

Abstract
The signaling events downstream of integrins that regulate cell attachment and motility are only partially understood. Using osteoclasts and transfected 293 cells, we find that a molecular complex comprising Src, Pyk2, and Cbl functions to regulate cell adhesion and motility. The activation of integrin αvβ3 induces the [Ca2+]i-dependent phosphorylation of Pyk2 Y402, its association with Src SH2, Src activation, and the Src SH3-dependent recruitment and phosphorylation of c-Cbl. Furthermore, the PTB domain of Cbl is shown to bind to phosphorylated Tyr-416 in the activation loop of Src, the autophosphorylation site of Src, inhibiting Src kinase activity and integrin-mediated adhesion. Finally, we show that deletion of c Src or c-Cbl leads to a decrease in osteoclast migration. Thus, binding of αvβ3 integrin induces the formation of a Pyk2/Src/Cbl complex in which Cbl is a key regulator of Src kinase activity and of cell adhesion and migration. These findings may explain the osteopetrotic phenotype in the Src−/− mice.
Keywords: Cbl, Src, Pyk2, osteoclast, vitronectin receptor (αvβ3)
Introduction
In many cell types, the attachment of cells to the extracellular matrix (ECM) via integrins leads to activation of several protein tyrosine kinases and the formation of focal adhesions, multiprotein complexes that anchor actin stress fibers to the cytoplasmic face of the plasma membrane (Longhurst and Jennings 1998). Highly motile cells such as osteoclasts, macrophages, and trophoblasts differ in their attachment, forming podosomes, specialized adhesion sites that are structurally and functionally distinct from focal adhesions (Marchisio et al. 1984, Marchisio et al. 1987; Tarone et al. 1985; Nermut et al. 1991). Although many of the same proteins are present in podosomes and focal adhesions, focal adhesions are relatively stable, whereas podosomes are dynamic attachment structures, undergoing assembly and disassembly within minutes (Stickel and Wang 1987; Chen 1989, Chen 1990). Interestingly, podosomes are also observed in v-src-transformed cells, where they contain high concentrations of Src as well as other tyrosine phosphorylated proteins (Marchisio et al. 1987; Ochoa et al. 2000). Activation of Src may therefore be associated with a shift from stable focal adhesions with actin stress fibers to more dynamic podosome assemblies, possibly regulating cell motility. Indeed, several papers have suggested a role for Src in cell spreading and migration (Kaplan et al. 1995; Meng and Lowell 1998; Felsenfeld et al. 1999). More recently, Felsenfeld et al. 1999 demonstrated that Src reduces the strength of links between the vitronectin receptor (VnR) and the cytoskeleton when the VnR is bound to extracellular matrix proteins. Modulation of these links is essential to generate the forces needed for cell migration, and reinforcement of these links in the absence of Src results in a decrease in both cell spreading and migration. Interestingly, the kinase domain of Src was not necessary for this process, a finding similar to the results of Schwartzberg et al. 1997 in genetic rescue experiments of the Src− osteopetrotic phenotype. Together, these results suggest that the predominant role of Src in these processes might be as an adaptor molecule, recruiting other proteins that are necessary for the modulation of integrin-cytoskeletal links, cell attachment, and migration.
c-Src also plays a critical role in bone resorption, and deletion of the gene in mice results in defective osteoclasts and osteopetrosis (Soriano et al. 1991; Horne et al. 1992; Tanaka et al. 1992; Lowe et al. 1993; Lowell and Soriano 1996). Normal osteoclasts have been demonstrated to contain podosomes (Marchisio et al. 1984, Marchisio et al. 1987), but the signaling pathway(s) that Src regulates in osteoclasts has not been identified. We have previously demonstrated that the phosphorylation of the proto-oncogene c-Cbl and its localization to the cytoskeleton-rich periphery of the osteoclast is Src dependent (Tanaka et al. 1996). Studies in a variety of other cell types have demonstrated that Cbl is tyrosine phosphorylated in response to the activation of a number of cell-surface receptors coupled to tyrosine kinases. These include growth factor receptors, cytokine receptors, and integrins, such as the αvβ3 vitronectin receptor (Miyake et al. 1997), which is the predominant integrin receptor in osteoclasts and essential for bone resorption (Rodan and Rodan 1997). Recent results from several laboratories have also revealed that Cbl can negatively regulate the tyrosine kinases it binds to (Ota and Samelson 1997; Lupher et al. 1998; Broome et al. 1999; Waterman et al. 1999) and participate in their proteosome-dependent degradation by recruiting ubiquitin-conjugating enzymes to the complex (Joazeiro et al. 1999; Levkowitz et al. 1999; Yokouchi et al. 1999).
Since integrins and c-Src play crucial roles in cell migration and are known to be essential for bone resorption (Soriano et al. 1991; Duong and Rodan 1999), we investigated whether integrin-dependent attachment and migration are defective in Src− osteoclasts, since such defects might contribute to the lack of resorptive activity in Src− osteoclasts. We show that while wild-type osteoclasts exhibit podosomes, Src− osteoclasts have attachment structures that are reminiscent of more stably attached cells. Furthermore, they exhibit a decrease in lamellipodia motility and cell migration, which is also true of osteoclasts from c-Cbl− mice.
We also show that Pyk2, a calcium-dependent tyrosine kinase that is the predominant FAK family member in the adhesion structures of osteoclasts (Levy et al. 1997; Duong et al. 1998) is autophosphorylated upon cell attachment, and subsequently forms a complex with activated Src and Cbl. In this complex, autophosphorylated Pyk2 binds to Src's SH2 domain, while Cbl interacts with Src's SH3 domain. In addition, the overexpression of Cbl's PTB domain inhibits Src kinase activity and VnR-mediated cell adhesion, apparently as a result of binding to the autophosphorylated Src Tyr416. We conclude that the Pyk2- and Src-dependent recruitment of Cbl to sites of adhesion may play a role in regulating cell attachment and motility.
Materials and Methods
Antibodies and Plasmids
The generation of plasmids containing myc-tagged Cbl mutants has previously been described (Bartkiewicz et al. 1999). To obtain stable transfections of cells already resistant to G418, these constructs were subcloned into the pcDNA3.1/Zeo(−) vector as Nhe1/Not1 fragments. The Src constructs were generous gifts from Dr. P. Schwartzberg (National Institutes of Health, Bethesda, MD) and Dr. J. Brugge (Harvard University, Cambridge, MA), and the Pyk2 construct was a generous gift from Dr. J. Schlessinger (New York University Medical Center, New York, NY). The G306E mutation of v-Cbl and c-Cbl, and the Y402F and K457A mutations of Pyk2 were performed as previously described (Lev et al. 1995; Dikic et al. 1996; Lupher et al. 1998; Sieg et al. 1998).
Anti–Pyk2 and –Cbl antibodies were obtained from Transduction Laboratories. Anti–avian Src, anti–phosphotyrosine (clone 4G10), and anti–vinculin antibodies were obtained from Upstate Biotechnology. Anti–v-Src antibody (clone Ab-1) was obtained from Calbiochem. Anti–Myc antibody (clone 9E10) was obtained from Santa Cruz Biotechnology, Inc. Anti–Pyk2 phosphotyrosine residue 402 antibody and anti–Src phosphotyrosine residue 416 were obtained from Biosource International. (Anti–Src pTyr 416 used in preliminary experiments was a kind gift of Dr. A. Laudano (University of New Hampshire, Durham, NH). Both anti–rabbit and anti–mouse secondary antibodies were obtained from Fisher Scientific.
Preparation of Authentic Osteoclasts and Osteoclast-like Cells
Authentic osteoclasts were obtained from the long bones of 2–4-d-old neonatal mice. Bones were dissected free of adherent tissues, placed in α-MEM containing 5% FBS, and minced into small pieces. After vigorous pipetting to release osteoclasts, the bone particles were allowed to sediment for 30 s and the remaining cell suspension containing osteoclasts was seeded onto serum-coated coverslips. To obtain large numbers of osteoclastic cells for biochemical analyses, osteoclast-like cells (OCLs) were generated in the murine coculture system (Tanaka et al. 1996) by culturing neonatal primary calvarial osteoblasts with spleen and marrow cells in the presence of 1,25 dihydroxyvitamin D3 [1,25(OH)2D3] and PGE2. Primary calvarial osteoblasts were isolated from neonatal mouse calvaria after sequential digestion with 0.1% collagenase and 0.2% dispase (Calbiochem), and then cultured in α-MEM containing 10% FBS. Cells obtained from spleens and bone marrow cells from Src− or Src+ mice (2 × 108 cells) were cocultured with osteoblastic cells (106) in α-MEM containing 10% FBS, 10−8 M 1,25(OH)2D3, and 10−6 M PGE2 (Sigma-Aldrich). After 4–5 d in culture, OCLs were purified by removing the osteoblast layer by repeated pipetting of media. Suspensions of serum-starved (1% FCS overnight) OCLs were obtained by treating the purified OCLs with 10 mM EDTA for 5 min at 37°C. Cells were then flushed off the culture dishes, washed once in serum-free α-MEM, and then resuspended in serum-free medium.
Confocal Microscopy
Cells were fixed in 3.7% (vol/vol) formaldehyde in PBS for 10 min, and then washed three times in PBS. Coverslips for cytoskeletal labeling were extracted in ice-cold acetone for 3–5 min and returned to PBS. All other coverslips were permeabilized in 0.05% saponin for 30 min. Coverslips for actin labeling were incubated in a 1:40 dilution (in PBS) of rhodamine phalloidin stock solution (Molecular Probes), for 20 min, washed several times with PBS, and mounted in FluorSave (Calbiochem). All other coverslips were blocked in 5% normal goat serum (Boehringer) for 30 min and incubated in appropriate primary antibodies, washed in PBS, incubated with fluorescein-conjugated secondary antibody, and finally washed in PBS and mounted in FluorSave. Cells were examined using a confocal imaging system (MRC-600; Bio-Rad Laboratories). Images were recorded and composites of the various time courses were compiled and total image enhancements were performed using Adobe Photoshop 4.0.
Osteoclast Motility and Migration Measurements
Authentic osteoclasts were freshly isolated, as previously described (Tanaka et al. 1996), from neonatal Src− and Cbl− mice and their wild-type littermates. Osteoclasts in α-MEM containing 10% FCS were placed in the incubation chamber of an inverted phase-contrast microscope linked via a charge-coupled device camera to a computer-assisted image analyzer (Trakcell; BIOCOM) and allowed to adhere and spread for 4 h before measurements were taken. Using a motorized stage and Trackcellwin software, digitized osteoclast images in several fields in parallel were recorded at set-time intervals. Over 50 freshly isolated osteoclasts were analyzed for each group in each experiment. Cells were recorded at 15-min intervals for 4 h. For each image of the sequence, the coordinated cell center of gravity was determined and the trajectory of each osteoclast was calculated. From these trajectories, the length of the cell path was calculated by determining the sum of distances between successive positions of the moving cell. Spreading was quantified as lamellipodia area divided by the total osteoclast area measured at 2-min intervals over a 90-min period. Motility was defined as the membrane area changes, quantified by calculating the nonoverlapping area between frames of two successive images recorded at 2-min intervals over a 90-min period. Statistical analysis was performed using Student's paired t test.
