

Abstract
Natural killer (NK) cells and dendritic cells (DCs) are two distinct cell types of innate immunity. It is known that the in vitro interaction of human NK cells with autologous DCs results in DC lysis. Here we show that contact-dependent interactions between activated human NK cells and immature DCs (iDCs) provides a “control switch” for the immune system. At low NK/DC ratios, this interaction dramatically amplifies DC responses, whereas at high ratios it completely turns off their responses. Specifically, culture of activated human NK cells with iDCs, at low NK/DC ratios (1:5), led to exponential increases in DC cytokine production, which were completely dependent on cell-to-cell contact. DC maturation was also driven by cognate interactions with NK cells and maturation was dependent on endogenously produced TNF-α in the culture. At slightly higher NK/DC ratios (5:1), inhibition of DC functions was the dominant feature due to potent killing by the autologous NK cells. Resting NK cells also stimulated autologous DC maturation in a TNF-α/contact-dependent manner, however, increasing the NK/DC ratio only led to an enhancement of this effect.
Keywords: NK cells, DCs, innate immunity, cytokines, CD86
Introduction
The innate immune response faces two mutually exclusive priorities. One is to respond to pathogenic threats with maximum speed and potency to initiate the immune response and mount the first line of defense against an infection. Competing with this goal is the requirement of controlling the initiation/amplification process to avoid overshoot and responses to benign stimuli. Failure to find a compromise between these two extremes can result in uncontrolled infection, for the former, or immune-mediated pathologies (e.g., septic shock or auto-immunity), for the latter. Cooperation between distinct immune cells and reciprocal coordination of their responses provides a robust mechanism for regulating the initiation and amplification of an immune response. NK cells and DCs are two specialized cell types of innate immunity and both act as liaisons between innate and antigen-specific responses (1, 2). DCs are considered sentinels of the immune system because of their role as initiators of both innate and antigen-specific immunity (1, 3). NK cells are lymphocytes active in innate responses against viruses, bacteria, and tumors, due to their potent cytotoxic activity and rapid production of cytokines (4). Although NK cells are part of innate immunity, they share many phenotypic and functional features with T cells (5–7). T cells cooperate with DCs in the initiation/amplification of antigen-specific responses (1, 3). We reasoned that NK cells may perform an analogous function during the initial phase of an immune response before activated T cells are generated. To address this hypothesis we developed an in vitro model to dissect the potential interactions between autologous NK cells and immature DCs (iDCs)* in the presence or absence of limiting concentrations of a bacterial stimulus (LPS). Using this culture system we have uncovered a new amplification/attenuation mechanism relevant to the regulation of innate immune responses in vivo.
Materials and Methods
Antibodies and Blocking Reagents.
Antibodies used for flow cytometry were: anti-CD86-FITC, -CD83-PE, -HLA-DR-PerCP, -CD69-FITC or -PE, -CD25-FITC, -CD3-FITC or -PE, -CD19-FITC, -CD14-FITC, -CD1a-FITC, -CD80-FITC or -PE, and -CD56-FITC or -PE (Becton Dickinson). Staining of cells was performed according to standard protocols and flow cytometric analysis was performed using a FACSCalibur™ cytometer. Antibodies used for magnetic activated cell sorting (MACS®) and subset purifications were: anti-CD3, anti-CD14, and anti-CD19 (Becton Dickinson). Blocking/neutralizing reagents used in different experiments were: anti–IFN-α (mAb MMHA-1 and sheep polyclonal anti–human; Ancell); anti–IFN-β (sheep polyclonal anti–human; Biosource International); r-hu-fas-ligand/Fc chimera (CD178; R&D Systems); hu-CTLA-4 muIg fusion protein (CD152; Ancell); anti–LFA-1 (CD11a, mAb G43–25B; Becton Dickinson); anti–TNF-α (mAb B154.2); anti–IL-12 p40 (mAb C8.6); and anti–IFN-γ (mAb B133.3) (all provided by G. Trinchieri, Schering Plough Corp., Lyon, France); humanized anti-CD40 (mAb 5H7; supplied by K. Chu, Chiron Corp., Emeryville, CA). All neutralizing antibodies and reagents were used at, at least, two different concentrations which were equal to and exceeded the manufacturers' instructions or their previously published levels for neutralization.
Cell Preparations and Cultures.
