

Abstract
Protein kinase Cs (PKCs) are activated by antigen receptors in lymphocytes, but little is known about proximal targets for PKCs in antigen receptor–mediated responses. In this report, we define a role for diacylglycerol-regulated PKC isoforms in controlling the activity of the serine/threonine kinase protein kinase D (PKD; also known as PKCμ) in T cells, B cells, and mast cells. Antigen receptor activation of PKD is a rapid and sustained response that can be seen in T cells activated via the T cell antigen receptor, B cells activated via the B cell antigen receptor, and in mast cells triggered via the high-affinity receptor for IgE (FcεR1). Herein, we show that antigen receptor activation of PKD requires the activity of classical/novel PKCs. Moreover, PKC activity is sufficient to bypass the requirement for antigen receptor signals in the induction of PKD activity. These biochemical and genetic studies establish a role for antigen receptor–regulated PKC enzymes in the control of PKD activity. Regulation of PKD activity through upstream PKCs reveals a signaling network that exists between different members of the PKC superfamily of kinases that can operate to amplify and disseminate antigen receptor signals generated at the plasma membrane.
Keywords: antigen receptor, diacylglycerol, protein kinase C, protein kinase D
Introduction
Antigen receptors control a network of signaling events that regulate lymphocyte function 1 2 3 4. Triggering of antigen receptors activates cytosolic tyrosine kinases that couple antigen receptors to a variety of different signaling molecules, including phospholipase Cγ1 (PLC) γ1. The activation of PLCγ1 results in the hydrolysis of phosphatidylinositol 4 5P2, producing inositol polyphosphates and diacylglycerol (DAG), which regulate intracellular calcium levels and activate the protein kinase C (PKC) family of serine/threonine kinases, respectively. Activation of PKCs is critical for leukocyte function. Targets for PKCs in lymphocytes include transcription factors such as nuclear factor κB 5, which are important for the regulation of cytokine gene expression. There is also evidence that PKCs control integrin function, with obvious implications for the regulation of lymphocyte adhesion and cell–cell contact 6 7. The potency of PKC signaling pathways for lymphocyte activation is underlined by the observation that phorbol esters, pharmacological agents that activate PKC, can mimic many aspects of antigen receptor triggering 8 9 10.
There are multiple related PKC isoforms, which are classified into three distinct groups: classical PKCs (α, βI, βII, and γ), which are regulated by calcium, DAG, and phospholipids; novel PKCs (δ, ε, η, and θ), which are regulated by DAG and phospholipids; and the atypical PKCs (ζ and λ), which are insensitive to both calcium and DAG 11 12 13. Mice lacking the classical PKC isoforms PKCβI/II exhibit B lymphocyte dysfunction and have impaired humoral responses to T cell–independent antigens 14. Novel PKCs have also been implicated in antigen receptor function in T cells on the basis that PKCθ localizes to the plasma membrane contact zone formed between T cells and APCs during antigen stimulation 15 16. Nevertheless, despite these intriguing localization data, there is little known about the biochemistry of PKC signaling pathways in lymphocytes. The present challenge is to identify targets for these enzymes that transmit signals to the cell interior and control the complex genetic processes that ultimately determine lymphocyte function.
A serine kinase protein kinase D (PKD) has been found to be highly expressed in hematopoietic cells 17 18. Several important functions have been reported for this kinase, including the regulation of nuclear factor κB transcription factors and in the control of Golgi complex organization and protein transport processes 19 20 21. PKD is distantly related to the PKC family through the presence of a conserved DAG-binding cysteine-rich domain (C1 domain) within its regulatory domain 17 22 23. However, the catalytic domain of PKD shows very low homology to the conserved kinase subdomains of the PKCs 17 and displays a unique substrate specificity 17 23. PKD is highly expressed in both T and B lymphocytes, and antigen receptor engagement rapidly stimulates PKD activity 24 25. The mechanism for activation of PKD in lymphocytes is not known. The C1 domain of PKD can bind DAG, and DAG production is essential for B cell antigen receptor (BCR) coupling to PKD 25. The C1 domain of PKD has an essential role in localizing PKD to the plasma membrane in phorbol etser–activated cells 26. Nevertheless, it is not clear whether DAG binding directly activates PKD in lymphocytes. In this context, studies in fibroblast cell lines have noted that DAG and phorbol esters do not directly activate PKD in intact cells 27 28. Instead, they activate classical and novel PKC enzymes, which subsequently act to regulate the catalytic activity of PKD by inducing the phosphorylation of two critical serine residues within the catalytic domain of PKD 29 30. Whether the regulatory mechanisms that control PKD activity in lymphocytes are the same as those operating in fibroblasts and epithelial cells is unknown. The reason for questioning a universal model for PKD activation is that PKD exhibits distinct patterns of subcellular localization in different cell types. In epithelial cells, PKD is localized within the Golgi compartment 20 31. In contrast, PKD is found localized to the cytosol of lymphocytes, not the Golgi, raising the possibility that the regulatory mechanisms that control the activity of this kinase may be different.
The object of this report was to use genetic and biochemical analysis to examine the regulation of PKD in lymphocytes, and in particular to study the mechanism of antigen receptor coupling to PKD. These studies identify a function for classical/novel PKC enzymes in lymphocytes as signaling intermediates that couple antigen receptors to the serine/threonine kinase, PKD.
Materials and Methods
cDNA Constructs.
Constitutively active PKCε and η mutants (containing pseudosubstate motif deletions) were provided by P. Parker (Protein Phosphorylation Laboratory, Imperial Cancer Research Fund, London, UK). cDNA constructs containing wild-type and P287G mutant PKD sequences in the pcDNA3 mammalian expression vector have been described previously 23 32. Chimeric fusion proteins between green fluorescent protein (GFP) and different PKD mutants were generated by subcloning the PKD constructs into the EcoR1 site of a pEF-plink2-GFPC3 expression vector. All constructs were purified by CsCl density gradient centrifugation before use in transient transfection experiments.
