

Abstract
Different T cell subsets exhibit distinct capacities to migrate into peripheral sites of inflammation, and this may in part reflect differential expression of homing receptors and chemokine receptors. Using an adoptive transfer approach, we examined the ability of functionally distinct subsets of T cells to home to a peripheral inflammatory site. The data directly demonstrate the inability of naive T cells and the ability of effector cells to home to inflamed peritoneum. Furthermore, interleukin (IL)-12 directs the differentiation of either CD4+ or CD8+ T cells into effector populations that expresses functional E- and P-selectin ligand and that are preferentially recruited into the inflamed peritoneum compared with T cells differentiated in the presence of IL-4. Recruitment can be blocked by anti–E- and –P-selectin antibodies. The presence of antigen in the peritoneum promotes local proliferation of recruited T cells, and significantly amplifies the Th1 polarization of the lymphocytic infiltrate. Preferential recruitment of Th1 cells into the peritoneum is also seen when cytokine response gene 2 (CRG-2)/interferon γ–inducible protein 10 (IP-10) is used as the sole inflammatory stimulus. We have also found that P-selectin binds only to antigen-specific T cells in draining lymph nodes after immunization, implying that both antigen- and cytokine-mediated signals are required for expression of functional selectin-ligand.
Keywords: T helper cells type 1, T helper cells type 2, lymphocyte homing receptors, interleukin 12, P-selectin
Awidely accepted model of T lymphocyte recirculation suggests that naive T cells preferentially migrate from lymph node to lymph node until they interact with antigen in the specialized lymph node microenvironment. Antigen and other signals provided in the lymph node induce differentiation of the naive T cells into effector and memory T cells that leave the lymph node and preferentially migrate to inflamed tissues (1–4). The distinct migration patterns of naive and effector/memory T cells is presumably due to differential expression of adhesion molecules, called homing receptors, on naive T cells and effector or memory T cells (5–8). For example, L-selectin on naive T cells binds to PNAd on high endothelial venules in lymph nodes and this molecular interaction mediates, at least in part, the homing of naive T cells to lymph nodes (9–11). The peripheral recruitment of helper T cells to inflammatory sites is governed in part by the expression of adhesion molecules on the T cells that bind to ligands induced on cytokine activated endothelium. Effector and memory T cells express higher levels of integrins (e.g., CD11aCD18 and CD29dCD49) and CD44 compared with naive T cells (5). In addition, functional selectin ligand expression appears to be absent on naive T cells and present on effector/memory T cells. For example, we have previously shown that in humans, CD45RA−RO+ memory but not CD45RA+RO− naive T cells roll efficiently on E- and P-selectin under flow (13). These differences in the surface phenotypes are likely to contribute to the preferential recruitment of effector or memory T cells, but not naive T cells to peripheral sites of inflammation. Analysis of expression of a number of cell surface markers indicates that naive T cells are normally absent from sites of inflammation in mice and humans (2–4). However, there are few reported studies in which comparative migration of naive versus activated T cells to inflammatory sites in vivo have been directly examined.
Naive CD4+ lymphocytes can differentiate into polarized subsets of effector/helper T cells, characterized by distinct profiles of cytokine production (14). The exogenous cytokine milieu at the time of initial antigen stimulation is a major determinant of the pathway of helper T cell differentiation into either Th1 cells, which produce IFN-γ, or Th2 cells, which produce IL-4 and IL-5 (14, 15). In addition to the distinct profiles of cytokine expression, Th1 and Th2 cells may also express distinct sets of surface structures that mediate recruitment patterns to peripheral inflammatory sites. Recent research has focused on expression of selectin ligands and chemokine receptor on Th1 and Th2 cells.
Functional ligands for E- and P-selectin appear to be more highly expressed on Th1 cells than on Th2 cells (16, 17), and this corresponds to an enhanced ability of Th1 cells to enter delayed-type hypersensitivity sites compared with Th2 cells (16). We and others have recently shown that IL-12 provides a necessary signal for induction of functional selectin ligands in vitro, and that IL-12 upregulates α3-fucosyltransferase VII, a key enzyme in the production of the carbohydrate ligands that bind to P- and E-selectin (18–20). The relative contributions of antigen versus IL-12 signals in inducing functional selectin ligand expression are not known. Furthermore, the appearance of functional selectin ligand on antigen activated T cells in vivo has not been described in any detail.
Th1 and Th2 cells also express distinct, but partially overlapping, sets of chemokine receptors (21). IP-10 is a CXC chemokine that is produced by endothelial cells and macrophages stimulated by IFN-γ (23). IP-10 induces rapid and transient adhesion of human IL-2–stimulated T lymphocytes to endothelial cells through its receptor CXCR3, which is selectively expressed on activated T cells (24, 25). In addition, CXCR3 is more abundantly expressed on human Th1 cells than on Th2 cells, and as a result IP-10 selectively attracts Th1 cells in in vitro migration assays (26–28). CRG-2 has been identified to be the murine homologue of IP-10 (29). However, the effect of CRG-2 on differentiated murine Th1 and Th2 cells is not known.
Subsets of CD8+ cytolytic T lymphocytes (CTLs)1 have been defined on the basis of the cytokines they produce (30–33). CTLs that predominantly secrete IFN-γ and IL-2 are called Tc1 cells, by analogy to Th1 cells, whereas CTLs that predominantly secrete IL-4, IL-5, and IL-10 are called Tc2 cells. Recent studies have indicated that Tc1 and Tc2 cells have stable phenotypes in vivo after adoptive transfer (34), and that these subsets may have different patterns of migration into inflammatory sites (35). The possibility that Tc1 and Tc2 cells differ in expression of functional selectin ligands has not been explored.
In the studies presented here, we have examined the differential recruitment of mouse T cell subsets to a peripheral inflammatory site. We have used adoptive transfer of naive or differentiated TCR transgenic CD4+ T cells into normal mice, and adoptive transfer of fluorescently labeled, Staphylococcus enterotoxin B (SEB)–stimulated CD4+ and CD8+ T cells. Our results directly demonstrate differences in recruitment capabilities among both CD4+ and CD8+ T cell subsets, and the importance of functional selectin ligand expression and chemokines in selective recruitment. Furthermore, we have exploited the DO11 TCR transgenic adoptive transfer model to examine the effect of antigen on both the induction of T cell selectin ligands in vivo and on the accumulation of Th1 cells at an inflammatory site.
Materials and Methods
Mice.
BALB/c mice, 4–6 wk of age, were purchased from The Jackson Laboratory. Transgenic mice expressing the DO11.10 TCR (DO11) (36), specific for the chicken OVA peptide OVA323–339 in the context of the MHC class II molecule I-Ad, were obtained from Dr. D. Loh (Hoffmann-LaRoche, Nutley, NJ). The DO11 mice were bred in our pathogen and viral antibody–free facility in accordance with guidelines of the Committee on Animals of the Harvard Medical School, and those prepared by the Committee on Care and Use of Laboratory Animals of the Institute of Laboratory Resources, National Research Council (Washington, D.C.). The mice were typed for the DO11 TCR by staining peripheral blood cells with antibodies against CD4 and Vβ8 as previously described (37).
Reagents.