Stable and Transient Transfections
Cells were transfected for 5 h using LipofectAMINE™ (Life Technologies) in α-MEM following the manufacturer's protocol. Stable 293-VnR cell lines expressing myc-tagged Cbl constructs were established by transferring transfected cells into culture medium supplemented with 100 μg/ml G418 (Life Technologies) and 200 μg/ml Zeocin (Invitrogen) 72 h after transfection. Cells were maintained in this medium until resistant colonies of cells were formed (∼2 wk). Resistant colonies were subsequently pooled (>20 colonies) and maintained in culture medium supplemented with 100 μg/ml G418 and 50 μg/ml Zeocin.
Transiently transfected cells were maintained in α-MEM containing 10% FCS for 72 h after transfection. Cells were then lysed in modified radioimmune precipitation assay (mRIPA) or subsequently used in adhesion assays.
Integrin Cross Linking and Integrin-mediated Adhesion-induced Signaling
Mouse osteoclasts were serum starved in α-MEM for 4–6 h before stimulation. OCLs generated in the coculture system were purified from contaminating cells as previously described, and then treated with trypsin-EDTA for 10 min before antibody cross-linking experiments. Cells were then washed with ice-cold α-MEM supplemented with 20 mM Hepes, pH 7.3, and then incubated with a 1:50 dilution of anti–murine VnR polyclonal antiserum (Gailit et al. 1997) for 30 min at 4°C. After washing with ice-cold α-MEM containing 100 μM sodium orthovanadate, the cells were incubated in the presence of a 1:100 dilution of anti–rabbit secondary antibody prepared in the presence of 20 mM Hepes and 100 μM sodium orthovanadate at 37°C for the times indicated. Cells were then rinsed in PBS and lysed in a buffer containing 50 mM Hepes, pH 7.2, 150 mM NaCl, 1% Triton X-100, 10% glycerol, 1 mM EGTA, 1.5 mM MgCl2, 1 mM sodium orthovanadate, 1 μg/ml pepstatin, 10 μg/ml leupeptin and aprotinin, 1 mM PMSF, and 50 mM sodium fluoride.
For experiments involving adhesion of OCLs and 293-VnR cells to ECM proteins, tissue culture–treated plastic or glass coverslips were coated with 10 μg/ml vitronectin, 10 μg/ml laminin, or 100% FCS overnight at 37°C. Plates or coverslips were washed three times in PBS before plating the cells. 293-VnR cells were starved in serum-free medium overnight before being harvested using trypsin-EDTA. To study the role of calcium in adhesion-induced signaling, the detached cells were treated with 100 μM EGTA (to chelate extracellular calcium) for 5 min, or 50 μM BAPTA (to chelate intracellular calcium) for 30 min, or both. Cells were then replated, allowed to adhere for the indicated period of time, and lysed in mRIPA buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.1% Nonidet P-40, 1% sodium deoxycholate, 10 μg/ml leupeptin, 10 μg/ml aprotinin, 1 mM sodium orthovanadate, 10 mM NaF, 1 μg/ml pepstatin, and 1 mM PMSF). Lysates were centrifuged for 30 min at 4°C at 16,000 g, and the supernatants were used for immunoprecipitation and Western blot analysis.
Coimmunoprecipitation and Western Blotting
Lysates were centrifuged for 30 min at 4°C at 16,000 g. Supernatants were used for immunoprecipitation assays. Typically, 5 μg of antibody was added to 500 μg of protein lysate and incubated at 4°C for 1 h. 40 μl of protein-G agarose slurry was added and the incubation continued for another 1 h. The immune complexes on the beads were washed three times in mRIPA buffer and once in PBS. Beads were boiled in 2× SDS-PAGE buffer and samples were electrophoresed on 8% SDS-PAGE gels. Proteins were then transferred to nitrocellulose membranes (BA85; pore size, 0.45 μm) (Schleicher & Schuell). To verify the quality of transfer, proteins were visualized on the filters by staining with 0.2% Ponceau S in 3% trichloroacetic acid. To block nonspecific binding, the filters were incubated for 2 h at room temperature in 5% milk, TBST buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.1% Tween 20). Antigens were visualized by immunoblotting with the appropriate primary antibody (1:1,000 dilution), and then a horseradish peroxidase–conjugated anti–mouse IgG or anti–rabbit IgG antibody. All blots were developed using enhanced chemiluminescence reagents from Amersham Pharmacia Biotech.
Adhesion Assay
24-well plates were coated overnight at 37°C with 10 μg/ml vitronectin. Before the adhesion assay, wells were washed three times with PBS. 293-VnR cells stably expressing the Cbl constructs or control vector were harvested by scraping followed by repeated pipetting in serum-free medium to obtain a single-cell suspension. Cells were then washed and resuspended in serum-free medium at 106 cells/ml. 1 ml of suspension was then added to each well and incubated at 37°C for 30 min, after which nonadherent cells were gently washed off with PBS. The remaining adherent cells were quantified by staining with MTT (3-[4,5-dimethyiazol-2-yl]-2,5-diphenyltetrazolium bromide).
Kinase Assay
c-Src was immunoprecipitated from cells as described above and the Src kinase activity assayed using a kit from Upstate Biotechnology, as described in the manufacturer's protocol. To ensure equal immunoprecipitation of Src, aliquots of beads were also subjected to Western blot analysis.
Results
Deletion of Src Leads to Alterations in Adhesion Structures, Lamellipodia Motility and Cell Migration in Osteoclasts
Punctate actin- and vinculin-containing podosomes are typically organized in a ring-like structure at the periphery of osteoclasts attached to coverslips. To investigate the role of Src in podosome formation and localization, Src+ and Src− OCLs were plated on vitronectin-coated coverslips and allowed to spread for relatively brief periods; then the distribution of vinculin and actin was analyzed by confocal microscopy (Fig. 1 A). After 1 h, punctate actin and vinculin colocalization was apparent in the central regions of both Src+ and Src− OCLs, with more podosomes in Src− cells. In Src+ OCLs, actin and vinculin were also colocalized in forming lamellipodia, which were not apparent in Src− OCLs. After 2 h, Src+ OCLs exhibited an increase in the number and length of lamellipodia. In contrast, dense concentrations of punctate actin and vinculin were seen in the interior of the Src− OCLs at 2 h, with no apparent formation of lamellipodia.
Figure 1.

Src deletion leads to alterations in adhesion structures and cell migration. (A) Immunofluorescence showing actin (red) and vinculin (green) colocalization (yellow) in Src+ and Src− OCLs replated on vitronectin for 1 and 2 h. Micrographs are composites of both channels together. Note the formation of vinculin-rich lamellipodia (blue arrows, Src+ 1 and 2 h), which is not seen in Src− OCLs, and the increased redistribution of actin from the central regions of the cell (white arrows) to the peripheral regions of the cell (red arrow, Src+ 2 h) in Src+ OCLs. These features were not observed in Src− OCLs where actin and vinculin colocalization was observed to significantly increase between 1 and 2 h. (B) Immunofluorescence showing actin (red) and vinculin (green) localization in Src+ and Src− authentic osteoclasts cultured on glass coverslips. Note the distinct ring-like assembly of podosomes that stain for both actin and vinculin in Src+ osteoclasts (white arrows), and the focal adhesion-like arrangement of vinculin in surrounding contaminating fibroblast-like cells (red arrows, Src+ vinculin). In Src− osteoclasts, the localization of actin to the periphery of the osteoclast is not observed (yellow arrow, Src− actin), and staining for vinculin demonstrates a more focal-adhesion–like arrangement (red arrow, Src− vinculin) reminiscent of that seen in the previously described nonosteoclastic cells. (C and D) The lengths of cell migration paths of Src− (C) and c-Cbl− (D) osteoclasts and their littermate wild-type controls were measured at regular intervals as described in Materials and Methods.
At a much longer time after plating (16–24 h), Src+ osteoclasts showed a characteristic and consistent peripheral ring-like organization of podosomes (Fig. 1 B), distinctly different in appearance from the characteristic focal adhesion-like structures seen in the surrounding contaminating fibroblastic cells. In contrast, all Src− osteoclasts lacked this dense ring-like pattern, and these proteins were relatively depleted at the cell periphery. Instead of the punctate podosomal localization seen in freshly purified Src+ osteoclasts, actin and vinculin were organized in focal adhesion-like structures, particularly visible with vinculin staining. Furthermore, actin stress fibers running through the cytoplasm and anchoring to the plasma membrane at the site of the focal adhesion-like structures were frequently observed in Src− cells. These observations suggested that, in the absence of Src, the dynamic and rapid assembly and disassembly of podosomes was compromised, a change that could possibly translate into a reduction in cell motility.
We therefore analyzed the motility (movement of the peripheral cell membrane lamellipodia) and migration (distance moved) of Src+ and Src− osteoclasts plated on serum-coated coverslips. Quantification revealed a 40% decrease (P < 0.01) in spreading after 4 h of plating and a 25% decrease in motility (P < 0.01) compared with Src+ osteoclasts (data not shown). When cells were tracked for a 4-h time period to measure cell migration, a dramatic 60% decrease in the migration of Src− osteoclasts was evident (Fig. 1 C, P < 0.001). The ratio of cells whose total migration path was >80 μm was ∼80% for Src+ as compared with only 20% for Src− cells. In addition, no extended migration of >160 μm was observed in Src− osteoclasts, compared with 45% of Src+ cells migrating distances >160 μm over the 4-h period (data not shown). These results indicate that the absence of Src in osteoclasts disrupts the dynamic regulation of podosomes, and that this change is associated with a decrease in lamellipodia motility and cell migration.
c-Cbl− Osteoclasts also Show Decreased Motility
We have previously shown that Src was also necessary for the tyrosine phosphorylation of Cbl and its normal localization to podosomes in osteoclasts (Tanaka et al. 1996), suggesting that Cbl is downstream of Src in osteoclasts and involved in bone resorption. If so, then Cbl might also be involved in osteoclast motility. We therefore determined whether osteoclasts from Cbl− mice, like those from Src− mice, exhibited changes in their ability to migrate. Deletion of c-Cbl significantly reduced the migration of osteoclasts (Fig. 1 D): in four separate experiments using a minimum of 70 osteoclasts per experiment, the distances traveled by Cbl− osteoclasts was 27% less than those of Cbl+ cells (P < 0.05). This effect is significantly smaller than that induced by Src deletion and is apparently not sufficient to give a detectable skeletal phenotype (Murphy et al. 1998; and data not shown). The lack of phenotype is probably a consequence of redundancies with Cbl-b, the expression of which we found to be highly upregulated in c-Cbl–deficient osteoclasts (data not shown). Nevertheless, these results indicate that the Cbl family of proteins is indeed, like Src, involved in the process of osteoclast migration.
Signaling through Integrin Receptors and c-Src in Osteoclasts
The altered adhesion structures and decreased cell motility are likely to reflect changes in signaling downstream of integrins. To elucidate how Src might regulate osteoclast adhesion and motility, we first determined whether Src was activated by integrins in OCLs. For this purpose, we used an antibody that specifically recognizes phosphorylated tyrosine 416, the autophosphorylation site of murine c-Src (Thomas and Brugge 1997). αvβ3 integrins, the predominant integrin expressed in osteoclasts, were cross linked with antibodies against the murine VnR (Gailit et al. 1997), and the cells were lysed and analyzed. Phosphorylation of tyrosine 416 was detected within 3 min of integrin clustering in Src+ OCLs, reached a maximum at 10 min, and then returned to control levels by 20 min (Fig. 2 A), demonstrating that c-Src is first activated and later deactivated upon stimulation of the αvβ3 integrin in OCLs.
