

Abstract
L-Selectin mediates rolling of lymphocytes in high endothelial venules (HEVs) of peripheral lymph nodes (PLNs). Cross-linking of L-selectin causes proteolytic shedding of its ectodomain, the physiological significance of which is unknown. To determine whether L-selectin shedding regulates lymphocyte migration, a mutant form that resists shedding (LΔP-selectin) was engineered. Transgenic mice expressing either LΔP or wild-type (WT) L-selectin on T cells were crossed with L-selectin knockout (KO) mice. The cellularity and subset composition of secondary lymphoid organs did not differ between LΔP and WT mice, however, they were different from C57BL/6. Plasma levels of soluble L-selectin in LΔP mice were reduced to <5% of WT and C57BL/6 mice. The rolling properties of T lymphocytes from LΔP and WT mice on immobilized L-selectin ligands were similar. Furthermore, similar numbers of LΔP and WT T lymphocytes were recruited from the bloodstream into PLNs in mice, although LΔP T cells transmigrated HEVs more slowly. WT, but not LΔP-selectin, underwent rapid, metalloproteinase-dependent shedding after TCR engagement, and LΔP T cells retained the capacity to enter PLNs from the bloodstream. These results suggest that the ability to shed L-selectin is not required for T cell recirculation and homing to PLNs. However, L-selectin shedding from antigen-activated T cells prevents reentry into PLNs.
Keywords: lymphocyte homing receptors, leukocyte rolling, vascular endothelium, matrix metalloproteinases, mice
Introduction
Naive lymphocytes constantly recirculate through lymphoid organs via the bloodstream and lymphatics to maximize encounters with antigen-presenting cells and initiate immune responses (1). Migration from the bloodstream into LNs is regulated by a multistep adhesion cascade, which involves different adhesion receptors and tissue-specific chemokines in the capture and arrest of lymphocytes from flowing blood (2, 3). L-Selectin mediates the tethering and rolling of naive lymphocytes in specialized high endothelial venules (HEVs) of peripheral LNs (PLNs), and rolling cells are arrested after activation of LFA-1 integrin by the CCR7 ligand secondary lymphoid chemokine (CCL21) expressed on the surface of HEVs (4–6). PLNs in L-selectin KO mice are smaller than their WT counterparts, and L-selectin KO T cells are unable to home to PLNs, confirming the critical role of L-selectin in lymphocyte trafficking (7). As a consequence, peripheral immune responses to antigens derived subcutaneously are suppressed or delayed in L-selectin KO mice, although systemic responses are not impaired (8–10).
Leukocyte rolling velocity is regulated by rapid formation and subsequent dissociation of bonds between L-selectin and its ligands (11). It has been demonstrated that ectodomain shedding of L-selectin is an important parameter in leukocyte (particularly neutrophil) rolling behavior in vitro and in vivo. Inclusion of synthetic hydroxamic acid–based matrix metalloproteinase (MMP) inhibitors, such as KD-IX-73-4, that block L-selectin shedding, reduces the rolling velocity, increases the number of cells that tether from flow, and increases the transit time through blood vessels in mice (12, 13). Further analyses using MMP inhibitors demonstrated that shedding regulated L-selectin–dependent β2-integrin activation in vitro and ICAM-1–dependent activation induced arrest in vivo in mice (14) and neutrophil-dependent inflammation in rats (15).
Like neutrophils, lymphocytes continually shed L-selectin, and shedding is accelerated by a variety of stimuli, including PMA and cross-linking by ligand mimetics (16–18). Constitutive and activation-induced shedding are both blocked by hydroxamic acid–based MMP inhibitors (19–22). A critical role for the level of L-selectin expression on naive lymphocytes in regulating migration into PLNs has been demonstrated previously (23). L-Selectin is rapidly down-regulated from the T cell surface within 30–60 min after antigen receptor engagement (16) and antigen-activated T cells that have migrated into nonlymphoid organs are generally L-selectinlow (24–26), but the precise contribution of L-selectin shedding to determining L-selectin expression patterns and trafficking of T cells has not been determined.
In this paper, we have generated transgenic mice that express a noncleavable form of L-selectin on T lymphocytes. We show here that the ability to shed L-selectin is not required for T cell recirculation and homing to LNs. However, L-selectin shedding after antigen receptor engagement prevents the reentry of activated T cells into PLNs.
Materials and Methods
Animals.
L-Selectin KO mice (7) were backcrossed onto the C57BL/6 background for >10 generations, recombination activating gene 1 (RAG-1) KO mice (27) were maintained on the C57BL/10 (H-2b) background, F5 TCR transgenic mice (F5) were maintained on the C57BL/10 background (28), and F5/RAG-1 KO mice (29) were bred and maintained under specific pathogen-free conditions at the National Institute for Medical Research. F5 mice were backcrossed to the L-selectin KO to generate L-selectin–deficient F5 mice (F5/L-selectin KO).
Construction of L-Selectin Mutants.
Domain deletions, domain swaps, and site-specific mutations were generated using PCR methods. WT L-selectin cDNA was generated by RT-PCR from BALB/c lymphocyte total RNA and cloned into the vector pEE6hCMV/neo (30) to give pEE6/WT. The 24-base deletion (ΔK-N eight–amino acid deletion) from the membrane proximal region (MPR) was engineered by overlap extension PCR to give pEE6/ΔK-N. A region of cDNA encoding the MPR of WT L-selectin was replaced with that encoding the mouse P-selectin equivalent by subcloning in two annealed synthesized complementary oligonucleotides to give pEE6/LΔP. The sequences of WT, ΔK-N, and LΔP cDNAs were verified by automated sequencing.
Generation of L-Selectin Transgenic Mice.
The strain-specific epitope Ly22 in the EGF domain of mouse L-selectin (31) provided a convenient marker to distinguish transgenic from endogenous L-selectin. The strains used to generate transgenic mice (CBA/Ca and C57BL/6) and WT L-selectin cDNA (BALB/c) are all Ly22+. Ala184 (GCA) was changed to Glu (GAA) by site-directed mutagenesis. The resultant Ly22− PCR product corresponding to a HindIII–ApaI fragment (encoding the lectin and part of the EGF domain) was subcloned into pEE6/WT, pEE6/ΔK-N, and pEE6/LΔP to give Ly22− vectors. Loss of the Ly22 allotype was confirmed by transient expression of WT, ΔK-N, and LΔP selectins in COS cells and loss of mAb T28 staining of MEL-14–positive transfectants (unpublished data). An XbaI recognition sequence was introduced immediately downstream of the HindIII sequence, and the Ly22− L-selectin cDNAs were subcloned into the human CD2 promoter and locus control region containing vector hCD2 (32) between XbaI and EcoRI to generate hCD2/WT, hCD2/ΔK-N, and hCD2/ LΔP. The cDNA sequences were verified by automated sequencing. The hCD2/WT/ΔK-N/LΔP digested fragments were separated from plasmid vector by gel electrophoresis, extracted from agarose, and further purified with Elutip columns. Transgenic mice were generated by independent microinjection of constructs into (CBA × C57BL/10)F2 fertilized eggs using standard procedures. Founder transgenic mice were identified by Southern blotting. Two independent lines for each transgene were crossed with C57BL/10 and H-2b offspring crossed to L-selectin KO mice. Inheritance of the transgene into the L-selectin KO background was determined by flow cytometry of PBL for MEL-14+/T28− staining and offspring were backcrossed to L-selectin KO mice for at least four generations. In some experiments, L-selectin transgenic mice were crossed with F5/L-selectin KO to generate mice coexpressing transgenic F5 TCR and either F5/WT or mutant (F5/LΔP) L-selectin.
Flow Cytometric Analysis.
