

Abstract
Dendritic cells (DCs) activated by CD40L-expressing CD4+ T cells act as mediators of “T helper (Th)” signals for CD8+ T lymphocytes, inducing their cytotoxic function and supporting their long-term activity. Here, we show that the optimal activation of DCs, their ability to produce high levels of bioactive interleukin (IL)-12p70 and to induce Th1-type CD4+ T cells, is supported by the complementary DC-activating signals from both CD4+ and CD8+ T cells. Cord blood– or peripheral blood–isolated naive CD8+ T cells do not express CD40L, but, in contrast to naive CD4+ T cells, they are efficient producers of IFN-γ at the earliest stages of the interaction with DCs. Naive CD8+ T cells cooperate with CD40L-expressing naive CD4+ T cells in the induction of IL-12p70 in DCs, promoting the development of primary Th1-type CD4+ T cell responses. Moreover, the recognition of major histocompatibility complex class I–presented epitopes by antigen-specific CD8+ T cells results in the TNF-α– and IFN-γ–dependent increase in the activation level of DCs and in the induction of type-1 polarized mature DCs capable of producing high levels of IL-12p70 upon a subsequent CD40 ligation. The ability of class I–restricted CD8+ T cells to coactivate and polarize DCs may support the induction of Th1-type responses against class I–presented epitopes of intracellular pathogens and contact allergens, and may have therapeutical implications in cancer and chronic infections.
Keywords: T helper subsets, dendritic cells, maturation, IL-12, CD8+ T cells
Introduction
Effective cell-mediated immunity, essential for the protection against chronic intracellular infections and cancer, depends on the concordant activity of CD8+ T cells (CTL) and CD4+ T cells, especially Th1 cells (1). Dendritic cells (DCs),* the carriers of pathogen-related information within the immune system (2), are essential for the interaction between the above T cell subsets. The ability of DCs to “translate” the CD40L-dependent helper signal from CD4+ T cells into an enhanced ability to induce and support effector functions of CD8+ T cells (3–5), allows them to act as a “spatial and temporal bridge” linking the responses of “helper” CD4+ and “effector” CD8+ T cells (3). In contrast to the well established role of DCs in mediating such CD4 to CD8 helper signals, it remains unclear whether they also mediate “reverse” CD8+ to CD4+ T cell communication. CD8+ T cells, recognizing MHC class I–restricted viral epitopes, can be effectively triggered by virus-infected cells (3) and are capable of activating the Ag-carrying DCs and bystander DCs in a CD40L and TNF-related activation-induced cytokine (TRANCE)/receptor activator of nuclear factor κB (RANK)-independent mechanism (6, 7). However, it remains unknown whether the DC–CD8+ T cell interaction results in any DC-carried signals that affect the character of CD4+ T cell responses.
Here, we show that in a sharp contrast to naive CD4+ T cells, naive CD8+ T cells are capable of IFN-γ and TNF-α production at early time-points of their priming, and efficiently synergize with CD40L-expressing naive Th cells in the optimal activation of DCs and the induction of IL-12 p70, the factor playing a key role in the CTL-, Th1-, and NK cell–dependent cell-mediated immunity (8). This novel “helper” function of CD8+ T cells in the DC-mediated polarization CD4+ T cell responses may provide a reliable mechanism of inducing the desirable Th1-type responses against intracellular pathogens. This additional Th1-inducing pathway is not contingent on the pathogen's intrinsic ability to induce IL-12 (9, 10) or IFN-α (11, 12), but utilizes the ability of CD8+ T cells to detect MHC class I–presented foreign epitopes as a cue for the pathogen's intracellular character.
Materials and Methods
Media, Reagents, and Transformed Cell Lines.
All cultures were performed in IMDM (Life Technologies) with gentamycin (250 ng/ml; Life Technologies) supplemented with 10% FCS (Hyclone). RhuGM-CSF and IL-4 were gifts from Schering-Plough. Human recombinant IL-2 was a gift from the Chiron Corporation (Emeryville, CA). rhTNF and rhIFN-γ were purchased from Strathman Biotech. LPS and Staphylococcus Enterotoxin B (SEB) were purchased from Sigma-Aldrich. CD3 mAb (OKT3), used as a surrogate Ag, was purchased from Ortho-Biotech. CD3 (CLB-T3/4.E) and CD28 (CLB-CD28/1) mAbs (both from CLB) were used to induce cytokine production in T cells. CD40L-transfected J558 plasmacytoma cells (J558-CD40L; reference 13), were a gift of Dr. P. Lane, University of Birmingham, Birmingham, UK. The HLA-DR4–transfected, HLA-class I A2 expressing T2 cell line was provided by Dr. J. Blum, University of Indiana, Indianapolis, IN.
Isolation of Peripheral Blood and Cord Blood T Cell Populations.
Mononuclear cells, obtained from peripheral blood of healthy donors or from cord blood, were isolated by LymphoPrep™ (Nycomed Pharma). Naive CD4+CD45RA+ T cells, and naive and bulk populations of CD8+ T cells were isolated by negative selection with StemSep™ (StemCell Technologies Inc.). CD8+ T cells were selected using the StemSep™ CD8 T cell enrichment cocktail. The selected population contained both memory (CD45RO+) and naive (CD45RA+) CD8+ T cells and are referred to as “bulk” CD8+ T cells. Isolation of naive CD4+ T cells and naive CD8+ was performed by the pretreatment of PBMCs or cord-blood-isolated mononuclear cells with biotin-labeled anti-CD45RO mAb, followed by the incubation with customized StemSep™ naive CD4 and CD8 T cell selection cocktails containing anti-biotin mAb tetramer. Percentage of double-positive CD45RA+/CD4+ and CD45RA+/CD8+ cells in the isolated populations ranged between 95–98%, as determined by FACS®. Contamination with memory CD45RO+ T cells in either of the isolated naive T cell populations was below 1% (see Fig. 1).
Figure 1.



Cord blood– and peripheral blood–isolated CD45R0−/RA+ CD4+ and CD8+ T cells show a mutually exclusive expression of CD40L and IFN-γ. (A) Expression of CD45RA and CD45RO on peripheral blood-isolated naive CD4+ and CD8+ T cells. Similar data were obtained using cord blood–isolated cells. (B) Differential ability of naive CD4+ and CD8+ T cell populations to produce IFN-γ. Freshly-isolated CD45R0−/RA+ CD4+ and CD8+ T cells (105 cells/well) from peripheral blood (white bars) or from cord blood (black bars) were stimulated for 24 h with the stimulatory combination of CD3 and CD28 mAbs (top) or with 2 × 104 TNF-α–preactivated SEB-coated DCs (bottom). Concentrations of IFN-γ in 24 h supernatants were analyzed with specific ELISA. The data (mean ± SD) represent one experiment of three (cord blood) or five (peripheral blood) that all yielded similar results. (C) The presence of CD40L on activated T cell populations from cord blood was analyzed in T cell cultures stimulated with CD3 and CD28 mAbs for 24 h. Similar data was obtained in case of peripheral blood–isolated T cells.
Class I–restricted Melanoma-specific and Influenza-specific CD8 Cells.