Murine IL-4 was obtained from the culture supernatant of the I3XL6 cell line (obtained from Dr. Abul Abbas, Brigham and Women's Hospital), which constitutively expresses a stably transfected murine IL-4 gene. Recombinant murine IL-12 was a gift of Dr. Stan Wolf (Genetics Institute, Cambridge, MA). IL-2–containing supernatants were obtained from the X63-IL2 cell line, which expresses a stably transfected murine IL-2 gene (obtained from Dr. Fritz Melchers, Basel Institute of Immunology, Basel, Switzerland). The hybridoma cell line producing anti–IL-4 (clone 11B11) was obtained from the American Type Culture Collection. Neutralizing anti–mouse IFN-γ (XMG1.2) was purchased from PharMingen. Human E- and P-selectin human IgG chimeric proteins (38, 39) were provided by Dr. R. Camphausen (Genetics Institute). OVA323–339 was produced by the peptide synthesis facility of the Center for Neurologic Diseases, Brigham and Women's Hospital. Murine recombinant IP-10/CRG-2 was obtained from R&D Systems.
Purification and In Vitro Differentiation of T Cells.
Naive CD4+ T cells were purified from pooled lymph nodes and spleen cells from DO11 mice by using CD4+ Dynalbeads and Detach-a-bead reagent (Dynal A.S., Norway) according to the manufacturer's instructions. More than 95% of the cells were CD4+ as assessed by flow cytometry using FITC-conjugated anti–mouse CD4+ mAbs. The CD4− fraction of the DO11 spleen and lymph node suspensions, or whole spleen cell suspensions from BALB/c mice, were treated with mitomycin C for 30 min at 37°C and used as APCs for DO11 T cells. T cell differentiation was induced by culturing 2 × 105 purified CD4+ TCR transgenic T cells in 1 ml of RPMI 1640 with 10% FCS with 2 × 106 APCs, 1 μM OVA323–339, and Th0 condition (no cytokines), Th1 condition (10 ng/ml IL-12 plus anti–IL-4 [11B11 hybridoma supernatant, 25% vol/vol]), or Th2 condition (1,000 U/ml IL-4 plus anti– IFN-γ [XMG 1.2, 1 μg/ml]). The cultures were fed with fresh medium containing 10 U/ml IL-2 after 4 d, and harvested for adoptive transfer on day 6.
In some experiments, suspensions of BALB/c mouse lymph node and spleen cells were differentiated in vitro by stimulation with 2 μg/ml SEB (Sigma Chemical Co.) in the presence of cytokine and antibody reagents used for Th1 and Th2 differentiation, as described above. 6 d later, cells were harvested, centrifuged over Ficoll-sodium metrizoate (Cappel), labeled with a fluorescent tracker dye (see below), and used in adoptive transfer studies.
Adoptive Transfers and Immunization.
Freshly prepared suspensions of whole lymph node and spleen cells from DO11 mice were used as a source of naive cells for adoptive transfer, as previously described (37). In vitro differentiated DO11 T cells were harvested at day 6 and centrifuged over Ficoll-sodium metrizoate before adoptive transfer. The number of T lymphocytes expressing the DO11 TCR in each population was measured by flow cytometric analysis of cell suspensions stained with the clonotypic antibody, KJ1-26 (40, 41) (see below). Naive or differentiated T cells (1.5–2.0 × 107 in 500 μl PBS) were adoptively transferred into unirradiated BALB/c recipients by tail vein injection. 6 h later, mice were given intraperitoneal injections of 500 μg OVA protein or PBS emulsified in IFA (Difco) or with IP-10/CRG-2 in PBS. 5, 48, or 72 h after adoptive transfer, spleen and lymph nodes (inguinal, axillary, and mesenteric) were removed and peritoneal cells were harvested by PBS lavage. Cell suspensions were prepared and analyzed by flow cytometry for surface antigen expression.
In some experiments, mice received naive DO11 T cells by tail vein injection, and were immunized 24 h later with OVA (100 μg) emulsified in CFA (Difco) through footpad injection. Draining popliteal and inguinal lymph nodes were harvested at varying times after immunization, and cell suspensions were prepared and were analyzed by flow cytometry.
SEB-stimulated T cell cultures were harvested at day 6, centrifuged over Ficoll-sodium metrizoate, and then incubated with 1 μM Cell Tracker Green (5-chloromethylfluorescein diacetate [CMFDA]; Molecular Probe) diluted in RPMI 1640 with 10% FBS for 20 min at 37°C. Cells were centrifuged and incubated for an additional 30 min at 37°C in fresh medium to allow elution of unreacted dye before transfer. Labeled cells (1.5–2.0 × 107) were then adoptively transferred into BALB/c mice through tail vein injection. 6 h later, mice were given intraperitoneal injections of 500 μl of PBS-IFA. 72 h after transfer, spleen, lymph nodes, and peritoneal cells were harvested and analyzed by flow cytometry.
FACS® Analysis.
In DO11 adoptive transfer studies, suspensions of cells from peritoneum, lymph node, or spleen were first incubated with 5 μg/ml anti-CD16/CD32 (PharMingen) to block Fc receptors. The cells were then stained with 1 μg/ml PE-labeled anti-CD4 antibody (PharMingen), and 1 μg/ml biotinylated KJ1-26 clonotypic antibody followed by Cy-Chrome– streptavidin (PharMingen), as previously described (37). These cells were also stained with 1 μg/ml P-selectin–human IgG chimera (38, 39), followed by FITC-conjugated anti–human IgG (Southern Biotechnology Associates). SEB-stimulated T cells were stained with 1 μg/ml biotinylated anti-Vβ8 antibody (PharMingen) followed by Cy-Chrome–streptavidin (PharMingen). SEB-stimulated cells were also stained with 1 μg/ml PE-labeled anti-CD8 or anti-CD4 antibodies (PharMingen), as well as with soluble P-selectin–IgG and FITC-conjugated anti-IgG, as described above for DO11 cells. More than 95% of the CD4− Vβ8+ T cells in the SEB-stimulated populations were CD8+. We therefore chose to analyze CD4+Vβ8+ and CD4−Vβ8+ T cells in the same staining tubes as a convenient way to compare CD4+ and CD8+ populations while still being able to use two additional fluorochromes for Vβ8 and P-selectin ligand analysis. All analyses were performed on a FACScan® flow cytometer (Becton Dickinson). In some studies, CMFDA (green tracker dye)-labeled T cells were detected by direct fluorescence in the FL1 channel on the flow cytometer.
Results
Naive versus Effector T Cell Recruitment to Inflammatory Sites.
We have recently shown that, in contrast to antigen-stimulated DO11 T cells, naive DO11 T cells fail to roll on selectins under flow conditions in vitro, nor do they bind soluble P- or E-selectin Ig chimeric molecules (18). To determine whether naive T cells are capable of entering a peripheral inflammatory site, we directly transferred spleen and lymph node cells from normal DO11 mice into BALB/c recipients and then further challenged the mice intraperitoneally with IFA. After 3 d, the recipient mice were assayed for the number of adoptively transferred cells in the peritoneum as well as in peripheral lymph node and spleen. As has been previously reported, naive DO11 T cells homed to lymph nodes, and to a lesser extent to spleen (Fig. 1 A). In contrast, DO11 CD4+ T cells were not detectable (<0.2%) in the peritoneum of either the control or IFA-treated mice (Fig. 1 A). The numbers of DO11 CD4+ T cells in both spleen and lymph nodes were comparable in control and IFA-treated mice, indicating that roughly equal number of cells were adoptively transferred into the circulation of each mouse.
Figure 1.


Recruitment of naive versus effector CD4+ T cells to inflamed peritoneum. 1.5 × 107 naive CD4+KJ126+ T cells (A) or 1.8 × 107 in vitro– activated CD4+ T cells (B) were adoptively transferred into BALB/c mice. After 6–24 h, mice were either left untreated (control) or were given intraperitoneal injections of IFA. 3 d later, spleens, lymph nodes, and peritoneal cells were collected and stained with PE-conjugated anti-CD4 and Cy-Chrome– labeled KJ126. The numbers adjacent to the upper right corner of each panel are the percentage of the total gated cells that are CD4+KJ126+ cells. One of two comparable experiments is shown.