PBMCs were prepared from healthy blood donors (buffy coats or lab volunteers) by separation on a Ficoll-Hypaque gradient. iDCs were generated as described by Sallusto and Lanzavecchia (8). Briefly, highly purified (>98%) monocyte populations were obtained from PBMCs by MACS® (Miltenyi Biotec), using a one step positive selection of CD14+ cells with anti-CD14–coated magnetic microbeads (Miltenyi Biotec). The monocytes were then cultured for 5–6 d in RPMI 1640 medium (Life Technologies) supplemented with 10% FCS (Hyclone); IL-4 (10% of supernatant from an IL-4–secreting cell line, provided by A. Lanzavecchia, Institute for Research in Biomedicine, Bellinzona, Switzerland) and 50 ng/ml of GM-CSF (Leucomax®). After culture the iDC populations were routinely checked for CD1a and CD80 expression and their lack of CD14 expression. Purified (85–95% CD56+/CD3−), resting NK cells were obtained from PBMCs by MACS® (Miltenyi Biotec) using a two step protocol: PBMCs were first incubated with purified anti-CD3, anti-CD14, and anti-CD19 mAb's (Becton Dickinson) for 30 min followed by negative selection with goat anti–mouse microbeads (Miltenyi Biotec). After which the NK cells (which contained <1% contamination with CD3+, CD19+, and CD14+ cells but between 5–15% lineage negative cells) were frozen in 90% FCS; 10% DMSO for 5–6 d. Prior to their use in experiments with cultured, autologous iDCs, they were gently thawed, washed, and resuspended in RPMI 1640/10% FCS medium (RPMIc). Purified (99% CD56+/CD3−), activated NK cells were obtained using the method of Perussia et al. with slight modification (9). Briefly, PBMCs were cultured with the irradiated (5,000 rads) EBV-transformed B cell line, RPMI 8866, in 24-well plates at a ratio of 5:1 (PBMC:8866). For experiments with buffy coats the PBMCs were cultured for 5–6 d, whereas for PBMCs from lab volunteers who donated blood twice (first for NK cell cultures and then for iDC cultures), they were cultured for 8 d under the published conditions (9). After culture, purified NK cells were obtained by negative selection by MACS® (Miltenyi Biotec) as described above for resting NK cells.
NK–iDC Cultures.
The specific details of the different experiments are described in the figure legends. In general, activated/cultured or resting NK cells were added to autologous iDCs at the ratios (NK/DC) indicated and the cells were cocultured in RPMIc medium for 24–48 h in the presence or absence of the indicated doses of LPS (Sigma-Aldrich). Prior to coculture, the iDCs were removed from their GM-CSF/IL-4 containing medium by washing with RPMI 1640 medium. Control experiments comparing NK activation or iDC maturation in the presence or absence of the DC culture medium (GM-CSF/IL-4) showed no differences.
Measurement of Cytokine Production.
Cytokine production was quantified in culture supernatants by specific sandwich ELISA's for TNF-α (capture/secondary; mAb's B154.9/B154.7), IL-12 p40 (mAb's C11.79/C8.6), and IL-10 (mAb's JES3–907/JES3–12G8). The antibodies for TNF-α and IL-12 p40 ELISA were provided by G. Trinchieri and those for IL-10 were obtained from BD PharMingen. Recombinant standards for all assays were obtained from R&D Systems.
Cell-mediated Cytotoxicity Assays.
NK cell killing of iDCs or K562 was determined using standard 4-h chromium release assays and a flow cytometric assay for NK cell killing developed by McGinnes et al. with slight modifications (10). Briefly, iDCs or K562 cells were loaded with 5 μM CFDA SE for 5 or 15 min, respectively, and incubated in 96-well plates (U-bottom; Costar) with activated/cultured NK cells at the indicated NK/target ratios. After the indicated times (4 or 24 h) each sample was resuspended in a final volume of 600 μl of PBS, to which 50 μl of Flow Check fluorospheres (Beckman Coulter) were added. Using the cytometer, a fixed number (700) of fluorescent beads was acquired and the percentage of killing was calculated using the following equation: (the number of CFDA SE+ cells acquired in the control without NK cells) − (the number of CFDA SE+ cells in the sample)/(the sample of CFDA SE+ cells in the control) × 100. Comparison of 4-h chromium release assays with the 4-h flow cytometric procedure revealed minor differences with the flow cytometric technique being more sensitive (unpublished data).
NK Cell Conjugate Formation Assays.
NK–iDC or NK-K562 conjugates were detected by flow cytometry using NK cells labeled with CFDA SE (Molecular Probes) and iDCs or K562 labeled with SNARF-1 (Molecular Probes). After dye loading (NK cells and K562, 15 min at 37°C with 5 μM of CFDA SE or 2 μM SNARF-1, respectively, and iDCs, 5 min at 37°C with 2 μM of SNARF-1) the cells were washed and incubated alone or together (NK–K562 or NK–iDC) at the indicated ratios for 30 min at 37°C. Prior to incubation the cells were centrifuged at 200 g for 3 min at room temperature. The percentage of conjugates formed was calculated on a FACSCalibur™ cytometer (Becton Dickinson) by gating on the FL3+ cells (SNARF-1/iDCs or K562) and determining the percent of this population also positive for FL1 (CFDA SE/NK).
Results
Contact-dependent and Reciprocal Activation of NK Cells and iDCs.
