

Abstract
Notch1 signaling is required for T cell development. We have previously demonstrated that expression of a dominant active Notch1 (ICN1) transgene in hematopoietic stem cells (HSCs) leads to thymic-independent development of CD4+CD8+ double-positive (DP) T cells in the bone marrow (BM). To understand the function of Notch1 in early stages of T cell development, we assessed the ability of ICN1 to induce extrathymic T lineage commitment in BM progenitors from mice that varied in their capacity to form a functional pre-T cell receptor (TCR). Whereas mice repopulated with ICN1 transduced HSCs from either recombinase deficient (Rag-2−/−) or Src homology 2 domain–containing leukocyte protein of 76 kD (SLP-76)−/− mice failed to develop DP BM cells, recipients of ICN1-transduced Rag-2−/− progenitors contained two novel BM cell populations indicative of pre-DP T cell development. These novel BM populations are characterized by their expression of CD3ε and pre-Tα mRNA and the surface proteins CD44 and CD25. In contrast, complementation of Rag-2−/− mice with a TCRβ transgene restored ICN1-induced DP development in the BM within 3 wk after BM transfer (BMT). At later time points, this population selectively and consistently gave rise to T cell leukemia. These findings demonstrate that Notch signaling directs T lineage commitment from multipotent progenitor cells; however, both expansion and leukemic transformation of this population are dependent on T cell–specific signals associated with development of DP thymocytes.
Keywords: leukemia, development, hematopoiesis, lymphocyte, stem cells
Introduction
Notch proteins comprise a family of transmembrane receptors that regulate responsiveness of a wide variety of cell types to environmental signals 1. Notch1, one of four family members, binds multiple ligands, including Delta/Serrate and Jagged 2. After ligand binding, the intracellular region of Notch1 is proteolytically cleaved from the transmembrane portion and translocated to the nucleus where it interacts with a transcriptional repressor, known as CSL (for CBF-1, suppressor of Hairless, and Lag-1) 3. After binding Notch, CSL becomes a transcriptional activator, and in conjunction with the activity of other cofactors, including Mastermind, SKIP, PCAF, and/or GCN5, induces transcription of downstream targets 4 5 6. Notch signaling also occurs through a poorly understood CSL-independent pathway that may involve Deltex 7.
Several studies examining Notch1 expression patterns and the effect of removing or deregulating Notch1 activity suggest that Notch1 signaling plays a key role at multiple steps in T cell development 8. The importance of Notch1 in commitment of early progenitors to the T cell lineage is underscored by two recent studies. T cells fail to develop in inducible Notch1 knockout mice 9. Reciprocally, we have demonstrated that expression of active Notch1 in stem cells induces ectopic CD4+CD8+ development in the bone marrow (BM) and inhibits early B cell development 10. Although these studies clearly show that Notch1 is required for T cell development, the signaling pathways and target genes underlying Notch1-mediated T cell commitment are unknown.
Early α/β T cell development in the thymus is characterized by lineage commitment followed by VβDβJβ recombination and differentiation of precursors expressing a functional pre-TCR into early CD4+CD8+ thymocytes (for a review, see reference 11). Lineage commitment and VβDβJβ recombination occur in discrete cell populations defined by differential surface expression of CD44 and CD25. Early thymic progenitors within the CD44+CD25− subset give rise to dendritic cells, NK cells, and T lineage–committed CD44+CD25+ early pro-T cells. Pre-TCR signaling is requisite for differentiation of late CD44−CD25+ pro-T cells into CD4+CD8+ thymocytes as evidenced by the arrest in T cell development at the CD44−CD25+ stage after disruption of pre-TCR assembly or signaling 11. This phenotype is shared by recombination activating gene (Rag)-1/2−/− mice and mice lacking the pre-TCR intracellular adapter protein Src homology 2 domain–containing leukocyte protein of 76 kD (SLP-76) 12 13 14 15. Pre-TCR signaling is also required for the exponential expansion of thymocyte numbers that occur after successful VDJ recombination, as evidenced by decreased thymic cellularity in Rag- and SLP-76–deficient mice 12 13 14 15.
To understand the function of Notch signaling at the earliest stages of T cell development, we used the ability of Notch1 to drive thymic-independent T cell development in the BM of recipients of ICN1-transduced hematopoietic stem cells (HSCs). When ICN1 is introduced into HSCs of either Rag-2−/− or SLP-76−/− mice, double-positive (DP) cells fail to develop in the BM. However, ICN1 does promote earlier stages of T cell development (CD44+CD25+ and CD44−CD25+) in the BM of Rag-2−/− mice. These progenitors constitute a small fraction of total BM cells in these chimeric mice, which remained healthy for over 1 yr. In contrast, rescue of the Rag-2−/− phenotype with a TCRβ transgene allows Notch to drive DP development in the BM within 3 wk after BM transfer (BMT). At later time points, these cells give rise to T cell leukemias. These results show that Notch is required for T cell commitment but insufficient to rescue the block in pre-TCR signaling necessary for either T cell expansion or DP development. Furthermore, signaling by an intact TCR provides a permissive environment for the development of T cell leukemia.
Materials and Methods
Mice.