The HLA-A2–restricted CD8+ CTL clone, recognizing melanoma antigen gp100 (209–217, ITDQVPFSV), was cloned from the TIL1520 cell line (14) provided by Drs. Steven Rosenberg and John Wunderlich (National Cancer Institute, Bethesda, MD). Influenza-specific CD8+ T cells, recognizing the influenza peptide (Flu M1; 58–66; GILGFVFTL) in context of HLA-A2.1, were obtained from the CD8+ T cell fraction of fresh blood by specific tetramer isolation (15). After isolation, the cells were cultured for 2 wk in the presence of IL-2 and IL-7 and stimulated with peptide-loaded autologous DCs, 10 d before use. The frequency of tetramer-binding cells was ∼50% (see Fig. 4 A).
Figure 4.

Short term-cultured class I–restricted influenza-specific CD8+ Th cells and freshly isolated naive CD8+ T cells enhance the activation status of DCs. (A) Short term cultured tetramer-isolated influenza-specific CD8+ T cells (2 × 105 cells/well; see the inset for the frequency of tetramer-binding cells) were cocultured with HLA-A2+ day 6 immature DCs in the presence of the relevant influenza peptide or irrelevant peptide (tyrosinase). (B) Day 6 DCs were cultured in the presence of SEB or OKT3 (CD3) mAb, in the absence or in the presence of bulk CD8+ T cells (2 × 105 cells/well) freshly isolated from peripheral blood. (C) Bystander activation of DCs by freshly isolated naive CD8+ T cells and gp100-specific CTL clone in the transwell system. CD8+ T cells (2 × 105 cells/well) and day 6 DCs (3 × 104 cells/well) were placed in the upper chamber in the presence of absence of Ag (SEB or gp100 peptide). Bottom chamber contained the DCs serving as a readout of bystander activation. (D) DC-activating properties of freshly isolated naive CD45RA/CD8+ T cells (2 × 105 cells/well) in the SEB model. 48 h direct cocultures.
Generation of DCs.
To obtain immature “day 6 DCs”, peripheral blood monocytes (5 × 105 cells/well) were cultured in FCS/IMDM with GM-CSF and IL-4 (both 1,000 U/ml) in 24-well plates (Costar), as described (16). At day 6, the cultures consisted uniformly of CD1a+CD14−, HLA-DR+, negative for CD83, the human DC maturation marker, and expressed intermediate levels of such costimulatory molecules as CD54 and CD86 (see Fig. 3). The number of such “day 6 DCs” varied between 2–4 × 105 cells/well, depending on the donor. In the initial set of experiments (Figs. 1 and 2 only), in addition to such “day 6 DC,” we also used the DCs that were stimulated overnight with TNF-α (50 ng/ml; Strathmann) to enhance their expression of costimulatory factors and their T cell stimulatory activity. Such cells are referred to as “TNF-α–preactivated DCs.” In the following experiments (DC activation and polarization), we used immature “day 6 DCs.”
Figure 3.
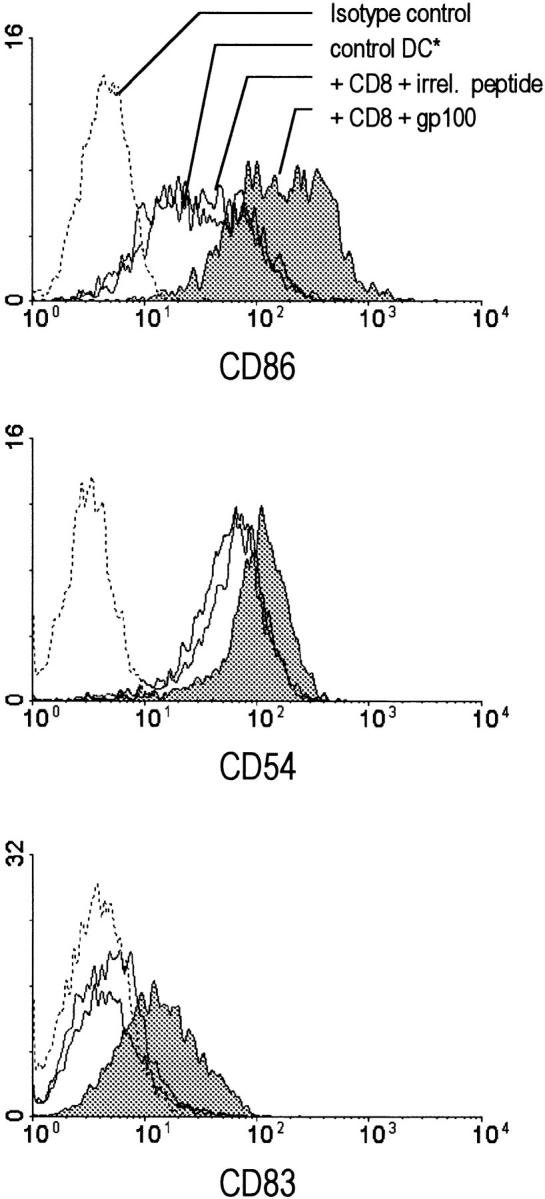
Class I–restricted melanoma (gp100)-specific CD8+ T cell clone induces DC activation. Day 6, immature HLA-A2+ DCs (see Materials and Methods) were cocultured with melanoma gp100-specific HLA-A2–restricted CD8+ T cells (2 × 105 cells/well), in the presence of the relevant gp100 (209–217) peptide (shaded histograms). The cells were harvested 48 h later and analyzed by FACScan® for the level of expression of costimulatory molecules, CD86 (B7.2) and CD54 (ICAM-1), and the maturation-associated marker CD83. Cultures containing DCs alone or DCs cultured with the CD8+ T cell clone in the presence of an irrelevant peptide, tyrosinase (368–376), were used as controls, as indicated. The data shown is from a representative experiment of five performed. *Similar expression of the maturation-associated markers was found on DCs exposed to Ag alone or to CD8+ T cells alone, in this and all other experiments. These additional negative controls are not shown for the sake of clarity of the figures.
Figure 2.


Complementary DC-activating signals from naive CD4+ and CD8+ T cells result in the primary induction of IL-12 and the resulting development of Th1 responses (A). TNF-α–preactivated, SEB-coated DCs (2 × 104 cells/well) were coincubated with naive CD45RA+CD4+T cells, naive CD8+, or bulk CD8+ T cells (105 cells/well), or their combination. Concentrations of IL-12 in 48 h supernatants were analyzed with a p70-specific ELISA. The data (mean ± SD) represents one experiment of four that yielded similar results. Similar results were obtained with day 6 DCs that had not been pretreated with TNF-α. (B) CD45RA+CD4+ naive Th cells (105 cells/well) were primed with TNF-α–preactivated, SEB-coated DCs (2 × 104 cells/well), either in the absence or in the presence of γ-irradiated CD45RA+CD8+ T cells (105 cells/well). At day 10, the cultures contained a uniform population of CD4+ Th (average of 100–300-fold expansion) without any detectable surviving CD8+ T cells (<1%, as determined by FACScan®). The cells were harvested, washed, and stimulated for 24 h with CD3 and CD28 mAbs.
Induction of IFN-γ and IL-12p70 in Triple Cultures of DCs, CD4+, and CD8+ T Cells.
Day 6 DCs were stimulated overnight with TNF-α (50 ng/ml; Strathmann), pulsed with SEB (1 ng/ml) for 30 min, subsequently washed, and plated (2 × 104 cells/well) in 96-well flat-bottom culture plates (Costar). Naive CD45RA+CD4+ T cells (105 cells/well) were added, either in the absence or in the presence of naive CD45RA+CD8+ T cells or bulk CD8+ T cells (105 cells/well in either case), up to a total volume of 200 μl. Cytokine-containing supernatants were collected after 48 h.
