

Abstract
The velocity of rolling leukocytes is thought to be determined by the expression of adhesion molecules and the prevailing wall shear stress. Here, we investigate whether rapid cleavage of L-selectin may be an additional physiologic regulatory parameter of leukocyte rolling. A unique protease in the membrane of leukocytes cleaves L-selectin after activation, resulting in L-selectin shedding. The hydroxamic acid–based metalloprotease inhibitor KD-IX-73-4 completely prevented L-selectin shedding in vitro and significantly decreased the rolling velocity of leukocytes in untreated wild-type C57BL/6 mice from 55 to 35 μm/s in vivo. When E-selectin was expressed on the endothelium (tumor necrosis factor [TNF]-α treatment 2.5–3 h before the experiment), rolling velocity was 4 μm/s and did not change after the application of KD-IX-73-4. However, KD-IX-73-4 decreased mean rolling velocity by 29% from 23 to 16 μm/s in E-selectin–deficient mice treated with TNF-α. The reduction of velocity caused by KD-IX-73-4 was immediate (<5 s) after injection of KD-IX-73-4 as shown by a novel method using a local catheter. These results establish a role for L-selectin shedding in regulating leukocyte rolling velocity in vivo.
Keywords: mouse, inflammation, protease inhibitor, trafficking, velocity
Leukocyte rolling is the initial step in the recruitment of leukocytes to the site of acute inflammation (1, 2). L-, E-, and P-selectin are transmembrane glycoproteins known to mediate leukocyte rolling on microvascular endothelial cells (3–5). L-selectin (CD62L), a molecule expressed exclusively on the surface of leukocytes, interacts with a yet unidentified ligand on venular endothelial cells and participates in mediating leukocyte rolling under physiologic conditions (5, 6). It has been hypothesized that leukocyte rolling is accomplished by rapid formation and subsequent release of bonds between selectins and their ligand(s) (7, 8). A high rate of bond detachment of L-selectin under tension has been predicted (8) and is supported by experimental evidence (9, 10). The present study is designed to explore whether rapid cleavage of bound L-selectin may be responsible for releasing the trailing end of rolling leukocytes from the endothelial surface during rolling.
L-selectin is rapidly shed from the surface of the leukocytes upon their activation (11, 12). Stimuli inducing endoproteolytic release of L-selectin include chemokines like IL-8 and other chemoattractants (N-formylmethionyl leucylphenylalanine, platelet activating factor [12]). L-selectin shedding may also be one of the relevant factors in the recruitment of polymorphonuclear granulocytes (PMNs)1 from the bone marrow, since a reduction of L-selectin expression on polymorphonuclear leukocytes has been reported after their release from bone marrow (13).
L-selectin is cleaved at a membrane-proximal site between Lys321 and Ser322 in a region that links the second short consensus repeat (SCR) and the transmembrane domain (14). The protease releases a large soluble extracellular fragment (sL-selectin) of 68 kD and a 6-kD transmembrane peptide fragment (15). Soluble L-selectin is functionally active, and can inhibit leukocyte attachment to the endothelium (16). Although other cell markers (CD14, CD16, CD43, CD44, and CD50) also show a downregulation upon activation, the shedding of L-selectin is distinct, because it is unusually rapid and resistant to common protease inhibitors (12, 17–19), including the tissue inhibitor of metalloproteinases (TIMP [20]). An unidentified metalloprotease is believed to be responsible for the shedding of L-selectin (20). The activity of this metalloprotease can be effectively inhibited by a hydroxamic acid–based metalloprotease, KD-IX-73-4 (21). A similar hydroxamate-based metalloprotease inhibitor, N-(d,l-[2-(hydroxyaminocarbonyl)-methyl]-4-methylpentano)-l-3-(tert-butyl)-alanyl- l-alanine, 2-aminoethyl amide, blocks the release of TNF, TNF receptors (CD120a and CD120b), IL-6 receptor, and L-selectin release from leukocytes (22). The hydroxamic acid–based inhibitor of zinc-dependent matrix metalloproteinases, Ro 31-9790, has also been shown to inhibit L-selectin shedding from leukocytes (20).
Previous in vitro studies have shown that incubation of neutrophils with KD-IX-73-4 alters their rolling and causes increased neutrophil accumulation on immobilized MECA-79 antigen coated on the lower wall of a flow chamber (23). However, later in vitro results from Allport et al. with human neutrophils on activated human umbilical vein endothelial cells (HUVECs) did not show any regulatory function for L-selectin shedding in terms of neutrophil attachment, rolling, or transmigration (24). Motivated by the controversial role of L-selectin shedding, and the lack of any plausible explanation for the abundance of soluble L-selectin in vivo, we hypothesized a role for L-selectin shedding in the mechanism of physiologic leukocyte rolling.
To determine whether and to what extent L-selectin shedding affects leukocyte rolling, we designed intravital microscopic observations of rolling leukocytes before and after inhibition of L-selectin shedding. We also developed a new method for local application of compounds directly into the microcirculation of mouse cremaster muscle, which allows observations of rolling leukocytes during or briefly after their contact with the protease inhibitor KD-IX-73-4 or appropriate control peptide. We used this method to assess the contribution of rapid L-selectin cleavage to physiologic leukocyte rolling.
Materials and Methods
Protease Inhibitor.
KD-IX-73-4 (HO-NH-CO-CH2-CH(CH2-CH(CH3)2)-CO-Nal-Ala-NH2) was a gift from Dr. T.K. Kishimoto (Boehringer Ingelheim Pharmaceuticals, Inc., Ridgefield, CT). 0.5 mg of the protease inhibitor KD-IX-73-4 was solubilized in 50 μl DMSO and diluted with saline to a final concentration of 1 mg/ml. 150 μg of KD-IX-73-4 was used in in vivo experiments for an estimated initial serum concentration of 50 μg/ml.