Single cell suspensions from pooled PLNs (axillary, brachial, inguinal, and cervical), spleen, mesenteric LNs (MLNs), Peyer's patches, thymus, and bone marrow of single mice were prepared in PBS–calcium and magnesium free at 4°C. RBCs in heparinized blood and spleen suspensions were lysed using 0.83% NH4Cl. In some experiments, down-regulation of L-selectin, which occurs during leukocyte isolation (33), was prevented by including 30 μM Ro31-9790 (Roche) in all buffers used to prepare lymphocyte suspensions and removed by washing. Cells were incubated with Ab for 20 min at 4°C, washed twice, and incubated for 20 min with a second layer. After washing, immunofluorescence was detected by flow cytometry (FACSCalibur™ or FACSVantage™), data acquired for at least 1–10 × 106 viable cells and analyzed using WinMDI software (Joseph Trotter, The Scripps Research Institute, La Jolla, CA). Abs used were as follows. FITC, PE or biotin-CD62L (MEL-14, rat IgG2a), biotin- or APC-CD45R (B220, rat IgG2a), FITC-CD19 (1D3, rat IgG2a), biotin-TCRβ (H57-597, Armenian hamster IgGλ), biotin-I-Ab (25-9-17, mouse IgG2a), and biotin–Pan-NK cells (DX5, rat IgM; BD Biosciences); PE-TCRβ (H57-597, Armenian hamster IgG; Southern Biotechnology Associates, Inc.); and anti–mouse APC-Ly6G (Gr-1, rat IgG2b), PE-CD4 (CT-CD4, rat IgG2a), and APC-CD8α (CT-CD8a, rat IgG2a; Caltag Laboratories). Biotinylated KT-11 (anti-Vβ11; reference 34) was a gift from D. Kioussis (National Institute for Medical Research, London, UK). mAb T28, which recognizes the Ly22 allotype in mouse L-selectin, was a gift from U. Hammerling (Sloan-Kettering Memorial Cancer Center, New York, NY). LAM-116 (mouse Ig G2a) mAb, which recognizes a conserved sequence in mouse and human L-selectin (35), was purchased from Ancell. Biotin-conjugated Abs were detected with streptavidin-PE (Southern Biotechnology Associates, Inc.) for three-color staining, and with streptavidin-Cy7 (Jackson ImmunoResearch Laboratories) or streptavidin red–670 (Invitrogen) for four-color staining. Stainings were always performed with samples from age-matched mice.
Purification of T Lymphocytes.
T lymphocytes were enriched from spleen and LN cell suspensions by depletion of NK cells, MHC II–positive cells, and B cells. 2 × 108 cell/ml cell suspensions were incubated with biotinylated anti–mouse PAN-NK cells, I-Ab, and CD45R (1.5 μg/ml) at 4°C for 30 min, washed twice, and incubated with M-280 streptavidin Dynabeads according to manufacturer's instructions (Dynal). Beads were removed using a magnet and cells were washed twice in PBS containing 1% FCS. The purity of T lymphocytes was >94% by staining with TCRβ mAb.
L-Selectin Shedding Assay.
PLN lymphocytes (1 × 108 cells/ml) from WT and LΔP transgenic mice and purified T cells from C57BL/6 mice were either stored at 4°C or incubated at 37°C for 60 min in the absence or presence of 300 nM PMA and excess PMA was removed by washing. Constitutive shedding was determined by comparing L-selectin expression on cells held at 4°C with cells incubated at 37°C and activation-induced shedding by comparing cells incubated in the presence and absence of PMA. Supernatants were collected and analyzed for the presence of soluble L-selectin (sL-selectin) by ELISA. To induce L-selectin shedding from the surface of T cells expressing the F5 TCR, cells from PLNs, spleen, or peripheral blood (5 × 107 cell/ml) were incubated at 37°C for 60 min in the presence of 0.1 μM NP-68 peptide (NP-[366–374]; ASNENMDAM) from nucleoprotein of influenza virus A/NT/60/68 (28) and L-selectin expression on F5-positive CD8 T cells determined by dual staining for L-selectin and either CD8α or Vβ11 chain of the F5 TCR.
Measurement of sL-Selectin by ELISA.
To measure soluble L-selectin transgenic mice, Ly22− L-selectin mAb LAM-116, which recognizes the lectin domain (35) and is nonoverlapping with mAb MEL-14, was used in place of mAb T28 in a sandwich ELISA as described previously (21).
Flow Assays.
9 cm2 Slide Flaskettes (Nunc) were spotted with 50 μl of 200 μg/ml yeast polyphosphomonoester core polysaccharide (PPME; provided C. Parish, University of Canberra, Canberra, Australia), incubated overnight at 4°C and incubated with 1% BSA in PBS for 2 h at room temperature. Slides were washed twice with PBS/0.05% Tween 20 before use and mounted in a parallel plate flow chamber (channel height, 0.15 cm). 106/ml lymphocytes in HBSS/2% FCS (viscosity 0.007 Poise) were perfused at 37°C over the substrate at a shear stress of 1.5 dyn/cm2. Where specified, lymphocytes were preincubated with 10 μg/ml MEL-14 or MAC193 mAbs for 15 min at room temperature before inclusion of mAbs in the flow assay or 5 mM EGTA was included during flow assays. Experiments were visualized as described previously (36) using an inverted fluorescence microscope (Diaphot 300; Nikon) connected to a color video camera and recorded on a video recorder (JVC TK-C1360B; Panasonic AG-6730 S-VHS; Microscope Service and Sales). 10 random fields were recorded for 15 s each using a 10× objective (800 × 600 mm2). Images were acquired into a video file (In Video PCI; Focus Enhancements) at 15 frames/s, and numbers of cells undergoing rolling and/or arrest, rolling velocity of individual cells, and mean velocity of the population were calculated using EML Motion Analysis software (Ed Marcus Laboratories). Rolling cells were defined as those with a mean velocity from 3 to 75 μm/s for at least 1 s and adhered cells were specified as those moving <2 μm in 10 s. Mean rolling velocities were calculated from measurements of at least 200 cells in each independent experiment.
In Vivo Trafficking Experiments.
The migration of T cells expressing transgenic L-selectins were compared directly in individual mice using different fluorescent labels to track the two populations in vivo, as described previously (37, 38). Pooled PLN cells from L-selectin transgenic mice (>90% T lymphocytes), or from F5/L-selectin transgenic mice (>85% CD8+ T cells) were used without purification. T cells were purified from PLNs and spleen of C57BL/6 or L-selectin KO mice. Lymphocyte populations to be compared were labeled either with carboxyfluorescein diacetate-succinimidyl ester (CFSE; Molecular Probes) or 5-(and 6)-([{4-chloromethyl}benzoyl]amino) tetramethylrhodamine (CMTMR; Molecular Probes). For labeling, cells were either incubated in 2 μM CFSE in PBS or in 2 μM CMTMR in RPMI 1640 (Invitrogen) for 15 min at 37°C in humidified atmosphere of 5% CO2, washed twice in PBS containing 1% FCS or in RPMI 1640, respectively. CMTMR-labeled cells were incubated in RPMI 1640 for another 20 min at 37°C. Cells were counted and mixed to give a 1:1 ratio of CFSE/CMTMR-labeled cells in the start population. Labeled cells were injected in 0.2 ml PBS into tail veins of 8–12-wk-old RAG-1 KO (20 × 106 cells) or C57BL/6 (40 × 106 cells) recipient mice (at least three age- and sex-matched recipients per experiment). After 1 h, recipient mice were killed by CO2 inhalation, and PLNs, spleen, and heparinized blood were collected. Single cell suspensions were prepared from PLNs and spleen, and the percentages of CFSE- and CMTMR-labeled cells in the total population of gated live lymphocytes in PLNs, spleen, and blood were determined by flow cytometry. The localization ratio was calculated from the percentages of CFSE- and CMTMR-labeled cells in each organ. Where the ratio of CFSE/CMTMR-labeled cell in the injected (start) population was not equal to 1, a correction coefficient was applied to normalize localization ratios and results expressed as means ± SE of three to seven recipient mice.
To study the migration of antigen-activated lymphocytes, T cells from PLNs of F5/WT and F5/LΔP transgenic mice were incubated in 0.1 μM NP-68 peptide for 60 min at 37C°, washed twice with IMDM containing 1% FCS, labeled with either CFSE or CMTMR, and injected into RAG-1 KO mice. The localization ratios of T cells in the PLNs, spleen, and blood were determined by flow cytometry after staining of F5-expressing T cells for CD8α or Vβ11.
Confocal Microscopy.