Figure 2.
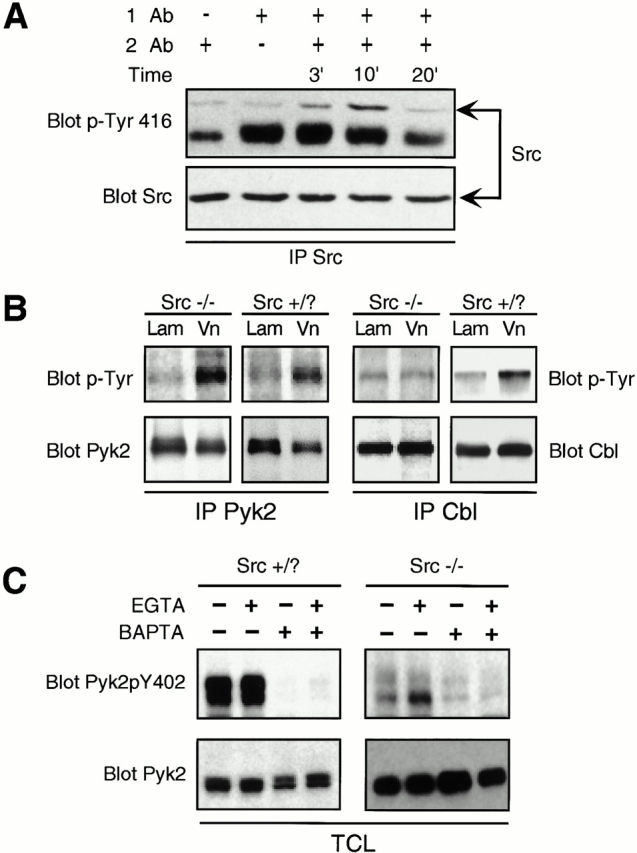
Integrin ligation activates Src and increases the tyrosine phosphorylation of a number of proteins, including c-Cbl, but not Pyk2, in a Src-dependent manner. (A) Src was immunoprecipitated from Src+ OCLs treated with VnR cross-linking antibodies and Western blotted with Src pTyr-416 antibody (top) that detects the activated form of Src. (Bottom) The same membrane stripped and reprobed with anti-Src Ab. (B) Pyk2 and Cbl immunoprecipitates from lysates of OCLs plated on laminin (Lam) or vitronectin (Vn) for 30 min were Western blotted with anti–p-Tyr antibody (top). Blots were stripped and reprobed with antibodies against Cbl or Pyk2 (bottom). (C) OCLs were pretreated with calcium chelators EGTA (100 μM) and/or BAPTA (50 μM), as described in Materials and Methods, and allowed to attach to vitronectin-coated plastic for 30 min. Total cell from lysates were Western blotted for Pyk2pY402 (top), then stripped and reprobed with anti–Pyk2 antibody (bottom).
Members of the FAK family are also activated by integrins, and Pyk2, a calcium-dependent FAK-related tyrosine kinase, is highly expressed in osteoclasts (Duong et al. 1998). To determine whether Pyk2 is tyrosine phosphorylated downstream of integrins in OCLs and whether this event occurs in a Src-dependent manner, we immunoprecipitated Pyk2 from lysates of Src+ and Src− OCLs that had been replated on vitronectin. After attachment of the cells, Pyk2 phosphorylation was increased in both Src+ and Src− cells (Fig. 2 B). In contrast, the tyrosine phosphorylation of Cbl was induced in Src+ cells but not in Src− cells by plating on vitronectin, confirming our previous observations (Tanaka et al. 1996). Neither Pyk2 nor Cbl were phosphorylated in response to adhesion to laminin, a ligand for the α2β1 integrin (Fig. 2 B). This is in agreement with Duong et al. 1998, who showed that Pyk2 does not get phosphorylated after cross linking of β1 or β2 integrins. Thus, both Pyk2 and Cbl are phosphorylated downstream of αvβ3 in OCLs, but only Cbl phosphorylation requires Src. This defect in Cbl phosphorylation was not a consequence of decreased spreading of the Src-deficient cells, since no differences in the spreading of Src+ and Src− cells could be detected (data not shown) at the time points used in these studies (up to 2 h).
Engagement of integrins also leads to increased intracellular Ca2+ levels (Sjaastad and Nelson 1997), a signal that could activate Pyk2 (Lev et al. 1995). Treatment of isolated osteoclasts with RGD-containing peptides produces transient increases in intracellular Ca2+ levels, which we found to be independent of the presence of Src (data not shown). Chelation of intracellular Ca2+ by BAPTA significantly decreased Pyk2 tyrosine phosphorylation in both Src+ and Src− OCLs (data not shown). In contrast, removal of extracellular Ca2+ by EGTA had no effect. Using antibodies to specifically detect Pyk2 phosphotyrosine 402, we determined that phosphorylation of this tyrosine was induced by adhesion and was also Ca2+, but not Src, dependent (Fig. 2 C). Thus, upon adhesion to ECM, the initial tyrosine phosphorylation at the autophosphorylation site Y402, reflecting the activation of Pyk2, results from changes in intracellular calcium levels and is not mediated by or dependent on Src. Since autophosphorylation of Pyk2 at Y402 creates a consensus binding site for the SH2 domain of Src (Dikic et al. 1996), we investigated whether Src, Pyk2, and possibly Cbl, which is known to interact with Src (Tanaka et al. 1996), were present in the same molecular complexes.
Interactions between Pyk2, Cbl and Src in Osteoclasts
Although Pyk2 and Cbl do not contain any presently identified domains that would mediate direct interaction, confocal microscopic analysis demonstrated that the two proteins colocalized in the podosome-rich periphery of osteoclasts as well as in the perinuclear Golgi regions (Fig. 3 A). Since both Pyk2 and Cbl can bind to Src via Src's SH2 and SH3 domains, respectively, we investigated the possibility that Src might act as an adaptor molecule mediating their association. Cbl coimmunoprecipitated with Pyk2 in Src+ OCLs, and this association was markedly decreased in Src− cells (Fig. 3 B). The residual association of Pyk2 and Cbl in the Src− OCLs is most likely due to the fact that other members of the Src kinase family are expressed in OCLs, and partially compensates for the absence of Src (Lowell et al. 1996). Thus, in osteoclasts, Pyk2 and Cbl are present in podosomes and associate via a mechanism that is predominantly Src dependent.
Figure 3.

Cbl associates with Pyk2 in a Src-dependent manner, and becomes tyrosine phosphorylated by Src in a reaction that also requires the presence of Pyk2. Immunocytochemistry showing Pyk2 and Cbl colocalization in Src+ authentic osteoclasts. Micrographs represent individual channels (red, Pyk2; green, Cbl) and a merging of the two channels. Pyk2 was immunoprecipitated from Src+ and Src− OCLs. Immune complexes were Western blotted for Cbl (top left), and then the membranes were stripped and reprobed with Pyk2 antibodies (bottom left) Total cell lysates (TCL) were also electrophoresed along with the immunoprecipitates. Reciprocal immunoprecipitation and blotting was performed using Cbl antibodies for immunoprecipitation and Pyk2 antibodies for blotting (top right). The membrane was stripped and reprobed with Cbl antibodies (bottom right).
Analysis of the Src/Pyk2/Cbl Complex in a Model System: Cbl Phosphorylation Downstream of the Vitronectin Receptor in 293 Cells Requires both Pyk2 and Src Kinase Activity
To analyze the molecular interactions between these three molecules in greater detail, we further examined the requirement for Pyk2 and Src in signaling downstream of the αvβ3 integrin using 293-VnR cells, HEK293 cells that were stably transfected with the vitronectin receptor (Chuntharapai et al. 1993), as a model system. In contrast to results obtained in osteoclasts or in other cell lines (Manie et al. 1997; Ojaniemi et al. 1997), we failed to observe Cbl tyrosine phosphorylation when 293-VnR cells were replated on vitronectin-coated dishes for up to 30 min, a time point at which we could demonstrate the vitronectin-induced phosphorylation of FAK, Cas, and paxillin (Fig. 4 A). Similar results were obtained after more prolonged incubations (data not shown). Cbl was tyrosine phosphorylated after treatment with EGF (data not shown), demonstrating that integrin-independent tyrosine phosphorylation of Cbl can occur in these cells. Interestingly, both FAK and Cbl are endogenously highly expressed in these cells, but coimmunoprecipitation experiments failed to demonstrate the association of Cbl and FAK in replated cells (Fig. 4 B). Thus, despite VnR-induced tyrosine phosphorylation events occurring in the presence of Cbl, Cbl tyrosine phosphorylation was not induced by integrin activation in 293-VnR cells.
Figure 4.
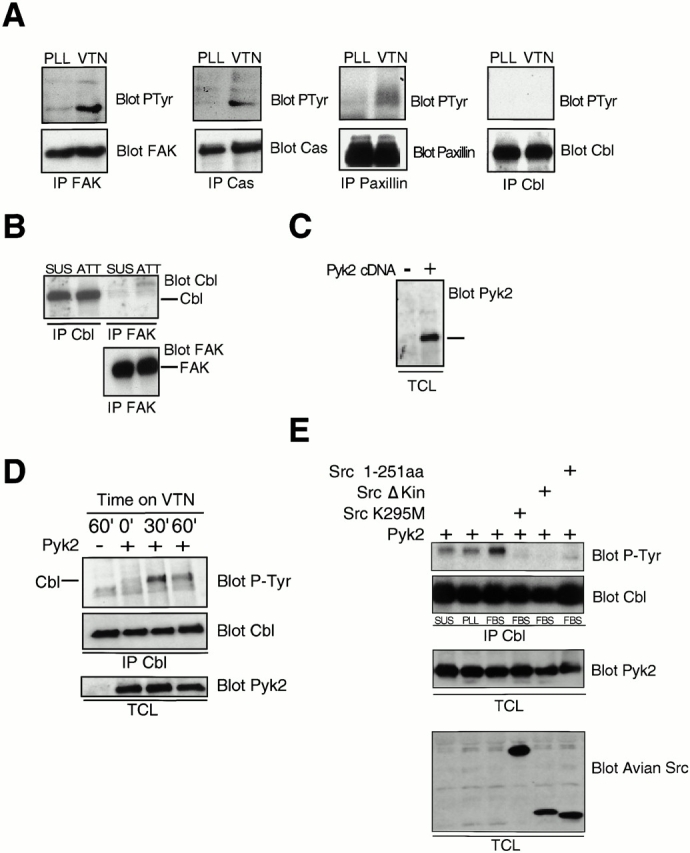
Cbl phosphorylation downstream of the vitronectin receptor requires Pyk2. (A) Serum-starved 293-VnR cells were replated on plastic coated with either polylysine (PLL) or vitronectin (VTN) for 30 min, and then lysed. FAK, Cas, paxillin, or Cbl were immunoprecipitated and analyzed by Western blotting for phosphotyrosine (top). The membranes were stripped and reprobed with antibodies against the immunoprecipitated antigen (bottom). (B) Cbl and FAK were immunoprecipitated from lysates of 293 VnR cells that had been replated on vitronectin-coated plastic (ATT) or kept in suspension (SUS), and the immune complexes analyzed by Western blotting for the presence of Cbl (top). (Bottom) Equal amounts of FAK in the FAK immunoprecipitates. (C) The total cell lysates (TCL) of 293 VnR cells transfected with vector control (−) or Pyk2 (+) were probed with Pyk2 antibodies to show expression of exogenous Pyk2 (+) (right). (D) 293-VnR cells transfected with 5 μg of either control vector (−) or Pyk2 (+) were harvested and either kept in suspension or plated on vitronectin-coated plastic for the indicated times. The cells were then lysed and Cbl was immunoprecipitated and Western blotted for P-Tyr (top). Membranes were then stripped and reprobed for Cbl (middle). Total cell lysates (TCL) were blotted for Pyk2 to ensure expression (bottom). (E) 293-VnR cells were transfected with a combination of 5 μg Pyk2 expression vector and 5 μg of cDNAs encoding various kinase-inactive Src mutants. The cells were harvested, and then either kept in suspension (SUS) or plated on plastic coated with polylysine (PLL) or serum (FBS) for 30 min. The cells were then lysed and Cbl was immunoprecipitated and Western blotted for p-Tyr (top). The membrane was reprobed for Cbl. Total cell lysate (TCL) blots were probed with antibodies to Pyk2 or avian Src to ensure expression.