We began our study by investigating the interactions between autologous iDCs and resting NK cells. These initial experiments demonstrated that both cell types become activated after coculture for 18–48 h. The iDCs showed evidence of maturation as determined by their increased expression of CD86 (Fig. 1 A), HLA-DR and CD83 (data not shown) which was augmented by the addition of increasing concentrations of LPS. The NK effect was dependent on cell-to-cell contact as separation of the cells by a permeable membrane (trans-well) completely abrogated DC maturation. Addition of a neutralizing antibody against TNF-α also blocked NK-mediated maturation of the iDC (Fig. 1 B). By contrast, the addition of other blocking reagents specific for IL-12, IFN-γ, IFN-α, IFN-β, fas (CD95), ICAM-3 (CD50), CD40L (CD154), CD80, CD86, and LFA-1 (CD11a) to the cultures did not inhibit maturation (data not shown). Raising the NK/DC ratio consistently led to increased maturation of the DCs (Fig. 1 C).
Figure 1.

Reciprocal stimulation of resting NK cells and autologous iDCs. Cultured iDCs and resting, purified NK cells were incubated alone or in combination for 18 h (E–G) or 48 h (A–D). After culture the cells were analyzed by flow cytometry for the expression of CD86 (DCs; A–C), CD69 (NK; D) or CD69 and CD25 (NK; E–G). Specific analysis of NK cell or DC subsets was achieved by a combination of gating on forward versus side scatter differences and staining with antibodies for CD56 (NK) or CD86 (DC). (A) iDCs were incubated alone (▪) or with NK cells (□) at a ratio of 5:1 (NK/DC) in the presence of the indicated doses of LPS. The percentage of CD86bright DC was determined by specific gating. (B) The representative experiment shown was performed in the presence of 6.25 ng/ml of LPS and the iDCs were incubated alone (gray bar); with NK cells at a 5:1 (NK/DC) ratio (black bar); with NK cells (5:1) plus 10 μg/ml of a neutralizing antibody for TNF-α (white bar); or with NK cells (5:1) but separated in trans-wells (striped bar). Changes in DC maturation were determined by measuring CD86 mean fluorescence intensity (MFI). (C) iDCs were incubated alone (DCs) or with NK cells at the indicated ratios (NK/DC) in the presence (▴) or absence (▵) of 6.25 ng/ml of LPS. DC maturation was determined as in A. (D) NK cells were incubated for 48 h either alone (NK) or with iDCs at the indicated ratios (NK/DC) in the presence (▴) or absence (▵) of 10 ng/ml of LPS. NK cell activation was determined by the percentage of cells expressing CD69. (E–G) Flow cytometry dot plots from a representative experiment measuring NK cell activation marker expression (CD69 and CD25). NK cells were incubated for 18 h: alone (E), with iDCs at an NK/DC ratio of 1:5 (F), or with iDCs (NK/DC, 1:5), but separated in trans-wells (G).
Under the same conditions, NK cells were also stimulated by the iDCs and upregulated their expression of the activation markers CD69 (Fig. 1 D) and CD25 (data not shown), however, the addition of LPS to the cultures did not enhance this activation. Unlike DC maturation, increasing the NK/DC ratio in favor of the NK cells led to decreased expression of these activation markers, and at the highest NK/DC ratios tested, the expression of CD69 returned to baseline. The activation of NK cells observed in these cultures was dependent on contact with the iDCs (Fig. 1 E–G). The blocking reagents used in the previous experiments on DC maturation also had no effect on iDC-mediated NK cell activation, and neither did a neutralizing antibody to TNF-α (data not shown). Thus, contact-dependent interactions between NK cells and iDCs could provide a novel positive feedback loop for the innate immune response which is reciprocally regulated by the number of NK–DC encounters.
Stimulation/Inhibition of iDC Functions by Activated NK Cells.
Two previous studies have shown that both resting and activated NK cells partially kill autologous DCs, as measured by 3–4-h chromium release assays using high NK/DC ratios (11, 12). However, in our long-term cultures (24–48 h) the NK cells stimulated the iDCs rather than lysing them. Based on these conflicting results, we examined in more detail the apparent stimulation of iDCs after contact with NK cells. We reasoned that activated NK cells would be more useful for these experiments because (i) an encounter in vivo between NK cells that normally traffic in the blood (4) and iDCs which reside in the tissues (1, 3), should only occur when both cells are activated; (ii) activated NK cells mediate more potent lysis of iDCs (11–13), thereby allowing for a more rigorous examination of a potential cytotoxic effect; and (iii) the number of NK cells obtained after short-term in vitro culture would be far greater providing more cells for analysis (9).