The T cells we isolated from normal DO11 mice express high levels of L-selectin and low levels of CD25 and CD44, which is a typical phenotype for naive T cells (Fig. 2). After 6 d in culture with APCs and OVA peptide (see Material and Methods, Th0 condition), DO11 T cells had acquired a typical activated phenotype: L-selectinlowCD25high CD44high (Fig. 2). Upon restimulation with OVA peptide, these activated CD4+ T cells produced both IFN-γ and IL-4, as assayed by ELISA (data not shown). After they were transferred into BALB/c mice, the activated DO11 CD4+ T cells were recruited to the IFA-treated peritoneum (Fig. 1 B). The frequency of CD4+KJ126+ cells in the peritoneum increased from 0.9 ± 0.1% to 3.4 ± 0.3% (n = 2 experiments) in the absence and presence of IFA, respectively. In contrast, the frequency of CD4+KJ126+ cells in the spleens remained comparable at 4.1 ± 0.4% and 3.9 ± 0.3% (n = 2 experiments) in the control and IFA-treated mice, respectively. Thus, we conclude that although both activated and naive CD4+ T cells are capable of entering secondary lymphoid tissues, only the activated subset is recruited to the inflamed peritoneum.
Figure 2.

Phenotype of naive and activated T cells. Naive cells represented in this figure were CD4+ T cells freshly isolated from DO11 mice, and activated cells represent DO11 T cells stimulated in vitro with OVA peptide and APCs, as described in Materials and Methods. The histograms depict staining for the indicated molecule using PE-labeled secondary antibody after gating on CD4+ (FITC) plus KJ126+ (Cy-Chrome) cells. One of two comparable experiments is shown.
Recruitment of Th1 versus Th2 CD4+ Cells to a Site of Inflammation.
To compare the recruitment of Th1 versus Th2 cells to an inflammatory site, we first generated differentiated Th1 and Th2 populations of DO11 T cells in vitro as described in Materials and Methods. Th1 cells produced abundant IFN-γ but no IL-4, whereas Th2 cells produced abundant IL-4 but little IFN-γ, as judged by cytokine ELISA assay and intracellular cytokine staining (reference 18 and data not shown).
DO11 Th1 or Th2 cells were adoptively transferred into syngeneic BALB/c recipient mice, the mice were challenged with intraperitoneal injection of IFA and the number of adoptively transferred CD4+KJ126+ cells in spleen, lymph nodes or peritoneum was determined 3 d later. Results of a typical experiment are shown in Fig. 3. In that experiment, adoptively transferred Th1 cells comprised 3.1% of total peritoneal cells, whereas adoptively transferred Th2 cells comprised only 0.7%. In contrast, Th1 and Th2 cell recruitment into spleen or lymph nodes was approximately equal. In four experiments, the frequency of adoptively transferred Th1 cells in IFA-treated peritoneum was 3.2 ± 1.0 times the frequency of adoptively transferred Th2 cells. When OVA and IFA were injected into the peritoneum, and cells were recovered 3 d later, the percentage of DO11 T cells was significantly enhanced compared with exudates from animals treated with IFA alone. Furthermore, there was a greater difference between the numbers of Th1 and Th2 cells in the peritoneum when OVA was present than when it was absent (Fig. 4). In the experiment shown, adoptively transferred Th1 cells comprised 19% of the total peritoneal cells in the IFA plus OVA-treated peritoneum, whereas Th2 cells comprised only 3.1%. The absolute numbers of DO11 T cells recruited in the peritoneum at various time points during this experiment are shown in Table I. With IFA alone at day 3, 106 DO11 Th1 cells were recruited to the peritoneum, which was five times more than the number of DO11 Th2 cells recruited under identical conditions. In the presence of IFA plus OVA, 3.2 × 106 of DO11 Th1 cells were present in peritoneum at day 3, but only 4 × 105 DO11 Th2 cells were recruited under identical conditions. The increased number of Th1 cells in the peritoneum when OVA is present may be due to increased recruitment and/or local proliferation in response to antigen after recruitment. When DO11 Th1 cells were labeled with tracker dye before transfer, the day 3 recovered peritoneal cells from OVA plus IFA treated mice had significantly less tracker dye than cells recovered from IFA-treated mice (Fig. 5). The loss of tracker dye correlates with cell division and dilution of the intracellular label among progeny cells (37, 42–44). This indicates local proliferation accounts for at least some of the accumulation of Th1 cells when antigen is present.
Figure 3.

In vivo recruitment of Th1 versus Th2 cells to adjuvant-treated peritoneum. DO11 Th1 or Th2 cells (2 × 107) were adoptively transferred into BALB/c mice. After 24 h, mice were given intraperitoneal injections of PBS-IFA. 3 d later, lymph nodes (inguinal, axillary, and mesenteric), spleen, and peritoneal cells were collected and stained with PE-conjugated anti-CD4 and Cy-Chrome–labeled KJ126. The numbers adjacent to the upper right corner of each panel are the percentage of the total gated cells that are CD4+KJ126+ cells. One of four comparable experiments is shown.
Figure 4.
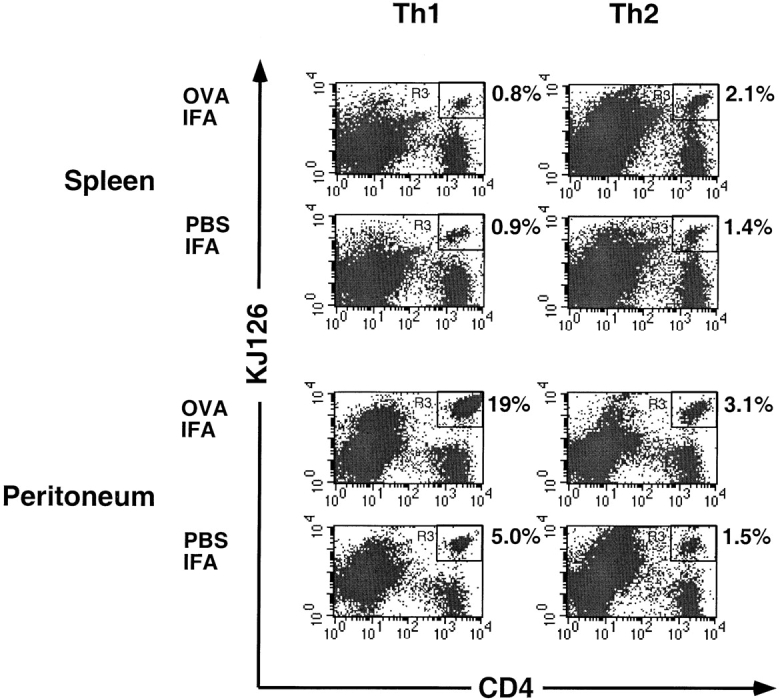
Influence of antigen on recruitment of Th1 versus Th2 cells to peritoneum. DO11 Th1 or Th2 cells (2 × 107) were adoptively transferred into BALB/c mice. After 24 h, mice were given intraperitoneal injections of either OVA-IFA or PBS-IFA. 3 d later, spleen and peritoneal cells were collected and stained with PE-conjugated anti-CD4 and Cy-Chrome–labeled KJ126. The numbers adjacent to the upper right corner of each panel are the percentage of the total gated cells that are CD4+KJ126+ cells. One of four comparable experiments is shown.
Table I.