Cell Culture and Transient Transfection.
The murine BALB/c mouse B lymphoma A20 cell line was maintained in RPMI 1640 medium supplemented with 10% fetal bovine serum and 50 μM β-mercaptoethanol. The rat basophilic leukemia (RBL) cell line 2H3 was maintained as described previously 33. Human peripheral blood–derived T lymphoblasts were generated and maintained as described previously 34. The Jurkat T lymphocyte cell line JH6.2 was maintained in RPMI 1640 medium supplemented with 10% fetal bovine serum. For transient expression of cDNA constructs, 1.5 × 107 A20 B cells or RBL 2H3 cells were electroporated with 20 μg of DNA at 310 V and 960 μF, resuspended in 5 ml of complete medium, and cultured overnight before experimental usage.
Cell Lysis and Western Blot Analysis.
After stimulation, cells were lysed for 20 min at 4°C in a buffer containing 50 mM Tris/HCl, pH 7.4; 2 mM EGTA; 2 mM EDTA; 1 mM dithiothreitol; 10 μg/ml aprotinin; 10 μg/ml leupeptin; 1 mM 4-(2′-aminoeth-yl)-benzenesulfonyl fluoride hydrochloride; and 1% Triton X-100. Lysates were clarified by centrifugation at 14,000 rpm for 10 min at 4°C, and proteins in the supernatant were precipitated with ice-cold acetone and resuspended in ×2 SDS-PAGE sample buffer. Samples were resolved under reducing conditions by 8% SDS-PAGE and transferred to polyvinylidene difluoride membranes (Millipore). Western blot analysis was performed, and immunoreactive bands were visualized by enhanced chemiluminescence.
In Vitro Kinase Assays.
Endogenous PKD was immunoprecipitated from cell lysates at 4°C for 2 h with the PA-1 antiserum (1:100 dilution) as described previously 23 and recovered with protein A–agarose beads. Immunocomplexes were washed twice in lysis buffer and once in kinase buffer (30 mM Tris/HCl, pH 7.4; 10 mM MgCl2), and PKD activity (as measured by autophosphorylation) was determined by incubating immunocomplexes with 20 μl of kinase buffer containing 100 μM [γ-32P]ATP final concentration at 30°C for 10 min. Reactions were terminated by the addition of ×2 SDS-PAGE sample buffer, and were analyzed by SDS-PAGE and autoradiography.
Materials.
Enhanced chemiluminescence reagents and [γ-32P]ATP (370 MBq/ml) were from Amersham Pharmacia Biotech. Phorbol dibutyrate (PDBu) was from Sigma-Aldrich. Protein A–agarose was from Boehringer. The PKC inhibitors Ro 31-8220 and GF 109203X were from LC Laboratories. A rabbit polyclonal PKD antibody directed against the COOH terminus of PKD used for Western blotting was from Santa Cruz Biotechnology, Inc. (sc-935). Phospho–extracellular signal–regulated kinase (Erk)-1/2 and pan–Erk-2 antibodies were from Promega and Transduction Laboratories, respectively. Recombinant human IL-2 was from Eurocetus. All other reagents were from standard suppliers, or as indicated in the text.
Results and Discussion
Activation of PKD in B Lymphocytes, Mast Cells, and T Lymphocytes after Antigen Receptor Ligation.
Previous results have shown that triggering of the BCR induces PKD catalytic activity 24 25. The BCR is one of a family of antigen receptors that includes the FcεR1 and the TCR. These receptors couple to different cytosolic tyrosine kinases and have the potential to couple to unique downstream signals. Therefore, in initial experiments, we examined whether PKD activation is common to the BCR, the FcεR1, and the TCR. PKD activity was monitored by quantitation of PKD autophosphorylation using in vitro kinase assays. Furthermore, we have recently shown that active PKD autophosphorylates on a COOH-terminal serine residue (S916), and that a specific antiserum recognizing PKD molecules phosphorylated on S916 can be used to efficiently monitor the catalytic activity of PKD in intact cells 24.
To study B cell responses, A20 B lymphoma cells were either left unstimulated or were activated by cross-linking the BCR with F(ab)′2 fragments of anti–mouse IgG for various amounts of time (0–10 min). Cross-linking of the BCR complex was found to induce a rapid ∼8-fold increase in PKD catalytic activity, as assessed by in vitro kinase assays that measure PKD autophosphorylation (Fig. 1 A). This response was maximal within 1 min of BCR triggering, and was sustained for >10 min. In addition to regulating the catalytic activity of PKD, ligation of the BCR complex was also found to induce autophosphorylation of the COOH-terminal S916 residue of PKD in a manner that paralleled the observed increase in PKD catalytic activity. Thus, Western blot analysis of lysates prepared from F(ab)′2-treated A20 B lymphoma cells showed a rapid, strong immunoreactivity of the pS916 antibody for active PKD present in these cells, but not with PKD isolated from nonstimulated cells (Fig. 1 A). PKD was also activated when B cells were treated with phorbol esters, pharmacological activators of PKC (Fig. 1 A).
Figure 1.


