance and near-resolution of visceral infection, these responses were iNOS dependent.
Figure 3.
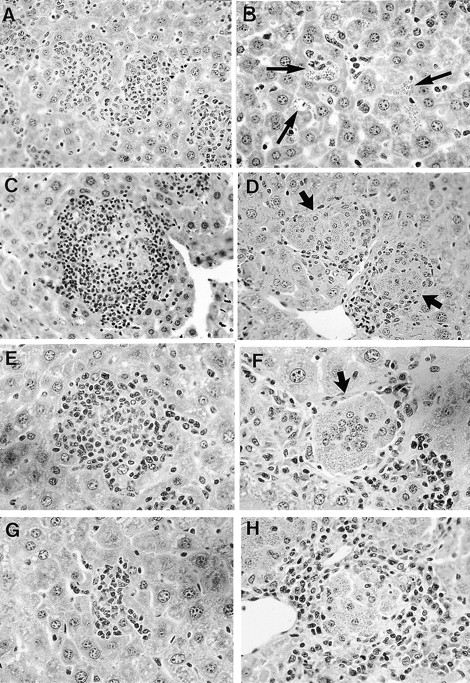
Histologic response in livers of infected iNOS KO mice and wild-type controls. At wk 2, granulomas are widespread and developed in controls (A) but absent at infected foci (arrows) in iNOS KO mice (B). At wk 4, granulomas are well-established or empty in control mice (C and E); in iNOS KO mice (D and F), granulomas are developing but heavily parasitized. At wk 8, inflammatory reaction in controls has involuted (G); in iNOS KO mice (H), granulomas are well-formed (mature) but contain numerous replicating amastigotes (see also Fig. 4). Original magnification: A, C, and D, ×200; B and E–H, ×315.
Antileishmanial Effect of iNOS in the Absence of Phox.
To determine if the capacity of X-CGD mice to control L. donovani infection involved iNOS-derived products, AG was used as an iNOS inhibitor (2, 25). Continuous AG treatment enhanced initial susceptibility of X-CGD mice at wk 2 (Table I), pointing to an active iNOS-dependent mechanism. At wk 4, liver parasite burdens in AG-treated X-CGD mice remained elevated; nevertheless, visceral infection began to come under control (Table I). Although this latter observation raised the possibility of an antimicrobial pathway independent of both phox and iNOS and although such pathways exist (9), we did not verify the degree of iNOS inhibition induced by AG. Since iNOS KO mice failed to control L. donovani, incomplete iNOS inhibition may well explain the decrease in parasite load at wk 4 in treated X-CGD animals.
Table I.
Effect of Aminoguanidine Treatment in X-CGD Mice
| Wk after infection | AG | Parasite burden | Percentage of infected foci showing granuloma formation | |||
|---|---|---|---|---|---|---|
| LDU | ||||||
| 2 | − | 3,065 ± 173 | 37 ± 5 | |||
| + | 5,214 ± 251* | 9 ± 2* | ||||
| 4 | − | 810 ± 146 | 88 ± 10 | |||
| + | 2,550 ± 128* | 93 ± 8 |
Starting 1 d after L. donovani infection, X-CGD mice received acidified drinking water alone or the same water containing AG at 1% (wt/vol) for 4 wk. Results indicate mean ± SEM values from two experiments, and for liver parasite burdens represent eight mice per group and for granuloma formation (percentage of liver-infected foci showing developing or mature granulomas; see Table II) represent six mice per group.
Significantly different (P < 0.05) from control mice.
Tissue Granulomatous Response.
In the liver, acquired resistance to L. donovani is expressed by a granulomatous response that encircles all initially parasitized resident macrophages (Kupffer cells) (1–4). This reaction is underway by wk 2 and fully evident at wk 4 as resistance develops. The surrounding mantle of the mature leishmanicidal granuloma is formed by influxing T cells and blood monocytes (4), a process governed by Th1 cell–derived cytokines (2– 4). During resolution of visceral infection (wk 8), the tissue reaction involutes and most cellular foci are devoid of visible amastigotes and can be scored as in Table II as microscopically “empty” (5).
Table II.