PLNs (axillary, brachial, or inguinal) from individual mice were fixed in 2% formaldehyde/5% sucrose in PBS for 2 h, transferred to 20% sucrose in PBS for 2 h, snap frozen in liquid nitrogen, and stored at −70C°. Acetone-fixed 8-μm cryostat sections of PLNs were stained with MECA 79 (rat IgM; American Type Culture Collection) and Cy-5–conjugated goat anti–rabbit Ig (Jackson ImmunoResearch Laboratories) to detect HEVs of PLNs. The position of CFSE (green)- and CMTMR (red)-labeled cells in relation to HEVs (blue) was visualized by fluorescence microscopy using a TCS SP1 confocal laser scanning microscope (40× 1.25 PL APO objective; Leica). One PLN from each of recipient was analyzed after collection of 10–13 images each containing 1–3 HEVs (20–25 HEVs/LNs). Images were analyzed using TCS NT-1.6.587 software (Leica) and Semper6w kernel software (Synoptics) was used to correct autofluorescence (unpublished data). Labeled cells attached to the luminal surface and within the HEV wall were recorded as “Inside HEVs” (see Fig. 7, arrows). The remaining labeled cells were scored as “Outside HEVs” (see Fig. 7, arrowheads). The total numbers of CFSE and CMTMR-labeled cells were counted, and the ratio of LΔP/WT cells inside and outside HEVs was calculated for each PLN. The ratio of CFSE/CMTMR-labeled cells in the injected population was determined by flow cytometry and a correction coefficient was applied to normalize localization data. Results are mean localization ratios ± SE (n = 3–6).
Figure 7.
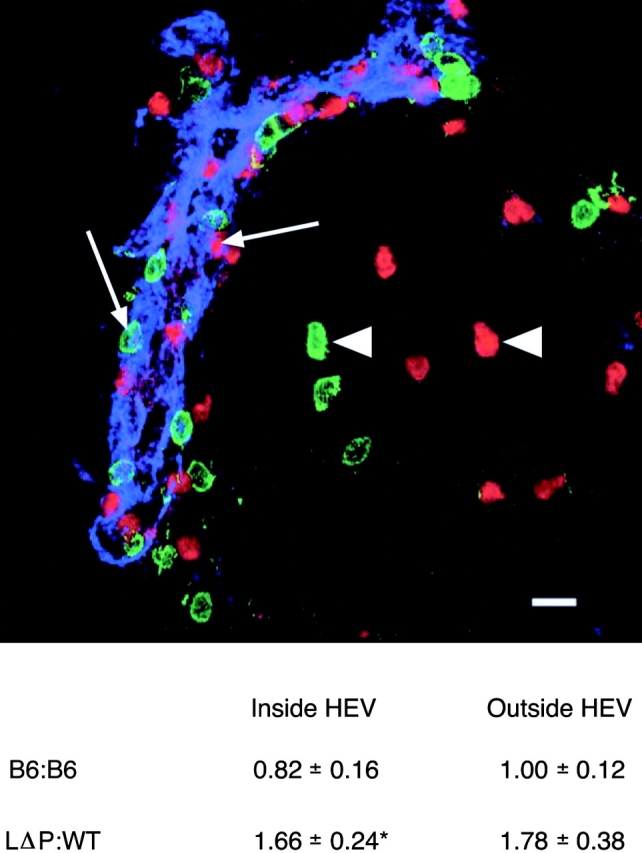
LΔP-Selectin–expressing lymphocytes migrate more slowly across HEVs into PLNs. HEVs in sections of PLNs from trafficking experiments described in Fig. 6 were stained with mAb MECA 79 (blue) and CFSE (green)- and CMTMR (red)-labeled T lymphocytes recorded as either Inside (arrows) or Outside (arrowheads) HEVs. A representative image shows the localization of CFSE (WT)- and CMTMR (LΔP)-labeled cells after injection into RAG-1 KO mice. The total numbers and localization of WT and LΔP lymphocytes were determined, and the ratio of LΔP/WT lymphocytes Inside and Outside HEVs was calculated. Results are pooled from five RAG-1 KO mice and one C57BL/6 recipient mouse. The total numbers of labeled cells counted were as follows: Inside HEVs, WT, 236, LΔP, 316; Outside HEVs, WT, 216, LΔP, 286. PLNs from mice that had been injected with equal numbers of C57BL/6 (B6) T cells labeled with either CFSE or CMTMR were also analyzed. The total numbers of CFSE- and CMTMR-labeled B6 cells counted were as follows: Inside HEVs, 60 and 77; and Outside HEVs, 165 and 182, respectively. The ratio of CFSE/CMTMR-labeled B6 cells was calculated and results pooled from three B6 recipient mice. Results are means ± SE. *, P < 0.05. Bar, 10 μm.
Statistical Methods.
All data presented are means ± SE. Localization ratios in trafficking experiments were compared with a theoretical ratio of one using paired Student's t test. Rolling velocities and numbers of rolling cells were compared using paired Student's t test. Other comparisons were made using unpaired Student's t test. Statistical significance was set at P < 0.05.
Results
Construction of Mouse L-Selectin Shedding Mutants and Generation of Transgenic Mice.
Proteolysis of L-selectin occurs within the MPR and a primary cleavage site has been mapped to K283S284 in human (39) and R283S284 in mouse (22) L-selectin (Fig. 1) . We generated two mutants of mouse L-selectin in which the MPR has been either truncated (ΔK-N, analogous to ΔM-N in human L-selectin; reference 40) or substituted with that of P-selectin (LΔP) and compared them with WT L-selectin. The Ly22 epitope, which is recognized by mAb T28, was deleted from cDNA's encoding WT, ΔK-N, and LΔP-selectin by site-directed mutagenesis and transgenic mice were generated using the hCD2 expression cassette. Two independent lines for each transgene were established by crossing founder mice to C57BL mice, which are Ly22+. Offspring were screened by flow cytometric analysis of PBLs, transgenic L-selectin being distinguished by the lack of T28 staining of MEL-14+ cells, and backcrossed to C57BL/6 L-selectin KO mice for at least four generations. PMA induced rapid shedding of endogenous L-selectin (C57BL/6) and transgenic WT L-selectin; however, both ΔK-N and LΔP transgenic L-selectins resisted PMA-induced shedding when incubated with up to 1 μM PMA for 1 h (unpublished data).
Figure 1.

Construction of mouse L-selectin shedding mutants. Schematic representation of the MPR of mouse L-selectin in relation to the whole molecule (SCR, short consensus repeat). Location of a primary cleavage site in wild-type L-selectin (WT) is shown together with amino acid sequences of corresponding regions of ΔK-N and LΔP mutants.
Inability to Shed L-Selectin Does Not Affect Cellularity or Subset Composition of LNs.
In PLNs of WT and LΔP transgenic mice, L-selectin expression was restricted to αβTCR+ T cells (Fig. 2 A). The percentage of T cells in PLNs of both WT and LΔP mice increased from 70% in C57BL/6 to >90% and B cell numbers were reduced to 5%, although the total yield of lymphocytes from PLNs did not differ between L-selectin transgenics and C57BL/6 (Table I). L-Selectin was largely restricted to αβTCR+ T cells in the spleens of WT and LΔP mice, although a small population of non-αβTCR cells also expressed transgenic L-selectin (∼3% of splenocytes). This population was distinguishable from the majority of B-220+ B cells, which did not express transgenic L-selectins. Gr-1+ myeloid cells in the bloodstream and granular cells in the bone marrow (Gr-1+ and Gr-1−) also did not express transgenic L-selectins (Fig. 2 A and not depicted). The total numbers and T/B subset composition of lymphocytes in spleens, MLNs, and Peyer's patches did not differ significantly between WT and LΔP transgenic mice and were similar to those in C57BL/6 (Table I). The ratios of CD4+/CD8+ T cells in PLNs, MLNs, spleen, and blood did not differ significantly between WT and LΔP mice (unpublished data). In accordance with activity of the CD2 cassette used (41), transgenic L-selectin was expressed early in thymocyte development at the double-negative stage and was sustained throughout development; thymocyte yield and subset composition were not altered in L-selectin transgenic mice (unpublished data).
Figure 2.