BALB/c, C57BL/6, Rag-2−/−, and Rag-2−/− × TCRβ (DO11.10) transgenics were purchased from Taconic. All experiments were conducted in accordance with National Institutes of Health guidelines for the care and use of animals and with an approved animal protocol from the University of Pennsylvania Animal Care and Use Committee.
Retroviral Transduction of BM Precursor Cells.
The retroviral vectors used in these experiments have been described previously 10 16. Transduction of BM cells from 5-fluorouracil (5-FU)–treated 4–8-wk-old mice with green fluorescent protein (GFP)-normalized retroviral supernatants and transplantation of these cells into lethally irradiated 4–8-wk-old female syngeneic BALB/c or C57BL/6 mice was performed as described 10 16. Spinoculations were performed in medium containing IL-3 (6 ng/ml; R&D Systems), IL-6 (10 ng/ml; R&D Systems), stem cell factor (SCF; 100 ng/ml; R&D Systems), and 5% WEHI-conditioned supernatant as described 10 16. On day 3 after BM harvest, 3 × 105 cells were injected into syngeneic 4–8-wk-old female mice that had been lethally irradiated (900 rads in a split dose). Furthermore, identical aliquots of MigR1 and Mig ICN1 retroviral supernatants were used to transduce BM progenitors from Rag-2−/− and Rag-2−/− × TCRβ (DO11.10) mice in two independent experiments.
Flow Cytometry and Cell Sorting.
BM cells were prepared and stained for flow cytometric analysis as described previously 10. In brief, 106 BM cells were stained with PE-conjugated anti-CD8 (Lyt2) and allophycocyanin (APC) anti-CD4 (L3T4) (BD PharMingen), then analyzed on a dual laser FACSCalibur™ equipped with CELLQuest™ software (Becton Dickinson). Cells derived from transduced precursors were distinguished from nontransduced cells by gating on cells with greater than two logs GFP expression over GFP− cells. For sorting, BM cells from recipients of Rag-2−/− ICN1-transduced HSCs were stained with PE–anti-CD25 and APC–anti-CD44, then sorted on a Becton Dickinson FACS Vantage™ equipped with argon 488-nm and HeNe 647-nm lasers. For cell cycle analysis, cells were permeabilized in EtOH and then stained with 50 μg/ml propidium iodide and 100 μg/ml RNaseA for 30 min before analysis. All flow cytometric data were analyzed by uploading forward scatter (FSC) files into FlowJo (Tree Star, Inc.).
Reverse Transcription PCR.
RNA and cDNA was prepared and amplified as described 17. In brief, 105 cells from each population were sorted directly into 400 μl of RNA lysis buffer consisting of 0.236 M guanidine isothyocyanate, 0.5% sarkosyl, 25 mM Na Citrate, pH 7.0, and 0.7% (vol/vol) 2-ME. After a phenol/chloroform extraction, RNA was precipitated with isopropanol using 1 μg of glycogen as a carrier then resuspended in 20 μl ddH2O. 4 μl of RNA was used for cDNA synthesis in a final volume of 20 μl with 1 μl of 1 μg/μl random hexamers, 1 μl M-MLV reverse transcriptase (200 U/μl), and 0.5 mM dNTPs. For amplification, 2–4 μl of cDNA was subjected to a 35 cycle PCR with the following oligonucleotides: CD3ε, 5′ TGGAACACTTTCTGGGGCATCCTG and 3′ TTTGAGGCTGGTGTGTAGCAGACG; pre-Tα, 5′ CTCGAGATGCTTCTCCACGAGT and 3′ GTTAACACTATGTCCAAATTCT; hypoxanthine phosphoribosyltransferase (HPRT), 5′ CACAGGACTAGAACACCTGC and 3′ GCTGGTGAAAAGGACCTCT. For a positive control for pre-Tα expression, cDNA from the SCID-derived early T cell line SCID.ADH 18 was provided by Dr. David Wiest (Fox Chase Cancer Center, Philadelphia, PA). To ensure that analyses were in the linear range, 10-μl aliquots were withdrawn at 25, 30, and 35 cycles for separate analysis on 1% agarose gels. HPRT served as a control to ensure that equal amounts of cDNA were added to each reaction.
Western Blot for Notch1 Expression.
Expression of Notch1 polypeptides was determined by Western blot analysis of whole cell detergent lysates using anti-Notch1 rabbit sera 16. DNA prepared from diseased and normal tissues by SDS extraction and proteinase K digestion were analyzed on Southern blots hybridized to probes for the internal ribosomal entry site (IRES) or murine TCRβ, as described previously 10 16.
Leukemogenesis Assay.
Mice were followed after transplantation by periodic peripheral blood sampling from retroorbital sinuses and daily physical inspection. Peripheral blood (PB) was assessed beginning 2 wk after transplant and every 2 wk thereafter for the presence of GFP+ immature T cells by flow cytometric analysis using antibodies to CD4 and CD8 (see below). Tissues were harvested from diseased and unaffected animals after asphyxiation with CO2. Mice were monitored for leukemia by enumerating the white blood cells (WBCs) once every 2 wk and monitoring the mice daily for signs of cachexia, inactivity, or ill health. Criteria for the development of leukemia were a WBC >50 × 106 cells/ml and/or the presence of circulating DP T cells 16.
Results
Notch1-induced Extrathymic T Cells Resemble Normal DP Thymocytes.