Liver Histologic (Granulomatous) Response to L. donovani Infection in X-CGD and iNOS KO Mice
| Weeks after infection | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| X-CGD | C57BL/6 | iNOS KO | WT Controls | |||||||||||||||||||||
| Tissue reaction | 2 | 4 | 8 | 2 | 4 | 8 | 2 | 4 | 8 | 2 | 4 | 8 | ||||||||||||
| None (%) | 70 | 10 | 0 | 19 | 0 | 0 | 81 | 32 | 1 | 22 | 0 | 0 | ||||||||||||
| Granuloma | ||||||||||||||||||||||||
| Developing (%) | 29 | 81 | 39 | 78 | 63 | 2 | 19 | 65 | 38 | 57 | 69 | 6 | ||||||||||||
| Mature (%) | 1 | 9 | 33 | 3 | 37 | 32 | 0 | 3 | 61 | 21 | 31 | 43 | ||||||||||||
| Empty (%) | 0 | 0 | 28 | 0 | 0 | 66 | 0 | 0 | 0 | 0 | 0 | 51 | ||||||||||||
All infected foci in 50 consecutive ×63 microscopic fields were scored as follows for granuloma formation: none, single or fused parasitized Kupffer cells with no mononuclear cell infiltrate; developing, core of fused infected Kupffer cells with some cell infiltrate; mature, fused Kupffer cell core surrounded by a well-developed mononuclear cell mantle; and empty, granuloma devoid of amastigotes (1–4). See Figs. 2 and 3. Results are from two experiments, and indicate mean values for four to five mice at each time point. SEM, omitted for clarity, were all <12%.
The kinetics of the granulomatous response and mature granuloma assembly were similar in control C57BL/6 (Fig. 2, A, C, and E) and wild-type C57BL/6 × 129/Sv (Fig. 3, A, C, and E, G) mice (Table II). In contrast, the tissue reaction in both X-CGD and iNOS KO animals was delayed and incomplete, particularly at the early stages. At wk 2, 70–81% of infected foci in X-CGD and iNOS KO livers had failed to attract any of the early mononuclear cell reaction already evident at 69–81% of parasitized Kupffer cells in control mice (Table II and Figs. 2, A and B, and 3, A and B). Thus, both phox-derived ROI and iNOS-derived RNI, presumably acting together, appear to participate in the early recruitment of T cells and monocytes into infected tissue. This observation further supports the expanding immunoregulatory spectrum of these molecules, particularly NO (10, 26–28), and in iNOS KO mice may also reflect suppressed induction of IL-12 and IFN-γ (17), key antileishmanial cytokines involved in both mononuclear cell influx and macrophage activation (2–4).
Figure 2.
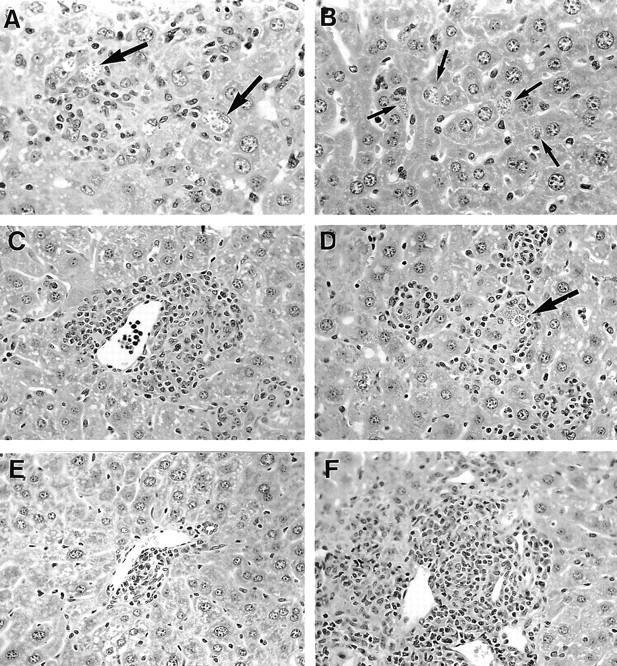
Histologic response to L. donovani infection in the liver in X-CGD versus C57BL/6 mice. 2 wk after challenge, granulomas are developing at infected foci (arrows) in C57BL/6 controls (A); in X-CGD mice (B), there is little or no reaction at well-parasitized foci (arrows). At wk 4, granulomas in C57BL/6 mice (C) in this field are mature and largely parasite free; (D) shows developing granulomas in X-CGD mice. At wk 8, (E) shows receding granulomas in C57BL/6 mice, whereas (F) illustrates intense, mature reaction in X-CGD mice. Original magnification: A and B, ×315; C–F, ×200.