These data suggested that specific protein(s) that are required for Cbl phosphorylation after adhesion and are present in some other cell types, including osteoclasts, were not expressed at sufficient levels in 293-VnR cells. One of the characteristics of osteoclasts, which exhibit adhesion-induced tyrosine phosphorylation of Cbl, is a high level of expression of Pyk2 rather than FAK (Levy et al. 1997; Duong et al. 1998). Examination of Pyk2 expression in 293-VnR cells showed that Pyk2 was expressed at extremely low levels in these cells (Fig. 4 C), suggesting that the absence of Pyk2 could be responsible for the failure of Cbl to be phosphorylated downstream of integrin engagement. We therefore determined the effect of transiently overexpressed Pyk2 on adhesion-induced Cbl phosphorylation, and found a time-dependent phosphorylation of Cbl in replated Pyk2-transfected 293-VnR cells (Fig. 4 D). Thus, the adhesion-induced tyrosine phosphorylation of Cbl downstream of the vitronectin receptor requires Pyk2.
To confirm that the adhesion-induced phosphorylation of Cbl was still dependent on Src in the 293-VnR cells that overexpressed Pyk2, we cotransfected 293-VnR cells with Pyk2 and one of a number of Src mutants/truncations that lacked kinase activity (Fig. 4 E). These dominant-negative Src mutants abolished Pyk2-dependent adhesion-induced Cbl phosphorylation, indicating that both Pyk2 and Src are required for adhesion-induced Cbl phosphorylation, and suggesting that Cbl phosphorylation is actually catalyzed by Src.
In 293 Cells, Pyk2 Is Autophosphorylated in an Adhesion- and Intracellular Calcium-dependent Manner
We then proceeded to characterize the specific molecular mechanism by which Pyk2 and Src mediated the adhesion-induced tyrosine phosphorylation of Cbl, confirming whenever possible that the model 293-VnR system was responding in a manner consistent with our findings in OCLs. To determine whether integrin-induced calcium signaling was activating Pyk2 in the 293-VnR cells as it does in OCLs, we examined the effects of BAPTA and EGTA on attachment-induced Pyk2 phosphorylation. As in OCLs (Fig. 2 C), chelation of intracellular Ca2+ by BAPTA significantly decreased Pyk2 tyrosine phosphorylation in replated cells, while removal of extracellular Ca2+ by EGTA had no affect (Fig. 5 A). Furthermore, and again as in OCLs, the attachment-induced phosphorylation of Pyk2 was not dependent on Src kinase activity since it was not inhibited by the overexpression of dominant-negative kinase-deficient or -deleted Src (Fig. 5 B). Thus, engagement of the αvβ3 integrin in the 293-VnR cells appears to induce Ca2+-dependent Pyk2 activation and autophosphorylation, as in untransfected OCLs.
Figure 5.

Pyk2 is autophosphorylated at Y402 after adhesion and associates with Src via the phosphorylated Y402. (A) 293-VnR cells transiently transfected with 5 μg of Pyk2 were treated with calcium chelators BAPTA and/or EGTA, and then replated on serum-coated plastic and allowed to attach to vitronectin-coated plastic for 30 min. Pyk2 IPs from lysates were Western blotted for P-Tyr (top), and then reprobed with anti–Pyk2 (bottom). (B) 293-VnR cells were transiently transfected with Pyk2 (5 μg) or cotransfected with Pyk2 (5 μg) and Src kinase-inactive mutants (10 μg). Pyk2 IPs from lysates were probed with P-Tyr antibodies (top). Membrane was stripped and reprobed with Pyk2 (bottom). (C) Pyk2 and Pyk2 mutants Y402F (PYF) and K457A (PKM) were transiently expressed in 293-VnR cells and total cell lysates (TCL) were blotted with an antibody specific for Pyk2 phosphorylated residue 402 (top). (Bottom) Transfected proteins. (D) Cells were processed as in A and Pyk2 IPs were blotted with an antibody specific for Pyk2 phosphorylated residue 402 (top). (Bottom) Pyk2 blot demonstrating equal loading. (E) 293-VnR cells were transiently transfected with wild-type Pyk2, kinase-dead Pyk2 (PKM) or Pyk2Y402F (PKF). The transfected cells were harvested, and then kept in suspension (SUS) or replated on serum-coated plastic (ATT) as in Fig. 4 E. Association of Src with mutant Pyk2 proteins was analyzed by Western blotting Src immunoprecipitates with Pyk2 antibodies (top). (Bottom) The membrane reprobed with Src antibodies. (F) Src IPs from cells treated with either EGTA or BAPTA were blotted with Pyk2 antibodies (top). (Bottom) The membrane reprobed with anti–Src. (G) 293-VnR cells transfected with 5 μg of Pyk2 or dominant-negative kinase-dead Pyk2 (PKM) were either kept in suspension or replated on vitronectin coated dishes for 30 min. Lysates were immunoprecipitated with anti–Src antibodies and probed with Src pTyr-416 antibody (top), which detects the activated form of Src. The membrane was stripped and reprobed with anti–Src (bottom). (H) Src kinase activity was quantified as an increase in pY416 Src using Scion Image1.62 C program.
To confirm that Pyk2 autophosphorylation was occurring, we examined the phosphorylation status of Y402. Phosphorylated Pyk2-Y402 was strongly detected in lysates from cells transfected with wild-type Pyk2, while only a very low level was present in lysates from cells transfected with the kinase-dead Pyk2K457A (PKM) mutant (Fig. 5 C), confirming that Y402 is largely phosphorylated by Pyk2 itself. Like the increase in total Pyk2 phosphorylation detected by the pan-phosphotyrosine antibody, the autophosphorylation of Y402 requires increased intracellular Ca2+ (Fig. 5 D). Thus, the increase in intracellular calcium that follows VnR binding during cell adhesion, both in untransfected OCLs and in Pyk2-transfected 293-VnR cells, leads to the activation of Pyk2 tyrosine kinase and its autophosphorylation on tyrosine 402.
Autophosphorylated Pyk2 Forms a Complex with Src and Cbl
Consistent with the prediction that the phosphorylation of tyrosine-402 in Pyk2 creates a consensus sequence for Src SH2 domain binding (Dikic et al. 1996), Src was coimmunoprecipitated with Pyk2 from lysates of attached cells, but not from lysates of cells kept in suspension (Fig. 5 E). Furthermore, and as predicted by our earlier results, the association of Src and Pyk2 was markedly diminished in cells cultured in the presence of BAPTA (Fig. 5 F) and in cells transfected with kinase-dead Pyk2 (PKM) or Pyk2 in which the autophosphorylation site was mutated (Pyk2Y402F or PYF) (E). Thus, VnR engagement induces the activation and autophosphorylation of Pyk2, resulting in the recruitment of Src to the adhesion site through an interaction of Src-SH2 and Pyk2 Tyr-402.
The binding of the autophosphorylated Pyk2Y402 to the Src SH2 domain will displace the inhibitory phosphotyrosine 527 of Src, which would be predicted to activate Src. To confirm that this is indeed the case in our model system, 293-VnR cells were transfected with wild-type Pyk2 or the kinase-dead mutant, PKM, and the adhesion-induced Src activation was assayed. As expected, replating the cells caused an increase in the level of autophosphorylated Src (approximately twofold) in cells transfected with wild-type Pyk2, but not in cells transfected with PKM or with the empty vector (Fig. 5G and Fig. H), indicating that Pyk2 kinase activity is required for the efficient adhesion-induced activation of Src.
We then determined whether Cbl, whose phosphorylation is dependent upon both the presence of Pyk2 and the kinase activity of Src (Fig. 4D and Fig. E), was present in the same molecular complex as Src and Pyk2. As shown in Fig. 6, Pyk2 and Cbl could be coimmunoprecipitated from lysates of replated cells. This association was independent of Src kinase activity; i.e., independent of Cbl's phosphorylation, since the overexpression of dominant-negative Src mutants that lacked kinase activity failed to block the association (Fig. 6 A).
Figure 6.
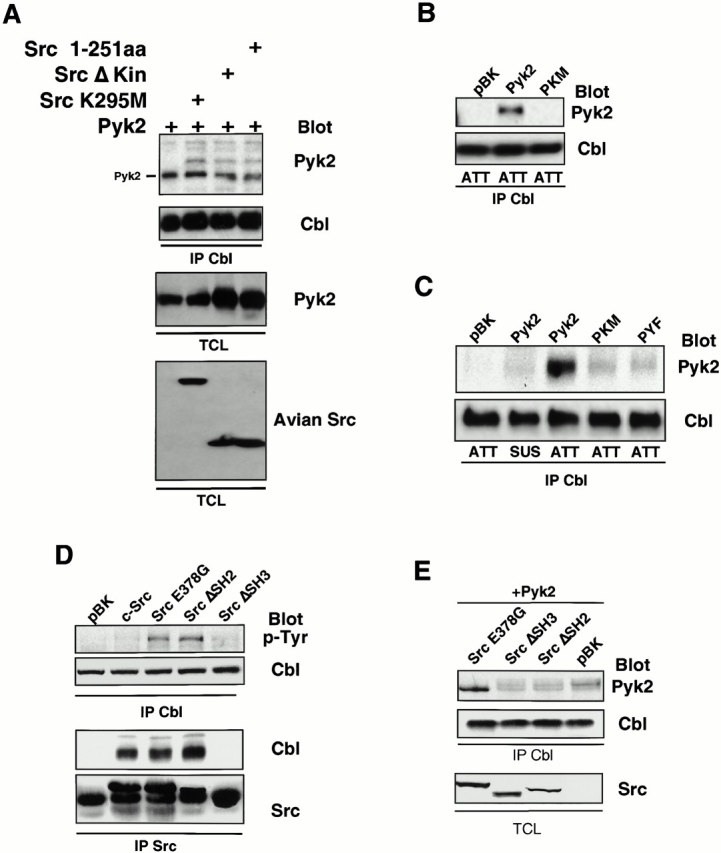
Src acts as an adaptor molecule mediating interactions between Cbl and Pyk2. (A) Association of Cbl with Pyk2 in presence of Src kinase mutants was analyzed by probing the blot of Cbl immunoprecipitates with Pyk2 antibodies (top). The blot was stripped and reprobed with Cbl antibodies. Total cell lysates (TCL) were Western blotted with Pyk2 and avian Src antibodies to check expression of transfected proteins. (B) 293-VnR cells were transfected with Pyk2, PKM, or empty vector (pBK). Cbl was immunoprecipitated from lysates of replated cells and analyzed for the presence of phosphotyrosine (top). (C) 293-VnR cells were transfected with Pyk2 mutants. The attachment-induced association of the mutant Pyk2 proteins with Cbl (top) was analyzed by blotting Cbl IPs with anti–Pyk2 antibody. (D) Cbl IPs from lysates of 293-VnR cells transfected with 10 μg of the indicated Src expression vector were blotted with an antiphosphotyrosine antibody to demonstrate the effects of these Src mutants of Cbl phosphorylation. Src was immunoprecipitated from the same lysates and the immune complexes were blotted for the presence of Cbl. (E) 293-VnR cells were transfected with a combination of 3 μg Pyk2 and 3 μg of the indicated Src expression vector. Cbl IPs were analyzed for the presence of Pyk2 (top). (Bottom) Expression of the Src mutants in total cell lysates (TCL).