The DC maturation induced by resting NK cells we observed was dependent on endogenous TNF-α production. Furthermore, TNF-α and other proinflammatory cytokines are important amplifiers of immune responses (14). Therefore, we used TNF-α production as a readout for DC stimulation in our culture system. Culturing activated NK cells with DCs at low NK/DC ratios (1:5) led to increases in TNF-α production, which were augmented dramatically by the addition of suboptimal doses (10 ng/ml) of LPS (Fig. 2 A). The increased TNF-α production was dependent on cell-to-cell contact because when the two cell types were separated in trans-wells, no stimulation was observed (Fig. 2 B). In separate experiments we observed this NK-stimulatory effect on TNF-α production at NK/DC ratios as low as 1:20 and 1:40 (data not shown). Raising the NK/DC ratio to 5:1 changes the interaction from one of potent DC stimulation to that of complete inhibition in the presence or absence of LPS (Fig. 2 C). An identical pattern is seen for IL-12 p40 production (Fig. 2 D). No IL-10 production in the cultures was detected under any of the conditions tested (data not shown). The massive production of TNF-α in the cultures at the lowest ratios was mediated primarily by DCs in the culture due to the low number of NK cells (2 × 104 cells per well) used in these experiments. In addition, efforts to stimulate NK cells directly with optimal doses of anti-CD16, rIL-2, rIL-12, and PMA/ionomycin alone or in combination induced only moderate (<500 pg/ml) TNF-α production by the NK cells alone which were present at 2 × 105 cells per well (data not shown).
Figure 2.

Activated/cultured NK cell–mediated amplification/inhibition of DC cytokine production. Results are presented on log scales. Cultured iDCs were incubated alone or with cultured/purified NK cells for 24 h (A and B) or 48 h (C–E) in the presence or absence of the indicated concentrations of LPS. After culture the supernatants were removed and analyzed by ELISA for TNF-α or IL-12 p40. (A) TNF-α production was measured in the supernatants from cultures of: DCs alone (gray bars); NK+DC (1:5) (black bars). LPS concentrations used were: 0, 10, and 1,000 ng/ml (DCs alone only). (B) Contact-dependent stimulation of iDC TNF-α production by NK cells was tested under the following conditions: DCs alone (gray bars); NK+DC (1:5) (black bars); NK/DC (1:5) trans-wells (striped bars). LPS concentrations used were: 10 ng/ml (donor 1) and 20 ng/ml (donor 2). (C and D) Effect of increasing the NK/DC ratio (DCs alone; NK:DC-1:5; 1:1; 5:1) on (C) TNF-α and (D) IL-12 p40 production. LPS concentrations used were: 0 ng/ml (▪); 20 ng/ml (•); 50 ng/ml (▴); and 1,000 ng/ml-LPS (□) (DCs alone only). (E) The effect of neutralizing IFN-γ production was tested under the following conditions: DCs alone (gray bars); NK+DC (1:5) (black bars); NK+DC (1:5) and 10 μg/ml of blocking anti–IFN-γ mAb (stippled bars). LPS concentrations used were: 0 ng/ml (NO LPS) and 20 ng/ml (LPS 20).
Blocking reagents for CD80, CD86, CD154, CD95, CD11a, CD50, or IL-12, had no effect on the NK cell–induced TNF-α or IL-12 p40 production, whereas a neutralizing antibody for IFN-γ while not affecting TNF-α production partially blocked (<20%) IL-12 p40 production induced by the NK cells (Fig. 2 E). This partial inhibition is consistent with numerous previous reports demonstrating a central role for an IL-12/IFN-γ positive feedback loop active between NK cells and IL-12–producing cells (15–17). Clearly, however, this well-described cytokine cross-talk mechanism is secondary to the contact-dependent activation of DCs by NK cells described here. Taken together these results indicate that novel mechanisms control cognate interactions between NK cells and DCs and that NK–DC contact has the potential to serve as an important control switch for both amplifying and attenuating innate immune responses in vivo.
We next tested directly whether NK cell killing was active in our system using both short-term (4-h) chromium release assays (data not shown) and a flow cytometric assay which allowed us to follow the survival of the autologous iDCs in the presence of NK cells over longer periods (Fig. 3). The results demonstrate that NK cells kill autologous iDCs (Fig. 3 A) at all NK/DC ratios tested after 4 h and the level of killing is similar to the lysis of K562 cells over the same time (Fig. 3 B). After 24 h, however, the presence of NK cells led to completely divergent effects on the DCs. At the 5:1 (NK/DC) ratio, NK cell–mediated destruction of the iDCs was the dominant feature, whereas at the low ratio (1:5) the same NK cells promoted iDCs survival compared with iDCs alone over the same period (Fig. 3 A). The equal ratio of NK/DC (1:1) fell between these two extremes. The NK cell–mediated lysis of the K562 targets increased at all ratios tested after the 24-h incubation. These striking differences between death and survival of DCs upon encountering NK cells correlate precisely with our previous results on TNF-α and IL-12 p40 production where increasing numbers of NK cells in the culture shifts the NK effect from potent stimulation to complete inhibition (Fig. 2).
Figure 3.