Transgenic L-selectin expression is restricted to T lymphocytes. L-Selectin expression on lymphocytes and peripheral blood leukocytes in either wild-type (WT) or mutant (LΔP) L-selectin transgenic mice was determined by flow cytometry using two- or three-color staining with MEL-14 and subset-specific markers and compared with expression of endogenous L-selectin in C57BL/6 mice. (A) L-Selectin expression on αβTCR+ T lymphocytes in PLNs and spleen, B220+ cells in spleen, and Gr-1+ myeloid cells in bone marrow and blood are shown. T cells have been excluded from the bone marrow and blood analysis by gating. (top right quadrant) The percentage of L-selectin–positive cells in each subset. (B) Histogram overlays show comparable levels of L-selectin on αβTCR+ T lymphocytes in PLNs of C57BL/6 mice (thin line) and in single transgenic mice (WT, heavy line; LΔP, dotted line). Gray line shows background fluorescence of unstained cells. Ro 31-9790 was included in all buffers used for lymphocyte isolation. Average levels of sL-selectin (ng/ml) in the blood of 12–20-wk-old normal C57BL/6 (B6), WT, and LΔP transgenic mice are also shown. Results are means ± SE of seven to nine mice. **, P < 0.001 compared with WT and B6 mice.
Table I.
T Lymphocyte Distribution in Peripheral Lymphoid Organs Is Similar in WT and LΔP Mice but Different from C57BL/6
| Total number of lymphocytes (×106) |
Percent T lymphocytes |
Percent B lymphocytes |
|||||||
|---|---|---|---|---|---|---|---|---|---|
| WT | LΔP | C57BL/6 | WT | LΔP | C57BL/6 | WT | LΔP | C57BL/6 | |
| PLN | 20 ± 2 | 16 ± 2 | 24 ± 3 | 92 ± 3a | 90 ± 2a | 70 ± 3 | 5 ± 1a | 4 ± 1a | 28 ± 4 |
| Spleen | 138 ± 12 | 146 ± 15 | 143 ± 16 | 31 ± 2 | 33 ± 2 | 31 ± 4 | 60 ± 4 | 59 ± 2 | 59 ± 4 |
| MLN | 13 ± 2 | 16 ± 1 | 19 ± 3 | 68 ± 4 | 71 ± 2 | 70 ± 3 | 37 ± 5 | 35 ± 4 | 29 ± 6 |
| PP | 7 ± 2 | 6 ± 1 | 5 ± 2 | 28 ± 2 | 34 ± 3 | 33 ± 1 | 67 ± 3 | 62 ± 4 | 65 ± 2 |
The total number and subset composition of PLNs, spleen, MLNs, and Peyer's patches (PP) in 6–12-wk-old WT and LΔP mice were determined using T and B cell–specific markers and compared with nontransgenic C57BL/6 mice. Results are mean ± SE (n = 16–22).
P < 0.001 between WT or LΔP and C57BL/6.
It appeared that WT transgenic and C57BL/6 mice showed slightly lower L-selectin expression in comparison with LΔP mice (Fig. 2 A). For example, mean levels of expression were 308 ± 39 and 368 ± 49 (MFI ± SE; n = 8) in age and sex-matched WT and LΔP mice, respectively. However, these differences were not statistically significant. For trafficking studies, it was important to use WT and LΔP mice both expressing levels of transgenic L-selectin comparable to each other and similar to that of endogenous L-selectin on T cells in C57BL/6 mice. Because L-selectin is down-regulated during leukocyte isolation (33), and this is likely due to shedding, we included the shedding inhibitor Ro31-9790 in all buffers used during the isolation of lymphocytes. We selected lines of WT and LΔP transgenic mice that showed similar levels of cell surface expression to each other and to C57BL/6 T cells (Fig. 2 B). Circulating levels of sL-selectin in the WT line were ∼30% lower than in C57BL/6 mice and reduced by 95% in the LΔP line (Fig. 2 B).
LΔP-Selectin Resists Activation-dependent and Constitutive Shedding.
PMA induced rapid shedding and release of L-selectin into the medium by PLN T cells from WT but not LΔP mice (Fig. 3, A and B) . Cell surface levels of L-selectin did not change after incubation of T cells from LΔP mice in medium for 1 h at 37°C (Fig. 3 D), whereas T cells from WT and C57BL/6 mice showed some shedding and release of L-selectin from the cell surface in the absence of stimulation (Fig. 3 C and not depicted).
Figure 3.

LΔP-Selectin resists basal and activation-dependent shedding. Lymphocytes isolated from PLNs of L-selectin transgenic mice were incubated for 1 h at 37°C in the absence and presence of 100 nM PMA, cell surface L-selectin expression was determined by flow cytometry, and L-selectin released into the medium determined by ELISA. (A and B) Histogram overlays show L-selectin expression on T cells in the absence (solid line) and presence (dotted line) of PMA from WT and LΔP mice. (C and D) Histogram overlays show L-selectin expression on lymphocytes from WT and LΔP mice incubated at 37°C (dotted line) or held at 4°C (heavy line). Results are representative of those obtained in at least five independent experiments. Levels of L-selectin (ng/ml) released into the medium are also shown. Results are means ± SE of three independent experiments.
T Lymphocyte Rolling in Hydrodynamic Flow Assays Is Not Regulated by L-Selectin Shedding.
To examine the rolling behavior of T cells expressing a nonshedding form of L-selectin, we used a natural L-selectin ligand mimetic from Hansenula hostii, PPME (42), in hydrodynamic flow assays at a wall shear stress of 1.5 dyn/cm2. L-Selectin expressing lymphocytes established stable rolling on PPME which was completely inhibited by 5mM EGTA and 10 μg/ml MEL-14 but not by an isotype-matched control mAb MAC 193 and L-selectin KO lymphocytes did not roll on PPME (unpublished data). We compared the number and velocity of rolling cells isolated from C57BL/6, WT, and LΔP mice. The number of rolling cells from WT mice at 45 ± 16 was not significantly different from C57BL/6 and they rolled with similar median velocities (Fig. 4) . The number and rolling velocity of lymphocytes from LΔP mice did not differ significantly from WT. To further analyze these results, we calculated the relative frequencies of individual cells rolling at defined speeds. The majority of T cells from C57BL/6 and WT mice (∼60%) rolled with velocities from 5 to 20 μm/s and <15% showed high rolling velocities of 30–75 μm/s; LΔP lymphocytes showed a similar velocity distribution (Fig. 4). In addition, we analyzed the jerkiness (i.e., the acceleration and deceleration in rolling velocities of individual cells at 0.07-s intervals; reference 14) and found no differences between WT and LΔP mice (Fig. 5) . Papers that have shown L-selectin shedding to regulate neutrophil rolling behavior used synthetic hydroxamic acid–based MMP inhibitors to block shedding. To determine whether these reagents exhibit effects on rolling that are unrelated to shedding, we studied rolling in the presence of Ro31-9790, which we have shown to inhibit constitutive, chemokine, and PMA-induced L-selectin shedding (21, 43). The jerkiness, number, median velocity, and velocity distribution of rolling cells from both LΔP and WT mice were not affected by pretreatment and inclusion of the MMP inhibitor in the flow assay (Fig. 5 and not depicted). Together, these data suggest that L-selectin–dependent rolling of T cells in hydrodynamic flow is not regulated by ectodomain shedding as shown using either a nonshedding mutant of L-selectin or exogenous shedding inhibitors.
Figure 4.

LΔP-Selectin does not affect the rolling behavior of T lymphocytes under flow conditions. 106/ml lymphocytes were perfused at 37°C over slides coated with PPME at constant shear stress of 1.5 dynes/cm2, and the number of rolling cells was determined after 2 min. The velocity of at least 300 rolling cells from WT (A), LΔP (B), and C57BL/6 (C) mice was measured and results presented as relative frequency. The median rolling velocity is also shown for each genotype. Results are means ± SE of data pooled from three to six independent experiments.
Figure 5.

Rolling behavior of T lymphocytes is not affected by L-selectin shedding inhibitors. Rolling behavior of T lymphocytes from WT and LΔP mice in the absence and presence of 30 μM Ro31-9790 was analyzed as described in Fig. 4. Microkinetic analysis was performed by measuring rolling velocity of individual cells at 0.07-s intervals over a period of 3 s. Velocity profiles of individual cells are shown and are representative of cells rolling in the absence and presence of Ro31-9790 from WT and LΔP mice.