BCR or FcεR1 ligation induces PKD catalytic activity. (A) Left: A20 B lymphocytes were left untreated (n/s) or were stimulated with 10 μg/ml F(ab)′2 fragments of anti–mouse IgG for various amounts of time (0–10 min) to activate the BCR complex. Alternatively, the cells were stimulated with 50 ng/ml PDBu for 10 min. The cells were subsequently lysed before PKD was immunoprecipitated using the PA-1 antiserum, and in vitro kinase assays were performed. PKD activity was measured by autophosphorylation (IVK). In parallel, total proteins from the cell lysates were precipitated with cold acetone, separated by SDS-PAGE, and subjected to Western blot analysis using the pS916 antiserum and a pan–COOH-terminal PKD antibody. Results are representative of four independent experiments. Right: the mean ± SE (n = 4) increase in PKD activity over time after BCR ligation (as measured by autophosphorylation [IVK] or by immunoreactivity with the pS916 antiserum) is shown, expressed as the fold increase in PKD activity over basal levels in nonstimulated cells. (B) RBL 2H3 cells were primed with 1 μg/ml IgE anti-DNP for 1 h at 37°C, and then either antigenic cross-linking of the bound IgE was performed using 500 ng/ml KLH-DNP for various amounts of time (0–10 min), or the cells were left unstimulated (−). Alternatively, RBL 2H3 cells were stimulated with 50 ng/ml PDBu for 10 min. PKD was immunoprecipitated from whole cell lysates, and the activity of PKD was measured by in vitro kinase assays (IVK). SDS-PAGE and Western blot analysis of the cell lysates show equivalent amounts of PKD in all the samples. Results are representative of two independent experiments. α-, anti-.
The antigen receptor present in mast cells is the high-affinity receptor for IgE, the FcεR1. To address whether the FcεR1 is also coupled to PKD, RBL 2H3 mast cells were left unstimulated or treated with IgE plus cognate antigen, KLH-DNP, for various amounts of time. The data presented in Fig. 1 B indicate that ligation of the FcεR1 induces a rapid increase in PKD catalytic activity, as measured by in vitro kinase assays.
To examine the effects of triggering the TCR complex on the catalytic activity of PKD, quiescent human peripheral blood–derived T lymphoblasts were activated with the CD3ε cross-linking mAb UCHT1. As demonstrated in Fig. 2 A, PKD exhibited a low basal level of kinase activity in quiescent peripheral T lymphocytes that was rapidly enhanced after cross-linking of the TCR complex with the CD3ε antibody UCHT1. Here, maximal stimulation of PKD catalytic activity was observed within ∼1 min of stimulation, and was sustained for >10 min. The data in Fig. 2 B shows that activation of PKD by the TCR complex was also observed in the leukemic Jurkat T cell line, JH6.2. These cells were stimulated with the UCHT1 mAb, and PKD activity was measured by in vitro kinase assays. PKD exhibited negligible basal activity in nonstimulated JH6.2 cells, and stimulation of the TCR complex led to a rapid increase in PKD activity, similar to that observed in peripheral blood–derived T cells.
Figure 2.



TCR stimulation induces PKD catalytic activity. (A) Peripheral blood T lymphoblasts were left nonstimulated (n/s) or were treated with 10 μg/ml UCHT1 for various amounts of time (1–10 min) to activate the TCR complex. Alternatively, the cells were stimulated with 50 ng/ml PDBu for 10 min. Top: endogenous PKD was immunoprecipitated from whole cell lysates using the PA-1 antiserum, and in vitro kinase assays were performed. PKD activity was measured by autophosphorylation (IVK). Bottom: total proteins from whole cell lysates were precipitated with cold acetone, separated by SDS-PAGE, and subjected to Western blot analysis using a pan–COOH-terminal PKD antibody. (B) JH6.2 T lymphocytes were left nonstimulated (n/s) or were treated with 10 μg/ml UCHT1 for various amounts of time (1–10 min) to activate the TCR complex. Endogenous PKD was immunoprecipitated from whole cell lysates using the PA-1 antiserum, and in vitro kinase assays (IVK) were performed. PKD expression levels were determined by Western blotting of cell lysates using a pan–COOH-terminal PKD antibody. Data are representative of a least two independent experiments. (C) T lymphocytes were left nonstimulated (n/s) or were treated with 10 μg/ml UCHT1 or 50 ng/ml PDBu for 10 min. Total proteins from the cell lysates were precipitated with cold acetone, analyzed by SDS-PAGE, and subjected to Western blot analysis using the pS916 antiserum, which recognizes active PKD and a pan–COOH-terminal PKD antibody. α-, anti-.
IL-2 Does Not Activate PKD in T Lymphocytes.
Hematopoietic cytokines such as IL-2 control T cell proliferation and differentiation. Several TCR-activated serine kinases are also activated by the IL-2 receptor, including the mitogen-activated protein kinases 35 and the phosphatidylinositol 3-kinase–dependent serine kinase PKB/Akt 36. However, IL-2 activation of classical and novel PKCs has not been generally observed. The data presented in Fig. 3 demonstrate that IL-2 stimulation of peripheral blood–derived T lymphoblasts does not increase PKD catalytic activity over basal levels. In contrast, a phospho-Erk polyclonal antibody (which specifically recognizes activated Erk-1/2 in Western blot analysis of whole cell lysates) reacted strongly with active Erks present in both IL-2– and PDBu-treated cells (Fig. 3), confirming the biological activity of the IL-2. Similar results were obtained after Western blot analysis with the pS916 antisera: here, even prolonged stimulation of peripheral blood–derived T lymphoblasts with IL-2 for >1 h did not activate PKD (data not shown). These experiments indicate that activation of PKD in T cells is a selective response to antigen receptor triggering and is not a general response to mitogenic stimuli.
Figure 3.