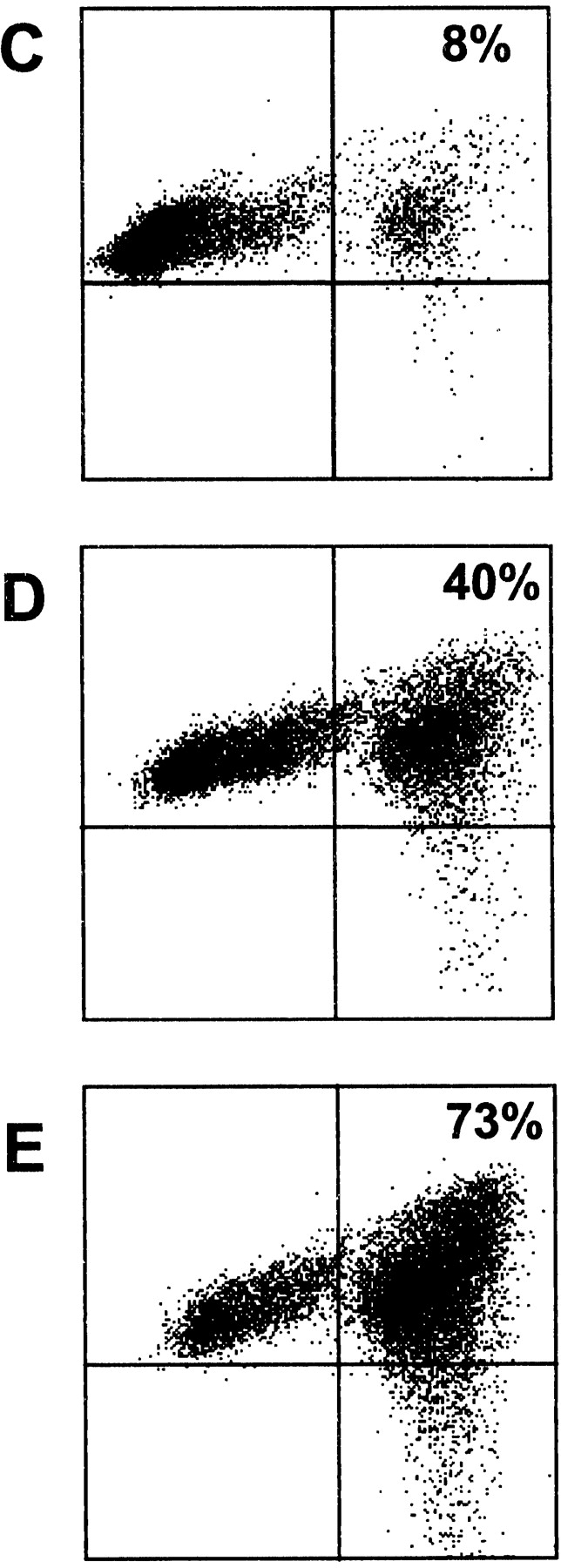
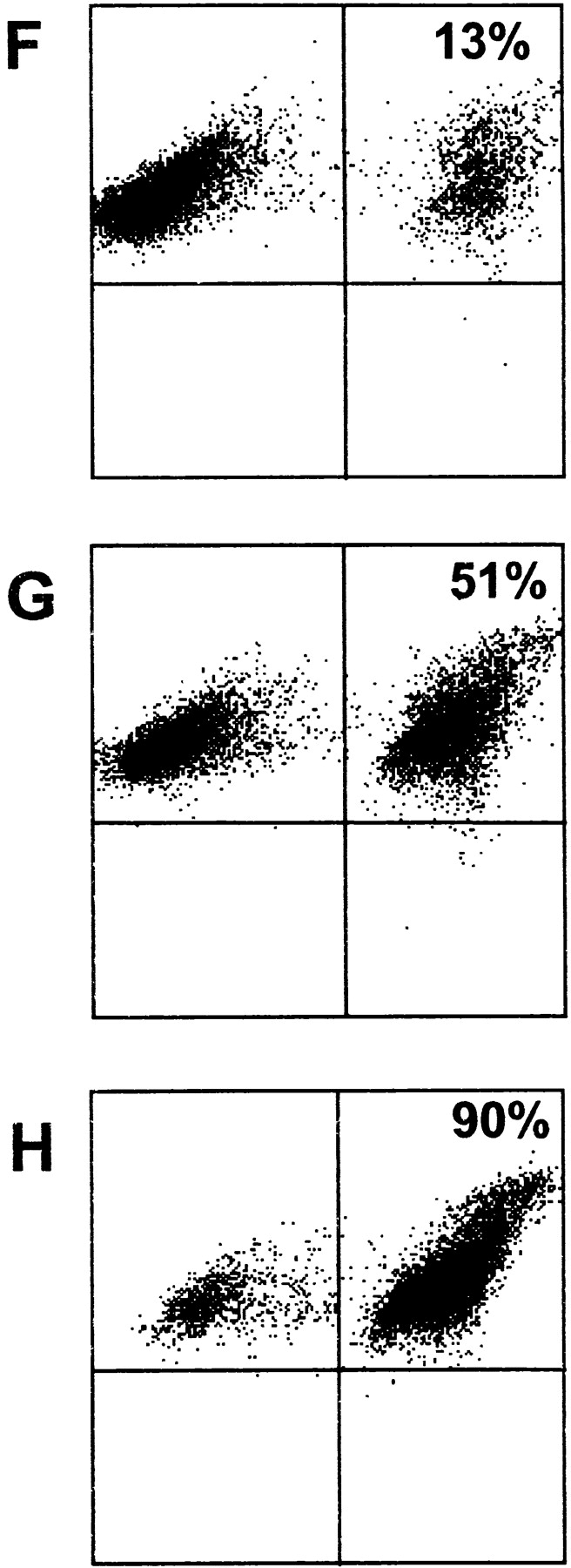
Activated/cultured NK cell–mediated death/survival of DCs and NK–DC conjugate formation. NK cell mediated killing/survival of CFDA.SE-loaded iDCs (A) and K562 cells (B) was measured by flow cytometry over 4 h (▪) or 24 h (•) at the indicated NK/DC ratios as described in Materials and Methods. Results are presented as percentage of specific lysis ± SEM (n = 3 independent experiments). NK–DC (C–E) or NK–K562 (F–H) conjugate formation was measured by flow cytometry, at the following ratios (NK/DC or NK/K562): 1:5 (C and F); 1:1 (D and G); and 5:1 (E and H). Snarf-1 (FL3)-labeled iDCs or K562 cells were incubated (30 min) with CFDA.SE (FL1)-labeled NK cells at the different ratios. The percentage of conjugates formed (top right quadrant) was calculated by determining the percentage of FL3+ (Snarf-1) events which were also positive for FL1 (CFDA.SE). The results are representative of four experiments performed with activated NK cells. Resting NK cells also readily formed conjugates with autologous iDCs (data not shown).