To dissect the role of Notch signaling in early events in T cell development, we exploited the ability of ICN1 to drive thymic-independent T cell development in the BM of recipients of ICN1-transduced HSCs 10. Between 3–4 wk after reconstitution, recipient BM contained T lineage cells that closely resembled normal immature DP thymocytes (Fig. 1). Both ICN1-induced DP BM cells and wild-type DP thymocytes expressed similar levels of TCR, CD3, and CD4/CD8 (Fig. 1A and Fig. B, and data not shown). Furthermore, both ICN1-induced and wild-type DP thymocytes were comprised of small resting cells (Fig. 1C and Fig. D). Our previous results showed that the ICN1-induced BM T cell population was oligoclonal 10. These results suggest that the ICN1-induced BM T cell population provides a physiologically relevant model to study T cell commitment from multipotential progenitors as well as the double negative (DN) to DP transition requisite for thymocyte development.
Figure 1.

Notch1-induced extrathymic T cells resemble normal DP thymocytes. (A) Flow cytometric analysis for CD4 and CD8 expression on normal thymocytes versus BM cells from a recipient of ICN1-transduced wild-type progenitor cells at 21 d after BMT. Also shown is TCRβ expression (B), forward scatter (C), and DNA content (D). The TCRβ expression (B) and forward scatter (C) results are based on DP GFP+ cells as shown in panel A. Because GFP is lost after cell permeabilization, the analysis in D used ungated BM cells; however, 95% of the cells in the BM at the time of analysis were DP GFP+. Data are representative of three recipients.
Notch1 Signaling Fails to Rescue T Cell Development in Rag-deficient Cells.
Thymocytes from Rag-deficient mice (Rag-2−/−) do not undergo VDJ recombination, and lacking pre-TCR signaling, fail to differentiate into DP cells 15. Instead, these thymocytes arrest at the CD44−CD25+ stage of development. The Rag-2−/− induced arrest can be rescued by activation of lck or the mitogen-activated protein (MAP) kinase pathway 19 20 21, suggesting that activation of appropriate downstream signaling pathways can compensate for lack of pre-TCR signaling. To evaluate the ability of activated Notch1 to rescue the Rag-2−/− block in T cell development, hematopoietic progenitors from Rag-2−/− BM were transduced with ICN1 and transferred into lethally irradiated recipients. Whereas mice repopulated with ICN1-transduced wild-type HSCs generated BM DP T cells within 3 wk after BMT, mice repopulated with ICN1 transduced Rag-2−/− HSCs did not, even when followed for 1 yr (Fig. 2, and data not shown). Similarly, progenitors from SLP-76−/− mice, which lack an essential component of the pre-TCR signaling pathway 12 14, failed to give rise to ICN1-induced DP T cells in the BM (Fig. 2). In contrast, the hematopoietic defects in SLP-76−/− mice were rescued by retroviral transduction of SLP-76−/− BM cells with SLP-76 (unpublished data). Most BM samples from recipients also contained both CD4+CD8− and CD4−CD8+ cells. However, these cells were host derived, as they were evident regardless of donor cell genotype, and uniformly expressed the host Ly5 allele in separate experiments using Ly5 congeneic donors and recipients (Fig. 2, and data not shown).
Figure 2.

Notch1 induction of extrathymic DP T cells requires pre-TCR assembly and signaling. Progenitor cells from wild-type, Rag-2−/−, Rag-2−/− × TCRβ, and SLP-76 mice were transduced with MigR1 control or MigR1-ICN1 viruses and then transferred to lethally irradiated C57BL/6 recipients. BM cells were stained 20–30 d after transfer with the indicated antibodies and analyzed by flow cytometry. GFP gates were set as described in Materials and Methods. The CD4+ and CD8+ SP thymocytes present in all GFP− populations are recipient derived, as shown by prior congenic analysis (data not shown). Data are representative of 3–12 recipients for each donor genotype.
To determine whether BM DP development could be restored by pre-TCR generated signals, chimeras were generated after ICN1 transduction of HSCs from Rag-2−/− mice complemented with the TCRβ transgene DO11.10 22. Within 3 wk after reconstitution with ICN1-expressing BM cells, these mice developed DP cells in the BM (Fig. 2). Together, these results demonstrate that pre-TCR–derived signals are necessary for the development of Notch1-induced extrathymic DP T cells.
Notch Signaling Drives T Cell Commitment but Not Expansion in the BM.
Although mice repopulated with ICN1-transduced Rag-2−/− HSCs failed to develop BM DP cells, these mice exhibited unique cell populations in the BM that parallel earlier stages of thymic T cell development. ICN1-transduced HSCs from Rag-2−/− mice developed into both CD44+CD25+ and CD44−CD25+ populations in the BM after adoptive transfer (Fig. 3 A). These populations were restricted to the GFP+ fraction of mice given ICN1-transduced HSCs, as they were absent from mice given MigR1-transduced HSCs. We showed previously that GFP expression mirrors ICN1 expression driven by the Mig ICN1 vector 10 23, and that the development of BM CD4+CD8+ cells induced by Mig ICN1 is thymus independent. To confirm that the CD44+CD25+ and CD44−CD25+ cells in the ICN1 mice are T lineage precursors, we isolated each population and performed reverse transcription (RT)-PCR for CD3ε and pre-Tα; two components of the pre-TCR complex 24. Both CD44+CD25+ and CD44−CD25+ populations express transcripts for CD3ε and pre-Tα, whereas other BM populations, which are common to cells derived from both MigR1 and ICN1 transduced HSCs, do not (Fig. 3 B). Together, these data support a model in which Notch1 activity is sufficient to induce commitment to the T cell lineage, instructing early multipotent progenitors in the BM to differentiate into T lineage precursor populations.