Inability to Shed L-Selectin Does Not Affect Homing of Naive T Cells to PLNs.
To determine whether L-selectin shedding regulates lymphocyte recruitment in vivo, the migration of T cells expressing WT and nonshedding L-selectin were compared. PLN lymphocytes from WT and LΔP mice (>90% T cells) were labeled with either CFSE or CMTMR, a mixture of labeled lymphocytes injected i.v. into C57BL/6 mice, and the ratio of injected cells in PLNs, spleen, and blood after 60 min were compared with that in the injected (start) population (Fig. 6) . Localization ratios in the organs of recipient mice that deviate from unity demonstrate nonidentical migration characteristic of the two lymphocytes populations under study. The ratio of WT/LΔP cells in PLNs of C57BL/6 mice at 1.15 ± 0.1 (Table II) suggests that L-selectin shedding does not regulate trafficking because WT and LΔP lymphocytes migrated equally well to PLNs. However, fluorescently labeled immigrant cells represented a minor fraction of total resident cells in C57BL/6 recipients (1–2%; unpublished data). RAG-1 KO mice, in which immigrant lymphocytes composed up to 20% of the resident population, were used as recipients in some experiments to increase the sensitivity. The localization ratio of WT and LΔP lymphocytes in RAG-1 KO was similar to that in C57BL/6 at 1.02 ± 0.1 (Table II), confirming that the inability to shed L-selectin does not affect lymphocyte homing to PLNs. Along with the absence of an effect on migration to PLNs, localization ratios of WT and LΔP T cells in the spleen and blood were near unity (Table II). Lymphocyte migration to PLN of RAG-1 KO mice was L-selectin dependent because L-selectin KO T cells did not enter PLNs and increased numbers were found in the spleen and blood of RAG-1 KO mice (Table II), as reported previously in C57BL/6 recipients (7). Transgenic Ly22− L-selectin supported normal trafficking because T cells from WT and C57BL/6 mice migrated equally well to PLN with a localization ratio of 1.06 ± 0.1 (Table II). Neither fluorescent label affected lymphocyte migration because the localization ratios of a population of C57BL/6 lymphocytes that was divided into two, labeled with either CFSE or CMTMR, mixed 1:1, and injected into C57BL/6 or RAG-1 KO mice were near unity in PLNs, spleen, and blood (unpublished data).
Figure 6.
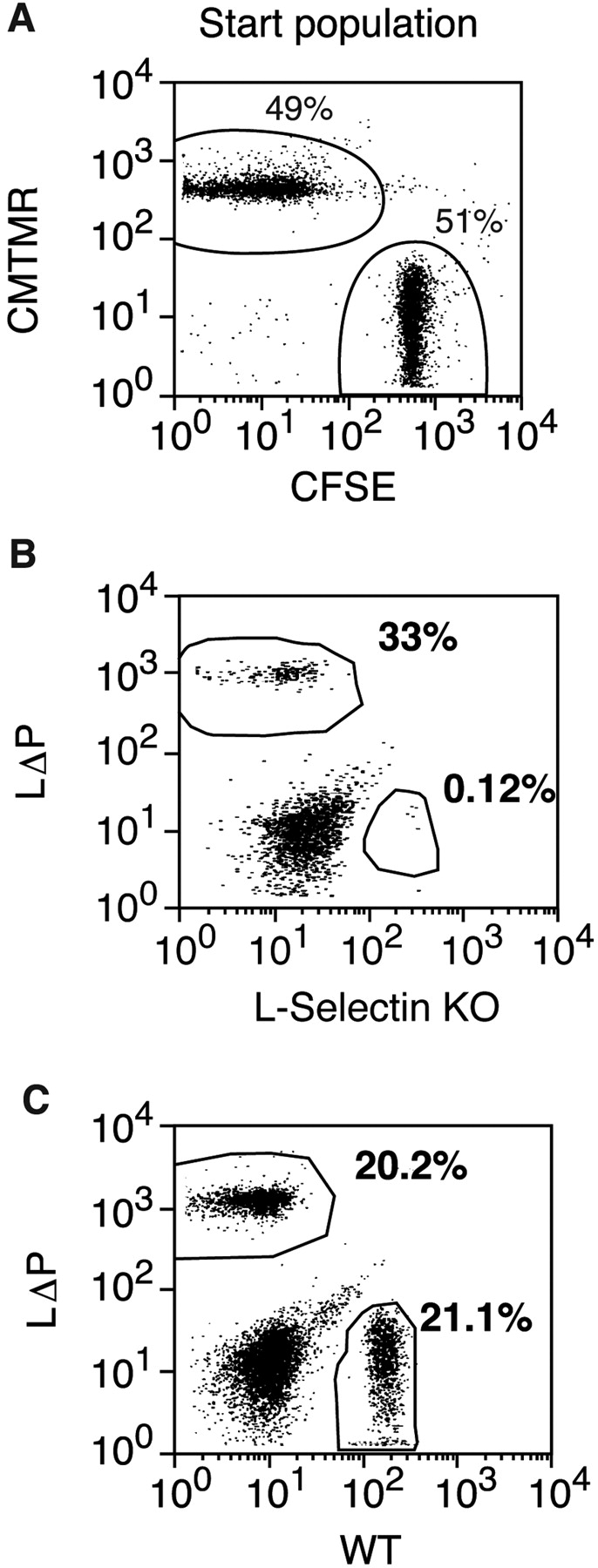
Inability to shed L-selectin does not affect homing to PLNs. (A) The two populations of T lymphocytes to be compared were labeled with either CFSE or CMTMR and mixed in equal numbers (start population) for injection into recipient mice. (B and C) LΔP-Selectin–expressing lymphocytes were mixed with either L-selectin KO (B) or WT (C) lymphocytes, injected into RAG-1 KO mice, and the presence of labeled cells in PLNs was determined after 60 min by flow cytometry. The percentages of CFSE- and CMTMR-labeled cells are shown on the dot plots. 1 representative experiment is shown from 3 using L-selectin KO and 16 using WT lymphocytes.
Table II.
Inability to Shed L-Selectin Does Not Affect Homing of Naive T Cells to Peripheral Lymph Nodes
| Injected cells | No. of mice |
Recipient | PLN | Spleen | Blood |
|---|---|---|---|---|---|
| WT/LΔP | 7 | C57BL/6 | 1.1 ± 0.1 | 0.9 ± 0.2 | 0.75 ± 0.1 |
| WT/LΔP | 9 | RAG-1 KO | 1.0 ± 0.1 | 1.2 ± 0.2 | 0.95 ± 0.1 |
| L-Selectin KO/LΔP |
3 | RAG-1 KO | 0.05 ± 0.02a | 1.3 ± 0.3 | 1.6 ± 0.2b |
| C57BL6/WT | 6 | RAG-1 KO | 1.0 ± 0.1 | 0.85 ± 0.1 | 1.1 ± 0.1 |
T lymphocytes from different transgenic mice were labeled with CFSE or CMTMR fluorescent dyes and equal numbers injected into C57BL/6 or RAG-1 KO recipient mice. After 60 min, single cell suspensions were obtained from different organs, and the numbers of fluorescent cells were determined by flow cytometry. The ratio of CFSE/CMTMR-labeled cells in each organ was determined and normalized to the ratio in the injected population. Results show mean normalized localization ratios ± SE for the two T cell populations studied.
P < 0.001.
P < 0.05.
We have reported that systemic treatment of mice with the MMP inhibitor Ro31-9790 did not affect the recruitment of lymphocytes from the bloodstream but arrested lymphocytes in the endothelial lining of HEVs, reduced migration into the paracortex, and blocked L-selectin shedding during PLN entry (44). These results suggested that transendothelial migration across HEVs is MMP-dependent and that L-selectin shedding might regulate this event. Therefore, we analyzed the distribution of fluorescently labeled LΔP T cells in PLNs of recipient mice. Cells attached to the lumen or in the process of transmigration across the endothelial lining of HEVs were identified by counterstaining HEV in PLN sections with mAb MECA 79 and recorded as Inside HEVs, the remaining labeled cells being recorded as outside HEVs (Fig. 7 ; reference 44). Labeling with different dyes did not differentially affect T cell binding and transmigration because the ratio of labeled cells did not deviate from unity either inside or outside HEVs after injection of a 1:1 mixture of CFSE- and CMTMR-labeled C57BL/6 T lymphocytes (Fig. 7). The number of LΔP T cells Inside HEVs was consistently greater than WT cells with a normalized LΔP/WT ratio of 1.66 ± 0.24 (P < 0.03). The number of LΔP T cells Outside HEVs was also greater (LΔP/WT ratio of 1.78 ± 0.38; P < 0.07). The accumulation of LΔP over WT lymphocytes suggests that the inability to shed L-selectin reduces the rate of T cell movement across HEVs and into PLNs.