Differential Priming of Naive Th Cells.
The priming cultures consisted of SEB-pulsed DCs (2 × 104 cells/well) and CD45RA+CD4+ Th cells (105 cells/well) with or without γ-irradiated (3,000 Rad) CD45RA+CD8+ T cells (105 cells/well). On day 3, rhIL-2 (50 IU/ml) was added to the cultures. At day 10–12, the expanded CD4+ cells (100–300-fold expansion of CD4+ cells, death of irradiated CD8+ cells) were washed, counted, plated in 96-well plates (105 cells/well) and stimulated with CD3 (1 μg/ml) and CD28 (2 μg/ml) mAb. The supernatants were collected after 24 h and analyzed for the presence of IL-4 and IFN-γ.
Direct Cocultures of DCs with CD8+ Cells and Transwell Experiments.
Freshly isolated bulk or naive CD8+ T cells, gp100-specific clone, or influenza-specific CD8+ T cells (2 × 105 cells in each case) were added to day 6 DC cultures, either in the absence or in the presence of the relevant Ag (gp100 [209–217] peptide, or influenza [Flu M1; 58–66] peptide) or antigen surrogates (SEB [1 ng/ml], or OKT3 mAb [1 μg], as indicated). After 48 h incubation, the cells were harvested, washed, and either analyzed by FACS®, or counted and stimulated (2 × 104 cells in 0.2 ml) in 96 wells with CD40L-transfected J558 cells (5 × 104 cells) for 24 h. When indicated, instead of adding CD8+ T cells directly to DC cultures, 2 × 105 CD8+ T cells and 3 × 104 APCs (autologous DCs or HLA-A2+DR4+ T2 cells) were placed in the upper chamber of Transwell™ culture inserts (Costar-3413; pore size of 0.4 μm), in the absence or the presence of relevant antigens. After 48 h, DCs from the lower chamber were harvested, washed, and analyzed as described above. To neutralize the biological activity of TNF-α and IFN-γ in CD8+ T cell–DC cocultures, human soluble TNF receptor I (sTNF RI; 1 μg/ml; R&D Systems) or IFN-γ receptor 1 (IFN-γ R1; 10 μg/ml; R&D Systems) were added at the beginning of cocultures.
Flow Cytometry.
Cell surface phenotypes were analyzed by FACScan™ (Becton Dickinson). The FITC- and PE-labeled isotype control (mouse IgG1 and IgG2a), anti–human CD86, CD54, CD3, CD4, CD8, HLA-DR, CD45RA, CD45RO, and CD14 mAbs were purchased from BD PharMingen. CD83 mAb was obtained from Immunotech. CD1a mAb was purchased from Diaclone.
Cytokine Detection.
Concentrations of IL-12p70, TNF-α, IFN-γ, and IL-4, were determined by specific ELISAs, performed with the matched reagents from Endogen.
Results
Naive CD8+ T Cells Cooperate with Naive CD4+ T Cells in the Induction of IL-12 in DCs and Promote the Development of Primary Th1 Responses.
An efficient induction of bioactive IL-12p70 during the interaction of CD4+ T cells with DCs depends on CD40–CD40L interaction, but requires a costimulatory signal which can be provided by such endogenous factors as IFN-γ (17, 18) or IL-4 (19, 20), or by pathogen-related signals (8–12, 16–20). Naive CD45RA+CD4+ T cells are poor producers of either factor within the first days of their priming (21–23), which allows for only limited induction of IL-12 during their interaction with DCs (17, 18). While memory/effector CD4+ T cells are known to produce substantial amounts of IFN-γ and IL-4 and effectively induce IL-12, the mechanism of endogenous IL-12 induction during primary Th1 responses remains elusive.
Unexpectedly, in sharp contrast to naive CD4+ CD45R0−/RA+ cells (Fig. 1 A), that did not produce detectable amounts of IFN-γ at the early time points of their priming (Fig. 1 B), and acquired the ability to produce this factor with a 72 h delay (data not shown), naive CD8+CD45R0−/RA+ proved to be highly efficient producers of IFN-γ, already within 24 h of their priming (Fig. 1 B). High IFN-γ–producing ability was similarly well-pronounced in CD8+CD45R0−/RA+ isolated from cord blood and from peripheral blood and was both observed after the stimulation with a pair of CD3 and CD28 stimulatory mAbs, or during the interaction with DC in a superantigen (SEB) model, that allows stimulation of a high proportion of both CD4+ and CD8+ T cells expressing the appropriate TCR Vβ chains (24; Fig. 1 B). Although the ability to induce IFN-γ production in naive CD8+ T cells was especially well pronounced in the case of TNF-α–pretreated DCs (Fig. 1 B), similar differences in the IFN-γ production were also observed when we used TNF-α–untreated, CD83−, day 6 monocyte-derived DCs (data not shown), consistent with their expression of significant amounts of costimulatory molecules already at that stage (reference 25, also see the current data below). CD40L expression by the two populations showed a pattern reciprocal to their IFN-γ–producing abilities, being restricted to activated naive CD4+ T cells and absent in CD8+ T cells (Fig. 1 C).
In accordance with the two-signal requirement for the induction of IL-12p70 in DCs (10, 17, 18) 48 h cocultures of SEB-presenting DCs with CD45RA+ CD4+ naive Th cells did not yield any detectable IL-12p70 production (Fig. 2 A), consistent with the ability of naive Th cells to provide a CD40L-mediated stimulus, but not the IFN-γ–mediated IL-12–coinducing signal (17, 18). The parallel cultures of naive CD45RA+CD8+ T cells with DCs yielded high concentrations of IFN-γ, comparable to these found in the cocultures of DCs with bulk CD8+ T cells, but neither naive nor memory CD8+ T cells were able to induce IL-12p70 by themselves, a finding consistent with the absence of CD40L on their surface (Fig. 1 C) and the inability of CD8+ T cells to provide DCs with the CD40-mediated activating signal (3–5). In sharp contrast, the simultaneous interaction of DCs with both naive CD8+ and naive CD4+ Th cells resulted in the effective induction of IL-12p70 production (Fig. 2 A).
To test the modulatory impact of CD8+ T cells on the development of naive Th cells into Th1- or Th2-type effector/memory cells, we compared the outcome of priming of CD45RA+ CD4+ T cells by SEB-primed DCs either in the absence or in the presence of 3,000 Rad-irradiated CD45RA+CD8+ T cells. After 10 d of growth (100- to 300-fold expansion of CD4+ cells and death of irradiated CD8+ cells), such differentially primed Th cells were restimulated with CD3 and CD28 mAb to analyze their cytokine production profiles. As shown in Fig. 2 B, CD4+ Th cells primed in the presence of naive CD8+ T cells developed strongly polarized Th1-type cytokine profile, characterized by the production of high amounts of IFN-γ, but only trace quantities of IL-4. In contrast, the priming cultures performed in the absence of irradiated CD8+ T cells yielded the memory/effector Th cells with strongly reduced IFN-γ and enhanced IL-4 production. Thus, the unique ability of naive CD8+ T cells to produce IFN-γ, a necessary cofactor of CD40L-induced IL-12 production in DCs (18), allows them to play a “helper” role during the induction of primary Th1-type responses.
Class I-restricted CD8+ T Cells Enhance the Activation Stage of Ag-bearing DCs.