Figure 3.
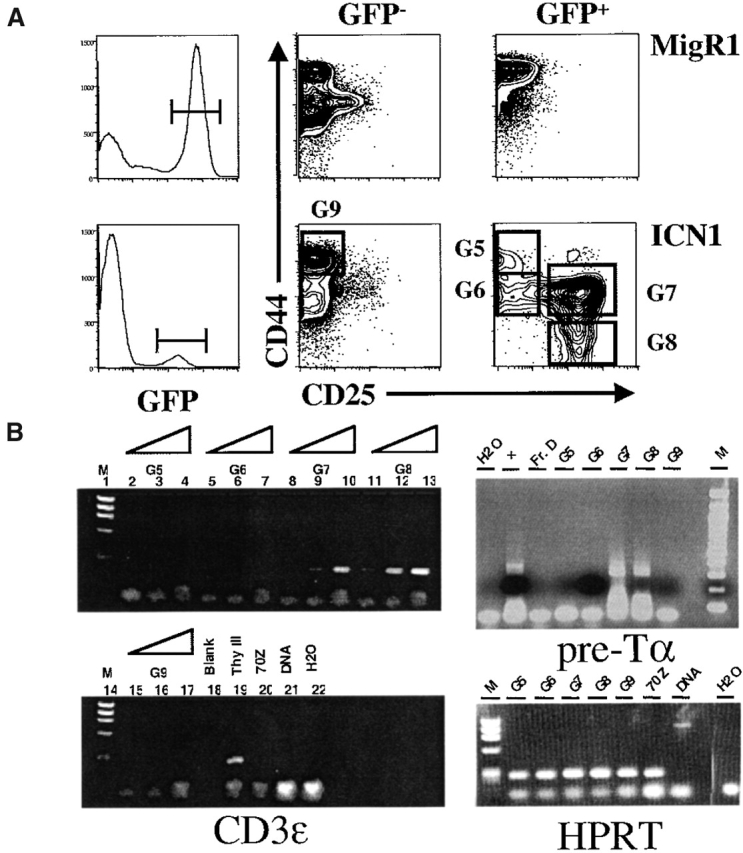
Notch1 signaling induces extrathymic T cell commitment. Progenitor cells from Rag-2−/− donors were transduced with MigR1 or MigR1-ICN1 then analyzed 30 d after transfer. (A) Flow cytometric analysis for CD44 and CD25 expression on BM cells from recipients. Data are representative of six recipients of ICN1-transduced Rag-2−/− progenitors. (B) RT-PCR analysis for CD3ε and pre-Tα mRNA was performed as described in Materials and Methods on sorted populations using the indicated gates. In the analysis of pre-Tα expression cDNA from the SCID-derived early T cell line SCID.ADH (reference 18) was included as a positive control (lane marked “+”), and from Hardy Fraction D (B220+CD43−IgM− pre-B cells) as a negative control (lane marked “Fr. D”). Thy III contains cDNA from purified CD25+CD44−CD3− thymocytes.
In wild-type mice, pre-TCR signaling results in exponential expansion of the T cell precursor pool. This is mimicked in our murine model as the ICN1-induced BM DP population in both wild-type mice and Rag-2−/− × TCRβ mice rapidly expanded to become the dominant cell population in the BM and spleen 10. In contrast, the ICN1-induced CD44+CD25+ and CD44−CD25+ BM populations constituted 1–4% of all BM cells, and were not detected in other organs (Fig. 4 B, and data not shown). These findings suggest that Notch activity commits lymphoid precursors to the T lineage; however, pre-TCR signals are required for the proliferative burst that accompanies thymocyte differentiation.
Figure 4.


Notch1 induction of T cell leukemia requires a functional TCRβ chain. Progenitor cells from wild-type, Rag-2−/−, and Rag-2−/− × TCRβ (DO11.10) donors were transduced with MigR1 or MigR1-ICN1 before transfer to irradiated adoptive hosts. (A) Leukemia onset in mice reconstituted with retrovirally transduced BM cells. Leukemia onset is defined as the time after BMT when CD4+CD8+ DP GFP+ T cells appear in the peripheral circulation, and/or the WBC exceeds 50 × 106 cells/ml. Mice typically survived for ∼1–2 mo after the onset of leukemia, during which time the fraction and number of DP GFP+ cells continued to increase (not shown). None of the mice reconstituted with the Rag-2−/− receiving Mig ICN or any of the mice receiving the empty MigR1 virus developed circulating DP GFP+ cells or any evidence of neoplasia. Each dot on the Mig ICN Rag-2−/− × TCRβ (DO11.10) curve represents the analysis of two mice. Although this figure shows leukemia-free survival to 100 d, these mice remained leukemia-free for the duration of this study (see Table ) (B). Western blot analysis of Notch1 expression in BM cells of recipients of ICN1-transduced progenitors from Rag-2−/− and Rag-2−/− × TCRβ (DO11.10) donors.