In Vivo Recruitment of Th1 versus Th2 Cells to Adjuvant-treated Peritoneum
| Time after adoptive transfer | Th1 | Th2 | ||||||
|---|---|---|---|---|---|---|---|---|
| OVA-IFA | PBS-IFA | OVA-IFA | PBS-IFA | |||||
| h | ||||||||
| 5 | 1.1 × 104 | 0.9 × 104 | 3 × 103 | 2 × 103 | ||||
| 48 | 8.3 × 104 | ND | ND | ND | ||||
| 72 | 3.2 × 106 | 106 | 4 × 105 | 2 × 105 | ||||
DO11 Th1 or Th2 cells (2 × 107) were adoptively transferred into BALB/c mice. After 24 h, mice were given intraperitoneal injections of either OVA-IFA or PBS-IFA. 3 d later, peritoneal cells were collected after 5, 48, or 72 h and stained with PE-conjugated anti-CD4 and Cy-Chrome–labeled KJ126. The cell numbers presented here are numbers of DO11 CD4+KJ126+ cells in the peritoneum according to percentage of CD4+KJ126+ cells and total number of peritoneal cells. One of two comparable experiments is shown.
Figure 5.

Antigen-induced proliferation of adoptively transferred Th1 cells in response to antigen in peritoneum. DO11 Th1 cells (2 × 107) were labeled with green tracker (CMFDA) before adoptive transfer into BALB/c mice. Recipient mice were given intraperitoneal injections of either OVA-IFA (OVA+) or PBS-IFA (OVA−), and 3 d later peritoneal cells were harvested and stained with PE-conjugated anti-CD4 and Cy-Chrome–labeled KJ126. The green tracker visualized in FL-1 shown here is gated on CD4+KJ126+ cells.
To determine if the cytokine secretory phenotype of the adoptively transferred Th1 and Th2 cells remained stable in vivo, the recovered peritoneal cells were stimulated with OVA peptide and culture supernatants were assayed for IFN-γ and IL-4 production. Consistent with previous findings (45), DO11 Th1 and Th2 cells maintain their effector cytokine profile in vivo (data not shown).
Role of Selectin Ligand Expression in T Cell Subset–specific Recruitment.
To dissect the mechanisms for differential recruitment of T cell subsets into the inflamed peritoneum, we first examined the role of P- and E-selectins and their ligands on T lymphocytes. T cells constitutively express PSGL-1, the best described receptor for P-selectin, which becomes functional only after posttranslational modification (46, 47). The P-selectin binding status of T cells was examined by FACS® analysis using soluble P-selectin–human IgG fusion protein. As shown in Fig. 6 A, naive CD4+ T cells failed to bind P-selectin. After in vitro differentiation with peptide and specific cytokines, Th1 cells bound much more P-selectin than Th2 cells did. To further investigate whether binding to P- and E-selectin is essential for the selective recruitment of CD4+ T cells to the inflamed peritoneum, we studied the migration of Th1 cells under the influence of antibodies against P- and E-selectin. As shown in Fig. 6 B, combination of antibodies against P- and E-selectin effectively blocked Th1 cells from entering IFA-treated peritoneum. The percentage of DO11 T cells in the peritoneum dropped from 7 to 1.6%, whereas the number of DO11 T cells in spleens remained comparable in these two groups of mice. This data indicates that the interaction of P- and E-selectin with their ligands is essential for maximal recruitment of Th1 cells into sites of inflammation.
Figure 6.


P- and E-selectin–mediated recruitment of Th1 cells into an inflammatory site. (A) Naive, Th1, or Th2 cells were stained with P-selectin Ig chimeric protein (Psel-IgG) or with human Ig control (HuIgG) followed by FITC-conjugated anti–human-Ig. Cells were also stained for CD4 (PE) and KJ126 (Cy-Chrome). The histogram shown represents cells gated on the CD4+KJ126+ population. One of three comparable experiments is shown. (B) Isotype control rabbit IgG or anti–P- and E-selectin antibodies were injected via tail vein together with Th1 cells into BALB/c mice. After 24 h, PBS-IFA was injected into the peritoneum of these mice. 3 d later, spleen and peritoneal cells were collected and stained with PE-conjugated anti-CD4 and Cy-Chrome–labeled KJ126. The numbers adjacent to the upper right corner of each panel are the percentage of the total gated cells that are CD4+KJ126+ cells. One of two comparable experiments is shown.
IP-10/CRG-2 Induced Preferential Recruitment of Th1 Cells to a Peripheral Site.
Chemokines activate integrins on leukocytes and trigger firm interaction of leukocytes to endothelium (48–50). Recently, human CXCR3, receptor for IP-10, was shown to be highly expressed in Th1 but not in Th2 cells (26–28). To demonstrate a possible role of chemokines in mediating selective recruitment of T cell subsets in vivo, we injected the mouse IP-10 homologue, CRG-2, into the peritoneum of mice after adoptive transfer of either DO11 Th1 or Th2 cells. In response to CRG-2 treatment, DO11 Th1 cells were recruited to the peritoneum more efficiently than Th2 cells. In the experiment shown in Fig. 7, Th1 cells comprised 7.9% of the total peritoneal population in CRG-2 treated mice, whereas Th2 cells comprised 2.3% in similarly treated mice. The total numbers of Th1 and Th2 cells harvested from the CRG-2–treated mice were 2.5 × 105 and 4 × 104 respectively. In two experiments in which CRG-2 treatment was performed, the frequency of adoptively transferred Th1 cells in the peritoneum was 3.7 ± 0.4 times the frequency of adoptively transferred Th2 cells. Therefore, differential expression of chemokine receptors and functional P/E-selectin ligand(s) contribute to differences in recruitment of T cell subsets.
Figure 7.

IP-10/CRG-2–mediated preferential recruitment of T helper cells in vivo. DO11 Th1 and Th2 cells were adoptively transferred into BALB/c mice through tail vein injection. After 24 h, mice either were left untreated or else received an intraperitoneal injection of 0.5 μg of CRG-2. 3 d later, spleen and peritoneal cells were collected and stained with PE-conjugated anti-CD4 and Cy-Chrome–labeled KJ126. The numbers adjacent to the upper right corner of each panel are the percentage of the total gated cells that are CD4+KJ126+ cells.
Recruitment of IL-12 versus IL-4 –stimulated CD8+ T Cells to a Site of Inflammation.
In vitro stimulation of BALB/c spleen cells with SEB for 6 d gave rise to activated CD4+ and CD8+ T cells. Over 80% of this mixed population was Vβ8+ (data not shown) and CD8+ T cells comprised from 50 to 66% of the Vβ8+ cells. Under the influence of Th1 promoting cytokines (IL-12 plus anti–IL-4), a majority of CD4+Vβ8+ and CD8+Vβ8+ T cells bound P-selectin (Fig. 8 A). The ratio of CD4+/CD8+ cells propagated under these condition was ∼1:2. On the other hand, under the influence of Th2 promoting cytokines (IL-4 plus anti– IFN-γ), both CD4+Vβ8+ and CD8+Vβ8+ populations failed to bind P-selectin (Fig. 8 A). The CD4+/CD8+ ratio of the cells propagated under these conditions was 1:1.
Figure 8.


The P-selectin binding and peritoneal recruitment of SEB-stimulated BALB/c splenic CD4+ and CD8+ T cells. (A) Spleen cells were stimulated in vitro with 2 μg/ml of SEB with either IL-12 plus anti–IL-4 antibody or with IL-4 plus anti–IFN-γ antibody. After 6 d of culture, cells were harvested and stained for CD4, Vβ8, and P-selectin Ig (Psel-IgG) or control human Ig (HuIgG) as described in previous figures. (B) The same cell populations as described in A were labeled with green tracker dye before tail vein injection into BALB/c mice. After 24 h, mice were injected in the peritoneum with PBS-IFA. 3 d later, spleen and peritoneal cells were collected and visualized by flow cytometry. The numbers in the plots represent the percentage of total gated cells that are tracker dye positive (CMFDA). One of four comparable experiments is shown.