NK cells can both efficiently kill and potently stimulate DCs in our model system and each of these opposing effects appear dependent on cell-to-cell contact. To quantify these apparent cognate interactions we measured the formation of NK–DC conjugates and compared them to conjugates formed by NK cells and tumor targets. Our analysis revealed that NK cells bound DC (Fig. 3 C–E) with an efficiency almost equal to that of NK cell binding to the classical NK-target, K562 (Fig. 3 F–H). Binding was detectable even at the lowest NK/DC ratio tested (1:5) indicating that NK–DC conjugates are formed in our other experiments measuring cytokine production and cell killing.
Our initial experiments using resting NK cells (Fig. 1) demonstrated a contact-dependent NK effect driving iDCs to mature. Therefore, we tested what effects activated NK cells mediated on DC maturation (Fig. 4). Similar to our previous results with activated NK cells (Figs. 2 and 3), addition of NK cells to the culture leads to opposing effects on the iDC population depending on the NK/DC ratio. Compared with the untreated DCs (Fig. 4 A–C), low NK/DC ratios (1:5 or 1:1) consistently augmented expression of the maturation markers: CD86 (Fig. 4 D–I), CD83, and HLA-DR (data not shown). At a 5:1 ratio (NK/DC) most of the DCs are nonviable (Fig. 4 J–L). Addition of suboptimal doses of LPS (Fig. 4 C, F, I, and L) augmented the NK-induced maturation at NK/DC ratios of 1:5 and 1:1, but had no effect at the 5:1 ratio. Again, DC maturation was dependent on NK cell contact and endogenously produced TNF-α, because it did not occur when the two cell types were separated in trans-wells or upon the addition of a neutralizing antibody for TNF-α (Fig. 5). Moreover, addition of blocking reagents for CD80, CD86, CD154, CD95, CD11a, CD50, IL-12, IFN-γ, IFN-α, or IFN-β had no effect on the NK-induced maturation (data not shown).
Figure 4.

Activated/cultured NK cell–mediated maturation/death of DCs. Cultured iDCs were incubated for 48 h, alone (A–C), or with NK cells at the following ratios (NK/DC), 1:5 (D–F), 1:1 (G–I), and 5:1 (J–L). Forward versus side scatter plots (A, D, G, and J) show the gating on the DC subset and demonstrate its disappearance at the 5:1 (NK/DC) ratio (J). Propidium iodide staining of the NK/DC (5:1) cultures confirmed the almost complete absence of viable DCs (data not shown). Staining for CD86 expression was performed as described in Fig. 1 and the histograms are shown for DCs cultured in the presence (C, F, I, and L) or absence (B, E, H, and K) of 1 ng/ml of LPS. Results are representative of six experiments performed.
Figure 5.

Activated/cultured NK cell-induced DC maturation is dependent on cell-to-cell contact and endogenous TNF-α production. Cultured iDCs were incubated alone (A, D, and G); with activated NK cells at an NK/DC ratio of 1:5 (B, E, and H); with activated NK cells (NK/DC; 1:5) separated in trans-wells (C and F); or with activated NK cells and 10 μg/ml of a neutralizing antibody for TNF-α (I). LPS concentrations used were: 0 ng/ml (A–C); 10 ng/ml (D–F) and 50 ng/ml (G–I). CD86 staining of the DC subset was performed as in Figs. 1 and 4. Results are representative of four experiments performed.