L-Selectin Shedding after Antigen Activation Regulates T Lymphocyte Entry into PLNs.
After activation by antigen, T cells show profoundly altered migration pathways characterized by loss of capacity to enter LNs and acquired capacity to enter nonlymphoid organs such as the lungs, skin, or gut and sites of inflammation. L-Selectin is down-regulated from the T cell surface within minutes after antigen receptor engagement and most antigen-activated T cells found in nonlymphoid organs are L-selectinlow (24, 26) but the roles of L-selectin shedding in regulating the expression and migration pathways of antigen-activated T cells have not been determined.
F5/RAG KO mice in which all lymphocytes express a class I restricted TCR receptor for influenza virus A/NT/60/68 nucleoprotein on CD8+ T cells (29) were used in the first instance to study antigen receptor induced shedding of endogenous L-selectin. The cognate peptide, NP-68, induced rapid, MMP-dependent shedding of endogenous L-selectin from CD8+ T cells in F5/RAG KO within 60 min. PMA induced shedding to a similar extent as NP-68 whereas a peptide from HIV GAG protein that does not bind the F5 TCR did not induce shedding (Fig. 8) . WT and LΔP mice were crossed with F5/L-selectin KO mice and offspring selected for expression of F5 TCR and transgenic L-selectin. In F5/WT and F5/LΔP mice, PLNs comprised ∼85% CD8+/F5+/L-selectin+ T cells, ∼10% CD4+/L-selectin+ T cells, and <5% L-selectin− B cells (unpublished data). NP-68 induced MMP-dependent shedding of transgenic L-selectin from CD8+ T cells in F5/WT mice but not from 10% CD8−/L-selectin+ cells found in these mice (unpublished data). PMA induced shedding from all L-selectin expressing cells and, therefore, was slightly greater than that induced by NP-68 (Fig. 8 A). In F5/LΔP mice, the number of CD8+ T cells expressing L-selectin and the mean fluorescence intensity were not altered after incubation with NP-68 peptide (Fig. 8 and not depicted).
Figure 8.



MMP-dependent shedding of L-selectin after TCR engagement regulates entry into PLNs. (A) PLN lymphocytes from F5/RAG-1 KO, F5/WT, and F5/LΔP mice were incubated with 0.1 μm cognate NP-68 or control peptide or 300 nM PMA at 37°C for 60 min in the absence and presence of 30 μM Ro31-9790. L-Selectin expression on F5-positive CD8 T cells was determined by flow cytometric analysis of dual-stained cells. Results are pooled from at least three independent experiments and are the mean percentage of F5 T cells expressing L-selectin ± SE. (B) CFSE-labeled PLN lymphocytes from F5/WT mice were incubated with 0.1 μM NP-68 peptide at 37°C for 60 min, mixed with equal numbers of either naive WT or NP68-activated LΔP lymphocytes labeled with CMTMR, and injected into RAG-1 KO mice. The percentages of immigrant CFSE- and CMTMR-labeled cells in PLNs after 60 min are shown on the dot plots. Representative data from a single experiment are shown. (C) PLN lymphocytes from F5/WT and F5/LΔP mice were incubated with or without NP-68 peptide and peptide removed by washing. In some experiments, Ro31-9790 was included during antigen activation to inhibit L-selectin shedding and removed before trafficking experiments. The lymphocyte populations to be compared were labeled with either CFSE or CMTMR and equal numbers injected into RAG-1 KO mice. After 60 min, the numbers of labeled cells in PLNs, spleen, and blood were measured by flow cytometry. The normalized ratio of CFSE/CMTMR-labeled cells in each organ was calculated as described in Materials and Methods. Results show mean localization ratios ± SE as follows: activated WT, naive F5/WT (black bars, n = 7); activated F5/WT, activated F5/LΔP (light gray bars, n = 7); activated F5/WT, activated F5/WT in the presence Ro31-9790 (white bars, n = 4); and activated LΔP, naive F5/LΔP (dark gray bars, n = 5). *, P < 0.05. **, P < 0.001.
After activation by antigen and shedding of transgenic L-selectin, the number of immigrant CD8 T cells from F5/WT mice in PLNs was reduced from 19% to 6%, giving a localization ratio of 0.31 ± 0.04 (Fig. 8). Antigen-activated CD8 T cells from F5/RAG KO mice showed a similar reduction in migration to PLNs with a localization ratio of 0.23 ± 0.07 (n = 6). The reduced migration to PLNs was not simply due to removal of cells from the circulation because equal numbers of antigen-activated and naive F5/WT cells were found in the bloodstream and spleen (Fig. 8). Antigen-activated CD8 T cells from F5/LΔP mice entered PLNs more efficiently than antigen-activated F5/WT (10 vs. 5% immigrant cells; localization ratio, 0.59 ± 0.02, Fig. 8) as did cells from F5/WT mice activated in the presence of Ro31-9790, showing that nonshed cell surface L-selectin on antigen-activated T cells functions as a PLN homing receptor. However, activated F5/LΔP cells migrated less efficiently than either naive WT cells (Fig. 8, 10 vs. 19% immigrant cells, respectively) or naive F5/LΔP (Fig. 8, localization ratio of 0.63 ± 0.06) even though they express similar levels of L-selectin suggesting that migration of antigen activated T cell into PLN is regulated by factors in addition to L-selectin expression.
Discussion
To study the physiological role of L-selectin shedding in lymphocyte biology, we have mutagenized the cleavage site of mouse L-selectin and directed the expression of mutant or WT L-selectin to T lymphocytes by transgenesis. L-Selectin transgenic mice were bred with L-selectin KO mice to generate lines in which either WT or nonshedding L-selectin was only expressed on T lymphocytes, and lines expressing physiological levels of L-selectin at the cell surface were selected for lymphocyte migration studies. We deleted the Ly22 epitope recognized by mAb T28 to distinguish transgenic from endogenous L-selectin during backcrossing to L-selectin KO mice. The anti-Ly22 antibody T28 inhibits L-selectin–dependent binding to PLNs in the frozen section assay (31). However, we could detect no differences in the function of transgenic Ly22− (WT) and endogenous Ly22+ (C57BL/6) L-selectin either in rolling assays or in short-term trafficking to PLNs. We have compared the migration pathways of T cells expressing either WT or a mutant form of L-selectin that resists constitutive and activation-induced shedding (LΔP) and show that L-selectin shedding does not regulate the constitutive trafficking of naive T cells to PLNs but controls the entry of antigen-activated T cells.
A major limitation to investigating the role of L-selectin shedding has been the reliance on a broad spectrum metalloproteinase inhibitors. These were originally developed to block matrix metalloproteinases (MMPs) but subsequently found to block related enzymes such as ADAM17/TACE (45). Their efficacy against the full spectrum of MMPs, ADAMs, and other related enzymes, such as the tolloid/astacins (46), is not known, therefore, proteolysis of substrates other than L-selectin could contribute to their effects on leukocyte recruitment (44). Interestingly, inhibition of L-selectin shedding does not affect neutrophil rolling or recruitment on endothelium when E-selectin is expressed (43), possibly because E-selectin–dependent rolling is significantly slower than that of L-selectin, even when shedding is blocked (13). Because entry of T cells into pLNs is completely dependent on L-selectin and the other selectins do not substitute, we chose to analyze the effects of expressing a nonshedding form of L-selectin on T cell rolling behavior and migration into PLNs.