To demonstrate in vivo recruitment of CD4+ and CD8+ SEB-stimulated T cells, the cells were labeled with green tracker dye (CMFDA) before adoptive transfer. As predicted based on P-selectin binding status shown above, a selective recruitment was clearly observed. After 3 d of IFA treatment, IL-12–treated cells were recruited to peritoneum more efficiently than those IL-4–treated cells. As shown in Fig. 8 B, IL-12–treated cells represented 16% of total peritoneal population, whereas IL-4–treated cells only represented 1.5%. In four experiments, the frequency of adoptively transferred cells in the peritoneum that were originally differentiated under Th1 conditions was 6.5 ± 2.8 times greater than the frequency of adoptively transferred cells originally differentiated under Th2 conditions. Furthermore, the CD4+/CD8+ ratio of the Vβ8+ cells recovered from the peritoneum was always the same as in the initial adoptively transferred population (data not shown). This indicates that conditions that promote CD4+ Th1 differentiation also act on CD8+ T cells to induce functional selectin ligands and the ability to migrate into inflammatory sites.
Induction of T Cell P-selectin Ligand Expression In Vivo.
We have demonstrated that naive T cells stimulated with antigen and IL-12 in vitro differentiate into P-selectin binding cells that are capable of entering an inflammatory site. To determine if naive T cells upregulate functional P-selectin ligand in response to antigen in vivo, we adoptively transferred naive DO11 T cells into BALB/c recipient mice. After 24 h, the mice were immunized by footpad injection with OVA peptide and CFA to provide specific antigen as well as a stimulus for IL-12 production. Draining lymph nodes were removed after 1, 2, and 3 d and cells were stained for CD4, DO11 TCR (KJ126), and P-selectin binding. As shown in Fig. 9 A, the DO11 CD4+KJ126+ T cells expanded in lymph nodes in response to OVA stimulation. The percentage of CD4+KJ126+ T cells increased from 3.1% to 8.7% by day 2 and further increased to 13% by day 3. At day 2 after immunization, there was a significant increase in the number of CD4+KJ126+ cells that bound P-selectin (Fig. 9 B). Interestingly, there was no increase in P-selectin binding by the CD4+KJ126− population indicating that cytokines in the lymph node cannot induce P-selectin ligand expression on T cells not directly stimulated by antigen. Forward scatter analysis indicated that the P-selectin binding cells were the largest, presumably most activated cells among the CD4+KJ126+ population (data not shown).
Figure 9.


Induction of functional P-selectin ligand expression on antigen-activated T cells in vivo. Naive (1.5 × 107) CD4+KJ126+ T cells were adoptively transferred into BALB/c mice. After 24 h, mice were either left untreated or further immunized with OVA and CFA by footpad injection. Draining lymph nodes were collected at days 1, 2, and 3, and were stained with PE-conjugated anti-CD4 and Cy-Chrome–labeled KJ126 and with P-selectin Ig chimeric protein (Psel-IgG) or with human Ig control (HuIgG) followed by anti–human Ig conjugated with FITC. The number of DO11 T cells in the draining lymph nodes is shown in A. The numbers adjacent to the upper right corner of each panel are the percentage of the total gated cells that are CD4+KJ126+ cells. P-selectin binding by CD4+KJ126+ or CD4+KJ126− populations is shown in B.
Discussion
In this study, we examined recruitment of different subsets of T lymphocytes into a peripheral inflammatory site. Using the adoptive transfer approach, we were able to follow specific populations of cells with distinct phenotypic characteristics. The peritoneal inflammatory model was chosen because recruitment of T cells can be easily quantified by flow cytometric analysis of cells harvested from the cavity, the recruitment process requires transmigration across an endothelial barrier, and different inflammatory stimuli can be administered by intraperitoneal injections. The injection of IFA into the peritoneum induces a mixed inflammatory infiltrate that changes over time, but includes activated neutrophils and lymphocytes at day 3 (data not shown).
Although there are many studies supporting the hypothesis that effector and naive T cells have different migratory patterns in vivo (3, 4, 51), there is little data directly comparing the abilities of effector and naive T cells to enter T peripheral inflammatory sites. The adoptive transfer of TCR transgenic T cells is a powerful method with which to address homing patterns of T cells. It has been used extensively to study homing of antigen specific T cells to secondary lymphoid tissues (52), but this technique has been applied only in a limited way to the study of effector T cell homing to inflammatory sites (35, 53). Our finding that naive DO11 T cells fail to enter the IFA-treated peritoneum, whereas activated effector DO11 T cells do, provides important direct evidence for the concept that effector T cells preferentially home to peripheral inflammatory sites but naive T cells do not.
We have also found that Th1 cells are more efficiently recruited to the inflamed peritoneum than are Th2 cells. This is consistent with previous studies indicating that Th1 cells enter dermal delayed-type hypersensitivity sites to a greater degree than Th2 cells (16). In our experiments, both Th1 and Th2 cells migrated to spleen and lymph node in equivalent numbers. This indicates that Th2 cells were as efficiently transferred into the circulation as Th1 cells, and that there is no intrinsic defect in the ability of in vitro– generated Th2 populations to transmigrate out of blood vessels into tissues. Furthermore, others have shown that Th2 cells can also migrate into lung tissue in mouse models of allergic airway inflammation (16, 54). On the other hand, in a murine model of autoimmune gastritis, activated Th2 cells are found in draining lymph nodes but not gastric mucosa, whereas Th1 cells are abundant in the mucosa (55). Thus, Th2 cells appear to be capable of entering only a subset of inflammatory sites. Differential recruitment of Th1 but not Th2 cells into inflammatory sites has been attributed to the expression of higher levels of functional selectin ligands on Th1 cells than Th2 cells (16–18, 20, 56). Consistent with those studies, we have found that the adoptively transferred Th1 cells express more P-selectin ligand than do the Th2 cells, and that anti–P- plus anti–E-selectin antibodies diminish Th1 recruitment into the peritoneum (Fig. 6 B). Unlike previous reports, we have examined the effect of the presence of antigen on the relative accumulation of antigen-specific Th1 and Th2 cells at the site. We have found that the difference between Th1 and Th2 cell accumulation seen without antigen is amplified when antigen is present (see Fig. 4 and Table I). The tracker-dye data indicate that the increased number of Th1 cells in the peritoneum in the presence of antigen is at least partially attributable to local proliferation. Thus the combination of preferential recruitment and then antigen-induced expansion of recruited cells appears to markedly polarize the phenotype of the helper T cell responses to peripheral antigen.
It is not clear if there are adhesion molecules that mediate selective recruitment of Th2 cells into inflammatory sites, such as in the asthmatic lung. A possible candidate is very late activation antigen (VLA)-4 on T cells interacting with vascular cell adhesion molecule (VCAM)-1 on inflamed endothelium. We have shown previously that an active form of VLA-4 that is capable of binding to VCAM-1 under physiological shear stress is expressed on memory but not naive human helper T cells (13). In addition, the Th2 cytokine, IL-4, can induce VCAM-1 expression on endothelial cells (57–59).