Discussion
Cognate interactions are a common feature of the antigen-specific immune response, where intimate cell-to-cell contact governs such critical events as antigen presentation, and the delivery of T cell help to CTLs and B cells (1, 3, 18–20). By contrast, coordinated control of innate immunity is thought to be mediated almost exclusively by cytokine feedback loops (2, 17, 21). Recently, one exception to this general rule has been proposed for the innate immune system and importantly, it involves contact-dependent activation of murine NK cells by iDCs (22). Consistent with this study, we also see evidence of NK cell activation in our system which is dependent on DC contact, although we have not investigated the extent and nature of this NK activation in further detail.
NK cells arrive at sites of infection within minutes to hours after pathogen invasion (4, 21, 23). Here, they should encounter resident DCs already responding to signals derived from invading pathogens and proinflammatory cytokines such as TNF-α (1, 3). We designed our in vitro model to recapitulate some of the most salient features of the early inflammatory process. This simple model cannot reproduce many aspects of the in vivo situation where the flow of immune cells, pathogens, and soluble factors in and out of the inflamed tissue is a dynamic process. Nevertheless, using this system, we have defined two extreme outcomes of an NK–DC encounter: (i) potent amplification and (ii) complete inhibition. During a normal functioning immune response, the majority of in vivo relevance of this NK–DC “control switch” likely lies at various points between the two in vitro extremes described here. However, it is tempting to speculate that the dramatic NK-mediated stimulation/inhibition of TNF-α production we observe reveals a novel positive/negative feedback loop applicable to TNF-α–mediated pathologies such as septic shock.
The inhibition of DC responses by NK cells at high NK/DC ratios, appears simply due to NK cell–mediated cytotoxicity which has already been described as a potential outcome of NK–DC encounters (11, 12). However, with the exception of endogenous TNF-α production which is necessary for NK-induced DC maturation, all other experiments using blocking reagents against numerous surface receptors and soluble factors failed to reveal a specific mechanism for the NK cell stimulatory effect. Although we cannot rule out the possibility that untested receptor/ligand pairs govern this interaction, we propose an alternative model founded on the more general phenomenon of NK cell killing of DCs. The model is based on the hypothesis that the majority of NK–DC interactions are fatal for the DCs. Thus, amplification of DC responses occurs by a bystander effect whereby killing of DCs by NK cells provides the activation signal(s) for their nonNK-binding neighbors. At low NK/DC ratios there are plenty of viable DCs to mediate this bystander effect, whereas, at higher ratios, there are no bystanders. This model fits nicely with our current understanding of the heterogeneous nature of NK cell triggering receptors (24) and is consistent with the results presented here. Moreover, it adds a new twist to “Stranger/Danger” models (25–28) for the initiation/amplification of innate immune responses. Thus, the arrival of activated NK cells (“stranger”) into infected tissues and their killing (“danger”) of resident DCs serves to amplify the “stranger/danger” signals derived from invading pathogens. It will be important to test this model with particular emphasis placed on understanding a role (or lack thereof) for NK cell inhibitory/stimulatory receptors for autologous HLA class I molecules (24). Given the exquisite sensitivity of autologous iDCs (11, 12) to NK cell cytotoxicity, it is likely that during an NK–DC encounter the diverse families of NK cell receptors for class I function differently than they do during encounters with more traditional target cell types.
Acknowledgments
We wish to thank Rino Rappuoli, Giuseppe Del Giudice, and Antonio Lanzavecchia for stimulating discussions and their critical reading of the manuscript. A special thanks to our volunteer blood donors because without their generosity and patience many of these studies would not have been possible.
S. Sbrana's present address is Istituto di Fisiologia Clinica CNR Ospedale Pediatrico Apuano, G. Pasquinucci Via Aurelia Sud, 54100 Massa, Italy.
Footnotes
Abbreviation used in this paper: iDC, immature DC.