Src Is an Adaptor Molecule Mediating the Interaction of Pyk2 and Cbl
Despite the presence of a number of protein binding domains in Pyk2 and Cbl, the two proteins lack known complementary domains that would mediate a direct interaction between them. They both bind to different Src domains, however, and we hypothesized that Src acts as an adaptor molecule mediating their association, with autophosphorylated Pyk2 Y402 binding to the Src SH2 domain and the Cbl proline-rich region binding to the Src SH3 domain. The adaptor role of Src was further supported by the fact that Src mutants in which the kinase domain or the whole COOH-terminal half were deleted could still associate with both Pyk2 and Cbl (Fig. 4 E and 6 A). The role of autophosphorylated Pyk2 Y402 was confirmed by the failure of either the kinase-dead PKM or the Pyk2Y402F mutant to associate with Src (Fig. 5 E) or with Cbl (Fig. 6B and Fig. C) in replated cells.
Finally, to verify the involvement of the Src SH2 and SH3 domains, we transfected 293-VnR cells with various mutants of Src, immunoprecipitated Src or Cbl, and analyzed the immunoprecipitates for the presence of Cbl and Pyk2, respectively. When Src was immunoprecipitated, Cbl was observed in all immune complexes except those from cells overexpressing Src ΔSH3 (Fig. 6 D), confirming that the SH3 domain of Src was necessary for the efficient association of these proteins. Cbl coimmunoprecipitated with Src independent of its kinase activity; i.e., with a constitutively active form of Src (E378G), which hyperphosphorylates Cbl, and a number of kinase dead/deleted Src mutants (data not shown), which are incapable of phosphorylating Cbl. Thus, although the Src kinase domain and its activity are required for Cbl phosphorylation, neither is necessary for the association between Cbl and Src.
The absence of either the SH2 or SH3 domains of Src resulted in a decrease in the amount of Pyk2 that coimmunoprecipitated with Cbl (Fig. 6 E), confirming that both domains of Src are necessary for the Src-mediated association of Pyk2 and Cbl, and suggesting that Pyk2 and Cbl were indeed associating via a single Src molecule. Thus, as in the OCLs (Fig. 3 B), Pyk2, Src, and Cbl form a trimolecular complex in the 293-VnR cells that is induced by the engagement and activation of the αvβ3 integrin. Subsequent Ca2+-dependent activation and autophosphorylation of Pyk2 leads to the recruitment and activation of Src via the Pyk2 pY402-Src SH2 interaction, while the proline-rich domain of Cbl binds to the Src SH3 domain.
Cbl Regulates Src Kinase Activity within the Complex
Cbl is known to be a negative regulator of several receptor and nonreceptor tyrosine kinases (Tanaka et al. 1995; Wang et al. 1996; Barber et al. 1997; Manie et al. 1997). We therefore hypothesized that while the binding of Cbl to the SH3 domain of Src might contribute to the full activation of Src kinase, as do other proline-rich ligands of the SH3 domains of several Src family members (Xu et al. 1999), the presence of Cbl in the complex might also lead to a secondary negative regulation. Thus, the presence of Cbl in the trimolecular complex could provide a mechanism to insure the transience of attachment-induced Src kinase activity. To determine whether Cbl in fact inhibited Src kinase activity, we transiently transfected a number of Cbl mutants, including the oncogenic 70Z-Cbl (missing part of the RING finger) and v-Cbl (truncation of the whole COOH-terminal half), into cells that overexpressed a constitutively active mutant of Src (Src Y527F). Full-length Cbl, 70Z-Cbl and v-Cbl inhibited the kinase activity of immunoprecipitated Src by 35, 28, and 37%, respectively, (P < 0.01, Fig. 7 A), comparable with the transfection efficiency (30–50%, as demonstrated using a green fluorescent protein expression vector) while transfection with empty vector or expression of the COOH-terminal half of Cbl had no effect.
Figure 7.

Cbl inhibits Src kinase activity by binding to Src. (A) NIH3T3 cells that stably express a constitutively active Src (SrcY527F) were transfected with 10 μg of the indicated Myc-tagged Cbl cDNA and expression was demonstrated by immunoblotting using an anti–Myc antibody (top). Src was immunoprecipitated and the kinase activity was measured and expressed as a percentage of the control kinase activity levels (pBK-transfected cells). The amount of immunoprecipitated Src in the kinase assay was assessed by immunoblotting. (B) 293-VnR cells that stably express the myc-tagged Cbl constructs (top) were transfected with 10 μg of active Src (SrcE378G). Src was immunoprecipitated and the immune complex was immunoblotted to detect myc-tagged proteins (middle) and Src (bottom).
Inhibition of Src Kinase by Cbl Is PTB Domain Dependent
The inhibition of Src kinase activity by v-Cbl suggested that the NH2 terminus of Cbl can bind to Src. We therefore sought to determine whether that is indeed true and, if so, to identify the requirements for such an interaction. As expected, full-length Cbl, 70Z-Cbl, and Cbl-COOH terminal, all of which contain the proline-rich domain and therefore bind to SH3, were present in Src immunoprecipitates (IPs). v-Cbl was also detected in Src IPs, albeit at lower levels, confirming that the NH2-terminal half of Cbl is capable of binding to Src (Fig. 7 B).
Since the PTB domain is the only functional domain identified in v-Cbl, we considered the possibility that this domain mediated the binding to Src and, possibly, the inhibition of its kinase activity. Interestingly, Src Y416, the autophosphorylated residue in Src's activation loop, occurs in the sequence DNEY, which, when phosphorylated on tyrosine, matches the preferred binding motif D(D/N)xpY for the Cbl PTB domain (Lupher et al. 1997), suggesting that the Cbl PTB domain might bind to phosphorylated Src Y416. We therefore sought to confirm that the Cbl PTB-Src pY416 interaction mediates the interaction of v-Cbl and Src.
A single point mutation, G306E, abolishes the binding ability of the PTB domain (Bonita et al. 1997). We therefore introduced this mutation into both c-Cbl and v-Cbl, and then determined the effect of the mutation on the binding of c-Cbl or v-Cbl to a constitutively active Src (SrcE378G). The G306E mutation had no effect on the association of full-length Cbl with Src (Fig. 8 A), consistent with our observation that the primary association of Src with Cbl is mediated by the proline-rich SH3 interaction. The mutation did, however, prevent the binding of v-Cbl to Src, confirming the involvement of Cbl's PTB domain in the v-Cbl–Src interaction.
Figure 8.
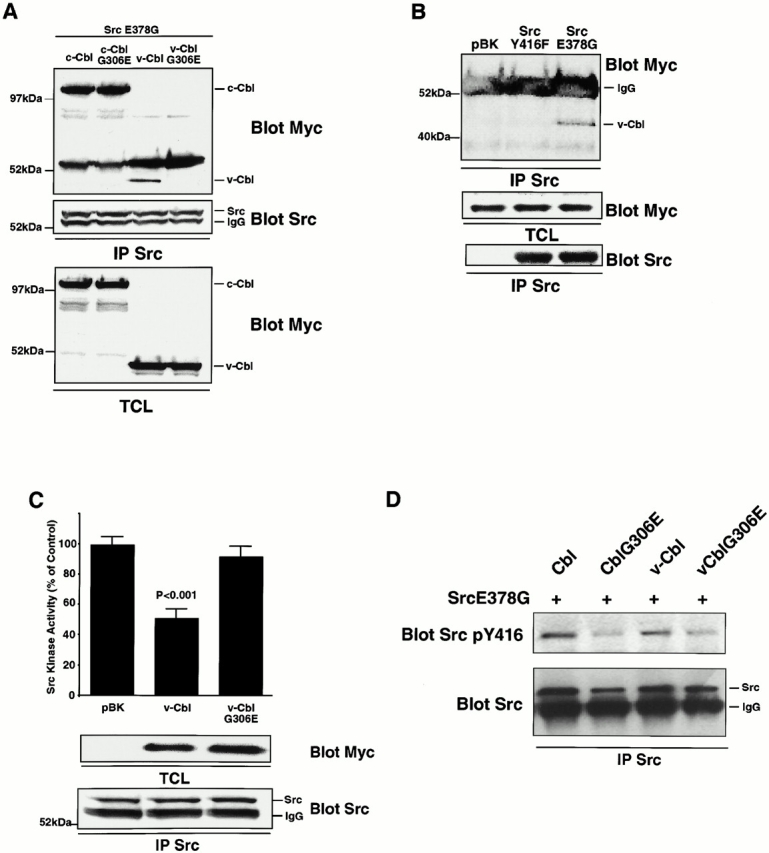
The association of the Cbl PTB domain with Src phosphotyrosine residue 416 results in an inhibition of Src kinase activity. (A) 293-VnR cells were transiently cotransfected with 5 μg Src E378G and 5 μg of normal or G306E-mutated forms of Cbl and v-Cbl. After immunoprecipitating Src, immune complexes were blotted for associated Cbl protein (top). The efficiencies of Src immunoprecipitation (middle) and Cbl expression (bottom) were determined. (B) 293-VnR v-Cbl cells were transiently transfected with active avian Src (SrcE378G) or avian Src Y416F. Avian Src was immunoprecipitated and blotted for the presence of myc-tagged v-Cbl. v-Cbl expression (middle) and efficiency of Src immunoprecipitation (bottom) were analyzed. (C) The kinase activity of immunoprecipitated Src from cells transfected with a combination of 5 μg SrcE378G and 5 μg of either v-Cbl or v-CblG306E was measured and expressed as a percentage of the control kinase activity levels (pBK-transfected cells). v-Cbl expression (middle) and efficiency of Src immunoprecipitation (bottom) were analyzed. (D) 293-Vnr cells were transiently cotransfected with 5 μg Src E378G and 5 μg normal or G306E mutated form of Cbl and v-Cbl. Src was immunoprecipitated and immune complex was immunoblotted with SrcY416 (top). The blot was reprobed with Src (bottom).