To test if the CD8+ T cell–dependent “help” in the development of polarized Th1 responses is indeed mediated by the modulation of DC functions, we analyzed the impact of CD8+ T cells on DC activation/maturation stage and the ability of DCs to produce IL-12. Day 6 monocyte-derived DCs expressed intermediate levels of costimulatory molecules, and lacked the expression of CD83, human DC maturation marker (Fig. 3). DCs exposed to CD8+ T cell clone recognizing the 209–217 epitope (ITDQVPFSV) of the gp100 melanoma-associated antigen in context of HLA-A2 (14) increased their surface expression of costimulatory molecules and acquired CD83 expression (Fig. 3). These effects required both the relevant class I–restricted antigenic peptide and the Ag-specific CD8+ T cells, indicating the specificity of the effect. A similar maturation-inducing activity was also observed using short-term cultured influenza-specific HLA-A2–restricted CD8+ T cells isolated from peripheral blood, using a specific tetramer (15) isolation (Fig. 4 A, see the inset for the frequency of the tetramer positive T cells).
To test the DC-activating properties of resting memory and naive CD8+ T cells, we used the SEB model and a CD3 mAb model, where low concentrations of OKT3 (IgG2a) mAb are captured by FcR on immature DCs and “presented” to T cells (26). Freshly isolated human peripheral blood CD8+ T cells induced the activation of DCs in each of these models (Fig. 4 B). Similar to the class I–restricted peptide models, this effect was also critically dependent on the presence of CD8+ T cells and an antigen surrogate. SEB or soluble CD3 mAb were completely ineffective in the absence of CD8+ T cells, demonstrating that the observed DC activation was not caused by any contaminating CD40L-expressing CD4+ T cells in DC preparation (<1%) or by a direct action of these factors on DCs. Experiments performed in a transwell system allowed us to determine that the DC maturation-inducing activity of CD8+ T cells is mediated by soluble factor(s). Maturation of DCs cultured in the bottom chamber critically depended on the ability of CD8+ T cells from the upper chamber to recognize class I–restricted peptide or SEB presented on the surface of DCs (Fig. 4 C).
The ability of both bulk CD8+ T cells ( Fig. 4 B) and naive CD8+ T cells isolated from cord blood (Fig. 4 D) or peripheral blood (data not shown) to contribute to the maturation of monocyte-derived DCs implicates that CD8+ T cells may affect DC activation status already during primary immune responses. Importantly, however, the release of the activating factor by resting naive CD8+ T cells required their activation by DCs, and could not be induced by nonprofessional APCs, such as the HLA-A2+, DR4-transfected T2 cells (Fig. 5 A). In contrast, effector-type CD8+ T cells, such as gp100-specific CTLs, induced DC maturation when stimulated either by peptide-presenting HLA-A2+ autologous DCs (Figs. 3 and 4 C) or by peptide-loaded T2 cells (Fig. 5 A). ELISA analysis revealed the presence of TNF-α, a known inducer of DC maturation, in the supernatants from 48 h cocultures of DCs with freshly isolated naive and bulk CD8+ T cells, and with effector-type CD8+ T cells (Fig. 5 B). TNF-α release was observed in the cocultures of resting CD8+ T cells with DCs, but not with T2 cells, while in the case of effector CD8+ cells (gp100-specific CTLs) it could be induced both by DCs and by the nonprofessional APCs (Fig. 5 B), a pattern similar to the release of the DC maturation–inducing activity. Indeed, TNF-α proved to be a critical factor for the CD8+ T cell induced DC maturation, as its neutralization with soluble TNF receptor I (sTNF-RI) prevented the CD8+ T cell–induced DC maturation, both in direct cocultures (Fig. 5 C) and in transwell experiments (Fig. 5 D).
Figure 5.


Different activation requirements of naive and effector-type CD8+ T cells to release TNF-α and induce DC maturation. (A) Ability of effector, but not naive CD8+ T cells to activate bystander DCs upon the interaction with nonprofessional APCs. Peripheral blood-isolated CD8+CD45RA T cells and gp100-specific CD8+ T cell clone (2 × 105 cells/well) were stimulated in the upper chamber of the transwell system with, respectively, the relevant gp100 peptide or SEB, presented by HLA-A2+/DR4+ T2 cells or day 6 DCs (3 × 104 cells/well), as indicated. The phenotype of the cells from the bottom chamber was analyzed after 48 h. The additional negative control groups (no antigen and no APCs) did not show any signs of DC maturation, compared with the DCs cultured alone (“control”) and were omitted from the figure, for the sake of its clarity. (B) Production of TNF-α in the cocultures of effector or naive CD8+ T cells with nonprofessional APCs or DCs. Peripheral blood–isolated CD8+CD45RA and gp100-specific CD8+ T cell clone (105 cells/well) were stimulated for 48 h with, respectively, the relevant gp100 peptide or SEB, presented by HLA-A2+/DR4+ T2 cells or by day 6 DCs (2 × 104 cells/well), as indicated. No TNF-α production was found in the supernatants from the additional negative control groups (no antigen and no APCs; data not shown). (C) Neutralization of TNF-α in the direct cocultures of SEB-presenting DCs with bulk isolated CD8+ T cells (2 × 105 cells/well) using sTNF-RI (1ug/ml) effectively blocks the CD8+ T cell–mediated DC maturation. The data shown is from one experiment of three, that all gave similar results. TNF-RI showed a similar ability to block DC maturation induced by in the gp100 specific CTL model (data not shown). (D). Neutralization of TNF-α with sTNF-RI (1 μg/ml) effectively blocks the CD8+ T cell–mediated DC maturation (transwell experiments). DCs in the bottom chamber were exposed to the soluble products released in the cocultures of SEB-presenting DCs (3 × 104 cells/well) with freshly-isolated bulk CD8+ T cells (2 × 105 cells), present in the upper chamber.
Class I–restricted CD8+ T Cells Induce Type-1 Polarized Effector DCs with Elevated IL-12–producing Capacity.
In addition to the induction of DC maturation, the recognition of MHC class I–presented Ag by CD8+ T cells also results in type-1 polarization of maturing DCs, manifested by their enhanced ability to produce IL-12p70 after subsequent DC stimulation. DCs cocultured for 48 h with gp100-specific CTLs in the presence of gp100 peptide, then harvested, washed, and stimulated with CD40L-transfected J558 cells (J558-CD40L) produced strongly increased amounts of IL-12p70 (Fig. 6 A). Similar effects were found in DCs cocultured with influenza-specific tetramer-isolated CD8+ T cells and the relevant influenza-derived peptide (Fig. 6 A), as well as in the cultures of DCs with freshly isolated CD8+ T cells in the presence of SEB (Fig. 6 B) or CD3 mAb (data not shown). In all four systems, DC polarization required the presence of both CD8+ T cells and class I–restricted antigenic peptides (or SEB/CD3 as Ag surrogates), confirming the specificity of the effect. Interestingly, both bulk and naive CD8+ T cells displayed the ability to induce type-1–polarized DCs (Fig. 6 B).
Figure 6.