The expression of functional P-selectin ligand on CD4+ (13, 47, 56, 60) and CD8+ (61) T cells has been described. An important finding in this study is that the conditions used to drive CD4+ Th1 differentiation in vitro, namely antigen, IL-12, and anti–IL-4, also induce functional P-selectin ligand expression on CD8+ cells, whereas Th2 differentiation conditions, namely antigen, IL-4, and anti–IFN-γ generate CD8+ T cells that do not bind P-selectin. Furthermore, P-selectin ligand expression on CD8+ T cells correlates with their ability to home to the inflamed peritoneum. It is not clear whether the effector functions of these two CD8+ populations are different or not, although Tc1 and Tc2 subsets of CD8+ T cells producing different cytokines have been described (31, 34, 62). Recent evidence does suggest that adoptively transferred influenza-specific Tc1 cells are better able to clear pulmonary influenza infection then are Tc2 cells, even though both populations have comparable cytolytic activity (35). This may reflect different recruitment kinetics between the two populations or the effects of the different cytokines they produce. Our data suggest that the microenvironmental conditions in which naive CD8+ T cells are differentiated into CTLs are likely to influence not only the effector functions of these cells, but also their selectin-dependent homing patterns to peripheral inflammatory sites, just as with CD4+ helper T cells.
The data showing induction of P-selectin ligand on lymph node T cells in vivo indicate one mechanism by which recruitment of effector T cells at inflammatory sites can be biased toward antigen-specific cells. We have shown that functional P-selectin ligand is induced on antigen-specific naive T cells in lymph nodes draining a site of exposure to the antigen, but not on bystander T cells that cannot recognize the antigen but are otherwise exposed to the same local cytokine milieu. Induction of P-selectin ligand on lymph node T cells has been reported recently, but that study did not use TCR-transgenic T cells and the antigen specificity of the ligand induction could not be addressed (1). The in vivo activated CD4+ DO11 T cells described in this paper appear to be Th1-type as they produce abundant IFN-γ but no IL-4 upon restimulation with OVA peptide. Because survival and propagation of naive TCR transgenic T cells in vitro requires antigen stimulation, it has not been possible to distinguish the necessity of T cell receptor signals from cytokine signals for the induction of functional selectin ligands. In the in vivo study reported here, viable T cells that cannot bind the administered antigen fail to acquire the ability to bind P-selectin, even though the microenvironment includes the necessary cytokines.
Although the acquisition of a selectin binding phenotype appears to distinguish effector T cell subsets and determines their capability of homing to peripheral inflammatory sites, the induction of other functional adhesion molecules on T cells during effector differentiation are also likely to be important in peripheral homing. For example, CD44–dependent hyaluronate binding increases specifically on Vβ8+ lymph node T cells in SEB-treated mice, and these hyaluronate-binding T cells are selectively recruited to SEB-containing peritoneum in a CD44-dependent manner (63). It is possible that CD44 or other adhesion molecules may also contribute selective recruitment of effector T cell subsets.
In summary, the results presented here support the following model. When naive T cells encounter antigen in the lymph node, they become activated, undergo clonal expansion, and they differentiate into one or another subset of effector cells depending on the local cytokine milieu. During this process of activation and differentiation, specific adhesion molecules, including functional selectin ligands, and chemokine receptors are induced on the surface of activated T cells, in subset-specific patterns. Therefore, after entering circulation, the T cells are capable of migrating to particular peripheral sites where appropriate adhesion molecules and chemokines (ligands to the receptors on the T cells) are expressed or displayed by endothelium. The requirement that both antigen and cytokine signals be provided in the draining lymph nodes in order to stimulate the expression of functional selectin ligands may contribute to relative enrichment antigen–specific T cells at peripheral inflammatory sites. Maintenance of a functionally polarized T cell response at a peripheral site appears to require subset-specific recruitment, but the polarization is amplified by antigen-driven proliferation of the recruited T cells.
Acknowledgments
The authors thank Dr. Cheryl London for technical help in establishing the adoptive transfer model.
This work was supported by National Institutes of Health grants HL53993 (to F.W. Luscinskas) and HL36028 (to A.H. Lichtman). H. Xie is supported by an Arthritis Foundation postdoctoral fellowship. Y.-C. Lim is supported by American Heart Association postdoctoral fellowship.
Abbreviations used in this paper
- CMFDA
5-chloromethylfluorescein diacetate
- CTL
cytolytic T lymphocyte
- SEB
Staphylococcus enterotoxin B
References
- 1.Tietz W, Allemand Y, Borges E, von Laer D, Hallmann R, Vestweber D, Hamann A. CD4+T cells migrate into inflamed skin only if they express ligands for E- and P-selectin. J Immunol. 1998;161:963–970. [PubMed] [Google Scholar]
- 2.Bradley LM, Watson SR. Lymphocyte migration into tissue: the paradigm derived from CD4 subsets. Curr Opin Immunol. 1996;8:312–320. doi: 10.1016/s0952-7915(96)80118-x. [DOI] [PubMed] [Google Scholar]
- 3.Mackay CR, Marston W, Dudler L. Altered patterns of T cell migration through lymph nodes and skin following antigen challenge. Eur J Immunol. 1992;22:2205–2210. doi: 10.1002/eji.1830220904. [DOI] [PubMed] [Google Scholar]
- 4.Mackay CR, Marston WL, Dudler L. Naive and memory T cells show distinct pathways of lymphocyte recirculation. J Exp Med. 1990;171:801–817. doi: 10.1084/jem.171.3.801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Swain SL, Bradley LM. Helper T cell memory: more questions than answers. Semin Immunol. 1992;4:59–68. [PubMed] [Google Scholar]
- 6.Pietschmann P, Cush JJ, Lipsky PE, Oppenheimer-Marks N. Identification of subsets of human T cells capable of enhanced transendothelial migration. J Immunol. 1992;149:1170–1178. [PubMed] [Google Scholar]
- 7.Brezinschek RI, Lipsky PE, Galea P, Vita R, Oppenheimer-Marks N. Phenotypic characterization of CD4+T cells that exhibit a transendothelial migratory capacity. J Immunol. 1995;154:3062–3077. [PubMed] [Google Scholar]
- 8.Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multiple paradigm. Cell. 1994;76:301–314. doi: 10.1016/0092-8674(94)90337-9. [DOI] [PubMed] [Google Scholar]