A related hydroxamic acid–based compound (slower isomer [SI]) HO-NH-CO-CH2-CH(CH2-CH(CH3)2)-CO-Nal-Ala-NH-CH2-CH2-NH2 was obtained from Peptides International. SI was solubilized, diluted, and aliquoted as described for KD-IX-73-4. SI was used in the experiments as a negative control. SI did not show any inhibitory effect on downregulation of L-selectin from neutrophils after activation with PMA (100 ng/ml; flow cytometry).
mAbs.
mAb RB40.34 (rat IgG1, 30 μg/mouse) is a blocking mAb against murine P-selectin and was purified from hybridoma supernatant (25). mAb 9A9 (rat IgG1, 30 μg/mouse) is a blocking mAb against murine E-selectin (26, 27). TNF-α (0.5 μg/ mouse) was obtained from Genzyme Corp.
Animals.
Experiments were performed on a total of 30 male mice 8–10 wk old and weighing 22–26 g. Mice included wild-type C57BL/6 (Hilltop), gene-targeted mice deficient in L-selectin (28), and gene-targeted mice deficient in E-selectin (29). Both L- and E-selectin knockout mice were backcrossed into a C57BL/6 background for at least seven generations.
Intravital Microscopy.
For intravital microscopy, mice were anesthetized with an intraperitoneal injection of ketamine hydrochloride (100 mg/kg, Ketalar; Parke-Davis) after pretreatment with xylazine (0.05 mg/kg i.p.) and atropine (0.1 mg/kg i.p.; Elkins-Sinn). Animals were kept at 37°C with a thermo-controlled heating pad. Some mice were pretreated 2 or 6 h before surgery with an intrascrotal injection of 0.5 μg murine TNF-α (Genzyme Corp.) in 0.3 ml isotonic saline as described (26). In the 6-h TNF-α model (30), the intrascrotally injected volume contained 30 U Heparin (Elkins-Sinn). Also, these mice were injected with 100 μg of mAb RB40.34 and 100 μg of mAb 9A9 intraperitoneally before the TNF-α treatment.
Local Catheter.
For local injection of the protease inhibitor KD-IX-73-4 and the negative control SI into the microcirculation of the cremaster muscle, a heparinized catheter was placed into the proximal part of the right femoral artery and advanced towards the branching section of the internal iliac artery from the common iliac artery, as shown in Fig. 1. The solutions (KD-IX-73-4 and SI) were injected as a 0.1-ml bolus containing 50 μg peptide. Upon injection of the bolus into the vasculature of the cremaster muscle, a brief hemodilution of the blood flowing through the observed venule indicated passage of the injected fluid. Centerline velocity and hematocrit returned to normal shortly after the passage of the bolus through the venule.
Figure 1.

Schematic of local catheter. Stream of the injected fluid (black arrow) mixes with blood from the arteria iliaca communis (gray arrow) into arteria iliaca interna (white arrows) to reach the microcirculation of the mouse cremaster muscle.
Cremaster.
The cremaster muscle was prepared for intravital microscopy as described (31) and superfused with thermo-controlled 35°C bicarbonate-buffered saline saturated with 95% N2/ 5% CO2 (26). The exposed cremaster microcirculation remained well perfused. Time 0 was set at the beginning of the cremaster surgery, which took ∼15 min. In the case of TNF-α–pretreated animals, time 0 was set at the time of scrotal injection with 500 ng TNF-α in 0.3 ml saline.
Microscopic observations were made on an Axioskope intravital microscope (Carl Zeiss) with a saline immersion objective (SW 40/0.75 numerical aperture). Each venule was observed for 90–120 s. Venules with diameters between 25 and 45 μm were observed, and video recordings were made through a CCD camera system (model VE-1000CD; Dage-MTI) on a Panasonic S-VHS recorder. Microvascular centerline red blood velocity was measured using a dual photodiode and a digital on-line cross-correlation program (32). Centerline velocities were converted to mean blood flow velocities by multiplying with an empirical factor of 0.625 (33). Shear rates were determined as γw = 2.12 × 8V b/d, where γw is the wall shear rate, V b is the mean blood flow velocity, d is the in vivo diameter of the vessel, and 2.12 is a median empirical correction factor obtained from actual velocity profiles measured in microvessels in vivo (34).
For continuous blood pressure monitoring and blood sampling, the carotid artery was cannulated with a heparinized PE-10 tubing. Blood samples of 10 μl volume each were taken before and 10 min after the systemic application of the protease inhibitor KD-IX-73-4 and SI to analyze systemic leukocyte concentrations (1:9 dilution with Kimura [11 ml of 5% (wt/wt) toluidine blue, 0.8 ml of 0.03% light green SF yellowish, 0.5 ml of saturated saponin, and 5 ml of 0.07 M phosphate buffer, pH 6.4; all reagents obtained from Sigma]). Three different vessels in each cremaster preparation were selected for observations before and 1–15 min after the application of the protease inhibitor KD-IX-73-4 or the negative control SI. All vessels had calculated wall shear rates between 600 and 800 s−1. Microvessel diameters and individual rolling leukocyte velocity were measured using a digital image processing system (32). Freeze-frame advancing was used to accurately monitor the movements of the individual rolling leukocytes. Each rolling leukocyte passing a line perpendicular to the vessel wall was followed for 0.5–1 s. Rolling velocities for individual leukocytes were calculated by dividing the traveled distance by the tracking time. Critical velocities were calculated for each vessel (35). The leukocyte rolling flux was determined from video recordings by counting all visible cells passing through a plane perpendicular to the vessel axis for 1 min before and 1 min after the systemic application of the protease inhibitor, starting 1 min after injection.
Flow Cytometry.
Mice were injected with KD-IX-73-4, or vehicle alone, and blood was obtained from the subclavian vein at 1 min after the injection. Mouse peripheral blood was stained with PE-conjugated anti–L-selectin mAb clone (cat. 01265B; PharMingen) or PE-conjugated isotype control mAb (rat IgG2a, cat. 11025A; PharMingen) 2 μl/ml at 4°C for 15 min. After lysis of the erythrocytes (lysing solution: 0.15 M NH4Cl, 0.01 M NaHCO3, and 0.001 M disodium EDTA), cells were washed once with BSA/PBS and fixed in 1% paraformaldehyde/PBS, and 0.5–1 × 104 cells/sample were analyzed by flow cytometry on a FACScan® (Becton Dickinson). The results reported are for granulocytes, gated by their characteristic forward and side scatter.