Pre-TCR Signals Provide a Permissive Environment for Notch Induction of T Cell Leukemia.
Although the Notch1-induced BM DP population in wild-type mice is initially composed of a oligoclonal population of small resting lymphocytes, additional events (presumably the acquisition of other genetic lesions) ensue that result in a lethal monoclonal leukemia. We compared the ability of ICN1 to induce T cell leukemia when expressed in Rag-2−/− HSCs, in which T lineage commitment but not DP development is intact, with Rag-2−/− × TCRβ DO11.10 HSCs, in which both T lineage commitment and DP development are unimpaired (Table ). Recipients of ICN1 transduced HSCs from Rag-2−/− mice and control MigR1 transduced HSCs from wild-type mice remained alive for >1 yr after transfer without developing any features of leukemia (Fig. 4 A). In sharp contrast, every mouse receiving ICN1 transduced HSCs from wild-type or Rag-2−/− mice expressing the DO11.10 TCRβ transgene developed T cell leukemia between 9 and 11 wk after BMT (Fig. 4 A). The T cell leukemias were similar in latency and phenotype to those described previously 16 25, being composed of cycling cells resembling either the immature single positive (ISP) or DP stages of T cell development (not shown). Southern blotting showed that all tumors contained proviral DNA of the expected size and were donor derived, as judged by the presence of the TCRβ transgene in each sample (data not shown). Recipients of ICN1-transduced progenitors from both Rag-2−/− and Rag-2−/− × TCRβ progenitors expressed readily detectable levels of Notch1 protein as assessed by Western blot analysis (Fig. 4 B). ICN1 protein expression in the Rag-2−/− mice was limited to the BM; whereas, ICN1 expression in the Rag-2−/− × TCRβ mice was found in all organs infiltrated by tumor cells (Fig. 4 B, and not shown). Thus, ICN1-mediated transformation of T cell progenitors required expression of a TCRβ chain and development of CD4+CD8+ T cells. More broadly, ICN1 expression in Rag2−/− HSCs did not induce malignancy in other hematopoietic lineages, suggesting that Notch1-mediated oncogenesis is sharply restricted to the context of CD4+CD8+ T cell progenitors.
Table 1.
Onset of Notch1-induced Leukemia in Recipients of Rag-2− /− versus Rag-2− /− × TCRβ Progenitors
| Donor cells | Construct | No. of BM recipients | No. devoloping leukemia | Mean onset (d) | Range of onset (d) |
|---|---|---|---|---|---|
| Rag2−/− | MigR1 | 6 | 0 | (455) | − |
| ICN | 12 | 0 | (455) | − | |
| Rag2−/− × TCRβ | MigR1 | 5 | 0 | (243) | − |
| ICN | 6 | 6 | 65 | 59–76 |
The number in parentheses in the “mean onset” column indicate the length of time that these mice were followed. None of these mice developed a malignancy.
Discussion
Our findings reveal two important facets regarding the induction of T cell development 10 and T cell leukemia 16 25 by Notch1 signaling. First, we show that Notch1 activity is sufficient to drive commitment of HSCs to early T cell progenitor subsets; however, an intact pre-TCR signaling pathway is required to both expand this population and induce differentiation into DP T cells. Thus, Notch1 signaling induces an extrathymic (but otherwise normal) T cell developmental program. Second, we show that Notch1-induced T cell leukemia requires pre-TCR signaling, as recipients of ICN1-transduced BM from SLP-76−/− or Rag-2−/− (but not Rag-2−/− × TCRβ) mice failed to develop malignancies and remained healthy for longer than 1 yr after BMT. In the presence of pre-TCR signaling, however, all mice developed fatal T cell leukemias. Together, these data suggest that among hematopoietic lineages, Notch1-transforming activity is limited to immature T cells with productive TCRβ rearrangements.
These studies clearly show that Notch1-induced T cell commitment is separate from the TCRβ-induced proliferative burst associated with the development of early DP thymocytes, and are consistent with an instructive role for Notch1 signaling in T cell commitment. Supporting this model, conditional Notch1−/− mice show that Notch1 is required for the earliest stage of T lineage commitment 9 26. Of note, another gene required for early T cell development, HES1, is a known target of activated Notch1 27. Although Notch appears to play an instructive role in early T cell development, it is difficult to rule out influences of Notch signaling on cell survival and/or proliferation. In fact, Notch signaling is associated with transcriptional upregulation of several genes among DN thymocytes including pre-Tα, CD25, Meltrin, Ifi-204, Hes1, and Deltex 28. In addition, Notch upregulates Bcl2 expression in some T cell hybridoma lines 29 and suppresses E2A activity 10 30, and both genes function at early stages of T cell development. Thus, because Notch1 appears to alter the expression of a wide array of genes with unrelated functions, one possibility is that Notch signaling integrates multiple aspects of early T cell differentiation including lineage commitment, proliferation, and survival.