- 9.Berg EL, Robinson MK, Warnock RA, Butcher EC. The human peripheral lymph node vascular addressin is a ligand for LECAM-1, the peripheral lymph node homing receptor. J Cell Biol. 1991;114:343–349. doi: 10.1083/jcb.114.2.343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hemmerich S, Butcher EC, Rosen SD. Sulfation-dependent recognition of high endothelial venules (HEV)-ligands by L-selectin and MECA 79, and adhesion-blocking monoclonal antibody. J Exp Med. 1994;180:2219–2226. doi: 10.1084/jem.180.6.2219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Girard JP, Springer TA. High endothelial venules (HEVs): specialized endothelium for lymphocyte migration. Immunol Today. 1995;16:449–457. doi: 10.1016/0167-5699(95)80023-9. [DOI] [PubMed] [Google Scholar]
- 12.Luster AD. Chemokines — chemotactic cytokines that mediate inflammation. N Engl J Med. 1998;338:436–445. doi: 10.1056/NEJM199802123380706. [DOI] [PubMed] [Google Scholar]
- 13.Lichtman AH, Ding H, Henault L, Vachino G, Camphausen R, Cumming D, Luscinskas FW. CD45 RA−RO+ (memory) but not CD45RA+RO−(naive) T cells roll efficiently on E- and P-selectin and vascular cell adhesion molecule-1 under flow. J Immunol. 1997;158:3640–3650. [PubMed] [Google Scholar]
- 14.Abbas AK, Williams ME, Burstein HJ, Chang TL, Bossu P, Lichtman AH. Activation and functions of CD4+T-cell subsets. Immunol Rev. 1991;123:5–22. doi: 10.1111/j.1600-065x.1991.tb00603.x. [DOI] [PubMed] [Google Scholar]
- 15.Hsieh CS, Macatonia SE, Tripp CS, Wolf SF, O'Garra A, Murphy KM. Development of TH1 CD4+ T cells through IL-12 produced by Listeria-induced macrophages. Science. 1993;260:547–549. doi: 10.1126/science.8097338. [DOI] [PubMed] [Google Scholar]
- 16.Austrup F, Vestweber D, Borges E, Lohning M, Brauer R, Herz U, Renz H, Hallmann R, Scheffold A, Radbruch A, Hamann A. P- and E-selectin mediate recruitment of T-helper-1 but not T-helper-2 cells into inflamed tissues. Nature. 1997;385:81–83. doi: 10.1038/385081a0. [DOI] [PubMed] [Google Scholar]
- 17.Borges E, Tietz W, Steegmaier M, Moll T, Hallmann R, Hamann A, Vestweber D. P-selectin glycoprotein ligand-1 (PSGL-1) on T helper 1 but not on T helper 2 cells binds to P-selectin and supports migration into inflamed skin. J Exp Med. 1997;185:573–578. doi: 10.1084/jem.185.3.573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lim YC, Henault L, Wagers AJ, Kansas GS, Luscinskas FW, Lichtman AH. Expression of functional selectin ligands on helper T cells is differentially regulated by interleukin-12 and interleukin-4. J Immunol. 1999;162:3193–3201. [PubMed] [Google Scholar]
- 19.Wagers AJ, Waters CM, Stoolman LM, Kansas GS. Interleukin 12 and interleukin 4 control T cell adhesion to endothelial selectins through opposite effects on alpha1, 3-fucosyltransferase VII gene expression. J Exp Med. 1998;188:2225–2231. doi: 10.1084/jem.188.12.2225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.van Wely CA, Blanchard AD, Britten CJ. Differential expression of alpha3 fucosyltransferases in Th1 and Th2 cells correlates with their ability to bind P-selectin. Biochem Biophys Res Commun. 1998;247:307–311. doi: 10.1006/bbrc.1998.8786. [DOI] [PubMed] [Google Scholar]
- 21.Ward SG, Bacon K, Westwick J. Chemokines and T lymphocytes: more than an attraction. Immunity. 1998;9:1–11. doi: 10.1016/s1074-7613(00)80583-x. [DOI] [PubMed] [Google Scholar]
- 22.Schall TJ, Bacon KB. Chemokines, leukocyte trafficking, and inflammation. Curr Opin Immunol. 1994;6:865–873. doi: 10.1016/0952-7915(94)90006-x. [DOI] [PubMed] [Google Scholar]
- 23.Luster AD, Unkeless JC, Ravetch JV. Gamma-interferon transcriptionally regulates an early-response gene containing homology to platelet proteins. Nature. 1985;315:672–676. doi: 10.1038/315672a0. [DOI] [PubMed] [Google Scholar]
- 24.Farber JM. Mig and IP-10: CXC chemokines that target lymphocytes. J Leukocyte Biol. 1997;61:246–257. [PubMed] [Google Scholar]
- 25.Piali L, Weber C, LaRosa G, Mackay CR, Springer TA, Clark-Lewis I, Moser B. The chemokine receptor CXCR3 mediates rapid and shear-resistant adhesion- induction of effector T lymphocytes by the chemokines IP10 and Mig. Eur J Immunol. 1998;28:961–972. doi: 10.1002/(SICI)1521-4141(199803)28:03<961::AID-IMMU961>3.0.CO;2-4. [DOI] [PubMed] [Google Scholar]
- 26.Bonecchi R, Bianchi G, Bordignon PP, D'Ambrosio D, Lang R, Borsatti A, Sozzani S, Allavena P, Gray PA, Mantovani A, Sinigaglia F. Differential expression of chemokine receptors and chemotactic responsiveness of type 1 T helper cells (Th1s) and Th2s. J Exp Med. 1998;187:129–134. doi: 10.1084/jem.187.1.129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sallusto F, Lenig D, Mackay CR, Lanzavecchia A. Flexible programs of chemokine receptor expression on human polarized T helper 1 and 2 lymphocytes. J Exp Med. 1998;187:875–883. doi: 10.1084/jem.187.6.875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Qin S, Rottman JB, Myers P, Kassam N, Weinblatt M, Loetscher M, Koch AE, Moser B, Mackay CR. The chemokine receptors CXCR3 and CCR5 mark subsets of T cells associated with certain inflammatory reactions. J Clin Invest. 1998;101:746–754. doi: 10.1172/JCI1422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Vanguri P, Farber JM. Identification of CRG-2. An interferon-inducible mRNA predicted to encode a murine monokine. J Biol Chem. 1990;265:15049–15057. [PubMed] [Google Scholar]
- 30.Mosmann TR, Sad S. The expanding universe of T-cell subsets: Th1, Th2 and more. Immunol Today. 1996;17:138–146. doi: 10.1016/0167-5699(96)80606-2. [DOI] [PubMed] [Google Scholar]
- 31.Sad S, Marcotte R, Mosmann TR. Cytokine-induced differentiation of precursor mouse CD8+ T cells into cytotoxic CD8+T cells secreting Th1 or Th2 cytokines. Immunity. 1995;2:271–279. doi: 10.1016/1074-7613(95)90051-9. [DOI] [PubMed] [Google Scholar]
- 32.Mosmann TR, Sad S, Krishnan L, Wegmann TG, Guilbert LJ, Belosevic M. Differentiation of subsets of CD4+ and CD8+T cells. Ciba Found Symp. 1995;195:42–50. doi: 10.1002/9780470514849.ch4. [DOI] [PubMed] [Google Scholar]
- 33.Mosmann TR, Li L, Sad S. Functions of CD8 T-cell subsets secreting different cytokine patterns. Semin Immunol. 1997;9:87–92. doi: 10.1006/smim.1997.0065. [DOI] [PubMed] [Google Scholar]
- 34.Cerwenka A, Carter LL, Reome JB, Swain SL, Dutton RW. In vivo persistence of CD8 polarized T cell subsets producing type 1 or type 2 cytokines. J Immunol. 1998;161:97–105. [PubMed] [Google Scholar]
- 35.Cerwenka A, Morgan TM, Harmsen AG, Dutton RW. Migration kinetics and final destination of type 1 and type 2 CD8 effector cells predict protection against pulmonary virus infection. J Exp Med. 1999;189:423–434. doi: 10.1084/jem.189.2.423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Murphy KM, Heimberger AB, Loh DY. Induction by antigen of intrathymic apoptosis of CD4+CD8+ TCRlothymocytes in vivo. Science. 1990;250:1720–1723. doi: 10.1126/science.2125367. [DOI] [PubMed] [Google Scholar]
- 37.London CA, Perez VL, Abbas AK. Functional characteristics and survival requirements of memory CD4+T lymphocytes in vivo. J Immunol. 1999;162:766–773. [PubMed] [Google Scholar]