Statistical Analysis.
Statistical comparisons were carried out using Kolmogorov-Smirnov two-sample t test. NCSS statistical software (http://www.icw.com/ncss) version 6.0.11 was used for the statistical analysis. Statistical significance was set at P < 0.05 or P < 0.01.
Results
Inhibition of L-selectin Shedding Lowers Leukocyte Rolling Velocity under Physiologic Conditions In Vivo.
Fig. 2 shows the results of inhibition of L-selectin shedding in wild-type mice. Velocities of 750 rolling leukocytes in 15 vessels of 5 wild-type mice were obtained before and after the application of KD-IX-73-4. Observations were made 1–8 min after the injection of the protease inhibitor and within the first 1 h after starting the cremaster preparation. The mean velocity of all observed cells before the inhibition of L-selectin shedding was 59 ± 5.6 μm/s and after the inhibition was 39 ± 3.6 μm/s, which represents a reduction of the mean velocity by 33 ± 2%. As an example, Fig. 3 shows a composite photomicrograph illustrating the change in rolling velocity in one representative vessel.
Figure 2.

Cumulative frequency of velocities of rolling leukocytes in wild-type mice. Each data point shows the fraction of all rolling leukocytes travelling below the indicated velocity. Open circles, velocities of rolling leukocytes on endothelium of cremaster muscle venules after acute exteriorization. Filled circles, velocities of rolling leukocytes after systemic application of 150 μg protease inhibitor, KD-IX-73-4 (significantly lower, P < 0.01). Data obtained between 1 and 8 min after intravenous injection of the protease inhibitor. Each data point stands for the mean of 15 cells, obtained from 5 animals.
Figure 3.
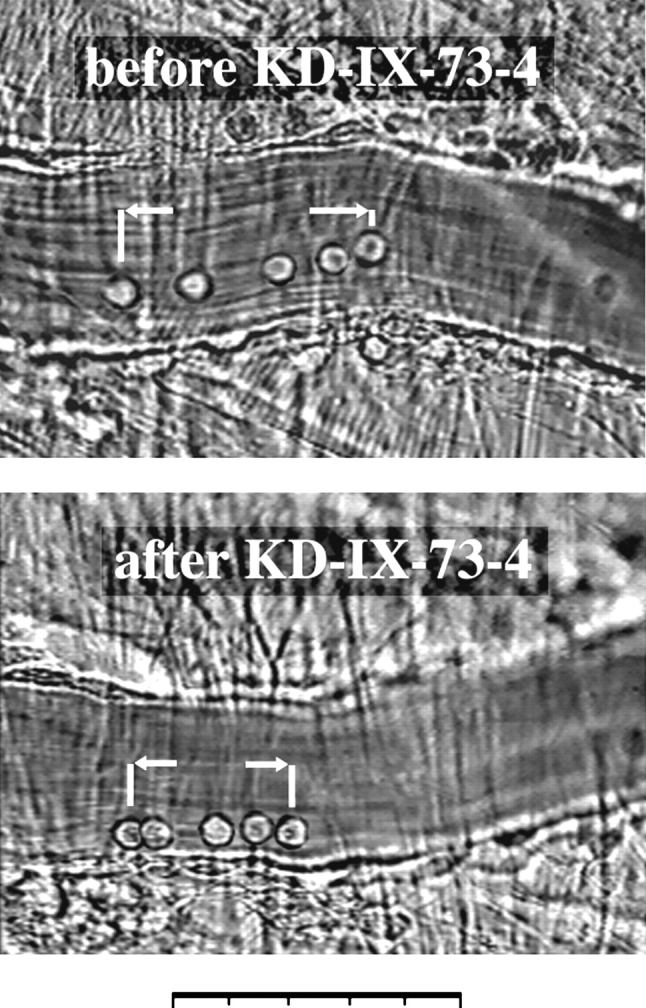
Composite of five superimposed video frames before and after application of KD-IX-73-4 from the same postcapillary venule of a wild-type mouse. Time elapsed between the first and last frame in each picture is 0.8 s. The distance traveled by a leukocyte in each panel illustrates the regulatory function of L-selectin shedding on leukocyte rolling velocity. Bar, 50 μm.
The related hydroxamic acid–based substance (SI) did not show any significant effect on the velocities of the rolling leukocytes. Interestingly, after the use of 150 μg SI (1 μg/ μl), the subsequent injection of 150 μg KD-IX-73-4 did not show any change in the velocity of rolling leukocytes, suggesting a possible competitive mechanism of interaction between the two hydroxamic acid–based substances and the L-selectin cleaving protease. Fig. 4 shows the results of the SI application followed by KD-IX-73-4 in wild-type mice.
Figure 4.

Cumulative frequency of velocities of rolling leukocytes in wild-type mice before and after application of SI and KD-IX-73-4, 150 μg. Open circles, after acute exteriorization; open triangles, after SI application; filled circles, after SI followed by KD-IX-73-4 application. Six measurements in two animals per data point. Velocities of 50 rolling cells in each vessel obtained before and after the injection of the control substance SI and after the subsequent injection of the protease inhibitor KD-IX-73-4 (no significant velocity difference).
To determine the duration of the effect of the protease inhibitor in vivo, single venule recordings were made in two wild-type animals for 15–20 min after the application of KD-IX-73-4. Rolling velocities of 10 cells were obtained in 1-min steps after the injection of 150 μg KD-IX-73-4. Within the first 1 min after the injection of 150 μg KD-IX-73-4, the reduction of rolling velocity could be observed and persisted for ∼9–10 min, after which the rolling velocity returned to control levels (Fig. 5).
Figure 5.

Leukocyte rolling velocities from single vessel recordings after application of 150 μg protease inhibitor KD-IX-73-4. Rolling velocities of 10 cells per vessel obtained at 1-min intervals to follow the change of the mean velocity of these cells over time. Each data point is the mean of 20 rolling leukocytes from 2 wild-type mice.