CD8+ T cells induce type-1–polarized DC phenotype. Day 6 DCs were cocultured for 48 h with CD8+ T cells either in the absence or presence of relevant Ag or Ag surrogate. Subsequently the cells were washed, counted, and stimulated with J558-CD40L, to analyze the production of IL-12p70 in 24 h supernatants. (A) HLA-A2+ DCs cocultured directly with HLA-A2–restricted gp100 (209–217)-specific (white bars; top) or influenza specific (black bars; bottom) CD8+ T cells, in the presence of the relevant class I–restricted peptides. Neither of the peptides showed any DC polarizing effect in the absence of specific T cells (not shown). (B) DCs were activated with peripheral blood-isolated bulk or naive CD8+ T cells in the presence of SEB. (C) Bystander induction of DC maturation by CD8+ T cells. gp100-specific CD8+ T cells, or CD8+ T cells freshly isolated from peripheral blood (2 × 105 cells/well), were stimulated in the top chamber of the transwell system with, respectively, gp100 or SEB, presented by (3 × 104 cells/well) HLA-A2/DR4–expressing T2 cells (white bars, top) or HLA-A2+ DC (black bars, bottom). The bottom chamber contained DCs only. DCs from the bottom chamber were harvested after 48 h, washed, and stimulated for 24 h with J558-CD40L to analyze their ability to produce IL-12. (D) Neutralization of either TNF-α or IFN-γ prevents type-1 DC polarization by gp100-specific CD8+ T cells in the direct coculture system (white bars) and in the transwell system (black bars). sTNF-R1 and sIFN-γ-R were used as neutralizing agents. The data shown is from one experiment of two (direct cocultures) or three (transwell system), which yielded similar results. Similar results were also obtained in the SEB model (not shown).
To test if the type-1–polarizing effect of CD8+ T cells required a direct cell-to-cell contact, we analyzed the ability of the HLA-A2–restricted, gp100-specific CTL clone, and freshly isolated CD8+ T cells to induce DC polarization in the transwell system. Similar to their ability to induce DC maturation, Ag-driven interaction of the effector-type CD8+ T cells with peptide-loaded T2 cells or DCs in the upper chamber resulted in the polarization of DCs in the bottom chamber, as evidenced by their strongly elevated production of IL-12 after the subsequent stimulation by J558-CD40L (Fig. 6 C). As in this model CD8+ T cells were no longer present during the CD40L-mediated DC stimulation, these experiments eliminated the possibility that the enhanced IL-12 production by CD8+ T cell–activated DCs resulted from the persistent production of IFN-γ by contaminating CD8+ T cells. Although resting CD8+ T cells were effective inducers of DC polarization upon a direct contact (Fig. 6 B) and could also polarize bystander DCs in a transwell model (Fig. 6 C), their ability to release the polarizing activity strictly depended on being activated by DCs, while T2 cells were completely ineffective.
The CD8+ T cell–dependent type-1 DC polarization was strongly inhibited by the addition of either sTNF-RI or soluble IFN-γ receptor (sIFN-γ-R), indicating the requirement for both of these CD8+ T cell–derived factors in DC polarization (Fig. 6 D). The effects of both agents were especially pronounced in the transwell system, but were also observed in the system of a direct DC–CD8+ T cell contact. The ability of neutralizing agents for either TNF-α and IFN-γ to interfere with DC polarization is in accordance with our previous observations that the simultaneous exposure of immature DCs to both a maturation-inducing agent and to recombinant IFN-γ, but not to either of these factors alone, can induce type-1 polarized DCs (27). In fact, TNF-α alone or another DC maturation–inducing factor, bacterial LPS, both reduce the ability of DCs to produce IL-12 after subsequent stimulation (Fig. 7 A), consistent with the previous reports (16, 27, 28). Similarly, the exposure of immature DCs to IFN-γ in the absence of any maturation-inducing stimulus does not induce type-1 polarized DC phenotype, but paradoxically reduces the ability of DCs to produce IL-12 (27). Although TNF-α and IFN-γ proved to be important mediators of the CD8+ T cell–dependent DC polarization, even high doses of these factors consistently resulted in a weaker polarization of DCs than CD8+ T cell contact (Fig. 7 B).
Figure 7.


Reciprocal regulation of the IL-12–producing capacity in maturing DCs by CD8+ T cells as opposed to bacterial LPS or an inflammatory mediator, TNF-α. (A) Day 6 DCs were either cultured for the additional 48 h in the absence of any stimuli (immature DCs), or activated in the transwell system (see Fig. 6) by the DC-stimulated gp100-specific CD8+ T cells (2 × 105 cells/well). Alternatively, DCs were preactivated by the exposure to LPS or TNF-α, as indicated. At day 8, DCs from the bottom chamber were harvested, washed, and stimulated for 24 h with J558-CD40L, before harvesting of the supernatants and analyzing them for IL-12 concentration. The data shown is from a representative experiment of five. (B) Comparison of the IL-12 production by CD8+ T cell–polarized DCs and by DCs polarized by the combination of exogenously added TNF-α and IFN-γ. Day 6 DCs were either exposed to gp100-specific CD8+ T cells (2 × 105 cells) and gp100 peptide (direct cocultures or transwell system) or to the standard concentrations of TNF-α and IFN-γ (see Materials and Methods) or the high concentrations of these factors (respectively, 100 ng/ml and 5,000 U/ml). At day 6, the cells were harvested, washed, and stimulated for 24 h with J558-CD40L. The data shown is from one experiment of four, that showed similar results. The inset: J558-CD40L-induced IL-12p70 production by TNF-α and IFN-γ–matured DCs and by DCs polarized by direct cocultures with gp100-specific CD8+ T cells. The data is shown as a relative increase in IL-12p70 production compared with control immature DC (n = four different donors). The level of significance was determined by Student's t test.
Discussion
Current models of the immune response recognize MHC class I–restricted CD8+ T cells as effector cells against intracellular pathogens, reserving T helper function for MHC class II–restricted CD4+ T cells, that activate DCs, instructing them to induce effector functions in CD8+ T cells (3–5). The present demonstration that human MHC class I–restricted CD8+ T cells contribute to DC activation, and induce Th1-promoting phenotype in DCs, indicates a novel role for human CD8+ T cells as “helper cells” during the induction of Th1-polarized inflammatory-type responses, and a role for DCs as effective carriers of such “reverse” (CD8+ to CD4+) helper signals.
While a large body of data supports the role of DCs in providing naive Th cells with the signals that drive immune responses to Th1- or Th2-direction (29–31), it is unclear how DCs distinguish between the different classes of pathogens that require Th1- versus Th2-type responses. One previously proposed mechanism rests on the ability of some intracellular pathogens to directly induce the production of IL-12 or of another Th1-driving factor, IFN-α (9–12, 32, 33). Another proposed scenario is a selective ability of distinct types of pathogens to interact with different Th1-inducing or Th2-inducing DC subpopulations (34), e.g., via different pattern recognition receptors. The current results suggest the existence of an additional “pathogen activity-driven” mechanism, that is not contingent on the ability of DCs themselves to interact with the pathogen and/or to judge its molecular structure. In contrast to DCs themselves, CD8+ T cells that recognize class I–presented peptides, mostly derived from the antigens present in cytoplasmic compartment of cells (35), have an intrinsic capacity to determine the intracellular character of pathogens. In addition to the antigens originating from their own cytoplasm, DCs are capable of class I presentation of pathogen-related antigens acquired from dying cells (36). This ability (cross-presentation), was previously postulated to allow DCs to induce CD8+ T cell responses against the pathogens that either do not infect DCs (37) or that developed the means to suppress the functions of directly infected APCs (38). The same ability, and the ability to receive polarizing signals in a bystander fashion, from CD8+ T cells detecting the infection of other cell types, may help DCs to induce the desirable Th1 pattern of CD4+ cell responses against such pathogens, reducing their chances to evade the immune system. The effectiveness of such a mechanism is augmented by the ability of CD8+ T cells to promote the production of IL-12 by either being physically present during DC–CD4+ T cell interaction, or by inducing type-1 polarized effector DCs that will produce enhanced IL-12 levels upon a subsequent interaction with CD40L-expressing CD4+ T cells. This allows the transmission of the DC-mediated Th1-inducing signal without the need for the Ag-specific CD8+ T cells and CD4+ T cells to interact with DCs at the same time or at the same site.