Notch signaling was unable to rescue DP T cell development in Rag-2– or SLP-76–deficient donors, both of which exhibit a complete block in pre-TCR signaling and a developmental arrest before the DP stage. These findings contrast with reports demonstrating that constitutive lck or ras signals complement defects in pre-TCR signaling 19 20 21. Thus, Notch1 signals do not result in sufficient activation of these pathways to rescue the Rag-2−/− block. Interestingly, it has been reported that Notch might suppress ras signaling 23 30. We do not, however, see clear evidence for such an effect in early DP T cell development, as inhibition of ras signaling would be expected to suppress DP T cell development 31, yet wild-type ICN1-transduced progenitors give rise to abundant numbers of extrathymic DP T cells. This is consistent with a model in which Notch signaling promotes T cell commitment from early progenitors; however, expansion and further differentiation of this pool is dependent on signals from the pre-TCR that are sufficient to overcome Notch inhibitory effects on the ras/MAP kinase signaling pathway. The inability of Notch1 signals to induce DP development from Rag-2−/− progenitors also contrasts with studies in which p53−/− × Rag-2−/− mice exhibit significant numbers of DP thymocytes 32. However, these cells likely result from the absence of p53-dependent apoptotic mechanisms, and thus bear no obvious connection with the ability of Notch1 signals to drive DP T cell development from earlier progenitor populations.
Our results also show that the oncogenic potential of Notch requires T cell–specific signals for leukemic transformation. Thus, recipients of ICN1-transduced Rag-2−/− HSCs remained healthy, whereas, recipients of ICN-transduced Rag-2−/− × TCRβ cells developed an aggressive T cell leukemia that was indistinguishable in both latency and phenotype to recipients of wild-type ICN1-transduced progenitors. This suggests that the additional genetic events required for Notch-induced leukemic transformation are either initiated or revealed by TCRβ-induced pre-TCR expression, rather than RAG-induced illegitimate recombination. A recent study suggests that ras signaling cooperates with Notch4 to transform murine breast cancer cell lines 33. It is possible that similar pathways activated by TCR signaling synergize with Notch to promote both T cell expansion and leukemic transformation.
Although unlikely to be oncogenic by itself, TCRβ signaling provides a permissive environment for ICN1-mediated T cell transformation. This contrasts with other mouse models for T cell malignancy. For instance, p53−/− mice develop thymic lymphomas with no dependence on active VβDβJβ recombination 34. Likewise, initial studies on thymoma incidence in ATM−/− mice suggested that tumor formation in these mice resulted from the inability to repair double-stranded breaks resulting from V(D)J rearrangements, as thymoma incidence was significantly reduced in both Rag-2−/− and Rag-2−/− × TCRβ mice 35. However, a more recent report indicates that thymomas also develop in ATM−/− mice (albeit with longer latencies) via a Rag-independent mechanism 36. Thus, the dependence of Notch1-induced leukemia on pre-TCR signaling and DP T cell development is unique among current models of T lymphoblastic malignancy.
Although the mechanisms underlying Notch1-mediated T cell leukemia remain to be elucidated, it is likely that additional Rag-1/2–independent genetic abnormalities outside the TCRβ locus play a significant role in this process. It was recently noted that chromosome abnormalities in thymoma prone ATM−/− mice are detectable in premalignant T cells 36. It would be of interest to determine whether similar aberrations can be detected within ICN1-induced resting DP or even CD44+CD25+ or CD44− CD25+ BM populations, or if recurrent chromosomal aberrations can be identified in fatal Notch1-induced leukemias. Regardless, our studies emphasize that developing T cells are uniquely sensitive to Notch-induced transformation. Our in vivo model should be useful to dissect both the Notch-induced signals required for T cell commitment and understanding the TCRβ-induced secondary genetic events that cause malignant transformation, and raise the possibility that interfering with TCR signaling may be of therapeutic benefit in treating some types of leukemia.
Acknowledgments
We thank Tom Kadesch and members of the Pear Lab for helpful comments and critical reading of the manuscript. We thank the members of the University of Pennsylvania John Morgan (Institute for Human Gene Therapy) mouse and flow cytometry facilities. The flow cytometry studies were performed in the University of Pennsylvania Cancer Center Flow Cytometry and Cell Sorting Shared Resource (supported in part by the Lucille B. Markey Trust and the National Institutes of Health).
Supported by grants from the National Institutes of Health to G.A. Koretzky, J.C. Aster, and W.S. Pear, and the National Science Foundation (NSF-RUI no. MCB-9728332) to J. Punt. W.S. Pear is a Scholar of the Leukemia and Lymphoma Society.