To confirm that the phosphotyrosine 416 was indeed the target of the PTB domain, coimmunoprecipitation experiments were performed with lysates from 293-VnR cells that stably express myc-tagged v-Cbl and were transiently transfected with Src E378G or Src Y416F. v-Cbl was present only in Src E378G immune complexes (Fig. 8 B), indicating that mutation of tyrosine 416 abolished the interaction between v-Cbl and Src in a manner similar to the G306E mutation in Cbl.
Finally, since the G306E mutation of Cbl abolished the binding of v-Cbl to Src, we examined the effect of the mutation on Cbl's inhibition of Src kinase activity. The mutation completely blocked the ability of overexpressed v-Cbl to inhibit the Src's kinase activity (Fig. 8 C), implying that while the initial and predominant interaction between Src and Cbl is likely via the Src SH3-Cbl proline-rich domains, the PTB domain of Cbl can subsequently bind to the autophosphorylation site on Src's activation loop to inhibit Src kinase activity. We also measured the kinase activity of activated Src in presence of c-Cbl and c-Cbl G306E proteins. Although the inhibition of Src kinase activity was not completely abolished by the mutation, as was the case with v-Cbl, the activity of Src from cells transfected with c-CblG306E was 66% greater than that of Src from cells transfected with Cbl (data not shown), confirming that the Cbl PTB/Src pY416 interaction contributes to the inhibition of Src by c-Cbl.
To further confirm that the Cbl PTB domain binds to the Src pY416, we characterized the effect of wild-type Cbl and CblG306E on the Src pY416 content, reasoning that binding to the PTB domain would protect the phosphotyrosine from phosphatase-catalyzed hydrolysis. Cells were cotransfected with SrcE378G and either c-Cbl, c-CblG306E, v-Cbl, or v-CblG306E, and then Src was immunoprecipitated and blotted for pY416. Mutation of both c-Cbl and v-Cbl resulted in less Src pY416 (Fig. 8 D), providing further support for the presence of an interaction between the Cbl PTB domain and phosphorylated Src Y416. Together, these data clearly demonstrate that Src and Cbl interact not only via Src SH3, but also through a Cbl PTB-Src kinase domain interaction. Most importantly, this latter interaction inhibits Src kinase activity. Thus, the recruitment of Cbl to Src serves to downregulate the kinase activity of Src, initially stimulated after integrin activation by the binding of both Pyk2 and Cbl to Src SH2 and SH3 domains, respectively.
Cbl also Inhibits Cellular Adhesion of 293 Cells to Vitronectin via its PTB Domain
Our results show that Cbl is recruited to a Pyk2- and Src-containing complex as a consequence of integrin engagement and activation, and that the association of Cbl with this complex leads to decreased Src kinase activity. Since Cbl regulates β1 integrin-mediated cell adhesion downstream of the CD28 cytoplasmic domain (Zell et al. 1998), we next sought to determine whether Cbl could regulate the VnR-mediated adhesion of 293-VnR cells. 293-VnR cells were stably transfected with various Cbl constructs, and the adhesion of the different cell lines to vitronectin was tested. All Cbl constructs that contained the PTB domain significantly inhibited the number of cells that adhered to vitronectin (Fig. 9), without affecting the expression of either subunit of the VnR (data not shown). In contrast, Cbl-C and empty vector controls had no significant effect. Interestingly, no inhibition of adhesion to fibronectin was observed (data not shown), suggesting that this effect of Cbl was specific for the VnR. Interestingly, the c-Cbl G306E mutant had the opposite effect (i.e., it led to a marked and significant increase in cell adhesion) (144%, P < 0.001) (Fig. 9), suggesting that a Cbl mutant that binds to the Src SH3 domain but cannot bind Src pY416 has a dominant-negative effect on the modulation of cell adhesion by endogenous c-Cbl, possibly by preventing the negative regulation of Src kinase. Taken together, these results suggest that Cbl molecules that contain a functional PTB domain inhibit both the activity of Src kinase and the adhesion of cells to vitronectin, which further suggests that the strength of VnR-matrix interactions may be directly or indirectly (i.e., via Src) modulated by Cbl.
Figure 9.

Cbl regulates vitronectin receptor–mediated 293-VnR adhesion. The adhesion of 293-VnR cells stably expressing a number of Cbl constructs was measured as described in Materials and Methods and expressed as a percentage of control (untransfected 293-VnR cells). The graph represents pooled data from two individual experiments.
Discussion
Cyclic attachment and detachment of individual podosomes is required as cells, particularly highly motile cells such as macrophages and osteoclasts, migrate over a substratum. Our data suggest that Pyk2, Src, and Cbl may play a pivotal role in these processes. First, we have demonstrated that deletion of Src significantly alters the initial distribution of podosomes in osteoclasts, eventually leading to the formation of focal adhesion-like structures. This shift from podosome to focal adhesion correlates with a decrease in the formation and motility of lamellipodia and in cell migration, although it is not possible to determine which comes first. We find that adhesion to vitronectin or activation of the αvβ3 receptor induces an increase in intracellular calcium that does not depend on the presence of Src. Using both osteoclasts and transfected 293-VnR cells in which this pathway has been reconstructed, we also show that Pyk2 is autophosphorylated at tyrosine 402 in a Src-independent, [Ca2+]i-dependent manner, creating a binding site for the SH2 domain and recruiting Src. Cbl is then recruited to this complex, binding to the SH3 domain of Src, or, alternatively, it may be constitutively bound to inactive Src but be phosphorylated only after Src is activated by binding to Pyk2. In either case, the formation of a kinase-rich trimolecular complex, which we observed in both OCLs and 293-VnR cells, results in the Pyk2- and Src-dependent phosphorylation of Cbl. Therefore, Pyk2, Src, and Cbl are all involved in the “outside-in” αvβ3 integrin signaling which, we propose, results in podosome assembly. In addition, we have also demonstrated that, in transfected 293-VnR cells, the PTB domain of Cbl binds to Tyr 416 in the activation loop of the Src kinase domain, and that this interaction downregulates both Src kinase activity and integrin-mediated adhesion. Thus, Cbl might also be crucial in “inside-out” signaling, playing a key role in podosome detachment and subsequent disassembly. In agreement with this hypothesis, we also found that deletion of the gene encoding Cbl, like the deletion of the gene encoding Src, leads to a significant, albeit smaller, decrease in osteoclast migration. The series of events described above could therefore form the basis for the cyclic attachment–detachment of single adhesion sites at the leading edge of lamellipodia in motile cells, and thereby participate in the assembly–disassembly of individual podosomes, thereby ensuring cell adhesion while still allowing cell motility.
This interpretation is consistent with the existing literature. It is known that, unlike FAK, Pyk2 can be activated by an increase in intracellular calcium (Lev et al. 1995), and we show here that such an event occurs in both osteoclasts and 293-VnR cells upon αvβ3 integrin ligation. We also confirm here that this results in the autophosphorylation of tyrosine 402, which generates a binding site for the SH2 domain of Src (Dikic et al. 1996). The binding of phosphotyrosine motifs to the Src family SH2 domains is known to cause a conformational change resulting in the partial activation of the Src kinase and autophosphorylation of a tyrosine in the activation loop (Xu et al. 1999), and we indeed demonstrate that integrin ligation leads to the phosphorylation of tyrosine 416 in Src. We also found that Cbl binds to the SH3 domain of Src. As demonstrated by Xu et al. 1999, the binding of proteins to both the SH3 and SH2 domains of Src family members induces the complete activation of their kinase. The binding of Cbl to Src results in the tyrosine phosphorylation of Cbl, which we demonstrate here is Src-dependent downstream of the αvβ3 integrin.
One major finding in this study is that Cbl plays a pivotal role in this complex, not only by binding to the Src SH3 domain and possibly thereby contributing to the activation of Src, or as an adaptor recruiting more proteins to the complex, but also as a kinase inhibitor, providing a mechanism that might ensure that the activity of Src kinase within the complex at the attachment site is transient. We show here that this function involves the binding of the Cbl PTB domain to the Src phosphotyrosine 416. Interestingly the PTB domain also modulates VnR-mediated attachment, regulating adhesion to vitronectin but not fibronectin. Furthermore, the G306E mutation of the PTB creates a form of c-Cbl that acts in a dominant-negative manner on the c-Cbl-induced downregulation of cell adhesion, providing further evidence that Cbl may regulate cell adhesion and that this occurs, at least in part, via the interaction of its PTB domain with Src Tyr-416. This specific inside-out effect of Cbl could be the result of the inhibition of Src kinase activity, of a direct modulation of integrin binding, of an indirect modulation via other tyrosine kinases or the cytoskeleton, or a combination of effects. Interestingly, our findings and those of Felsenfeld et al. 1999 show that both Src and Cbl negatively regulate the adhesion of VnR to vitronectin, whereas adhesion to fibronectin is unaffected. This suggests a certain level of specificity of this molecular complex for the vitronectin receptor, possibly linked to the recruitment and calcium-dependent activation of Pyk2 as opposed to FAK. Further work will be required to address these questions.
Cbl's inhibition of several other tyrosine kinases, including EGFR, PDGFR, and Syk has been reported (Ota and Samelson 1997; Lupher et al. 1998; Waterman et al. 1999). In all cases, Cbl's PTB domain has been shown to be responsible for Cbl's recruitment and binding. However, our findings illustrate the fact that Cbl can also be recruited to the activated tyrosine kinase Src via the SH3 domain and that it is the specific binding of Cbl's PTB domain to the activation loop in Src's kinase domain that is responsible for inhibition of the kinase activity. Nearly all other members of the Src family possess the Cbl PTB binding consensus motif (DNEY) at their autophosphorylation site and, therefore, by analogy to Src, could also be inhibited by association with Cbl. Although v-Cbl was able to associate with Src in a cellular overexpression system, the association of the COOH-terminal half of Cbl (containing the proline-rich domain) was observed to be much greater, so that association of full-length Cbl with Src was unaffected by the G306E mutation. It therefore seems likely that, under normal conditions (i.e., endogenous levels of protein expression), the Src SH3 interaction with the Cbl proline-rich domain occurs first and that the interaction between the Cbl PTB domain and the autophosphorylated tyrosine occurs only secondarily, possibly only transiently and as a consequence of Cbl phosphorylation. This represents a novel mechanism by which Cbl might modulate Src tyrosine kinase-dependent signaling events and VnR-mediated cell adhesion and motility. The smaller decrease in the motility of Cbl− cells (relative to Src− cells) is probably due to the partial compensation for the absence of c-Cbl in osteoclasts by Cbl-b, which contains identical functional domains (Keane et al. 1995), is overexpressed in c-Cbl− OCLs, and coimmunoprecipitates with Pyk2 from OCL lysates (our unpublished observations).