IL-2 does not regulate PKD activity in T lymphocytes (A) Peripheral blood T lymphoblasts were left nonstimulated (–) or were treated with either 50 ng/ml PDBu for 10 min or with 20 ng/ml recombinant IL-2 (rIL-2) for 2 or 15 min, as indicated. Left: endogenous PKD was immunoprecipitated from whole cell lysates using the PA-1 antiserum, and in vitro kinase assays (IVK) were performed. Extracts from cell lysates were also analyzed by Western blotting with a pan–COOH-terminal PKD antibody. Right: lysates from the recombinant IL-2 (rIL-2)– or PDBu-treated T lymphoblasts were also analyzed by Western blotting with a phosphorylation state–specific Erk-1/2 antibody, and subsequently with a pan–Erk-2 antibody. Data are representative of three independent experiments. (B) Peripheral blood T lymphoblasts were left nonstimulated (−) or were treated with either 10 μg/ml UCHT1 or 20 ng/ml recombinant IL-2 (rIL-2) for the indicated amount of time. Total proteins from the cell lysates were precipitated with cold acetone, analyzed by SDS-PAGE, and subjected to Western blot analysis using the pS916 antiserum (which recognizes active PKD), a pan–COOH-terminal PKD antibody, a specific phosphoserine (Ser473) PKB antibody, and a pan-PKB antibody. a-, anti-.
PKD Activation by Antigen Receptors Is Not Regulated by Direct Binding of DAG in Intact Cells.
Previous work has shown that antigen receptor activation of PKD in B cells is dependent on PLCγ1 activation 25. The regulatory domain of PKD contains a C1 domain that is capable of binding DAG and that mediates translocation of the enzyme to the plasma membrane after antigen receptor engagement. In vitro, DAG and acidic phospholipids can activate PKD synergistically 18 23, but in intact lymphocytes the importance of DAG binding to the C1 domain of PKD for enzyme activation by antigen receptors is not known. The C1 domain in PKD contains two cysteine-rich motifs (C1A and C1B) that are not functionally equivalent: C1B acts as the major DAG binding site within PKD both in vitro and in vivo 32. A highly conserved proline residue is conserved in all DAG-binding C1 domains described to date and is essential for high-affinity binding of DAG or phorbol esters to these domains. Accordingly, a single amino acid substitution (P287G) within the C1B motif of PKD inhibits phorbol ester binding in vitro and in vivo 32. In this study, we used the PKD P287G mutant to explore the role of the C1 domain in antigen receptor–mediated PKD activation.
A20 B cells were transiently transfected with either wild-type PKD or the P287G mutant of PKD. The loss of in vivo DAG/phorbol ester binding to the PKD P2887G mutant is demonstrated by the inability of this mutant to translocate to the plasma membrane in response to antigen receptor stimulation or phorbol ester treatment. The data presented in Fig. 4 compare the activity of wild-type PKD and the PKD P287G mutant and demonstrate that PKD P287G, when transiently expressed in A20 B lymphocytes, displays identical kinetics and magnitude of activation as wild-type PKD after BCR ligation. Thus, the integrity of the C1 domain is not essential for antigen receptor–mediated activation of PKD in intact cells.
Figure 4.

Direct DAG binding is not essential for PKD activation in intact cells. A20 B lymphocytes were transiently transfected with 20 μg of GFP-tagged wild-type (WT) or P287G PKD cDNA. After 16 h incubation at 37°C, the cells were left nonstimulated (−) or were treated with 10 μg/ml F(ab)′2 fragments of anti–mouse IgG for the indicated amounts of times to activate the BCR complex. Ectopically expressed PKD was immunoprecipitated from cell lysates using a monoclonal GFP antibody (Imperial Cancer Research Fund), and PKD activity (autophosphorylation) was measured by in vitro kinase assays (IVK). In addition, expression levels of the GFP-PKD constructs were verified by Western blot analysis with a pan–COOH-terminal PKD antibody. Data are representative of two individual experiments. a-, anti-.
PKD Activation Is Regulated by a PKC Cascade in T Lymphocytes, B Lymphocytes, and Mast Cells.
The ability of the BCR to stimulate PKD mutants with defective C1/DAG binding domains must be reconciled with previous studies showing that DAG production is essential for PKD activation 25 37. Recent studies in fibroblasts have placed PKD downstream of classical/novel PKC enzymes in a hierarchical signaling cascade regulated by DAG (for reviews, see references 30, 38). To examine the role of PKCs in the regulation of PKD by the BCR, A20 cells were preincubated with Ro 31-8220, an inhibitor of classical/novel PKCs that has no direct inhibitory effect on PKD kinase activity 28 39. Subsequently, the BCR complex was activated, and PKD activity was measured by in vitro kinase assays. BCR-induced activation of PKD was severely impaired in B lymphocytes that had been pretreated with Ro 31-8220, compared with the marked increase in PKD activity observed in control BCR-stimulated cells (Fig. 5 A). BCR engagement poorly induced PKD S916 phosphorylation in B lymphocytes that had been pretreated with Ro 31-8220 (Fig. 5 B). Moreover, phorbol ester–induced PKD S916 phosphorylation was also blocked by the Ro 31-8220 inhibitor (Fig. 5 B).
Figure 5.