References
- 1.Banchereau, J., and R. Steinman. 1998. Dendritic cells and the control of immunity. Nature. 392:245–252. [DOI] [PubMed] [Google Scholar]
- 2.Scott, P., and G. Trinchieri. 1995. The role of natural killer cells in host-parasite interactions. Curr. Opin. Immunol. 7:34–40. [DOI] [PubMed] [Google Scholar]
- 3.Cella, M., F. Sallusto, and A. Lanzavecchia. 1997. Origin, maturation and antigen presenting function of dendritic cells. Curr. Opin. Immunol. 9:10–16. [DOI] [PubMed] [Google Scholar]
- 4.Trinchieri, G. 1989. Biology of natural killer cells. Adv. Immunol. 47:187–376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sanchez, M.J., M.O. Muench, M.G. Roncarolo, L.L. Lanier, and J.H. Phillips. 1994. Identification of a common T/natural killer cell progenitor in human fetal thymus. J. Exp. Med. 180:569–576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lanier, L.L., H. Spits, and J.H. Phillips. 1992. The developmental relationship between NK cells and T cells. Immunol. Today. 13:392–395. [DOI] [PubMed] [Google Scholar]
- 7.Valiante, N.M., and P. Parham. 1996. NK cells and CTL: opposite sides of the same coin. Chem. Immunol. 64:146–163. [PubMed] [Google Scholar]
- 8.Sallusto, F., and A. Lanzavecchia. 1994. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor α. J. Exp. Med. 179:1109–1118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Perussia, B., C. Ramoni, I. Anegon, M.C. Cuturi, J. Faust, and G. Trinchieri. 1987. Preferential proliferation of natural killer cells among perpheral blood mononuclear cells co-cultured with B lymphoblastoid cell lines. Nat. Immun. Cell Growth Reg. 6:171–182. [PubMed] [Google Scholar]
- 10.McGinnes, K., G. Chapman, R. Marks, and R. Penny. 1986. A fluorescence NK assay using flow cytometry. J. Immunol. Methods. 86:7–15. [DOI] [PubMed] [Google Scholar]
- 11.Carbone, E., G. Terrazzano, G. Ruggiero, D. Zanzi, A. Ottaiano, C. Manzo, K. Karre, and S. Zappacosta. 1999. Recognition of autologous dendritic cells by human NK cells. Eur. J. Immunol. 29:4022–4029. [DOI] [PubMed] [Google Scholar]
- 12.Wilson, J., L. Heffler, J. Charo, A. Scheynius, M. Bejarano, and H. Ljunggren. 1999. Targeting of human dendritic cells by autologous NK cells. J. Immunol. 1999. 163:6365–6370. [PubMed] [Google Scholar]
- 13.Phillips, J.H., and L.L. Lanier. 1986. Dissection of the lymphokine-activated killer phenomenon. J. Exp. Med. 164:814–825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Tracey, K., and A. Cerami. 1994. Tumor necrosis factor: a pleiotropic cytokine and therapeutic target. Annu. Rev. Med. 45:491–503. [DOI] [PubMed] [Google Scholar]
- 15.Scharton, T.M., and P. Scott. 1993. Natural killer cells are a source of IFN-γ that drives differentiation of CD4+ T cell subsets and induces resistance to Leishmania major of mice. J. Exp. Med. 178:567–577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Scott, P. 1993. IL-12: initiation cytokine for cell-mediated immunity. Science. 260:496–497. [DOI] [PubMed] [Google Scholar]
- 17.Trinchieri, G. 1997. Cytokines acting on or secreted by macrophages during intracellular infection (IL-10, IL-12, IFN-γ). Curr. Opin. Immunol. 9:17–23. [DOI] [PubMed] [Google Scholar]
- 18.Davis, M.M., and P.J. Bjorkman. 1988. T-cell antigen receptor genes and T-cell recognition. Nature. 334:395–402. [DOI] [PubMed] [Google Scholar]
- 19.Scherle, P., and W. Gerhard. 1986. Functional analysis of influenza-specific helper T cell clones in vivo. T cells specific for internal viral proteins provide cognate help for B cell responses to hemagglutinin. J. Exp. Med. 164:1114–1128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Krowka, J., B. Singh, A. Fotedar, T. Mossman, M. Giedlin, and L. Pilarski. 1986. A requirement for physical linkage between determinants recognized by helper molecules and cytotoxic T cell precursors in the induction of cytotoxic T cell responses. J. Immunol. 136:3561–3566. [PubMed] [Google Scholar]
- 21.Unanue, E.R. 1997. Inter-relationship among macrophages, natural killer cells and neutrophils in early stages of Listeria resistance. Curr. Opin. Immunol. 9:35–43. [DOI] [PubMed] [Google Scholar]
- 22.Fernandez, N., A. Lozier, C. Flament, P. Ricciardi-Castagnoli, D. Bellet, M. Suter, M. Perricaudet, T. Tursz, E. Maraskovsky, and L. Zitvogel. 1999. Dendritic cells directly trigger NK cell functions: cross-talk relevant in innate anti-tumor immune responses in vivo. Nat. Med. 5:405–411. [DOI] [PubMed] [Google Scholar]
- 23.Scharton-Kersten, T.M., and A. Sher. 1997. Role of natural killer cells in innate resistance to protozoan infections. Curr. Opin. Immunol. 9:44–51. [DOI] [PubMed] [Google Scholar]
- 24.Lanier, L.L. 1998. NK cell receptors. Annu. Rev. Immunol. 16:359–393. [DOI] [PubMed] [Google Scholar]
- 25.Janeway, C.J., C. Goodnow, and R. Medzhitov. 1996. Danger-pathogen on the premises! Immunological tolerance. Curr. Biol. 6:519–522. [DOI] [PubMed] [Google Scholar]
- 26.Vance, R. 2000. A Copernican Revolution? Doubts about the danger theory. J. Immunol. 165:1725–1728. [DOI] [PubMed] [Google Scholar]
- 27.Matzinger, P. 1994. Tolerance, danger, and the extended family. Annu. Rev. Immunol. 12:991–1045. [DOI] [PubMed] [Google Scholar]
- 28.Ridge, J., E. Fuchs, and P. Matzinger. 1996. Neonatal tolerance revisited: turning on newborn T cells with dendritic cells. Science. 271:1723–1726. [DOI] [PubMed] [Google Scholar]