A clear and surprising result of this work was that L-selectin shedding does not regulate T lymphocyte rolling behavior in vitro. LΔP T cells behaved identically to WT and C57BL/6 T cells according to all rolling parameters measured, including the jerkiness of rolling. Moreover, inclusion of the MMP inhibitor Ro31-9790 did not affect the number, velocity, or jerkiness of rolling T cells. It could be argued that the high valency and clustering of PPME would mask any effects of L-selectin shedding on rolling. However, it has recently been reported that T lymphocyte rolling on GlyCAM-1 was not affected by inhibitors of L-selectin shedding, such as Ro31-9790 and KD-IX-73-4 (47). Our results contrast sharply with papers about neutrophils using KD-IX-73-4 under flow (12–14). Inhibition of L-selectin shedding was found to increase neutrophil binding to L-selectin ligands, to decrease the rolling velocity, to enhance β2 integrin activation in vitro, and to increase ICAM-1–dependent neutrophil recruitment in vivo. From these studies, it was concluded that threshold levels of L-selectin on neutrophils regulate rolling behavior such that lowering of cell surface L-selectin increases the velocity and, therefore, decreases exposure to endothelial-expressed ligands for β2 integrin activation and the subsequent arrest of rolling cells (14). The lack of regulation of T lymphocyte rolling behavior by L-selectin shedding suggests that, if shedding occurs, it is not sufficient to reduce cell surface expression below threshold levels that may regulate rolling. However, L-selectin–dependent rolling of neutrophils and T cells differ significantly in that there is an energy requirement for T cells, but not neutrophils, and lymphocyte rolling is slower and more resistant to shear stress (48). Therefore, it is possible that changes in the cell surface levels of L-selectin during rolling are not sufficient to affect the already slow, more stable rolling of T cells in comparison with neutrophils.
Our results also show clearly that the inability to cleave cell surface L-selectin does not significantly affect the recruitment of naive T lymphocytes from the bloodstream into PLNs. In short-term trafficking studies, T cells expressing LΔP-selectin entered PLNs as efficiently as T cells expressing either transgenic (WT) or endogenous (C57BL/6) wild-type L-selectin. LΔP T cells migrated across the walls of HEVs and into the LN paracortex, however, the number of LΔP cells was consistently higher than WT cells both in the vessel wall and in the paracortex. Because the total numbers of immigrated WT and LΔP T cells were comparable in trafficking experiments, the increased ratio of LΔP/WT cells suggests that LΔP lymphocytes move more slowly. Although L-selectin is shed during entry into PLNs, immigrant cells remain L-selectin–positive (44). Therefore, it is possible that in the absence of shedding, higher levels of L-selectin on immigrant LΔP T cells may alter the quality of signals induced by cross-linking L-selectin that, for example, control β1 integrin activation (49) and, thereby, rates of migration. Ligands for L-selectin are expressed on the basolateral as well as the luminal surface of high endothelial cells (50), which raises the possibility of a role for L-selectin engagement in events after the rolling stages of recruitment, as already suggested for neutrophils (51). It will be interesting to analyze traffic routes distal to HEVs to determine whether shedding controls subsequent movement through the LNs.
Despite slower movement, the inability to shed L-selectin does not impact on overall steady-state distribution of T cells in PLNs because the cellularities and subset compositions were similar in LΔP and WT mice. Although the ratios of B/T cells in PLNs of WT and LΔP transgenic mice were similar, they were reduced in comparison with C57BL/6 mice, confirming the critical role for L-selectin in B cell trafficking into PLNs (7). However, the B/T ratio and cellularity of gut-associated lymphoid organs of L-selectin transgenics did not differ from C57BL/6. Healthy LΔP mice have been maintained in specific pathogen-free conditions for >12 mo without obvious signs of ill health.
Normal mouse serum contains functionally active sL-selectin in levels sufficient to inhibit lymphocyte migration into PLNs (52). Circulating sL-selectin could arise from constitutive shedding or from activation induced shedding during leukocyte recruitment (12, 14, 44). Because monocytes, neutrophils, and T and B lymphocytes shed L-selectin in response to different stimuli, circulating sL-selectin may arise from all or a combination of these cell types. We demonstrate here that a significant fraction of sL-selectin can arise from T lymphocytes because the concentration of sL-selectin in serum of WT mice, which only express L-selectin on T cells, was ∼70% of that in C57BL/6 mouse. These data are in agreement with previous analyses in which it was shown that ∼70% of serum sL-selectin is derived from lymphocytes (52). We further show that the sL-selectin concentration in LΔP mice was <5% of that WT mice, which suggests that a significant fraction of sL-selectin in normal mice is generated by ectodomain shedding.
Many papers have reported that L-selectin expression is low or absent on antigen-activated T cells, particularly on those harvested from sites of inflammation (24, 26). L-Selectin is rapidly down-regulated from the surface of T cells within 60 min after stimulation by anti-CD3 (53) or superantigen (54), is reexpressed within 12 h, and expression is finally lost 2–3 d after anti-CD3. It is assumed that the loss of L-selectin is due to shedding and that it is sufficient to redirect T cells away from LNs but this has not been demonstrated directly. Therefore, we used LΔP transgenic mice to determine the contribution of shedding to the complex pattern of L-selectin expression and the migration pathways of antigen-activated T cells. A transgenic TCR and its cognate peptide antigen were used to activate T cells from WT and LΔP mice. This activation resulted in rapid, MMP-dependent shedding from the surface of WT but not LΔP cells. The kinetics and magnitude of shedding were similar to that of endogenous L-selectin in F5 mice and to that induced by PMA. In short-term trafficking assays, the entry of antigen-activated WT T cells into PLNs was reduced by 70% in comparison with naive T cells, consistent with loss of cell surface L-selectin and confirming a role for L-selectin shedding in regulating the migration of antigen-activated T cells. Antigen-activated T cells, on which cell surface L-selectin expression has been maintained either by mutation (LΔP) or by shedding inhibitor Ro31-9790, migrated more efficiently than their WT counterparts, which shed L-selectin normally after TCR engagement. This difference is most likely due to retention of L-selectin enabling antigen-activated cells to enter PLNs in increased numbers. Interestingly, antigen-activated T lymphocytes (LΔP or WT + Ro31-9790) expressing similar levels of L-selectin as naive T cells did not enter LNs as efficiently as naive cells (localization ratios of 0.6 ± 0.06 and 0.6 ± 0.02, respectively) suggesting that factors in addition to the level of L-selectin expression affect entry into PLNs. A number of other cell adhesion molecules known to regulate T cell migration pathways (CD44, LFA-1, VLA-4, and α4β7 integrin) as well as chemokine receptors are altered in function on activated T cells and may regulate PLN homing.
The LΔP-selectin transgenic mice that we have generated show clearly that the ability to shed L-selectin does not regulate T lymphocyte recirculation or homing to LNs. However, these mice provide the first proof-of-principle that shedding of L-selectin from the surface of antigen-activated T cells prevents reentry into PLNs from the bloodstream. Further studies will be required to determine whether shedding regulates the migration pathways of antigen-activated T cells that have undergone proliferation and differentiation in vivo. For example, it will be interesting to determine whether shedding regulates the differential migration of central and effector memory cells to PLNs. LΔP-Selectin transgenic mice will be useful to study the consequences, if any, of aberrant migration of antigen-activated T cells for host immunity.
Acknowledgments
We thank A. Docherty and P. Stephens for advice on mutagenesis and cloning, A. O'Garra for discussions and enthusiastic support and A. Hamann for helpful comments on the manuscript. We thank and acknowledge the expertise of H. Boyes for maintaining the transgenic lines, C. Faveeuw for advice on tissue preparation for multicolor fluorescence microscopy, J. Adler and S. Pagakis for assistance with confocal microscopy, and L. MacNeill and J. Brock for the figures.
This work was supported by the Medical Research Council (MRC) and in part by European Union grant CT-1999-01036. E. Galkina was supported by a Wellcome Trust Travelling Fellowship and K. Tanousis was supported by an MRC Collaborative Studentship with Celltech (G78/5223).
E. Galkina and K. Tanousis contributed equally to this work.
Abbreviations used in this paper: CFSE, carboxyfluorescein diacetate-succinimidyl ester; CMTMR, 5-(and 6)-([{4-chloromethyl}benzoyl]amino) tetramethylrhodamine; HEV, high endothelial venule; MLN, mesenteric LN; MMP, matrix metalloproteinase; MPR, membrane proximal region; PLN, peripheral LN; sL-selectin, soluble L-selectin; PPME, polyphosphomannan ester.