In models using genetically modified cells, the short-term strength of adhesive contacts has been demonstrated to regulate migration rates as net traction forces result from an unequal distribution of adhesive contacts between the leading and trailing edges (Lauffenburger and Horwitz 1996; Palecek et al. 1998). The adhesive strength of integrin-mediated substrate contacts, and thus migration rates, can be modulated by altering either integrin–ligand affinity, cell-surface integrin expression levels, substrate–ligand concentrations, or integrin–cytoskeletal coupling (Palecek et al. 1997). Interestingly, the forces generated by the cytoskeleton are increased in the absence of Src (Felsenfeld et al. 1999), and this results in decreased spreading (Kaplan et al. 1995) and, as shown here, decreased motility. Our data suggest that the motility defect exists before the formation of focal adhesions, since all motility measurements were made using osteoclasts within 4 h of initial attachment, a time frame in which we do not yet see the formation of stress fibers and focal adhesions in Src− OCLs, but rather observe a defect in the localization of podosomes. Since we found podosomes to be more numerous in freshly plated OCLs in the absence of Src, we propose that Src deletion affects podosome disassembly rather than assembly.
We further suggest that the apparent stronger attachment of Src-deleted cells (Felsenfeld et al. 1999) results from Cbl failure to be recruited and/or phosphorylated at the adhesion sites. This view is supported by the fact that the absence of Src causes a significant reduction in the amount of Cbl associated with Pyk2 and with podosomes at the periphery of the cell (Tanaka et al. 1996). As in Felsenfeld et al. 1999, the recruitment of Cbl is apparently independent of Src's kinase activity, suggesting that Src's role in this process is primarily as an adaptor protein. A role for Src as an adaptor molecule has been suggested previously by the rescue of the Src− osteopetrotic phenotype by osteoclast-specific expression of kinase-deficient Src mutants (Schwartzberg et al. 1997). Thus, tyrosine phosphorylation of Cbl, which requires an active Src kinase, may not be critical for podosome assembly and/or disassembly. It should, however, be noted that Src251 was not able to rescue and that the K295M kinase-dead Src only partially rescued the phenotype. The best rescue was obtained with SrcY416F (Schwartzberg et al. 1997), which retains some kinase activity, albeit less than wild-type Src. Interestingly, our model would predict that SrcY416F cannot be downregulated by Cbl since it lacks the Cbl PTB binding site. The fact that the kinase activity of this mutant is low may have made further downregulation unnecessary for a sufficient degree of cell motility. Hence, the most likely interpretation is that some Src kinase activity is required for efficient assembly and/or disassembly of podosomes and, thereby, bone resorption.
Thus, we propose that the Src-dependent recruitment of Cbl to adhesion sites acts to modulate the binding of the vitronectin receptor. Failure to recruit either Src or Cbl, in Src− or Cbl− osteoclasts, respectively, results in a decrease in lamellipodia motility and cell migration. The observations reported here may therefore explain, at least in part, the osteopetrotic phenotypes of Src-deleted (Soriano et al. 1991) and Pyk2-deleted (Sims et al. 1999) mice.
It should be emphasized here that Pyk2 and Cbl recruit other essential proteins to the Pyk2/Src/Cbl complex, modulating cytoskeletal interactions and the turnover of adhesion structures, although for the sake of simplification we did not incorporate this information in our model. We have shown that Cbl is able to homodimerize in a leucine zipper-dependent manner (Bartkiewicz et al. 1999) so that the recruitment of Cbl to sites of adhesion would create a large number of potential binding sites for other proteins such as PI3 kinase (Miyake et al. 1997). Similarly, Pyk2 has been shown to be constitutively associated with p130Cas in OCLs (Palecek et al. 1997). More intriguing is our recent observation that the RING finger of Cbl recruits the ubiquitin-conjugating enzyme UbCH7 (Yokouchi et al. 1999) to EGF-induced molecular complexes, leading to the ubiquitination and degradation of the activated EGFR complex. Thus, Cbl recruitment to the podosomes might also bring about ubiquitination, and subsequently degradation, of one or more adhesion-related proteins. Indeed, activation-dependent ubiquitination of Src has recently been reported in other systems. Further experiments will be required to determine whether these events also play a part in the regulation of adhesion structures and cell motility.
Acknowledgments
The authors thank Dr. Sarah Bodary (Genentech, San Francisco, CA) for providing the 293-VnR cell line, to Dr. A. Laudano for providing the antibodies to Y416 of Src, and to Karen Ford for maintaining the Src-deficient colony of mice.
This work was supported by National Institutes of Health grant AR42927 (National Institute of Arthritis and Musculoskeletal and Skin Diseases, and Office of Research on Women's Health) and by a grant from Ariad Pharmaceuticals (R. Baron). Both A. Houghton and A. Sanjay are recipients of Arthritis Council Postdoctoral Fellowships.
Footnotes
Drs. Sanjay and Houghton contributed equally to this work and should be considered co-first authors.
Abbreviations used in this paper: ECM, extracellular matrix; IP, immunoprecipitate; mRIPA, modified radioimmune precipitation assay; OCL, osteoclast-like cell; VnR, vitronectin receptor.
References
- Barber D.L., Mason J.M., Fukazawa T., Reedquist K.A., Druker B.J., Band H., D'Andrea A.D. Erythropoietin and interleukin-3 activate tyrosine phosphorylation of CBL and association with CRK adaptor proteins. Blood. 1997;89:3166–3174. [PubMed] [Google Scholar]
- Bartkiewicz M., Houghton A., Baron R. Leucine zipper-mediated homodimerization of the adaptor protein c-Cbla role in c-Cbl's tyrosine phosphorylation and its association with epidermal growth factor receptor. J. Biol. Chem. 1999;274:30887–30895. doi: 10.1074/jbc.274.43.30887. [DOI] [PubMed] [Google Scholar]
- Bonita D.P., Miyake S., Lupher M.L., Jr., Langdon W.Y., Band H. Phosphotyrosine binding domain-dependent upregulation of the platelet-derived growth factor receptor α signaling cascade by transforming mutants of Cblimplications for Cbl's function and oncogenicity. Mol. Cell. Biol. 1997;17:4597–4610. doi: 10.1128/mcb.17.8.4597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broome M.A., Galisteo M.L., Schlessinger J., Courtnedge S.A. The protooncogene c-Cbl is a negative regulator of DNA synthesis initiated by both receptor and cytoplasmic tyrosine kinases. Oncogene. 1999;18:2908–2912. doi: 10.1038/sj.onc.1202873. [DOI] [PubMed] [Google Scholar]
- Chen W.T. Proteolytic activity of specialized surface protrusions formed at rosette contact sites of transformed cells. J. Exp. Zool. 1989;251:167–185. doi: 10.1002/jez.1402510206. [DOI] [PubMed] [Google Scholar]
- Chen W.T. Transmembrane interactions at cell adhesion and invasion sites. Cell Differ. Dev. 1990;32:329–336. doi: 10.1016/0922-3371(90)90047-z. [DOI] [PubMed] [Google Scholar]
- Chuntharapai A., Bodary S., Horton M., Kim K.J. Blocking monoclonal antibodies to αVβ3 integrina unique epitope of αVβ3 integrin is present on human osteoclasts. Exp. Cell Res. 1993;205:345–352. doi: 10.1006/excr.1993.1096. [DOI] [PubMed] [Google Scholar]
- Dikic I., Tokiwa G., Lev S., Courtneidge S.A., Schlessinger J. A role for Pyk2 and Src in linking G-protein–coupled receptors with MAP kinase activation. Nature. 1996;383:547–550. doi: 10.1038/383547a0. [DOI] [PubMed] [Google Scholar]
- Duong L.T., Lakkakorpi P.T., Nakamura I., Machwate M., Nagy R.M., Rodan G.A. PYK2 in osteoclasts is an adhesion kinase, localized in the sealing zone, activated by ligation of αvβ3 integrin, and phosphorylated by Src kinase. J. Clin. Invest. 1998;102:881–892. doi: 10.1172/JCI3212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duong L.T., Rodan G.A. The role of integrins in osteoclast function. J. Bone Miner. Metab. 1999;17:1–6. doi: 10.1007/s007740050055. [DOI] [PubMed] [Google Scholar]
- Felsenfeld D.P., Schwartzberg P.L., Venegas A., Tse R., Sheetz M.P. Selective regulation of integrin-cytoskeleton interactions by the tyrosine kinase Src. Nat. Cell Biol. 1999;1:200–206. doi: 10.1038/12021. [DOI] [PubMed] [Google Scholar]
- Gailit J., Clarke C., Newman D., Tonnesen M.G., Mosessen M.W., Clark R.A.F. Human fibroblasts bind directly to fibrinogen at RGD sites through integrin αVβ3. Exp. Cell Res. 1997;232:118–126. doi: 10.1006/excr.1997.3512. [DOI] [PubMed] [Google Scholar]
- Horne W.C., Neff L., Chatterjee D., Lomri A., Levy J.B., Baron R. Osteoclasts express high levels of pp60c-srcin association with intracellular membranes. J. Cell Biol. 1992;119:1003–1013. doi: 10.1083/jcb.119.4.1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joazeiro C.A.P., Wing S.S., Huang H.-K., Leverson J.D., Hunter T., Liu Y.-C. The tyrosine kinase negative regulator c-Cbl as a RING-type, E2-dependent ubiquitin-protein ligase. Science. 1999;286:309–312. doi: 10.1126/science.286.5438.309. [DOI] [PubMed] [Google Scholar]
- Kaplan K.B., Swedlow J.R., Morgan D.O., Varmus H.E. c-Src enhances the spreading of src−/− fibroblasts on fibronectin by a kinase-independent mechanism. Genes Dev. 1995;9:1505–1517. doi: 10.1101/gad.9.12.1505. [DOI] [PubMed] [Google Scholar]
- Keane M.M., Rivero-Lezcano O.M., Mitchell J.A., Robbins K.C., Lipkowitz S. Cloning and characterization of cbl-ba SH3 binding protein with homology to the c-cbl proto-oncogene. Oncogene. 1995;10:2367–2377. [PubMed] [Google Scholar]
- Lauffenburger D.A., Horwitz A.F. Cell migrationa physically integrated molecular process. Cell. 1996;84:359–369. doi: 10.1016/s0092-8674(00)81280-5. [DOI] [PubMed] [Google Scholar]