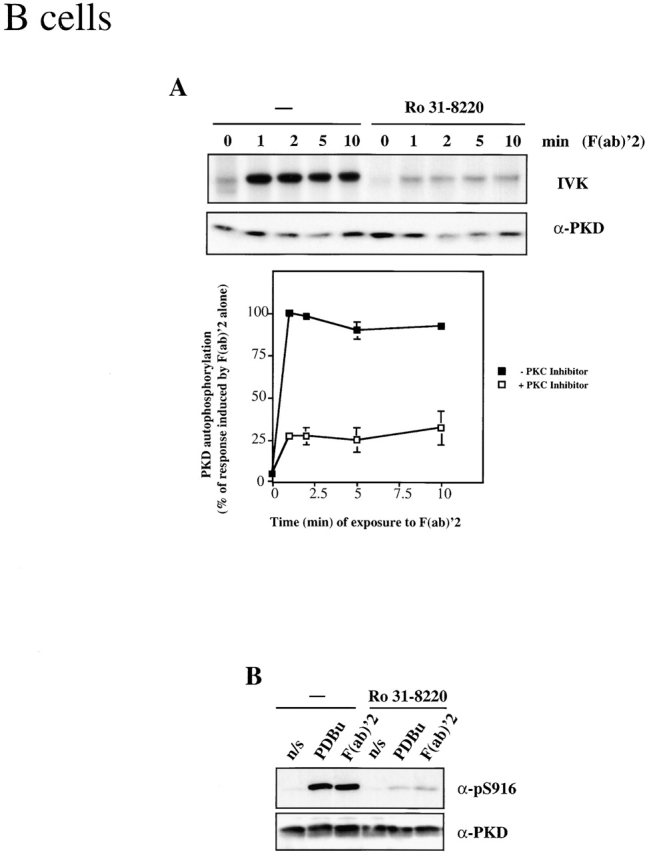
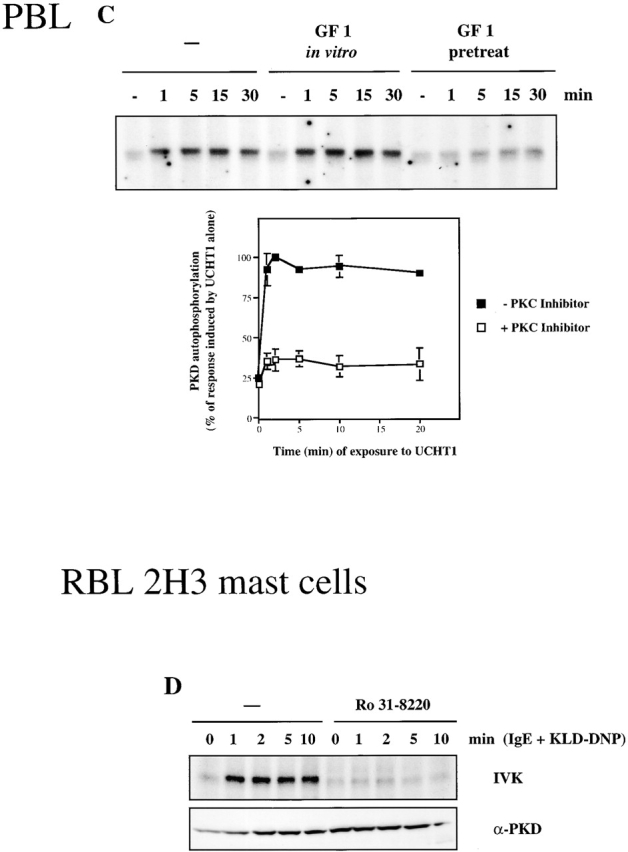
Inhibition of classical/novel PKCs prevents activation of PKD by the BCR, the TCR, and the FcεR1. (A) A20 B lymphocytes were pretreated with 2.5 μm Ro 31-8220 or an equal volume of solvent (−) for 1 h before activation of the BCR complex using 10 μg/ml F(ab)′2 fragments of anti–mouse IgG for the indicated amounts of time. PKD was immunoprecipitated from cell lysates, and PKD activity was measured by in vitro kinase assays (IVK) measuring PKD autophosphorylation. Data are expressed as the percentage of the maximal response induced by F(ab)′2 alone, and are the mean ± SE of three independent experiments. A representative autoradiogram is shown. (B) A20 B lymphocytes were pretreated with 2.5 μm Ro 31-8220 or an equal volume of solvent (−) for 1 h. The cells were subsequently left nonstimulated (n/s), or were treated with either 10 μg/ml F(ab)′2 fragments of anti–mouse IgG for 2 min or with 50 ng/ml PDBu for 10 min. Whole cell lysates were analyzed by Western blotting with the pS916 antiserum and a pan–COOH-terminal PKD antibody. Similar results were seen in two independent experiments. (C) Peripheral blood–derived T lymphoblasts were pretreated with 3.5 μM GF 109203X (GF 1 pretreat) or an equal volume of solvent (−) for 1 h. Cells were subsequently left unstimulated (−) or were treated with 10 μg/ml UCHT1 for various amounts of time, as indicated. PKD was immunoprecipitated from whole cell lysates, and PKD activity measured by in vitro kinase assays. Data are expressed as the percentage of the maximal response induced by UCHT1 alone and are the mean ± SE of three independent experiments. A representative autoradiogram is shown. In some cases, PKD immunoprecipitates were incubated in the presence of 3.5 μM GF 109203X during in vitro kinase assays (GF 1 in vitro). (D) RBL 2H3 mast cells were pretreated with 2.5 μm Ro 31-8220 or an equal volume of solvent (−) for 1 h before stimulation of the FcεR1, as described in the legend to Fig. 1. PKD activity was measured by in vitro kinase assays (IVK). Results are representative of two independent experiments. a-, anti-.
Subsequent experiments determined whether antigen receptor–mediated activation of PKD was also regulated via classical/novel PKCs in T lymphocytes and mast cells. GF 109203X is a second inhibitor of classical and novel PKC enzymes that also has no direct inhibitory effect on PKD 28 39. Peripheral blood–derived T lymphoblasts were preincubated with GF 109203X, the TCR complex was activated, and PKD activity was measured by in vitro kinase assays. As indicated in Fig. 5 C, pretreatment of T lymphoblasts with GF 109203X did not effect the constitutive low basal activity of PKD present in these cells, but severely abrogated TCR-induced activation of PKD. Addition of this PKC inhibitor directly to PKD during in vitro kinase assays did not effect the catalytic activity of PKD (Fig. 5 C), confirming that this inhibitor has no direct action towards PKD.