The DC-modulating impact of CD8+ T cells may explain their immunoregulatory function postulated by Kemeny (39). More recently, murine Ag-specific CD8+ T cells were shown to suppress the production of Th2 cytokines and IgE in response to OVA (40). Furthermore, the depletion of CD8+ cells was recently shown to abrogate the protective Th1-mediated responses after the DNA vaccination against Leishmania (41). CD8+ T cells were also demonstrated to shift the pattern of Th1/Th2-dependent antibody isotypes during the response against Respiratory Syncytial Virus (42).
CD8+ T cell–dependent DC activation and polarization may be also responsible for the different cytokine patterns of the responses against Th1-inducing contact allergens as opposed to the Th2-inducing atopic allergens. In contrast to proteinaceous atopic allergens, presented mainly to CD4+ T cells in context of MHC class II, contact allergens are effectively presented both by MHC class II and class I molecules, and activate both CD4+ and CD8+ T cells (43). This may lead to their perception by the immune system as intracellular pathogens, resulting in the induction of IL-12 and mobilization of the inflammatory-type Th1-driven immunity, manifested as contact sensitization.
CD8+ T cell–induced mature DCs are functionally different than DCs matured under the influence of bacterial LPS or inflammatory cytokines, including IL-1β and TNF-α, which all show a maturation-related decrease in the ability to produce IL-12 and to induce Th1 responses (16, 28). We have reported that the exposure of DCs to recombinant IFN-γ during the first hours of their TNF−α/IL-1β- or LPS-induced maturation reverses the maturation-related decrease in IL-12 producing capacity (27). The resulting type-1 polarized DCs produce up to 100-fold more IL-12p70 than DCs matured by LPS or TNF−α/IL-1β alone. Although both IFN-γ and TNF-α proved to be involved in the currently described CD8+ T cell–induced DC polarization, the exposure of DCs to these two factors was consistently giving weaker results than the exposure of DCs to CD8+ T cells (Fig. 7 B). These observations may have several possible explanations: first, they may suggest a participation of additional T cell–derived factors. In line with this possibility, the neutralization of IFN-γ and TNF-α was more efficient in the transwell experiments than during direct cocultures (Fig. 6 D), suggesting perhaps the additional role for membrane-bound molecules. Alternatively, it may be the particular kinetic of the production of IFN-γ and/or TNF-α during CD8+ T cell–DC interaction, particular concentrations of these cytokines or their matched ratio at the contact area of the interacting cells, that result in their optimal activity. The above possibilities are being currently tested.
In sharp contrast to the observations made in the CD4+ T cell system (21–23), the ability to produce IFN-γ and the resulting ability to polarize DCs is present not only in effector CD8+ T cells and freshly isolated memory CD8+ T cells, but also in the cord blood– or peripheral blood–isolated naive CD8+ T cells, at the earliest stages of their interaction with DCs. This suggests that CD8+ T cells may play a Th1-driving role already during the induction of primary immune responses. However, in contrast to effector CD8+ T cells that can release the DC-polarizing activity after the stimulation by nonprofessional APCs, such as T2 cells, naive CD8+ T cells need to be activated by an APC providing a certain level of costimulation, such as the GM-CSF– and IL-4–cultured (even if not fully mature) monocyte-derived DCs (see Fig. 3 for the expression of costimulatory molecules on control CD83− DCs). This suggests that, similar to naive CD4+ T cells, naive CD8+ T cells depend on an initial pathogen-dependent DC activation in order to further boost the activation status of DCs and induce their type-1 polarization. Further studies are needed to define what is the minimal level of activation permitting DCs to effectively interact with naive CD8+ T cells. Using a TCR transgenic mouse model, Ruedl and colleagues have shown that the injection of specific class I–restricted peptide results in a massive activation of both the Ag-presenting and bystander DCs (6). These observations indicated the ability of naive CD8+ T cells to activate resting DCs in vivo, in a soluble factor-mediated mechanism. A more recent study (7) showed that the transfer of 20 × 103 TCR-transgenic CD8+ T cells allowed them to activate DCs and function in a CD4+ T cell–independent fashion in recipient mice, while lower amounts of CD8+ T cells were unable to function autonomously. This requirement for relatively high numbers of Ag-specific cells may explain a strict requirement for additional DC-activating stimuli, including the signals from CD4+ T cells, in most of CD8+ T cell responses.
In our current experimental setup, we observed only a marginal degree of DC killing in the experiments with resting CD8 T cells, as well as in all cases of bystander DC polarization, while even the direct cocultures of DCs with gp100-specific CTL resulted with only up to 30% lower DC recovery, compared with other modes of DC maturation (data not shown). A recent paper showed that at least some CTL clones may be more effective killers of immature bone marrow–derived mouse DCs and of the DC-like D1 cells (44). It remains to be established whether any particular activation stages of CD8+ T cells or particular subsets of CD8+ T cells, exert predominantly cytotoxic, rather than DCs activating and polarizing roles. If this is the case, helper signals from CD40L-expressing CD4+ T cells, that can protect DCs from Fas-mediated killing (45, 46), may have additional importance.
The current results suggest a consensual, “socratic,” rather than a hierarchical, “instructive,” model of the interaction between DCs, CD4+, and CD8+ T cells, during the induction of cell-mediated immune responses. In both models, the activation of resting DCs by the pathogen (47) or by the pathogen-induced tissue response (48) is required for the migration of DCs to the lymph nodes and their ability to activate naive T cells. The currently accepted “hierarchical” model postulates that a subsequent step is the CD40-CD40L–mediated interaction with Ag-specific CD4+ T cells, allowing DCs to acquire a “second-step” level of activation and to effectively interact with naive CD8+ T cells (3–5). The current data and the results obtained in the mouse (6, 7) suggest an alternative “socratic” model, where CD4+ and CD8+ T cells act in a network-like system, controlling each-others' activity via bidirectional DC-mediated signals. In this model, the combination of TNF-α- and IFN-γ–dependent “CD8 help” and CD40L-dependent “CD4 help” allows the pathogen-mobilized DCs to acquire an optimal stimulatory function and to produce IL-12, an important factor in the induction of the effector functions and the long-term survival of CTL, NK, and Th1 cells (8).
The DC-modulating function of CD8+ T cells may have implications for the design of vaccination strategies in cancer, chronic infections, and allergy. KLH, a “heterologous” CD4+ Th cell antigen, has been used to enhance the efficacy of DC-based vaccination against melanoma (49). The current results raise a possibility of using the heterologous help from CD8+ T cells specific for common pathogens to optimize the character of DC-induced anti-tumor responses, or to correct the pathogenic Th2-bias in atopy. The feasibility of these approaches is currently being evaluated.
Acknowledgments
The authors would like to thank Steven Rosenberg and John Wunderlich for providing gp100-specific CTL line, Peter Lane for providing CD40L-transfected J558 cells, Janice Blum for T2 cells, and Kathy Rakow for secretarial assistance. We thank Polly Matzinger, Penny Morel, Stefania Galluci, and Young-Jun Liu for critically reading the manuscript and for stimulating discussions.