To assess whether the injected dose of KD-IX-73-4 resulted in complete inhibition of L-selectin shedding in vivo, we investigated the expression of L-selectin on neutrophils in peripheral blood by flow cytometry (Fig. 6). All neutrophils in peripheral blood of untreated mice expressed L-selectin, which was almost entirely downregulated after PMA activation. In contrast, PMNs obtained from mice 1 min after KD-IX-73-4 injection did not shed L-selectin in response to PMA activation. The inhibitory effect of KD-IX-73-4 in vivo was no longer detectable at 15 min after the injection. SI, the peptide used as a control, did not show any inhibitory effect on L-selectin shedding after PMA activation of the cells. Taken together, these data show that KD-IX-73-4 transiently inhibits L-selectin shedding in vivo. The time course of this inhibitory effect parallels the time course of reduction of rolling velocity by KD-IX-73-4.
Figure 6.


Flow cytometry of mouse PMNs for expression of L-selectin with PE-conjugated mAb, Mel-14. a, PMNs, no mAb; b, L-selectin expression without activation; c, L-selectin expression after activation with PMA (100 ng/ml, 20 min at 37°C); d, 1 min after KD-IX-73-4 injection (150 μg); e, 1 min after KD-IX-73-4 injection and subsequent PMA activation; f, 15 min after KD-IX-73-4 injection and subsequent PMA activation; and g, 1 min after SI injection and subsequent PMA activation.
Systemic application of 150–300 μg KD-IX-73-4 did not cause an alteration of the systemic blood pressure, which remained between 80 and 95 mmHg throughout the experiments. Likewise, centerline velocities and diameters (and consequently the shear rates) of the observed venules were not affected by the systemic injections of KD-IX-73-4 or SI. Also, systemic white blood cell counts remained unchanged after systemic injection of KD-IX-73-4 (Table I).
Table I.
Leukocyte Counts and Hemodynamic Parameters, Mean ± SEM
| White blood cell counts | Velocity | Diameter | Wall shear rate, γ | |||||
|---|---|---|---|---|---|---|---|---|
| Before KD-IX-73-4 | 4.3 ± 0.4 | 2.8 ± 0.3 | 39 ± 5 | 758 ± 67 | ||||
| After KD-IX-73-4 | 4.2 ± 0.3 | 2.7 ± 0.3 | 40 ± 4 | 770 ± 58 | ||||
| After SI | 4.4 ± 0.3 | 2.6 ± 0.2 | 46 ± 3 | 610 ± 32 | ||||
To determine whether the injected amount of KD-IX-73-4 (150 μg) in vivo was sufficient to cause maximal reduction of velocity, subsequent injections of KD-IX-73-4 (150–300 μg) were made in two wild-type animals to achieve an accumulation of the protease inhibitor in plasma. Such injections did not cause a further shift of velocities of the rolling cells compared with the cell rolling velocities after the first injection of 150 μg KD-IX-73-4 (data not shown). Therefore, an injection of 150 μg KD-IX-73-4 into a mouse produced complete inhibition of L-selectin shedding and maximal reduction of rolling velocity in vivo.
As another parameter relevant for leukocyte trafficking, the flux of rolling leukocytes (number of rolling cells passing per unit of time) was measured in eight vessels of wild-type mice. Inhibition of L-selectin shedding caused a significant 22% increase of rolling flux compared with the mean of the rolling flux of the same vessels before the KD-IX-73-4 treatment (118 cells/min). This finding suggests that inhibition of L-selectin shedding not only reduces the velocity of rolling leukocytes, but also aids in promoting leukocyte capture from the free stream and/or stabilizes their rolling along the venular tree. Furthermore, the number of firmly adherent leukocytes was significantly increased after inhibition of L-selectin shedding in mice treated with TNF-α for 6 h (data not shown). In this model, rolling is primarily L-selectin dependent (30). Concomitant with increased adhesion, inhibition of L-selectin shedding also increased the number of transmigrating cells, apparent by the characteristic change of shape during their transendothelial passage (data not shown). This suggests that, beyond its prominent and immediate effect on the velocity of rolling leukocytes, L-selectin shedding significantly impacts leukocyte recruitment.
To investigate whether the effect of KD-IX-73-4 on the leukocyte velocities in vivo is specific to the inhibition of L-selectin shedding or whether other variables contribute to the shift of velocities, experiments with L-selectin–deficient mice were made. Fig. 7 shows leukocyte rolling velocities obtained in L-selectin–deficient mice before and after systemic application of KD-IX-73-4. There was no significant change of the mean rolling velocities measured after application of KD-IX-73-4 (37 ± 1.4 μm/s before and 38.2 ± 1.5 μm/s after the inhibition; 100 cells from 2 animals). The absence of the reduction of velocity in L-selectin–deficient mice shows that inhibition of L-selectin shedding with KD-IX-73-4 does not alter the rolling velocity of leukocytes unless L-selectin is expressed. Flux also remained unchanged after injection of KD-IX-73-4 in L-selectin–deficient mice (data not shown).
Figure 7.

Cumulative frequency of velocities of rolling leukocytes in L-selectin–deficient mice before and after application of KD-IX-73-4. Open circles, after acute exteriorization; filled circles, after KD-IX-73-4 application (150 μg). No reduction of rolling velocity in L-selectin–deficient mice. Mean velocity of 100 rolling cells, obtained from 2 male wild-type animals, is 37 ± 1.4 μm/s before the inhibition of L-selectin shedding, and 38.2 ± 1.5 μm/s after the injection of KD-IX-73-4 (not significant).