The obligatory role played by classical/novel PKCs in mediating the activation of PKD by antigen receptors was also demonstrated in mast cells. Here, pretreatment of RBL 2H3 cells with the PKC inhibitor Ro 31-8220 abrogated the ability of the FcεR1 to induce PKD catalytic activity (Fig. 5 D). Together, these results indicate that antigen receptor–induced activation of PKD is regulated through a PKC-dependent signaling pathway in T and B lymphocytes and in mast cells.
PKC Activation Is Sufficient to Induce PKD Activity in Lymphocytes.
To address whether PKC activity was sufficient to induce the activation of PKD in lymphocytes, we transiently expressed a constitutively active PKCη mutant in B lymphocytes and examined its effect on the activity of endogenous PKD. Control A20 B lymphocytes, transfected with an empty expression vector, exhibited a low basal level of PKD activity (as shown by Western blotting with the pS916 antibody) that was markedly enhanced upon triggering of the BCR complex (Fig. 6 A). In contrast, A20 B cells transfected with the constitutively active PKCη mutant displayed a high basal level of PKD activity that was only slightly further enhanced by activation of the BCR complex (Fig. 6 A). Similar results were observed in the RBL 2H3 mast cell line, where ectopic expression of constitutively activated mutants of PKCη or PKCε was found to induce near-maximal PKD activity in the absence of FcεR1 engagement (Fig. 6 B), as measured by in vitro kinase assays. In contrast, control RBL 2H3 cells transfected with an empty expression vector exhibited a low basal level of PKD activity (Fig. 6 B). Thus, a PKC-dependent signaling pathway is both required and sufficient to control PKD activation in lymphocytes.
Figure 6.


Constitutively active PKC enzyme mutants induce PKD activation. (A) A20 B lymphocytes were transiently transfected with 20 μg of either vector control (V) or activated PKCη (PKCη*) cDNA. After 16 h of incubation at 37°C, the cells were left nonstimulated (−) or were treated with 10 μg/ml F(ab)′2 fragments of anti–mouse IgG for 2 min (+). Whole cell lysates were analyzed by Western blotting with the pS916 antiserum and a pan–COOH-terminal PKD antibody. (B) RBL 2H3 mast cells were transiently transfected with 20 μg of either vector control (V), activated PKCε (PKCε*), or activated PKCη (PKCη*) cDNA. After 16 h of incubation at 37°C, the cells were left nonstimulated (−) or were stimulated with IgE plus KLH-DNP (IgE+KLH-DNP) for 2 min (+). PKD was immunoprecipitated from cell lysates, and PKD activity was measured by in vitro kinase assays (IVK) measuring PKD autophosphorylation. All data are representative of two independent experiments. a-, anti-.
Conclusions.
Historically, PKC enzymes were positioned in the earliest models of antigen receptor signal transduction, but there has been little real progress in identifying targets for PKC signals in lymphocytes. This study identifies a PKC signaling network in T cells, B cells, and mast cells that couples antigen receptors to the serine kinase PKD. The regulatory region of PKD contains a conserved DAG-binding C1 domain, but the integrity of this domain is not required for antigen receptor activation of PKD. Rather, PKD activity is dependent on activation of PKCs; specifically, inhibitors of classical and novel PKCs (that do not directly effect PKD activity) block the activation of PKD after BCR, TCR, or FcεR1 ligation. Furthermore, expression of constitutively active mutants of novel PKC enzymes (PKCη and PKCε) was sufficient to maximally stimulate PKD catalytic activity in lymphocytes. The phosphorylation of two critical serine residues within the catalytic domain of PKD is essential for activation of this enzyme 29 30. Future studies will determine whether these sites in PKD are directly phosphorylated by PKC or whether an intermediate kinase is involved. The role of PKCs in the regulation of PKD identify a novel mechanism by which a DAG signal generated by antigen receptor ligation can be disseminated via a PKC to PKD serine kinase cascade. The data presented here establish PKD as a PKC-regulated kinase in lymphocytes. Antigen receptors in T cells, B cells, and mast cells activate unique tyrosine kinases. They may also activate unique PKC isoforms; PKCβ is associated with BCR function, whereas PKCθ is linked to the TCR signaling. Herein, we establish PKD as a common downstream target for PKCs in lymphocytes and identify the PKC/PKD serine kinase cascade as a common element in antigen receptor signaling.
Acknowledgments
We thank P. Parker for the constitutively active PKCη and PKCε constructs. We also thank members of the Lymphocyte Activation Laboratory for useful discussions.
This work was supported by the Imperial Cancer Research Fund. E. Rozengurt is supported by National Institutes of Health grant DK-55003-01.
Footnotes
Abbreviations used in this paper: BCR, B cell antigen receptor; Erk, extracellular signal–regulated kinase; GFP, green fluorescent protein; PLC, phospholipase C; PKC, protein kinase C; PKD, protein kinase D; RBL, rat basophilic leukemia.