The current study was supported by the National Cancer Institute grants: 1RO1CA82016, 1RO1CA57840, and 1PO1CA74343, and by the 2000 Kathy Salling Memorial Melanoma Research Foundation Award.
Footnotes
Abbreviations used in this paper: DC, dendritic cell; SEB, Staphylococcus Enterotoxin B.
References
- 1.Mosmann, T.R., and S. Sad. 1996. The expanding universe of T-cell subsets: Th1, Th2 and more. Immunol. Today. 17:138–146. [DOI] [PubMed] [Google Scholar]
- 2.Banchereau, J., and R.M. Steinman. 1998. Dendritic cells and the control of immunity. Nature. 392:245–252. [DOI] [PubMed] [Google Scholar]
- 3.Ridge, J.P., F. Di Rosa, and P. Matzinger. 1998. A conditioned dendritic cell can be a temporal bridge between a CD4+ T- helper and a T-killer cell. Nature. 393:474–478. [DOI] [PubMed] [Google Scholar]
- 4.Schoenberger, S.P., R.E. Toes, E.I. van der Voort, R. Offringa, and C.J. Melief. 1998. T-cell help for cytotoxic T lymphocytes is mediated by CD40-CD40L interactions. Nature. 393:480–483. [DOI] [PubMed] [Google Scholar]
- 5.Bennett, S.R., F.R. Carbone, F. Karamalis, R.A. Flavell, J.F. Miller, and W.R. Heath. 1998. Help for cytotoxic-T-cell responses is mediated by CD40 signalling. Nature. 393:478–480. [DOI] [PubMed] [Google Scholar]
- 6.Ruedl, C., M. Kopf, and M.F. Bachmann. 1999. CD8+ T cells mediate CD40-independent maturation of dendritic cells in vivo. J. Exp. Med. 189:1875–1884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wang, B., C.C. Norbury, R. Greenwood, J.R. Bennink, J.W. Yewdell, and J.A. Frelinger. 2001. Multiple paths for activation of naive CD8+ T cells: CD4-independent help. J. Immunol. 167:1283–1289. [DOI] [PubMed] [Google Scholar]
- 8.Trinchieri, G. 1995. Interleukin-12: a proinflammatory cytokine with immunoregulatory functions that bridge innate resistance and antigen-specific adaptive immunity. Annu. Rev. Immunol. 13:251–276. [DOI] [PubMed] [Google Scholar]
- 9.Sousa, C.R., S. Hieny, T. Scharton-Kersten, D. Jankovic, H. Charest, R.N. Germain, and A. Sher. 1997. In vivo microbial stimulation induces rapid CD40 ligand-independent production of interleukin 12 by dendritic cells and their redistribution to T cell areas. J. Exp. Med. 186:1819–1829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Schulz, O., D.A. Edwards, M. Schito, J. Aliberti, S. Manickasingham, A. Sher, and C. Sousa. 2000. CD40 triggering of heterodimeric IL-12 p70 production by dendritic cells in vivo requires a microbial priming signal. Immunity. 13:453–462. [DOI] [PubMed] [Google Scholar]
- 11.Siegal, F.P., N. Kadowaki, M. Shodell, P.A. Fitzgerald-Bocarsly, K. Shah, S. Ho, S. Antonenko, and Y.J. Liu. 1999. The nature of the principal type 1 interferon-producing cells in human blood. Science. 284:1835–1837. [DOI] [PubMed] [Google Scholar]
- 12.Cella, M., D. Jarrossay, F. Facchetti, O. Alebardi, H. Nakajima, A. Lanzavecchia, and M. Colonna. 1999. Plasmacytoid monocytes migrate to inflamed lymph nodes and produce large amounts of type I interferon. Nat. Med. 5:919–923. [DOI] [PubMed] [Google Scholar]
- 13.Cella, M., D. Scheidegger, K. Palmer-Lehmann, P. Lane, A. Lanzavecchia, and G. Alber. 1996. Ligation of CD40 on dendritic cells triggers production of high levels of interleukin-12 and enhances T cell stimulatory capacity: T-T help via APC activation. J. Exp. Med. 184:747–752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Clay, T.M., M.C. Custer, M.D. McKee, M. Parkhurst, P.F. Robbins, K. Kerstann, J. Wunderlich, S.A. Rosenberg, and M.I. Nishimura. 1999. Changes in the fine specificity of gp100(209-217)-reactive T cells in patients following vaccination with a peptide modified at an HLA-A2.1 anchor residue. J. Immunol. 162:1749–1755. [PubMed] [Google Scholar]
- 15.Dunbar, P.R., G.S. Ogg, J. Chen, N. Rust, B.P. van der, and V. Cerundolo. 1998. Direct isolation, phenotyping and cloning of low-frequency antigen- specific cytotoxic T lymphocytes from peripheral blood. Curr. Biol. 8:413–416. [DOI] [PubMed] [Google Scholar]
- 16.Kalinski, P., J.H. Schuitemaker, C.M. Hilkens, E.A. Wierenga, and M.L. Kapsenberg. 1999. Final maturation of dendritic cells is associated with impaired responsiveness to IFN-gamma and to bacterial IL-12 inducers: decreased ability of mature dendritic cells to produce IL-12 during the interaction with Th cells. J. Immunol. 162:3231–3236. [PubMed] [Google Scholar]
- 17.Hilkens, C.M., P. Kalinski, M. de Boer, and M.L. Kapsenberg. 1997. Human dendritic cells require exogenous interleukin-12-inducing factors to direct the development of naive T-helper cells toward the Th1 phenotype. Blood. 90:1920–1926. [PubMed] [Google Scholar]
- 18.Snijders, A., P. Kalinski, C.M. Hilkens, and M.L. Kapsenberg. 1998. High-level IL-12 production by human dendritic cells requires two signals. Int. Immunol. 10:1593–1598. [DOI] [PubMed] [Google Scholar]
- 19.Kalinski, P., H.H. Smits, J.H. Schuitemaker, P.L. Vieira, M. van Eijk, E.C. de Jong, E.A. Wierenga, and M.L. Kapsenberg. 2000. IL-4 is a mediator of IL-12p70 induction by human Th2 cells: reversal of polarized Th2 phenotype by dendritic cells. J. Immunol. 165:1877–1881. [DOI] [PubMed] [Google Scholar]
- 20.Hochrein, H., M. O'Keeffe, T. Luft, S. Vandenabeele, R.J. Grumont, E. Maraskovsky, and K. Shortman. 2000. Interleukin (IL)-4 is a major regulatory cytokine governing bioactive IL-12 production by mouse and human dendritic cells. J. Exp. Med. 192:823–833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kristensson, K., M. Dohlsten, H. Fischer, P.O. Ericsson, G. Hedlund, H.O. Sjogren, and R. Carlsson. 1990. Phenotypical and functional differentiation of CD4+ CD45RA+ human T cells following polyclonal activation. Scand. J. Immunol. 32:243–253. [DOI] [PubMed] [Google Scholar]
- 22.Salmon, M., G.D. Kitas, and P.A. Bacon. 1989. Production of lymphokine mRNA by CD45R+ and CD45R- helper T cells from human peripheral blood and by human CD4+ T cell clones. J. Immunol. 143:907–912. [PubMed] [Google Scholar]