At wk 4, granuloma formation emerged at 90% of infected sites in X-CGD mice (Fig. 2 D) and at 68% of foci in iNOS KO mice (Fig. 3, D and F, and Table II). However, further assembly into mature granulomas (present at 31–37% of foci in control mice; Figs. 2 C and 3, C and E) was retarded and <10% of infected foci in either group of mutant mice could be scored as mature granulomas (Table II). In X-CGD mice, AG treatment inhibited early granuloma assembly (Table I), supporting an active role for iNOS in the initial tissue response. However, granulomas developed by wk 4 despite AG treatment, suggesting a mechanism unrelated to phox or iNOS or suboptimal pharmacologic inhibition of iNOS.
Results at wk 8 indicated that neither phox nor iNOS alone was ultimately required for cell recruitment and assembly, since granulomas developed in both types of deficient mice (Table II). In X-CGD animals, capable of killing L. donovani, the tissue reaction was widespread, comprised of both mature granulomas and inflammatory foci devoid of parasites (Fig. 2 F), and as in a prior study (29) was exaggerated at some perivascular areas (data not shown). However, despite the presence of a similarly well-developed inflammatory reaction, mature-appearing granulomas in iNOS-deficient mice contained overwhelming numbers of replicating amastigotes defining a novel structure, the “ineffective granuloma” (Figs. 3 H and 4).
Together, these results demonstrate clear-cut, probable interdigitating roles for both the respiratory burst and endogenous iNOS–derived RNI as macrophage antimicrobial mechanisms in the initial host defense response to L. donovani. However, the activity of the respiratory burst in inflammatory cell recruitment and in limiting intracellular replication is early and transient, since granulomas formed and infection resolved in X-CGD mice. Similar findings of enhanced susceptibility followed by control in X-CGD mice have been reported in a short-term model of Listeria monocytogenes infection (30).
It is possible that mechanisms related to neither phox nor iNOS, such as other sources of ROI and RNI, also emerge to complete the induction of the tissue cellular immune response to L. donovani and contribute additional antileishmanial effects. However, our results clearly illustrate an obligatory role for iNOS in intracellular killing in vivo and resolution of visceral infection. The presence in iNOS-deficient livers of numerous granulomas, well-established but heavily-laden with intracellular parasites (Fig. 4), graphically illustrates the tissue consequences of the absence of this specific macrophage antimicrobial mechanism.
Figure 4.
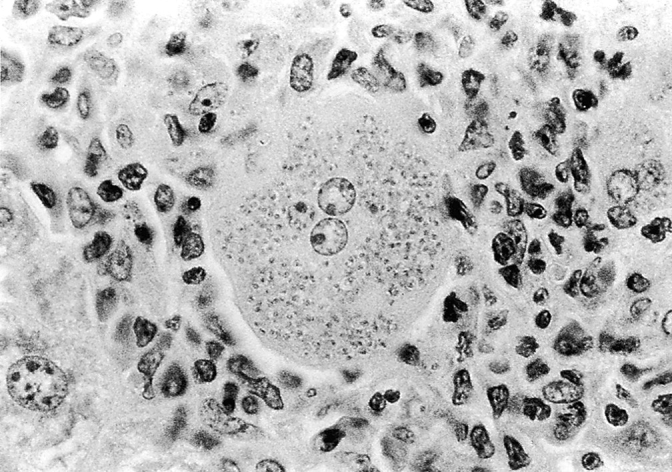
The ineffective granuloma. Photomicrograph shows well-developed tissue structure in 8-wk-infected iNOS KO liver containing a striking load of intracellular amastigotes. Original magnification: ×500.
Acknowledgments
We thank M. Dinauer for providing X-CGD breeders; M. Shiloh, S. Nicholson and S. Potter for their help in establishing the animal colonies; D. Sacks for providing L. donovani; and A. Delph and S. Delph-Etienne for technical assistance.
This work was supported by National Institutes of Health grants AI16963 (to H.W. Murray) and HL51967 (to C.F. Nathan).