References
- DeFranco A.L. The complexity of signaling pathways activated by the BCR. Curr. Opin. Immunol. 1997;9:296–308. doi: 10.1016/s0952-7915(97)80074-x. [DOI] [PubMed] [Google Scholar]
- Kinet J.-P. The high affinity IgE receptor (FcεR1)from physiology to pathology. Annu. Rev. Immunol. 1999;17:931–972. doi: 10.1146/annurev.immunol.17.1.931. [DOI] [PubMed] [Google Scholar]
- Qian D., Weiss A. T cell antigen receptor signal transduction. Curr. Opin. Cell Biol. 1997;9:205–212. doi: 10.1016/s0955-0674(97)80064-6. [DOI] [PubMed] [Google Scholar]
- Weiss A., Littman D.R. Signal transduction by lymphocyte antigen receptors. Cell. 1994;76:263–274. doi: 10.1016/0092-8674(94)90334-4. [DOI] [PubMed] [Google Scholar]
- Williams D.H., Woodrow M., Cantrell D.A., Murray J.E. Protein kinase C is not a downstream effector of p21ras in activated T cells. Eur. J. Immunol. 1995;25:42–47. doi: 10.1002/eji.1830250109. [DOI] [PubMed] [Google Scholar]
- Fahraeus R., Lane D.P. The p16INK4α tumour suppressor protein inhibits αvβ3 integrin-mediated cell spreading on vitronectin by blocking PKC-dependent localisation of αvβ3 to focal contacts. EMBO (Eur. Mol. Biol. Organ.) J. 1999;18:2106–2118. doi: 10.1093/emboj/18.8.2106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ng T., Shima D., Squire A., Bastiaens P.I., Gschmeissner S., Humphries M.J., Parker P.J. PKCa regulates β1-integrin-dependent cell motility through association and control of integrin traffic. EMBO (Eur. Mol. Biol. Organ.) J. 1999;18:3909–3923. doi: 10.1093/emboj/18.14.3909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ozawa K., Szallis Z., Kazanietz M.G., Blumberg P.M., Mischak H., Mushinski J.F., Beaven M.A. Calcium dependent and independent isozymes of protein kinase C mediate exocytosis in antigen stimulated rat basophilic RBL 2H3 cells. J. Biol. Chem. 1993;268:1749–1756. [PubMed] [Google Scholar]
- Valge V.E., Wong J.G., Datlof B.M., Sinskey A.J., Rao A. Protein kinase C is required for responses to T cell ligands but not to interleukin-2 in T cells. Cell. 1988;55:101–112. doi: 10.1016/0092-8674(88)90013-x. [DOI] [PubMed] [Google Scholar]
- Genot E.M., Parker P.J., Cantrell D.A. Analysis of the role of protein kinase C-alpha, -epsilon, and -zeta in T cell activation. J. Biol. Chem. 1995;270:9833–9839. doi: 10.1074/jbc.270.17.9833. [DOI] [PubMed] [Google Scholar]
- Toker A. Signaling through protein kinase C. Front. Biosci. 1998;3:D1134–D1147. doi: 10.2741/a350. [DOI] [PubMed] [Google Scholar]
- Newton A.C. Regulation of protein kinase C. Curr. Opin. Cell Biol. 1997;9:161–167. doi: 10.1016/s0955-0674(97)80058-0. [DOI] [PubMed] [Google Scholar]
- Mellor H., Parker P.J. The extended protein kinase C superfamily. Biochem. J. 1998;332:281–292. doi: 10.1042/bj3320281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leitges M., Schmedt C., Guinamard R., Davoust J., Schaal S., Stabel S., Tarakhovsky A. Immunodeficiency in protein kinase Cβ-deficient mice. Science. 1996;273:788–791. doi: 10.1126/science.273.5276.788. [DOI] [PubMed] [Google Scholar]
- Monks C.R., Kupfer H., Tamir I., Barlow A., Kupfer A. Selective modulation of protein kinase C-theta during T-cell activation. Nature. 1997;385:83–86. doi: 10.1038/385083a0. [DOI] [PubMed] [Google Scholar]
- Monks C.R., Freiberg B.A., Kupfer H., Sciaky N., Kupfer A. Three-dimensional segregation of supramolecular activation clusters in T cells. Nature. 1998;395:82–86. doi: 10.1038/25764. [DOI] [PubMed] [Google Scholar]
- Valverde A.M., Sinnett-Smith J., Van Lint J., Rozengurt E. Molecular cloning and characterization of protein kinase Da target for diacylglycerol and phorbol esters with a distinctive catalytic domain. Proc. Natl. Acad. Sci. USA. 1994;91:8572–8576. doi: 10.1073/pnas.91.18.8572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johannes F.J., Prestle J., Eis S., Oberhagemann P., Pfizenmaier K. PKCμ is a novel, atypical member of the protein kinase C family. J. Biol. Chem. 1994;269:6140–6148. [PubMed] [Google Scholar]
- Johannes F.J., Horn J., Link G., Haas E., Siemienski K., Wajant H., Pfizenmaier K. Protein kinase Cμ downregulation of tumor-necrosis-factor-induced apoptosis correlates with enhanced expression of nuclear-factor-kappaB-dependent protective genes. Eur. J. Biochem. 1998;257:47–54. doi: 10.1046/j.1432-1327.1998.2570047.x. [DOI] [PubMed] [Google Scholar]
- Prestle J., Pfizenmaier K., Brenner J., Johannes F.-J. Protein kinase C μ is located at the golgi compartment. J. Cell Biol. 1996;134:1401–1410. doi: 10.1083/jcb.134.6.1401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jamora C., Yamanouye N., Van Lint J., Vandenheede J.R., Faulkner D.J., Malhotra V. Gβγ-mediated regulation of golgi organization is through the direct activation of protein kinase D. Cell. 1999;98:59–68. doi: 10.1016/S0092-8674(00)80606-6. [DOI] [PubMed] [Google Scholar]