- 23.Kalinski, P., C.M. Hilkens, E.A. Wierenga, T.C. Pouw-Kraan, R.A. van Lier, J.D. Bos, M.L. Kapsenberg, and F.G. Snijdewint. 1995. Functional maturation of human naive T helper cells in the absence of accessory cells. Generation of IL-4-producing T helper cells does not require exogenous IL-4. J. Immunol. 154:3753–3760. [PubMed] [Google Scholar]
- 24.Fraser, J.D. 1989. High-affinity binding of staphylococcal enterotoxins A and B to HLA-DR. Nature. 339:221–223. [DOI] [PubMed] [Google Scholar]
- 25.Sallusto, F., and A. Lanzavecchia. 1994. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J. Exp. Med. 179:1109–1118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hilkens, C.M., A. Snijders, H. Vermeulen, P.H. van der Meide, E.A. Wierenga, and M.L. Kapsenberg. 1996. Accessory cell-derived IL-12 and prostaglandin E2 determine the IFN- gamma level of activated human CD4+ T cells. J. Immunol. 156:1722–1727. [PubMed] [Google Scholar]
- 27.Vieira, P.L., E.C. de Jong, E.A. Wierenga, M.L. Kapsenberg, and P. Kalinski. 2000. Development of Th1-inducing capacity in myeloid dendritic cells requires environmental instruction. J. Immunol. 164:4507–4512. [DOI] [PubMed] [Google Scholar]
- 28.Langenkamp, A., M. Messi, A. Lanzavecchia, and F. Sallusto. 2001. Kinetics of dendritic cell activation: impact on priming of Th1, Th2 and nonpolarized T cells. Nat. Immunol. 1:311–316. [DOI] [PubMed] [Google Scholar]
- 29.Kalinski, P., C.M. Hilkens, E.A. Wierenga, and M.L. Kapsenberg. 1999. T-cell priming by type-1 and type-2 polarized dendritic cells: the concept of a third signal. Immunol. Today. 20:561–567. [DOI] [PubMed] [Google Scholar]
- 30.Liu, Y.J., N. Kadowaki, M.C. Rissoan, and V. Soumelis. 2000. T cell activation and polarization by DC1 and DC2. Curr. Top. Microbiol. Immunol. 251:149–159. [DOI] [PubMed] [Google Scholar]
- 31.Pulendran, B., J. Banchereau, E. Maraskovsky, and C. Maliszewski. 2001. Modulating the immune response with dendritic cells and their growth factors. Trends Immunol. 22:41–47. [DOI] [PubMed] [Google Scholar]
- 32.von Stebut, E., Y. Belkaid, T. Jakob, D.L. Sacks, and M.C. Udey. 1998. Uptake of Leishmania major amastigotes results in activation and interleukin 12 release from murine skin-derived dendritic cells: implications for the initiation of anti-Leishmania immunity. J. Exp. Med. 188:1547–1552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kadowaki, N., S. Antonenko, J.Y. Lau, and Y.J. Liu. 2000. Natural interferon alpha/beta-producing cells link innate and adaptive immunity. J. Exp. Med. 192:219–226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Liu, Y.J., H. Kanzler, V. Soumelis, and M. Gilliet. 2001. Dendritic cell lineage, plasticity and cross-regulation. Nat. Immunol. 2:585–589. [DOI] [PubMed] [Google Scholar]
- 35.Ploegh, H., and C. Watts. 1998. Antigen recognition. Curr. Opin. Immunol. 10:57–58. [DOI] [PubMed] [Google Scholar]
- 36.Albert, M.L., B. Sauter, and N. Bhardwaj. 1998. Dendritic cells acquire antigen from apoptotic cells and induce class I-restricted CTLs. Nature. 392:86–89. [DOI] [PubMed] [Google Scholar]
- 37.Larsson, M., J.F. Fonteneau, and N. Bhardwaj. 2001. Dendritic cells resurrect antigens from dead cells. Trends Immunol. 22:141–148. [DOI] [PubMed] [Google Scholar]
- 38.Bhardwaj, N. 1997. Interactions of viruses with dendritic cells: a double-edged sword. J. Exp. Med. 186:795–799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kemeny, D.M., A. Noble, B.J. Holmes, and D. Diaz-Sanchez. 1994. Immune regulation: a new role for the CD8+ T cell. Immunol. Today. 15:107–110. [DOI] [PubMed] [Google Scholar]
- 40.Holmes, B.J., P.A. MacAry, A. Noble, and D.M. Kemeny. 1997. Antigen-specific CD8+ T cells inhibit IgE responses and interleukin-4 production by CD4+ T cells. Eur. J. Immunol. 27:2657–2665. [DOI] [PubMed] [Google Scholar]
- 41.Gurunathan, S., L. Stobie, C. Prussin, D.L. Sacks, N. Glaichenhaus, D.J. Fowell, R.M. Locksley, J.T. Chang, C.Y. Wu, and R.A. Seder. 2000. Requirements for the maintenance of Th1 immunity in vivo following DNA vaccination: a potential immunoregulatory role for CD8+ T cells. J. Immunol. 165:915–924. [DOI] [PubMed] [Google Scholar]
- 42.Hyland, L., S. Hou, C. Coleclough, T. Takimoto, and P.C. Doherty. 1994. Mice lacking CD8+ T cells develop greater numbers of IgA-producing cells in response to a respiratory virus infection. Virology. 204:234–241. [DOI] [PubMed] [Google Scholar]
- 43.Kalish, R.S., and P.W. Askenase. 1999. Molecular mechanisms of CD8+ T cell-mediated delayed hypersensitivity: implications for allergies, asthma, and autoimmunity. J. Allergy Clin. Immunol. 103:192–199. [DOI] [PubMed] [Google Scholar]
- 44.Medema, J.P., D.H. Schuurhuis, D. Rea, J. van Tongeren, J. de Jong, S.A. Bres, S. Laban, R.E. Toes, M. Toebes, T.N. Schumacher, et al. 2001. Expression of the serpin serine protease inhibitor 6 protects dendritic cells from cytotoxic T lymphocyte-induced apoptosis: differential modulation by T helper type 1 and type 2 cells. J. Exp. Med. 194:657–667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bjorck, P., J. Banchereau, and L. Flores-Romo. 1997. CD40 ligation counteracts Fas-induced apoptosis of human dendritic cells. Int. Immunol. 9:365–372. [DOI] [PubMed] [Google Scholar]
- 46.Koppi, T.A., T. Tough-Bement, D.M. Lewinsohn, D.H. Lynch, and M.R. Alderson. 1997. CD40 ligand inhibits Fas/CD95-mediated apoptosis of human blood-derived dendritic cells. Eur. J. Immunol. 27:3161–3165. [DOI] [PubMed] [Google Scholar]
- 47.Medzhitov, R., and C.A. Janeway, Jr. 1997. Innate immunity: impact on the adaptive immune response. Curr. Opin. Immunol. 9:4–9. [DOI] [PubMed] [Google Scholar]
- 48.Matzinger, P. 1994. Tolerance, danger, and the extended family. Annu. Rev. Immunol. 12:991–1045. [DOI] [PubMed] [Google Scholar]
- 49.Nestle, F.O., S. Alijagic, M. Gilliet, Y. Sun, S. Grabbe, R. Dummer, G. Burg, and D. Schadendorf. 1998. Vaccination of melanoma patients with peptide- or tumor lysate-pulsed dendritic cells. Nat. Med. 4:328–332. [DOI] [PubMed] [Google Scholar]