References
- Artavanis-Tsakonas S., Rand M.D., Lake R.J. Notch signalingcell fate control and signal integration in development. Science. 1999;284:770–776. doi: 10.1126/science.284.5415.770. [DOI] [PubMed] [Google Scholar]
- Weinmaster G. Notch signal transductiona real rip and more. Curr. Opin. Genet. Dev. 2000;10:363–369. doi: 10.1016/s0959-437x(00)00097-6. [DOI] [PubMed] [Google Scholar]
- Mumm J.S., Kopan R. Notch signalingfrom the outside in. Dev. Biol. 2000;228:151–165. doi: 10.1006/dbio.2000.9960. [DOI] [PubMed] [Google Scholar]
- Wu L., Aster J.C., Blacklow S.C., Lake R., Artavanis-Tsakonas S., Griffin J.D. MAML1, a human homologue of drosophila mastermind, is a transcriptional co-activator for NOTCH receptors. Nat. Genet. 2000;26:484–489. doi: 10.1038/82644. [DOI] [PubMed] [Google Scholar]
- Kurooka H., Honjo T. Functional interaction between the mouse notch1 intracellular region and histone acetyltransferases PCAF and GCN5. J. Biol. Chem. 2000;275:17211–17220. doi: 10.1074/jbc.M000909200. [DOI] [PubMed] [Google Scholar]
- Zhou S., Fujimuro M., Hsieh J.J., Chen L., Miyamoto A., Weinmaster G., Hayward S.D. SKIP, a CBF1-associated protein, interacts with the ankyrin repeat domain of NotchIC to facilitate NotchIC function. Mol. Cell. Biol. 2000;20:2400–2410. doi: 10.1128/mcb.20.7.2400-2410.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsuno K., Eastman D., Mitsiades T., Quinn A.M., Carcanciu M.L., Ordentlich P., Kadesch T., Artavanis-Tsakonas S. Human deltex is a conserved regulator of Notch signalling. Nat. Genet. 1998;19:74–78. doi: 10.1038/ng0598-74. [DOI] [PubMed] [Google Scholar]
- Osborne B., Miele L. Notch and the immune system. Immunity. 1999;11:653–663. doi: 10.1016/s1074-7613(00)80140-5. [DOI] [PubMed] [Google Scholar]
- Radtke F., Wilson A., Stark G., Bauer M., van Meerwijk J., MacDonald H.R., Aguet M. Deficient T cell fate specification in mice with an induced inactivation of Notch1. Immunity. 1999;10:547–558. doi: 10.1016/s1074-7613(00)80054-0. [DOI] [PubMed] [Google Scholar]
- Pui J.C., Allman D., Xu L., DeRocco S., Karnell F.G., Bakkour S., Lee J.Y., Kadesch T., Hardy R.R., Aster J.C., Pear W.S. Notch1 expression in early lymphopoiesis influences B versus T lineage determination. Immunity. 1999;11:299–308. doi: 10.1016/s1074-7613(00)80105-3. [DOI] [PubMed] [Google Scholar]
- Haks M.C., Oosterwegel M.A., Blom B., Spits H.M., Kruisbeek A.M. Cell-fate decisions in early T cell developmentregulation by cytokine receptors and the pre-TCR. Semin. Immunol. 1999;11:23–37. doi: 10.1006/smim.1998.0153. [DOI] [PubMed] [Google Scholar]
- Clements J.L., Yang B., Ross-Barta S.E., Eliason S.L., Hrstka R.F., Williamson R.A., Koretzky G.A. Requirement for the leukocyte-specific adapter protein SLP-76 for normal T cell development. Science. 1998;281:416–419. doi: 10.1126/science.281.5375.416. [DOI] [PubMed] [Google Scholar]
- Mombaerts P., Iacomini J., Johnson R.S., Herrup K., Tonegawa S., Papaioannou V.E. RAG-1-deficient mice have no mature B and T lymphocytes. Cell. 1992;68:869–877. doi: 10.1016/0092-8674(92)90030-g. [DOI] [PubMed] [Google Scholar]
- Pivniouk V., Tsitsikov E., Swinton P., Rathbun G., Alt F.W., Geha R.S. Impaired viability and profound block in thymocyte development in mice lacking the adapter protein SLP-76. Cell. 1998;94:229–238. doi: 10.1016/s0092-8674(00)81422-1. [DOI] [PubMed] [Google Scholar]
- Shinkai Y., Rathbun G., Lam K.P., Oltz E.M., Stewart V., Mendelsohn M., Charron J., Datta M., Young F., Stall A.M. RAG-2-deficient mice lack mature lymphocytes owing to inability to initiate V(D)J rearrangement. Cell. 1992;68:855–867. doi: 10.1016/0092-8674(92)90029-c. [DOI] [PubMed] [Google Scholar]
- Aster J.C., Xu L., Karnell F.G., Patriub V., Pui J.C., Pear W.S. Essential roles for ankyrin repeat and transactivation domains in induction of T-cell leukemia by Notch1. Mol. Cell. Biol. 2000;20:7505–7515. doi: 10.1128/mcb.20.20.7505-7515.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y.S., Hayakawa K., Hardy R.R. The regulated expression of B lineage associated genes during B cell differentiation in bone marrow and fetal liver. J. Exp. Med. 1993;178:951–960. doi: 10.1084/jem.178.3.951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carleton M., Ruetsch N.R., Berger M.A., Rhodes M., Kaptik S., Wiest D.L. Signals transduced by CD3epsilon, but not by surface pre-TCR complexes, are able to induce maturation of an early thymic lymphoma in vitro. J. Immunol. 1999;163:2576–2585. [PubMed] [Google Scholar]
- Mombaerts P., Anderson S.J., Perlmutter R.M., Mak T.W., Tonegawa S. An activated lck transgene promotes thymocyte development in RAG-1 mutant mice. Immunity. 1994;1:261–267. doi: 10.1016/1074-7613(94)90077-9. [DOI] [PubMed] [Google Scholar]