The systemic application of the protease inhibitor into the jugular vein is likely to produce a homogenous serum concentration of the inhibitor throughout the vascular system. However, in order to be able to make a statement about the onset of the effect of KD-IX-73-4 on rolling velocity, it was necessary to observe rolling leukocytes from the moment of their first contact with the inhibitor. For this purpose, the protease inhibitor was injected as a small bolus (50 μg/0.1 ml) through a local catheter (Fig. 1) into the microcirculation of cremaster muscle. With the passage of the solution through the cremaster vasculature, the inhibitor washed over rolling leukocytes in the cremaster venules and might inhibit L-selectin shedding in these cells within the field of view. A brief hemodilution effect caused by injection of the bolus furnished evidence for the passage of the solution through the observed venules. Within the first 5 s after the passage of the bolus, velocities of 25 rolling leukocytes/venule were obtained. The mean velocity of 50 rolling leukocytes from 2 animals before contact with the protease inhibitor was 40.2 ± 3.1 μm/s, which was lowered to 29.9 ± 2.7 μm/s immediately after the injection of KD-IX-73-4. This equals a 26% reduction of leukocyte rolling velocity.
To explore the role of L-selectin shedding during cytokine-induced inflammation, we used intrascrotal injection of TNF-α 3 h before the observation period, as described previously (6). Treatment of wild-type mice with intrascrotal injections of 0.5 μg TNF-α 2 h before the experiments causes the expression of E-selectin and increased expression of P-selectin on the endothelial cells (36). Presence of E-selectin on the endothelium of the cremaster venules lowers the mean leukocyte rolling velocity by almost one order of magnitude to under 5 μm/s (37). Fig. 8 A shows results of inhibition of L-selectin shedding with KD-IX-73-4 in TNF-α–treated wild-type mice. The mean velocity of 300 rolling leukocytes (4.5 μm/s) before application of the protease inhibitor did not change significantly compared with the mean of 300 rolling leukocytes (4.1 μm/s) obtained between 1 and 6 min after the application of 150 μg KD-IX-73-4.
Figure 8.
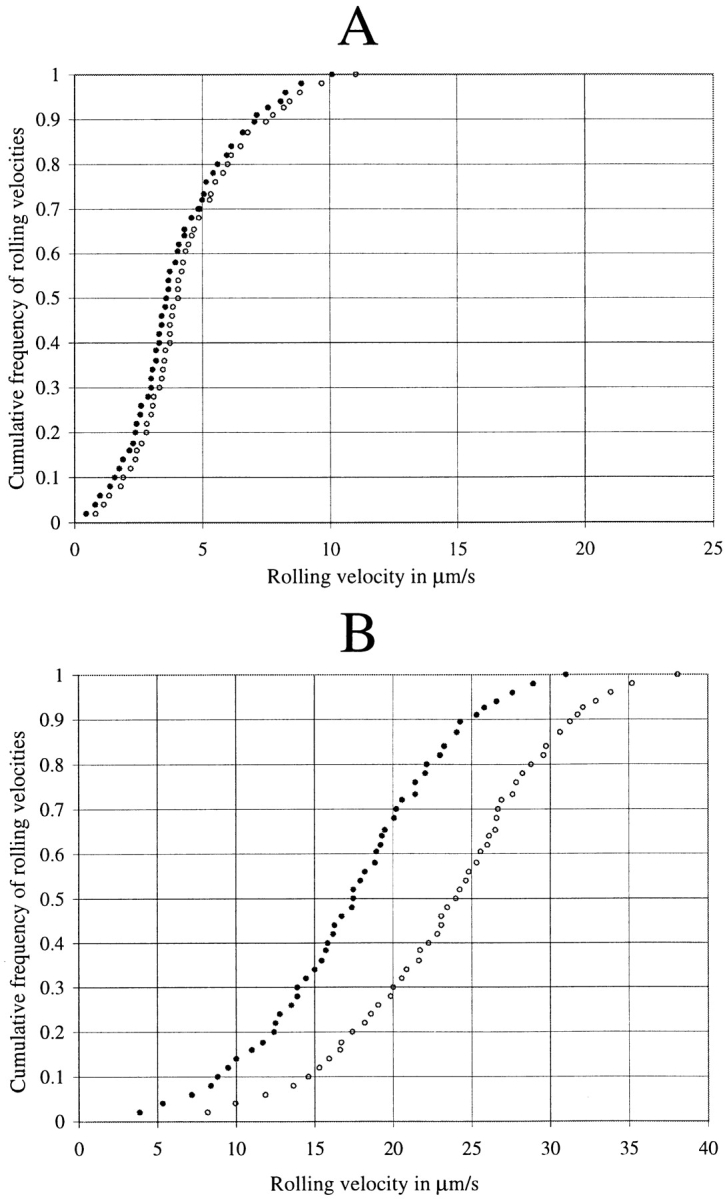
(A) Cumulative frequency of velocities of rolling leukocytes in TNF-α–treated wild-type mice (3 h before the injection of the protease inhibitor). Open circles, before; filled circles, after KD-IX-73-4 injection (150 μg). Mean value of six measurements from two animals per data point. (B) Cumulative frequency of velocities of rolling leukocytes in TNF-α–treated E-selectin–deficient mice. Mean value of four measurements from two animals per data point.
We hypothesized that E-selectin–mediated leukocyte rolling proceeded at such low velocities that inhibition of L-selectin shedding did not influence the actual velocity because the low off-rate of E-selectin (38) would determine and dominate the rolling velocity, no matter whether L-selectin was being shed or not. Previous studies had shown that rolling velocities were significantly higher in E-selectin– deficient mice treated with TNF-α than in wild-type mice (37). To explore the reason for the apparent absence of an effect of inhibiting L-selectin shedding in TNF-α–treated mice, we used TNF-α–treated E-selectin–deficient mice. In these mice, mean rolling velocity was 24 μm/s and was reduced to 17 μm/s after treatment with KD-IX-73-4 (a 29% reduction of mean rolling velocity; Fig. 8 B).
Discussion
Although proteolytic cleavage of L-selectin from the surface of leukocytes was described as early as 1989 (11, 12), the physiologic relevance of L-selectin shedding in vivo has remained unclear and the subject of controversial speculations. The results of this study establish L-selectin shedding as a novel parameter, in addition to adhesion molecule expression and hemodynamic forces, relevant for regulation of leukocytes rolling. Continuous L-selectin shedding appears necessary for physiologic leukocyte rolling at typical velocities. Though Walcheck et al. have noted that inhibition of L-selectin shedding lowers the rolling velocity of isolated neutrophils on immobilized MECA-79 antigen (23), the results of other in vitro experiments using cultured endothelial cells seemed to contradict their conclusion (24). In experiments with TNF-α–treated HUVECs, a role for L-selectin shedding in neutrophil attachment, rolling, or transmigration could not be shown. Our results in untreated and TNF-α–treated wild-type mice can explain this discrepancy and establish the role of L-selectin shedding for leukocyte trafficking.