References
- 1.Tumang M, Keogh C, Moldawer LL, Teitelbaum RF, Hariprashad J, Murray HW. The role and effect of tumor necrosis factor–alpha in experimental visceral leishmaniasis. J Immunol. 1994;153:768–775. [PubMed] [Google Scholar]
- 2.Taylor A, Murray HW. Intracellular antimicrobial activity in the absence of interferon-γ: effect of interleukin 12 in experimental visceral leishmaniasis in interferon-γ gene-disrupted mice. J Exp Med. 1997;185:1231–1239. doi: 10.1084/jem.185.7.1231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Murray HW. Endogenous interleukin 12 regulates acquired resistance in experimental visceral leishmaniasis. J Infect Dis. 1997;175:1477–1479. doi: 10.1086/516482. [DOI] [PubMed] [Google Scholar]
- 4.Murray, H.W. 1999. Granulomatous inflammation: host antimicrobial defense in the tissues in visceral leishmaniasis. In Inflammation: Basic Principles and Clinical Correlates. J.I. Gallin, R. Snyderman, D.T. Fearon, B.F. Haynes, and C.F. Nathan, editors. Lippincott-Raven, Philadelphia. In press.
- 5.Wilson ME, Sandor M, Blum AM, Younbag B, Metwali A, Elliot D, Lynch RG, Weinstock JV. Local suppression of IFN-γ in hepatic granulomas correlates with tissue-specific replication of Leishmania chagasi. . J Immunol. 1996;156:2231–2239. [PubMed] [Google Scholar]
- 6.Murray HW. Cell-mediated immune response in experimental visceral leishmaniasis. II. Oxygen-dependent killing of intracellular Leishmania donovaniamastigotes. J Immunol. 1982;129:351–357. [PubMed] [Google Scholar]
- 7.Murray HW. Effect of continuous administration of interferon-gamma in experimental visceral leishmaniasis. J Infect Dis. 1990;161:992–994. doi: 10.1093/infdis/161.5.992. [DOI] [PubMed] [Google Scholar]
- 8.Roach TI, Kiderlen AF, Blackwell JM. Role of inorganic nitrogen oxides and tumor necrosis factor-alpha in killing Leishmania donovani amastigotes in gamma-interferon lipopolysaccharide-activated macrophages from Lsh s and Lsh rcongenic mouse strains. Infect Immun. 1991;59:3935–3944. doi: 10.1128/iai.59.11.3935-3944.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Shiloh, M.U., J.D. MacMicking, S. Nicholoson, J.E. Brause, S. Potter, M. Marino, F. Fang, M. Dinauer, and C. Nathan. 1999. Phenotype of mice and macrophages deficient in both phagocyte oxidase and inducible nitric oxide synthase. Immunity. In press. [DOI] [PubMed]
- 10.Fang FC. Mechanisms of nitric oxide-related antimicrobial activity. J Clin Invest. 1997;99:2818–2825. doi: 10.1172/JCI119473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Murray HW, Cartelli DM. Killing of intracellular Leishmania donovaniby human mononuclear phagocytes: evidence for oxygen-dependent and -independent leishmanicidal activity. J Clin Invest. 1983;72:32–39. doi: 10.1172/JCI110972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Murray HW, Teitelbaum RF. L-arginine- dependent reactive nitrogen intermediates and the antimicrobial effect of activated human mononuclear phagocytes. J Infect Dis. 1992;165:513–517. doi: 10.1093/infdis/165.3.513. [DOI] [PubMed] [Google Scholar]
- 13.Murray HW, Szuro-Sudol A, Wellner D, Oca MJ, Granger AM, Libby DM, Rothermel CD, Rubin BY. Role of tryptophan degradation in respiratory burst- independent antimicrobial activity of gamma interferon-stimulated human macrophages. Infect Immun. 1989;57:845–849. doi: 10.1128/iai.57.3.845-849.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Evans TG, Thai L, Granger DL, Hibbs JB. Effect of in vivo inhibition of nitric oxide production in murine leishmaniasis. J Immunol. 1993;151:907–915. [PubMed] [Google Scholar]
- 15.Wei XQ, Charles IG, Smith A, Ure J, Feng GJ, Huang FP, Xu D, Muller W, Moncada S, Liew FY. Altered immune responses in mice lacking inducible nitric oxide synthase. Nature. 1995;375:408–411. doi: 10.1038/375408a0. [DOI] [PubMed] [Google Scholar]