- Lev S., Moreno H., Martinez R., Canoll P., Peles E., Musacchio J.M., Plowman G.D., Rudy B., Schlessinger J. Protein tyrosine kinase PYK2 involved in Ca2+-induced regulation of ion channel and MAP kinase functions. Nature. 1995;376:737–745. doi: 10.1038/376737a0. [DOI] [PubMed] [Google Scholar]
- Levkowitz G., Waterman H., Ettenberg S.A., Katz M., Tsygankov A.Y., Alroy I., Lavi S., Iwai K., Reiss Y., Ciechanover A. Ubiquitin ligase activity and tyrosine phosphorylation underlie suppression of growth factor signaling by c-Cbl/Sli-1. Mol. Cell. 1999;4:1029–1040. doi: 10.1016/s1097-2765(00)80231-2. [DOI] [PubMed] [Google Scholar]
- Levy J.B., DiDomenico E., Neff L., Dikic I., Schlessinger J., Baron R. The FAK-related kinase, Pyk2, is tyrosine phosphorylated in response to integrin stimulation in osteoclasts. J. Bone Miner. Res. 1997;12:S109. [Google Scholar]
- Longhurst C.M., Jennings L.K. Integrin-mediated signal transduction. Cell. Mol. Life Sci. 1998;54:514–526. doi: 10.1007/s000180050180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lowe C., Yoneda T., Boyce B.F., Chen H., Mundy G.R., Soriano P. Osteopetrosis in Src-deficient mice is due to an autonomous defect of osteoclasts. Proc. Natl. Acad. Sci. USA. 1993;90:4485–4489. doi: 10.1073/pnas.90.10.4485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lowell C.A., Niwa M., Soriano P., Varmus H.E. Deficiency of the Hck and Src tyrosine kinases results in extreme levels of extramedullary hematopoiesis. Blood. 1996;87:1780–1792. [PubMed] [Google Scholar]
- Lowell C.A., Soriano P. Knockouts of Src-family kinasesstiff bones, wimpy T cells, and bad memories. Genes Dev. 1996;10:1845–1857. doi: 10.1101/gad.10.15.1845. [DOI] [PubMed] [Google Scholar]
- Lupher M.L., Jr., Songyang Z., Shoelson S.E., Cantley L.C., Band H. The Cbl phosphotyrosine-binding domain selects a D(N/D)XpY motif and binds to the Tyr292 negative regulatory phosphorylation site of ZAP-70. J. Biol. Chem. 1997;272:33140–33144. doi: 10.1074/jbc.272.52.33140. [DOI] [PubMed] [Google Scholar]
- Lupher M.L., Jr., Rao N., Lill N.L., Andoniou C.E., Miyake S., Clark E.A., Drucker B., Band H. Cbl-mediated negative regulation of the Syk tyrosine kinase. A critical role for Cbl phosphotyrosine-binding domain binding to Syk phosphotyrosine 323. J. Biol. Chem. 1998;273:35273–35281. doi: 10.1074/jbc.273.52.35273. [DOI] [PubMed] [Google Scholar]
- Manie S.N., Sattler M., Astier A., Phifer J.S., Canty T., Morimoto C., Druker B.J., Salgia R., Griffin J.D., Freedman A.S. Tyrosine phosphorylation of the product of the c-cbl protooncogene is induced after integrin stimulation. Exp. Hematol. 1997;25:45–50. [PubMed] [Google Scholar]
- Marchisio P.C., Cirillo D., Naldini L., Primavera M.V., Teti A., Zambonin-Zallone A. Cell-substratum interaction of cultured avian osteoclasts is mediated by specific adhesion structures. J. Cell Biol. 1984;99:1696–1705. doi: 10.1083/jcb.99.5.1696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marchisio P.C., Cirillo D., Teti A., Zambonin-Zallone A., Tarone G. Rous sarcoma virus-transformed fibroblasts and cells of monocytic origin display a peculiar dot-like organization of cytoskeletal proteins involved in microfilament-membrane interactions. Exp. Cell Res. 1987;169:202–214. doi: 10.1016/0014-4827(87)90238-2. [DOI] [PubMed] [Google Scholar]
- Meng F., Lowell C.A. A β1 integrin signaling pathway involving Src-family kinases, Cbl and PI-3 kinase is required for macrophage spreading and migration. EMBO (Eur. Mol. Biol. Organ.) J. 1998;17:4391–4403. doi: 10.1093/emboj/17.15.4391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyake S., Lupher M.L., Jr., Andoniou C.E., Lill N.L., Ota S., Douillard P., Rao N., Band H. The Cbl protooncogene productfrom an enigmatic oncogene to center stage of signal transduction. Crit. Rev. Oncog. 1997;8:189–218. doi: 10.1615/critrevoncog.v8.i2-3.30. [DOI] [PubMed] [Google Scholar]
- Murphy M.A., Schnall R.G., Venter D.J., Barnett L., Bertoncello I., Thien C.B.F., Langdon W.Y., Bowtell D.D. Tissue hyperplasia and enhanced T-cell signalling via ZAP-70 in c-Cbl-deficient mice. Mol. Cell. Biol. 1998;18:4872–4882. doi: 10.1128/mcb.18.8.4872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nermut M.V., Eason P., Hirst E.M.A., Kellie S. Cell/substratum adhesions in RSV-transformed rat fibroblasts. Exp. Cell Res. 1991;193:382–397. doi: 10.1016/0014-4827(91)90111-7. [DOI] [PubMed] [Google Scholar]
- Ochoa G.-C., Slepnev V.I., Neff L., Ringstad N., Takei K., Daniell L., Kim W., Cao H., McNiven M., Baron R., De Camili P. A functional link between dynamin and the actin cytoskeleton at podosomes. J. Cell Biol. 2000;150:377–389. doi: 10.1083/jcb.150.2.377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ojaniemi M., Martin S.S., Dolfi F., Olefsky J.M., Vuori K. The proto-oncogene product p120cbl links c-Src and phosphatidylinositol 3′-kinase to the integrin signaling pathway. J. Biol. Chem. 1997;272:3780–3787. doi: 10.1074/jbc.272.6.3780. [DOI] [PubMed] [Google Scholar]
- Ota Y., Samelson L.E. The product of the proto-oncogene c-cbla negative regulator of the Syk tyrosine kinase. Science. 1997;276:418–420. doi: 10.1126/science.276.5311.418. [DOI] [PubMed] [Google Scholar]
- Palecek S.P., Loftus J.C., Ginsberg M.H., Lauffenburger D.A., Horwitz A.F. Integrin-ligand binding properties govern cell migration speed through cell-substratum adhesiveness. Nature. 1997;385:537–540. doi: 10.1038/385537a0. [DOI] [PubMed] [Google Scholar]
- Palecek S.P., Huttenlocher A., Horwitz A.F., Lauffenburger D.A. Physical and biochemical regulation of integrin release during rear detachment of migrating cells. J. Cell Sci. 1998;111:929–940. doi: 10.1242/jcs.111.7.929. [DOI] [PubMed] [Google Scholar]
- Rodan S.B., Rodan G.A. Integrin function in osteoclasts. J. Endocrinol. 1997;154:S47–S56. [PubMed] [Google Scholar]
- Schwartzberg P.L., Xing L., Hoffmann O., Lowell C.A., Garrett L., Boyce B.F., Varmus H.E. Rescue of osteoclast function by transgenic expression of kinase-deficient Src in src−/− mutant mice. Genes Dev. 1997;11:2835–2844. doi: 10.1101/gad.11.21.2835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sieg D.J., Ilic D., Jones K.C., Damsky C.H., Hunter T., Schlaepfer D.D. Pyk2 and Src-family protein-tyrosine kinases compensate for the loss of FAK in fibronectin-stimulated signaling events but Pyk2 does not fully function to enhance FAK− cell migration. EMBO (Eur. Mol. Biol. Organ.) J. 1998;17:5933–5947. doi: 10.1093/emboj/17.20.5933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sims N.A., Aoki K., Bogdanovic Z., Maragh M., Okigaki M., Logan S., Neff L., DiDomenico E., Sanjay A., Schlessinger J., Baron R. Impaired osteoclast function in Pyk2 knockout mice and cumulative effects in Pyk2/Src double knockout. J. Bone Miner. Res. 1999;14:S183. [Google Scholar]
- Sjaastad M.D., Nelson W.J. Integrin-mediated calcium signaling and regulation of cell adhesion by intracellular calcium. Bioessays. 1997;19:47–55. doi: 10.1002/bies.950190109. [DOI] [PubMed] [Google Scholar]
- Soriano P., Montgomery C., Geske R., Bradley A. Targeted disruption of the c-src proto-oncogene leads to osteopetrosis in mice. Cell. 1991;64:693–702. doi: 10.1016/0092-8674(91)90499-o. [DOI] [PubMed] [Google Scholar]
- Stickel S.K., Wang Y.L. Alpha-actinin-containing aggregates in transformed cells are highly dynamic structures. J. Cell Biol. 1987;104:1521–1526. doi: 10.1083/jcb.104.6.1521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka S., Takahashi N., Udagawa N., Sasaki T., Fukui Y., Kurokawa T., Suda T. Osteoclasts express high levels of pp60c-src, preferentially on ruffled border membranes. FEBS Lett. 1992;313:85–89. doi: 10.1016/0014-5793(92)81190-w. [DOI] [PubMed] [Google Scholar]
- Tanaka S., Neff L., Baron R., Levy J.B. Tyrosine phosphorylation and translocation of the c-Cbl protein after activation of tyrosine kinase signaling pathways. J. Biol. Chem. 1995;270:14347–14351. doi: 10.1074/jbc.270.24.14347. [DOI] [PubMed] [Google Scholar]
- Tanaka S., Amling M., Neff L., Peyman A., Uhlmann E., Levy J.B., Baron R. c-Cbl is downstream of c-Src in a signalling pathway necessary for bone resorption. Nature. 1996;383:528–531. doi: 10.1038/383528a0. [DOI] [PubMed] [Google Scholar]
- Tarone G., Cirillo D., Giancotti F.G., Comoglio P.M., Marchisio P.C. Rous sarcoma virus-transformed fibroblasts adhere primarily at discrete protrusions of the ventral membrane called podosomes. Exp. Cell Res. 1985;159:141–157. doi: 10.1016/s0014-4827(85)80044-6. [DOI] [PubMed] [Google Scholar]
- Thomas S.M., Brugge J.S. Cellular functions regulated by Src family kinases. Annu. Rev. Cell. Dev. Biol. 1997;13:513–609. doi: 10.1146/annurev.cellbio.13.1.513. [DOI] [PubMed] [Google Scholar]
- Wang Y., Yeung Y.G., Langdon W.Y., Stanley E.R. c-Cbl is transiently tyrosine-phosphorylated, ubiquitinated, and membrane-targeted following CSF-1 stimulation of macrophages. J. Biol. Chem. 1996;271:17–20. doi: 10.1074/jbc.271.1.17. [DOI] [PubMed] [Google Scholar]
- Waterman H., Levkowitz G., Alroy I., Yarden Y. The RING finger of c-Cbl mediates desensitization of the epidermal growth factor receptor. J. Biol. Chem. 1999;274:22151–22154. doi: 10.1074/jbc.274.32.22151. [DOI] [PubMed] [Google Scholar]
- Xu W., Doshi A., Lei M., Eck M.J., Harrison S.C. Crystal structures of c-Src reveal features of its autoinhibitory mechanism. Mol. Cell. 1999;3:629–638. doi: 10.1016/s1097-2765(00)80356-1. [DOI] [PubMed] [Google Scholar]
- Yokouchi M., Kondo T., Houghton A., Bartkiewicz M., Horne W.C., Zhang H., Yoshimura A., Baron R. Ligand-induced ubiquitination of the epidermal growth factor receptor involves the interaction of the c-Cbl RING finger and UbcH7. J. Biol. Chem. 1999;274:31707–31712. doi: 10.1074/jbc.274.44.31707. [DOI] [PubMed] [Google Scholar]
- Zell T., Warden C.S., Chan A.S.H., Cook M.E., Dell C.L., Hunt S.W., III, Shimizu Y. Regulation of β1-integrin-mediated cell adhesion by the Cbl adaptor protein. Curr. Biol. 1998;8:814–822. doi: 10.1016/s0960-9822(98)70323-9. [DOI] [PubMed] [Google Scholar]