When E-selectin is expressed on endothelial cells, inhibition of L-selectin shedding has no effect on leukocyte rolling velocity. However, when E-selectin is not expressed, either in wild-type mice not treated with an inflammatory cytokine, or in E-selectin–deficient mice, the modulatory role of L-selectin shedding is evident. In TNF-α–treated cultured HUVECs (24), E-selectin is likely to be expressed. This would explain why Allport et al. (24) did not find a velocity difference between human neutrophils with or without incubation with a zinc-dependent metalloproteinase inhibitor (Ro 31-9790). In the experiment of Walcheck et al. (23), E-selectin is not present, because a purified L-selectin ligand (MECA-79) is used as the sole adhesive substrate. We investigated the effect of inhibiting L-selectin on rolling in vivo both in the presence and absence of E-selectin. In the absence of E-selectin, inhibition of L-selectin shedding causes a significant decrease of rolling velocity (as in reference 23), which is not detectable when E-selectin is involved in mediating leukocyte rolling (as in reference 24).
As presented in Fig. 2, a shift towards lower leukocyte rolling velocities can be seen with the inhibition of L-selectin shedding in wild-type mice without intentional induction of inflammation. The amount of protease inhibitor used in these animals provided a maximum inhibition of L-selectin shedding, since further injections of KD-IX-73-4 did not lower the rolling velocity of the cells further. The shift of velocity in these animals shows that normal leukocyte rolling requires continuous shedding of L-selectin. Consistent with this, Palecanda et al. detected soluble L-selectin in the plasma of healthy adults, whose peripheral blood leukocytes did not demonstrate any obvious signs of activation (39), and Schleiffenbaum et al. found high physiologic levels of soluble L-selectin in the serum of healthy volunteers (16). Since no soluble or secreted splice variants of L-selectin are known, we hypothesize that continuous L-selectin shedding during physiologic leukocyte rolling is a likely source for soluble L-selectin in the plasma of healthy individuals.
The fact that no change in the rolling velocity can be measured with the protease inhibitor KD-IX-73-4 in L-selectin– deficient mice shows that the observed shift of velocities in wild-type mice was specifically and exclusively caused by the inhibition of L-selectin shedding. Other molecules such as P- or E-selectin, which also mediate leukocyte rolling, do not contribute to the shift of velocity caused by the application of KD-IX-73-4.
As a negative control, we used the hydroxamic acid– based substance (SI), which is structurally closely related to the protease inhibitor KD-IX-73-4 but does not inhibit L-selectin shedding as shown by flow cytometry. Interestingly, after the use of SI, subsequent injections of KD-IX-73-4 did not show a shift of velocity. Therefore, SI may interact with the protease responsible for L-selectin shedding at the same binding site as KD-IX-73-4, thus competing with KD-IX-73-4 binding. PMNs from KD-IX-73-4– treated wild-type mice do not shed L-selectin in response to PMA activation. However, the inhibitory effect of KD-IX-73-4 in vivo is not permanent, as PMNs obtained 15 min after KD-IX-73-4 treatment responded to PMA activation by shedding L-selectin. This parallels our observations from the functional assay (Fig. 5) that the shift of velocity lasts for ∼10 min after one injection. The inhibition of L-selectin shedding in vivo and the velocity shift in the functional assay can both be observed after the injection of the protease inhibitor KD-IX-73-4. These data are consistent with a causal relationship between the injection of KD-IX-73-4, the inhibition of the L-selectin shedding, and the shift of rolling velocity.
The current findings show that proteolytic cleavage of L-selectin shedding is relevant for physiologic leukocyte rolling. Since enzymatic processes often require metabolic energy, it may be worthwhile exploring whether L-selectin–mediated leukocyte rolling is modulated by cell metabolism or whether it is as independent of cell metabolism as formerly believed (40, 41). Furthermore, in light of the present study, a revision of the calculated dissociation rate of L-selectin appears necessary, which was made with the assumption that during stable rolling the average number of bonds that form and break will be equal (9). This assumption may prove erroneous, since inhibition of L-selectin shedding decreases rolling velocity. Therefore, we believe that the 7.5–11.5-fold faster L-selectin–mediated rolling does not necessarily require a 7–10-fold more rapid bond dissociation, as proposed by Alon et al. (9). Rather, continuous cleavage of L-selectin and breakage of bonds are likely to occur at the same time and thus cause short duration of L-selectin–mediated tethering events.
In conclusion, inhibition of L-selectin shedding decreases the leukocyte rolling velocity and increases the leukocyte rolling flux in vivo. A decreased rolling velocity increases the transit time of rolling leukocytes. Leukocyte transit times have recently been shown to be an important determinant of leukocyte recruitment in vivo (42). Modulation of L-selectin shedding is a novel and influential parameter in the leukocyte trafficking by virtue of its regulatory function on rolling velocity, rolling flux, and transit time of leukocytes.
Acknowledgments
We wish to thank Dr. T.K. Kishimoto for the gift of KD-IX-73-4 and for insightful discussion. We thank William Ross for help with flow cytometry experiments, and Nicholas Douris and Jennifer Bryant for mouse husbandry. L-selectin–deficient mice were from a colony based on breeders provided by Dr. T.F. Tedder, Duke University, Durham, NC. E-selectin–deficient mice were from a colony based on breeders provided by Dr. D.C. Bullard and Dr. A.L. Beaudet, Baylor College of Medicine, Houston, TX.
This work was supported by National Institutes of Health grant HL54136 to K. Ley.