- Dieterich S., Herget T., Link G., Bottinger H., Pfizenmaier K., Johannes F.J. In vitro activation and substrates of recombinant, baculovirus expressed human protein kinase C mu. FEBS Lett. 1996;381:183–187. doi: 10.1016/0014-5793(96)00116-0. [DOI] [PubMed] [Google Scholar]
- Van Lint J., Sinnett-Smith J., Rozengurt E. Expression and characterisation of PKD, a phorbol ester and diacylglycerol-stimulated serine protein kinase. J. Biol. Chem. 1995;270:1445–1461. doi: 10.1074/jbc.270.3.1455. [DOI] [PubMed] [Google Scholar]
- Matthews S.A., Rozengurt E., Cantrell D. Characterisation of S916 as an in vivo autophosphorylation site for PKD/PKCμ. J. Biol. Chem. 1999;274:23543–26549. doi: 10.1074/jbc.274.37.26543. [DOI] [PubMed] [Google Scholar]
- Sidorenko S.P., Law C.L., Klaus S.J., Chandran K.A., Takata M., Kurosaki T., Clark E.A. Protein kinase Cμ (PKCμ) associates with the B cell antigen receptor complex and regulates lymphocyte signaling. Immunity. 1996;5:353–363. doi: 10.1016/s1074-7613(00)80261-7. [DOI] [PubMed] [Google Scholar]
- Matthews S., Iglesias T., Cantrell D., Rozengurt E. Dynamic redistribution of protein kinase D (PKD) as revealed by a GFP-PKD fusion proteindissociation from PKD activation. FEBS Lett. 1999;457:515–521. doi: 10.1016/s0014-5793(99)01090-x. [DOI] [PubMed] [Google Scholar]
- Zugaza J.L., Waldron R.T., Sinnett-Smith J., Rozengurt E. Bombesin, vasopressin, endothelin, bradykinin, and platelet-derived growth factor rapidly activate protein kinase D through a protein kinase C-dependent signal transduction pathway. J. Biol. Chem. 1997;272:23952–23960. doi: 10.1074/jbc.272.38.23952. [DOI] [PubMed] [Google Scholar]
- Zugaza J.L., Sinnett-Smith J., Van Lint J., Rozengurt E. Protein kinase D (PKD) activation in intact cells through a protein kinase C-dependent signal transduction pathway. EMBO (Eur. Mol. Biol. Organ.) J. 1996;15:6220–6230. [PMC free article] [PubMed] [Google Scholar]
- Iglesias T., Waldron R.T., Rozengurt E. Identification of in vivo phosphorylation sites required for protein kinase D activation. J. Biol. Chem. 1998;273:27662–27667. doi: 10.1074/jbc.273.42.27662. [DOI] [PubMed] [Google Scholar]
- Waldron R.T., Iglesias T., Rozengurt E. Phosphorylation-dependent protein kinase D activation. Electrophoresis. 1999;20:382–390. doi: 10.1002/(SICI)1522-2683(19990201)20:2<382::AID-ELPS382>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- Moore E.D.W., Ring M., Scriven D.R.L., Smith V.C., Meloche R.M., Buchan A.M.J. The role of protein kinase C isozymes in bombesin-stimulated gastrin release from human antral gastrin cells. J. Biol. Chem. 1999;274:22493–22501. doi: 10.1074/jbc.274.32.22493. [DOI] [PubMed] [Google Scholar]
- Iglesias T., Matthews S., Rozengurt E. Dissimilar phorbol ester binding properties of the individual cysteine-rich motifs of protein kinase D. FEBS Lett. 1998;437:19–23. doi: 10.1016/s0014-5793(98)01189-2. [DOI] [PubMed] [Google Scholar]
- Turner H., Cantrell D.A. Distinct Ras effector pathways are involved in Fc epsilon R1 regulation of the transcriptional activity of Elk-1 and NFAT in mast cells. J. Exp. Med. 1997;185:43–53. doi: 10.1084/jem.185.1.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beadling C., Guschin D., Wittuhn B.A., Ziemiecki A., Ihle J.N., Kerr I.M., Cantrell D.A. Activation of JAK kinases and STAT proteins by interleukin-2 and interferon-α, but not the T cell antigen receptor, in human T lymphocytes. EMBO (Eur. Mol. Biol. Organ.) J. 1994;13:5605–5615. doi: 10.1002/j.1460-2075.1994.tb06898.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Izquierdo Pastor M., Reif K., Cantrell D. The regulation and function of p21ras during T cell activation and growth. Immunol. Today. 1995;16:159–164. doi: 10.1016/0167-5699(95)80134-0. [DOI] [PubMed] [Google Scholar]
- Reif K., Burgering B.M.T., Cantrell D.A. Phosphatidylinositol 3-kinase links the interleukin-2 receptor to protein kinase B and p70 S6 kinase. J. Biol. Chem. 1997;272:14426–14438. doi: 10.1074/jbc.272.22.14426. [DOI] [PubMed] [Google Scholar]
- Van Lint J., Ni Y., Valius M., Merlevede W., Vandenheede J.R. Platelet-derived growth factor stimulates protein kinase D through the activation of phospholipase Cgamma and protein kinase C. J. Biol. Chem. 1998;273:7038–7043. doi: 10.1074/jbc.273.12.7038. [DOI] [PubMed] [Google Scholar]
- Rozengurt E., Sinnett-Smith J., Zugaza J.L. Protein kinase Da novel target for diacylglycerol and phorbol esters. Biochem. Soc. Trans. 1997;25:565–571. doi: 10.1042/bst0250565. [DOI] [PubMed] [Google Scholar]
- Matthews S.A., Pettit G.R., Rozengurt E. Bryostatin 1 induces biphasic activation of protein kinase D in intact cells. J. Biol. Chem. 1997;272:20245–20250. doi: 10.1074/jbc.272.32.20245. [DOI] [PubMed] [Google Scholar]