- Fehling H.J., Iritani B.M., Krotkova A., Forbush K.A., Laplace C., Perlmutter R.M., von Boehmer H. Restoration of thymopoiesis in pT alpha−/− mice by anti-CD3epsilon antibody treatment or with transgenes encoding activated Lck or tailless pT alpha. Immunity. 1997;6:703–714. doi: 10.1016/s1074-7613(00)80446-x. [DOI] [PubMed] [Google Scholar]
- Iritani B.M., Alberola-Ila J., Forbush K.A., Perlmutter R.M. Distinct signals mediate maturation and allelic exclusion in lymphocyte progenitors. Immunity. 1999;10:713–722. doi: 10.1016/s1074-7613(00)80070-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinkai Y., Koyasu S., Nakayama K., Murphy K.M., Loh D.Y., Reinherz E.L., Alt F.W. Restoration of T cell development in RAG-2-deficient mice by functional TCR transgenes. Science. 1993;259:822–825. doi: 10.1126/science.8430336. [DOI] [PubMed] [Google Scholar]
- Izon D.J., Punt J.A., Xu L., Karnell F.G., Allman D., Myung P.S., Boerth N.J., Pui J.C., Koretzky G.A., Pear W.S. Notch1 regulates maturation of CD4+ and CD8+ thymocytes by modulating TCR signal strength. Immunity. 2001;14:253–264. doi: 10.1016/s1074-7613(01)00107-8. [DOI] [PubMed] [Google Scholar]
- Berger M.A., Dave V., Rhodes M.R., Bosma G.C., Bosma M.J., Kappes D.J., Wiest D.L. Subunit composition of pre-T cell receptor complexes expressed by primary thymocytesCD3 delta is physically associated but not functionally required. J. Exp. Med. 1997;186:1461–1467. doi: 10.1084/jem.186.9.1461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pear W.S., Aster J.C., Scott M.L., Hasserjian R.P., Soffer B., Sklar J., Baltimore D. Exclusive development of T cell neoplasms in mice transplanted with bone marrow expressing activated Notch alleles. J. Exp. Med. 1996;183:2283–2291. doi: 10.1084/jem.183.5.2283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomita K., Hattori M., Nakamura E., Nakanishi S., Minato N., Kageyama R. The bHLH gene Hes1 is essential for expansion of early T cell precursors. Genes Dev. 1999;13:1203–1210. doi: 10.1101/gad.13.9.1203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarriault S., Brou C., Logeat F., Schroeter E.H., Kopan R., Israel A. Signalling downstream of activated mammalian Notch. Nature. 1995;377:355–358. doi: 10.1038/377355a0. [DOI] [PubMed] [Google Scholar]
- Deftos M.L., Huang E., Ojala E.W., Forbush K.A., Bevan M.J. Notch1 signaling promotes the maturation of CD4 and CD8 SP thymocytes. Immunity. 2000;13:73–84. doi: 10.1016/s1074-7613(00)00009-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deftos M.L., He Y.W., Ojala E.W., Bevan M.J. Correlating notch signaling with thymocyte maturation. Immunity. 1998;9:777–786. doi: 10.1016/s1074-7613(00)80643-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ordentlich P., Lin A., Shen C.P., Blaumueller C., Matsuno K., Artavanis-Tsakonas S., Kadesch T. Notch inhibition of E47 supports the existence of a novel signaling pathway. Mol. Cell. Biol. 1998;18:2230–2239. doi: 10.1128/mcb.18.4.2230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gartner F., Alt F.W., Monroe R., Chu M., Sleckman B.P., Davidson L., Swat W. Immature thymocytes employ distinct signaling pathways for allelic exclusion versus differentiation and expansion. Immunity. 1999;10:537–546. doi: 10.1016/s1074-7613(00)80053-9. [DOI] [PubMed] [Google Scholar]
- Haks M.C., Krimpenfort P., van den Brakel J.H., Kruisbeek A.M. Pre-TCR signaling and inactivation of p53 induces crucial cell survival pathways in pre-T cells. Immunity. 1999;11:91–101. doi: 10.1016/s1074-7613(00)80084-9. [DOI] [PubMed] [Google Scholar]
- Fitzgerald K., Harrington A., Leder P. Ras pathway signals are required for notch-mediated oncogenesis. Oncogene. 2000;19:4191–4198. doi: 10.1038/sj.onc.1203766. [DOI] [PubMed] [Google Scholar]
- Liao M.J., Zhang X.X., Hill R., Gao J., Qumsiyeh M.B., Nichols W., Van Dyke T. No requirement for V(D)J recombination in p53-deficient thymic lymphoma. Mol. Cell. Biol. 1998;18:3495–3501. doi: 10.1128/mcb.18.6.3495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao M.J., van Dyke T. Critical role for Atm in suppressing V(D)J recombination-driven thymic lymphoma. Genes Dev. 1999;13:1246–1250. doi: 10.1101/gad.13.10.1246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petiniot L.K., Weaver Z., Barlow C., Shen R., Eckhaus M., Steinberg S.M., Ried T., Wynshaw-Boris A., Hodes R.J. Recombinase-activating gene (RAG) 2-mediated V(D)J recombination is not essential for tumorigenesis in Atm-deficient mice. Proc. Natl. Acad. Sci. USA. 2000;97:6664–6669. doi: 10.1073/pnas.97.12.6664. [DOI] [PMC free article] [PubMed] [Google Scholar]