Abbreviations used in this paper
- HUVEC
human umbilical vein endothelial cell
- PMN
polymorphonuclear granulocyte
- SI
slower isomer (negative control)
References
- 1.Ley K, Tedder T. Leukocyte interactions with vascular endothelium. New insights into selectin-mediated attachment and rolling. J Immunol. 1995;155:525–528. [PubMed] [Google Scholar]
- 2.Springer T. Traffic signals on endothelium for lymphocyte recirculation and leukocyte emigration. Annu Rev Physiol. 1995;57:827–872. doi: 10.1146/annurev.ph.57.030195.004143. [DOI] [PubMed] [Google Scholar]
- 3.Dore M, Korthuis R, Granger D, Entman M, Smith C. P-selectin mediates spontaneous leukocyte rolling in vivo. Blood. 1993;82:1308–1316. [PubMed] [Google Scholar]
- 4.Kansas G. Selectins and their ligands: current concepts and controversies. Blood. 1996;88:3259–3287. [PubMed] [Google Scholar]
- 5.Ley K, Gaehtgens P, Fennie C, Singer M, Lasky L, Rosen S. Lectin-like cell adhesion molecule 1 mediates leukocyte rolling in mesenteric venules in vivo. Blood. 1991;77:2553–2555. [PubMed] [Google Scholar]
- 6.Ley K, Bullard D, Arbones M, Bosse R, Vestweber D, Tedder T, Beaudet A. Sequential contribution of L-and P-selectin to leukocyte rolling in vivo. J Exp Med. 1995;181:669–675. doi: 10.1084/jem.181.2.669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hammer D, Apte S. Simulation of cell rolling and adhesion on surfaces in shear flow: general results and analysis of selectin-mediated neutrophil adhesion. Biophys J. 1992;63:35–57. doi: 10.1016/S0006-3495(92)81577-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tözeren A, Ley K. How do selectins mediate leukocyte rolling in venules? . Biophys J. 1992;63:700–709. doi: 10.1016/S0006-3495(92)81660-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Alon R, Chen S, Puri K, Finger E, Springer T. The kinetics of L-selectin tethers and the mechanics of selectin-mediated rolling. J Cell Biol. 1997;138:1169–1180. doi: 10.1083/jcb.138.5.1169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Puri K, Chen S, Springer T. Modifying the mechanical property and shear threshold of L-selectin adhesion independently of equilibrium properties. Nature. 1998;392:930–933. doi: 10.1038/31954. [DOI] [PubMed] [Google Scholar]
- 11.Jutila M, Rott L, Berg E, Butcher E. Function and regulation of the neutrophil MEL-14 antigen in vivo: comparison with LFA-1 and MAC-1. J Immunol. 1989;143:3318–3324. [PubMed] [Google Scholar]
- 12.Kishimoto T, Jutila M, Berg E, Butcher E. Neutrophil Mac-1 and MEL-14 adhesion proteins inversely regulated by chemotactic factors. Science. 1989;245:1238–1241. doi: 10.1126/science.2551036. [DOI] [PubMed] [Google Scholar]
- 13.van Eeden S, Miyagashima R, Haley L, Hogg J. A possible role for L-selectin in the release of polymorphonuclear leukocytes from bone marrow. Am J Physiol. 1997;272:H1717–H1724. doi: 10.1152/ajpheart.1997.272.4.H1717. [DOI] [PubMed] [Google Scholar]
- 14.Chen A, Engel P, Tedder TF. Structural requirements regulate endoproteolytic release of the L-selectin (CD62L) adhesion receptor from the cell surface of leukocytes. J Exp Med. 1995;182:519–530. doi: 10.1084/jem.182.2.519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kahn J, Ingraham R, Shirley F, Migaki G, Kishimoto T. Membrane proximal cleavage of L-selectin: identification of the cleavage site and a 6-kD transmembrane peptide fragment of L-selectin. J Cell Biol. 1994;125:461–470. doi: 10.1083/jcb.125.2.461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Schleiffenbaum B, Spertini O, Tedder T. Soluble L-selectin is present in human plasma at high levels and retains functional activity. J Cell Biol. 1992;119:229–238. doi: 10.1083/jcb.119.1.229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bazil V, Strominger J. Metalloprotease and serine protease are involved in cleavage of CD43, CD44, and CD16 from stimulated human granulocytes. Induction of cleavage of L-selectin via CD16. J Immunol. 1994;152:1314–1322. [PubMed] [Google Scholar]
- 18.Kishimoto T, Kahn J, Migaki G, Mainolfi E, Shirley F, Ingraham R, Rothlein R. Regulation of L-selectin expression by membrane proximal proteolysis. Agents Actions Suppl. 1995;47:121–134. doi: 10.1007/978-3-0348-7343-7_11. [DOI] [PubMed] [Google Scholar]
- 19.Shipp M, Stefano G, Switzer S, Griffin J, Reinherz E. CD10 (CALLA)/neutral endopeptidase 24.11 modulates inflammatory peptide-induced changes in neutrophil morphology, migration, and adhesion proteins and is itself regulated by neutrophil activation. Blood. 1991;78:1834–1841. [PubMed] [Google Scholar]
- 20.Preece G, Murphy G, Ager A. Metalloproteinase-mediated regulation of L-selectin levels on leucocytes. J Biol Chem. 1996;271:11634–11640. doi: 10.1074/jbc.271.20.11634. [DOI] [PubMed] [Google Scholar]
- 21.Feehan C, Darlak K, Kahn J, Walcheck B, Spatola A, Kishimoto T. Shedding of the lymphocyte L-selectin adhesion molecule is inhibited by a hydroxamic acid-based protease inhibitor. Identification with an L-selectin-alkaline phosphatase reporter. J Biol Chem. 1996;271:7019–7024. doi: 10.1074/jbc.271.12.7019. [DOI] [PubMed] [Google Scholar]