- 38.Sako D, Chang XJ, Barone KM, Vachino G, White HM, Shaw G, Veldman GM, Bean KM, Ahern TJ, Furie B, et al. Expression cloning of a functional glycoprotein ligand for P-selectin. Cell. 1993;75:1179–1186. doi: 10.1016/0092-8674(93)90327-m. [DOI] [PubMed] [Google Scholar]
- 39.Sako D, Comess KM, Barone KM, Camphausen RT, Cumming DA, Shaw GD. A sulfated peptide segment at the amino terminus of PSGL-1 is critical for P-selectin binding. Cell. 1995;83:323–331. doi: 10.1016/0092-8674(95)90173-6. [DOI] [PubMed] [Google Scholar]
- 40.Haskins K, Kubo R, White J, Pigeon M, Kappler J, Marrack P. The major histocompatibility complex– restricted antigen receptor on T cells. I. Isolation with a monoclonal antibody. J Exp Med. 1983;157:1149–1169. doi: 10.1084/jem.157.4.1149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Marrack P, Shimonkevitz R, Hannum C, Haskins K, Kappler J. The major histocompatibility complex– restricted antigen receptor on T cells. IV. An antiidiotypic antibody predicts both antigen and I-specificity. J Exp Med. 1983;158:1635–1646. doi: 10.1084/jem.158.5.1635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Van Parijs L, Peterson DA, Abbas AK. The Fas/Fas ligand pathway and Bcl-2 regulate T cell responses to model self and foreign antigens. Immunity. 1998;8:265–274. doi: 10.1016/s1074-7613(00)80478-1. [DOI] [PubMed] [Google Scholar]
- 43.Kurts C, Kosaka H, Carbone FR, Miller JF, Heath WR. Class I–restricted cross-presentation of exogenous self-antigens leads to deletion of autoreactive CD8+T cells. J Exp Med. 1997;186:239–245. doi: 10.1084/jem.186.2.239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bird JJ, Brown DR, Mullen AC, Moskowitz NH, Mahowald MA, Sider JR, Gajewski TF, Wang CR, Reiner SL. Helper T cell differentiation is controlled by the cell cycle. Immunity. 1998;9:229–237. doi: 10.1016/s1074-7613(00)80605-6. [DOI] [PubMed] [Google Scholar]
- 45.Swain SL. Generation and in vivo persistence of polarized Th1 and Th2 memory cells. Immunity. 1994;1:543–552. doi: 10.1016/1074-7613(94)90044-2. [DOI] [PubMed] [Google Scholar]
- 46.Luscinskas FW, Ding H, Lichtman AH. P-selectin and vascular cell adhesion molecule 1 mediate rolling and arrest, respectively, of CD4+T lymphocytes on tumor necrosis factor α–activated vascular endothelium under flow. J Exp Med. 1995;181:1179–1186. doi: 10.1084/jem.181.3.1179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Vachino G, Chang XJ, Veldman GM, Kumar R, Sako D, Fouser LA, Berndt MC, Cumming DA. P-selectin glycoprotein ligand-1 is the major counter-receptor for P-selectin on stimulated T cells and is widely distributed in non-functional form on many lymphocytic cells. J Biol Chem. 1995;270:21966–21974. doi: 10.1074/jbc.270.37.21966. [DOI] [PubMed] [Google Scholar]
- 48.Carr MW, Alon R, Springer TA. The C-C chemokine MCP-1 differentially modulates the avidity of beta 1 and beta 2 integrins on T lymphocytes. Immunity. 1996;4:179–187. doi: 10.1016/s1074-7613(00)80682-2. [DOI] [PubMed] [Google Scholar]
- 49.Lloyd AR, Oppenheim JJ, Kelvin DJ, Taub DD. Chemokines regulate T cell adherence to recombinant adhesion molecules and extracellular matrix proteins. J Immunol. 1996;156:932–938. [PubMed] [Google Scholar]
- 50.Weber C, Alon R, Moser B, Springer TA. Sequential regulation of alpha 4 beta 1 and alpha 5 beta 1 integrin avidity by CC chemokines in monocytes: implications for transendothelial chemotaxis. J Cell Biol. 1996;134:1063–1073. doi: 10.1083/jcb.134.4.1063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mackay CR, Andrew DP, Briskin M, Ringler DJ, Butcher EC. Phenotype, and migration properties of three major subsets of tissue homing T cells in sheep. Eur J Immunol. 1996;26:2433. doi: 10.1002/eji.1830261025. [DOI] [PubMed] [Google Scholar]
- 52.Pape KA, Kearney ER, Khoruts A, Mondino A, Merica R, Chen ZM, Ingulli E, White J, Johnson JG, Jenkins MK. Use of adoptive transfer of T-cell-antigen-receptor-transgenic T cell for the study of T-cell activation in vivo. Immunol Rev. 1997;156:67–78. doi: 10.1111/j.1600-065x.1997.tb00959.x. [DOI] [PubMed] [Google Scholar]
- 53.Chen ZM, Jenkins MK. Revealing the in vivo behavior of CD4+ T cells specific for an antigen expressed in Escherichia coli. . J Immunol. 1998;160:3462–3470. [PubMed] [Google Scholar]
- 54.Randolph DA, Carruthers CJ, Szabo SJ, Murphy KM, Chaplin DD. Modulation of airway inflammation by passive transfer of allergen-specific Th1 and Th2 cells in a mouse model of asthma. J Immunol. 1999;162:2375–2383. [PubMed] [Google Scholar]
- 55.Katakai T, Mori KJ, Masuda T, Shimizu A. Differential localization of Th1 and Th2 cells in autoimmune gastritis. Int Immunol. 1998;10:1325–1334. doi: 10.1093/intimm/10.9.1325. [DOI] [PubMed] [Google Scholar]
- 56.Wagers AJ, Stoolman LM, Kannagi R, Craig R, Kansas GS. Expression of leukocyte fucosyltransferases regulates binding to E-selectin: relationship to previously implicated carbohydrate epitopes. J Immunol. 1997;159:1917–1929. [PubMed] [Google Scholar]
- 57.Briscoe DM, Cotran RS, Pober JS. Effects of tumor necrosis factor, lipopolysaccharide, and IL-4 on the expression of vascular cell adhesion molecule-1 in vivo. Correlation with CD3+T cell infiltration. J Immunol. 1992;149:2954–2960. [PubMed] [Google Scholar]
- 58.Luscinskas FW, Kansas GS, Ding H, Pizcueta P, Schleiffenbaum BE, Tedder TF, Gimbrone MA., Jr Monocyte rolling, arrest and spreading on IL-4–activated vascular endothelium under flow is mediated via sequential action of L-selectin, beta 1-integrins, and beta 2-integrins. J Cell Biol. 1994;125:1417–1427. doi: 10.1083/jcb.125.6.1417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Schleimer RP, Sterbinsky SA, Kaiser J, Bickel CA, Klunk DA, Tomioka K, Newman W, Luscinskas FW, Gimbrone MA, Jr, McIntyre BW, et al. IL-4 induces adherence of human eosinophils and basophils but not neutrophils to endothelium. Association with expression of VCAM-1. J Immunol. 1992;148:1086–1092. [PubMed] [Google Scholar]
- 60.Alon R, Rossiter H, Wang X, Springer TA, Kupper TS. Distinct cell surface ligands mediate T lymphocyte attachment and rolling on P and E selectin under physiological flow. J Cell Biol. 1994;127:1485–1495. doi: 10.1083/jcb.127.5.1485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Knibbs RN, Craig RA, Maly P, Smith PL, Wolber FM, Faulkner NE, Lowe JB, Stoolman LM. Alpha(1,3)-fucosyltransferase VII-dependent synthesis of P- and E- selectin ligands on cultured T lymphoblasts. J Immunol. 1998;161:6305–6315. [PubMed] [Google Scholar]
- 62.Li L, Sad S, Kagi D, Mosmann TR. CD8Tc1 and Tc2 cells secrete distinct cytokine patterns in vitro and in vivo but induce similar inflammatory reactions. J Immunol. 1997;158:4152–4161. [PubMed] [Google Scholar]
- 63.DeGrendele HC, Estess P, Siegelman MH. Requirement for CD44 in activated T cell extravasation into an inflammatory site. Science. 1997;278:672–675. doi: 10.1126/science.278.5338.672. [DOI] [PubMed] [Google Scholar]