- 16.Stenger S, Thuring H, Rollinghoff M, Bogdan C. Tissue expression of inducible nitric oxide synthase is closely associated with resistance to Leishmania major. . J Exp Med. 1994;180:783–793. doi: 10.1084/jem.180.3.783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Diefenbach A, Schindler H, Donhauser N, Lorenz E, Laskay T, McMicking J, Rollinghoff M, Gresser I, Bogdan C. Type I interferon (IFNα/β) and type 2 nitric oxide synthase regulate the innate immune response to a protozoan parasite. Immunity. 1998;8:77–87. doi: 10.1016/s1074-7613(00)80460-4. [DOI] [PubMed] [Google Scholar]
- 18.Evans TG, Reed SS, Hibbs JB. Nitric oxide production in murine leishmaniasis: correlation of progressive infection with increasing systemic synthesis of nitric oxide. Am J Trop Med Hyg. 1996;54:486–489. doi: 10.4269/ajtmh.1996.54.486. [DOI] [PubMed] [Google Scholar]
- 19.Huang F-P, Xu D, Esfandiari E-O, Sands W, Wei X, Liew FY. Mice defective in Fas are highly susceptible to Leishmania majorinfection despite elevated IL-12 synthesis, strong Th1 responses, and enhanced nitric oxide production. J Immunol. 1998;160:4143–4147. [PubMed] [Google Scholar]
- 20.Melby PC, Yang Y-Z, Cheng J, Zhao W. Regional differences in the cellular immune response to experimental cutaneous or visceral leishmaniasis with Leishmania donovani. . Infect Immun. 1998;66:18–27. doi: 10.1128/iai.66.1.18-27.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bories C, Scherman E, Bories PN. Serum and tissue nitrate levels in murine visceral leishmaniasis correlate with parasite load but not with host protection. Trans R Soc Trop Med Hyg. 1997;91:433–436. doi: 10.1016/s0035-9203(97)90271-6. [DOI] [PubMed] [Google Scholar]
- 22.Nicholson, S., M. da G. Bonecini-Almeida, J.R. Lapa e Silva, C. Nathan, Q.W. Xie, R. Mumford, J.R. Weidner, J. Calaycay, J. Geng, N. Boechat, et al. 1996. Inducible nitric oxide synthase in pulmonary alveolar macrophages from patients with tuberculosis. J. Exp. Med. 183:2293–2302. [DOI] [PMC free article] [PubMed]
- 23.McMicking JD, Nathan C, Hom G, Chartrain N, Fletcher DS, Trumbauer M, Stevens K, Xie Q, Sokol K, Hutchinson N, et al. Altered responses to bacterial infection and endotoxic shock in mice lacking inducible nitric oxide synthase. Cell. 1995;81:641–650. doi: 10.1016/0092-8674(95)90085-3. [DOI] [PubMed] [Google Scholar]
- 24.Pollock JD, Williams PA, Gifford G, Li LL, Du X, Fisherman J, Orkin SH, Doershuck CM, Dinhauer MC. Mouse model of X-linked chronic granulomatous disease, an inherited defect in phagocyte superoxide production. Nat Genet. 1995;9:202–208. doi: 10.1038/ng0295-202. [DOI] [PubMed] [Google Scholar]
- 25.Chan J, Tanaka K, Carroll D, Flynn J, Bloom BR. Effect of nitric oxide synthase inhibitors on murine infection with Mycobacterium tuberculosis. . Infect Immun. 1995;63:736–740. doi: 10.1128/iai.63.2.736-740.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Nathan C. Inducible nitric oxide synthase: what difference does it make? . J Clin Invest. 1997;100:2417–2423. doi: 10.1172/JCI119782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hogaboam CM, Chensue SW, Steinhauser ML, Huffnagle GB, Lukacs NW, Strieter RM, Kunkel SL. Alteration of the cytokine phenotype in an experimental lung granuloma model by inhibiting nitric oxide. J Immunol. 1997;159:5585–5593. [PubMed] [Google Scholar]
- 28.Bogdan C. The multiplex function of nitric oxide in (auto)immunity. J Exp Med. 1998;187:1361–1365. doi: 10.1084/jem.187.9.1361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Morgenstern DE, Gifford MAC, Li LL, Doerschuk CM, Dinauer MC. Absence of respiratory burst in X-linked chronic granulomatous disease mice leads to abnormalities in both host defense and inflammatory response to Aspergillus fumigatus. . J Exp Med. 1997;185:207–218. doi: 10.1084/jem.185.2.207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Dinauer MC, Deck MB, Unanue ER. Mice lacking reduced nicotinamide adenine dinucleotide phosphate oxidase activity show increased susceptibility to early infection with Listeria monocytogenes. . J Immunol. 1997;158:5581–5583. [PubMed] [Google Scholar]