- 22.Bennett T, Lynam E, Sklar L, Rogelj S. Hydroxamate-based metalloprotease inhibitor blocks shedding of L-selectin adhesion molecule from leukocytes: functional consequences for neutrophil aggregation. J Immunol. 1996;156:3093–3097. [PubMed] [Google Scholar]
- 23.Walcheck B, Kahn J, Fisher J, Wang B, Fisk R, Payan D, Feehan C, Betageri R, Darlak K, Spatola A, Kishimoto T. Neutrophil rolling altered by inhibition of L-selectin shedding in vitro. Nature. 1996;380:720–723. doi: 10.1038/380720a0. [DOI] [PubMed] [Google Scholar]
- 24.Allport J, Ding H, Ager A, Steeber D, Tedder T, Luscinskas F. L-selectin shedding does not regulate human neutrophil attachment, rolling, or transmigration across human vascular endothelium in vitro. J Immunol. 1997;158:4365–4372. [PubMed] [Google Scholar]
- 25.Bosse R, Vestweber D. Only simultaneous blocking of the L- and P-selectin completely inhibits neutrophil migration into mouse peritoneum. Eur J Immunol. 1994;24:3019–3024. doi: 10.1002/eji.1830241215. [DOI] [PubMed] [Google Scholar]
- 26.Kunkel E, Jung U, Bullard D, Norman K, Wolitzky B, Vestweber D, Beaudet A, Ley K. Absence of trauma-induced leukocyte rolling in mice deficient in both P-selectin and intercellular adhesion molecule 1. J Exp Med. 1996;183:57–65. doi: 10.1084/jem.183.1.57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Norton C, Rumberger J, Burns D, Wolitzky B. Characterization of murine E-selectin expression in vitrousing novel anti-mouse E-selectin monoclonal antibodies. Biochem Biophys Res Commun. 1993;195:250–258. doi: 10.1006/bbrc.1993.2037. [DOI] [PubMed] [Google Scholar]
- 28.Arbones M, Ord D, Ley K, Ratech H, Maynard-Curry C, Otten G, Capon D, Tedder T. Lymphocyte homing and leukocyte rolling and migration are impaired in L-selectin-deficient mice. Immunity. 1994;1:247–260. doi: 10.1016/1074-7613(94)90076-0. [DOI] [PubMed] [Google Scholar]
- 29.Bullard D, Kunkel E, Kubo H, Hicks M, Lorenzo I, Doyle N, Doerschuk C, Ley K, Beaudet A. Infectious susceptibility and severe deficiency of leukocyte rolling and recruitment in E-selectin and P-selectin double mutant mice. J Exp Med. 1996;183:2329–2336. doi: 10.1084/jem.183.5.2329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Jung U, Ramos C, Bullard D, Ley K. Gene-targeted mice reveal importance of L-selectin-dependent rolling for neutrophil adhesion. Am J Physiol. 1998;43:H1785–H1791. doi: 10.1152/ajpheart.1998.274.5.H1785. [DOI] [PubMed] [Google Scholar]
- 31.Baez S. An open cremaster muscle preparation for the study of blood vessels by in vivomicroscopy. Microvasc Res. 1973;5:384–394. doi: 10.1016/0026-2862(73)90054-x. [DOI] [PubMed] [Google Scholar]
- 32.Pries A. A versatile video image analysis system for microcirculatory research. Int J Microcirc Clin Exp. 1988;7:327–345. [PubMed] [Google Scholar]
- 33.Lipowsky H, Zweifach B. Application of the “two-slit” photometric technique to the measurement of microvascular volumetric flow rates. Microvasc Res. 1978;15:93–101. doi: 10.1016/0026-2862(78)90009-2. [DOI] [PubMed] [Google Scholar]
- 34.Reneman, R., B. Woldhuis, M. oude Egbrink, D. Slaaf, and G. Tangelder. 1992. Concentration and velocity profiles of blood cells in the microcirculation. In Advances in Cardiovascular Engineering. N. Hwang, V. Turitto, and M. Yen, editors. Plenum Publishing Corp., New York. 25–40.
- 35.Ley K, Gaehtgens P. Endothelial, not hemodynamic, differences are responsible for preferential leukocyte rolling in rat mesenteric venules. Circ Res. 1991;69:1034–1041. doi: 10.1161/01.res.69.4.1034. [DOI] [PubMed] [Google Scholar]
- 36.Jung U, Ley K. Regulation of E-selectin, P-selectin, and intercellular adhesion molecule 1 expression in mouse cremaster muscle vasculature. Microcirculation. 1997;4:311–319. doi: 10.3109/10739689709146794. [DOI] [PubMed] [Google Scholar]
- 37.Kunkel E, Ley K. Distinct phenotype of E-selectin-deficient mice. E-selectin is required for slow leukocyte rolling in vivo. Circ Res. 1996;79:1196–1204. doi: 10.1161/01.res.79.6.1196. [DOI] [PubMed] [Google Scholar]
- 38.Kaplanski G, Farnarier C, Tissot O, Pierres A, Benoliel AM, Alessi MC, Kaplanski S, Bongrand P. Granulocyte-endothelium initial adhesion. Analysis of transient binding events mediated by E-selectin in a laminar shear flow. Biophys J. 1993;64:1922–1933. doi: 10.1016/S0006-3495(93)81563-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Palecanda A, Walcheck B, Bishop D, Jutila M. Rapid activation-independent shedding of leukocyte L-selectin induced by crosslinking of the surface antigen. Eur J Immunol. 1992;22:1279–1286. doi: 10.1002/eji.1830220524. [DOI] [PubMed] [Google Scholar]
- 40.Lalor P, Clements J, Pigott R, Humphries M, Spragg J, Nash G. Association between receptor density, cellular activation, and transformation of adhesive behavior of flowing lymphocytes binding to VCAM-1. Eur J Immunol. 1997;27:1422–1426. doi: 10.1002/eji.1830270619. [DOI] [PubMed] [Google Scholar]
- 41.Lawrence M, Springer T. Neutrophils roll on E-selectin. J Immunol. 1993;151:6338–6346. [PubMed] [Google Scholar]
- 42.Jung U, Norman K, Scharffetter-Kochanek K, Beaudet A, Ley K. Transit time of leukocytes rolling through venules controls cytokine-induced inflammatory cell recruitment in vivo. J Clin Invest. 1998;102:1526–1533. doi: 10.1172/JCI119893. [DOI] [PMC free article] [PubMed] [Google Scholar]