References
- 1.Butcher, E.C., and L.J. Picker. 1996. Lymphocyte homing and homeostasis. Science. 272:60–66. [DOI] [PubMed] [Google Scholar]
- 2.Springer, T.A. 1994. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell. 76:301–314. [DOI] [PubMed] [Google Scholar]
- 3.Butcher, E.C. 1991. Leukocyte-endothelial cell recognition: three (or more) steps to specificity and diversity. Cell. 67:1033–1036. [DOI] [PubMed] [Google Scholar]
- 4.Forster, R., A. Schubel, D. Breitfeld, E. Kremmer, I. Renner-Muller, E. Wolf, and M. Lipp. 1999. CCR7 coordinates the primary immune response by establishing functional microenvironments in secondary lymphoid organs. Cell. 99:23–33. [DOI] [PubMed] [Google Scholar]
- 5.Gunn, M.D., S. Kyuwa, C. Tam, T. Kakiuchi, A. Matsuzawa, L.T. Williams, and H. Nakano. 1999. Mice lacking expression of secondary lymphoid organ chemokine have defects in lymphocyte homing and dendritic cell localization. J. Exp. Med. 189:451–460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Warnock, R.A., S. Askari, E.C. Butcher, and U.H. von Andrian. 1998. Molecular mechanisms of lymphocyte homing to peripheral lymph nodes. J. Exp. Med. 187:205–216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Arbones, M.L., D.C. Ord, K. Ley, H. Ratech, C. Maynard-Curry, G. Otten, D.J. Capon, and T.F. Tedder. 1994. Lymphocyte homing and leukocyte rolling and migration are impaired in L-selectin-deficient mice. Immunity. 1:247–260. [DOI] [PubMed] [Google Scholar]
- 8.Catalina, M.D., M.C. Carroll, H. Arizpe, A. Takashima, P. Estess, and M.H. Siegelman. 1996. The route of antigen entry determines the requirement for L-selectin during immune responses. J. Exp. Med. 184:2341–2351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Steeber, D.A., N.E. Green, S. Sato, and T.F. Tedder. 1996. Humoral immune responses in L-selectin-deficient mice. J. Immunol. 157:4899–4907. [PubMed] [Google Scholar]
- 10.Xu, J., I.S. Grewal, G.P. Geba, and R.A. Flavell. 1996. Impaired primary T cell responses in L-selectin–deficient mice. J. Exp. Med. 183:589–598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Alon, R., S. Chen, K.D. Puri, E.B. Finger, and T.A. Springer. 1997. The kinetics of L-selectin tethers and the mechanics of selectin-mediated rolling. J. Cell Biol. 138:1169–1180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Walcheck, B., J. Kahn, J.M. Fisher, B.B. Wang, R.S. Fisk, D.G. Payan, C. Feehan, R. Betageri, K. Darlak, A.F. Spatola, and T.K. Kishimoto. 1996. Neutrophil rolling altered by inhibition of L-selectin shedding in vitro. Nature. 380:720–723. [DOI] [PubMed] [Google Scholar]
- 13.Hafezi-Moghadam, A., and K. Ley. 1999. Relevance of L-selectin shedding for leukocyte rolling in vivo. J. Exp. Med. 189:939–948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hafezi-Moghadam, A., K.L. Thomas, A.J. Prorock, Y. Huo, and K. Ley. 2001. L-Selectin shedding regulates leukocyte recruitment. J. Exp. Med. 193:863–872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Strausbaugh, H.J., P.G. Green, E. Lo, K. Tangemann, D.B. Riechling, S.D. Rosen, and J.D. Levine. 1999. Painful stimulation suppresses joint inflammation by inducing shedding of L-selectin from neutrophils. Nat. Med. 5:1057–1061. [DOI] [PubMed] [Google Scholar]
- 16.Jung, T.M., W.M. Gallatin, I.L. Weissman, and M.O. Dailey. 1988. Down-regulation of homing receptors after T cell activation. J. Immunol. 141:4110–4117. [PubMed] [Google Scholar]
- 17.Jung, T.M., and M.O. Dailey. 1990. Rapid modulation of homing receptors (gp90(MEL-14)) induced by activators of protein kinase C. Receptor shedding due to accelerated proteolytic cleavage at the cell surface. J. Immunol. 144:3130–3136. [PubMed] [Google Scholar]
- 18.Palecanda, A., B. Walcheck, D.K. Bishop, and M.A. Jutila. 1992. Rapid activation-independent shedding of leukocyte L-selectin induced by cross-linking of the surface antigen. Eur. J. Immunol. 22:1279–1286. [DOI] [PubMed] [Google Scholar]
- 19.Bennett, T.A., E.B. Lynam, L.A. Sklar, and S. Rogelj. 1996. Hydroxamate-based metalloprotease inhibitor blocks shedding of L-selectin adhesion molecule from leukocytes: functional consequences for neutrophil aggregation. J. Immunol. 156:3093–3097. [PubMed] [Google Scholar]
- 20.Feehan, C., K. Darlak, J. Kahn, B. Walcheck, A.F. Spatola, and T.K. Kishimoto. 1996. Shedding of the lymphocyte L-selectin adhesion molecule is inhibited by a hydroxamic acid-based protease inhibitor. Identification with an L-selectin-alkaline phosphatase reporter. J. Biol. Chem. 271:7019–7024. [DOI] [PubMed] [Google Scholar]
- 21.Preece, G., G. Murphy, and A. Ager. 1996. Metalloproteinase-mediated regulation of L-selectin levels on leucocytes. J. Biol. Chem. 271:11634–11640. [DOI] [PubMed] [Google Scholar]
- 22.Zhao, L.C., J.B. Edgar, and M.O. Dailey. 2001. Characterization of the rapid proteolytic shedding of murine L-selectin. Dev. Immunol. 8:267–277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tang, M.L., D.A. Steeber, X.Q. Zhang, and T.F. Tedder. 1998. Intrinsic differences in L-selectin expression levels affect T and B lymphocyte subset-specific recirculation pathways. J. Immunol. 160:5113–5121. [PubMed] [Google Scholar]
- 24.Harris, N.L., V. Watt, F. Ronchese, and G. Le Gros. 2002. Differential T cell function and fate in lymph node and nonlymphoid tissues. J. Exp. Med. 195:317–326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Weninger, W., M.A. Crowley, N. Manjunath, and U.H. von Andrian. 2001. Migratory properties of naive, effector, and memory CD8+ T cells. J. Exp. Med. 194:953–966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Roman, E., E. Miller, A. Harmsen, J. Wiley, U.H. Von Andrian, G. Huston, and S.L. Swain. 2002. CD4 effector T cell subsets in the response to influenza: heterogeneity, migration, and function. J. Exp. Med. 196:957–968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Spanopoulou, E., C.A. Roman, L.M. Corcoran, M.S. Schlissel, D.P. Silver, D. Nemazee, M.C. Nussenzweig, S.A. Shinton, R.R. Hardy, and D. Baltimore. 1994. Functional immunoglobulin transgenes guide ordered B-cell differentiation in Rag-1-deficient mice. Genes Dev. 8:1030–1042. [DOI] [PubMed] [Google Scholar]
- 28.Mamalaki, C., T. Norton, Y. Tanaka, A.R. Townsend, P. Chandler, E. Simpson, and D. Kioussis. 1992. Thymic depletion and peripheral activation of class I major histocompatibility complex-restricted T cells by soluble peptide in T-cell receptor transgenic mice. Proc. Natl. Acad. Sci. USA. 89:11342–11346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Corbella, P., D. Moskophidis, E. Spanopoulou, C. Mamalaki, M. Tolaini, A. Itano, D. Lans, D. Baltimore, E. Robey, and D. Kioussis. 1994. Functional commitment to helper T cell lineage precedes positive selection and is independent of T cell receptor MHC specificity. Immunity. 1:269–276. [DOI] [PubMed] [Google Scholar]
- 30.Stephens, P.E., and M.I. Cockett. 1989. The construction of a highly efficient and versatile set of mammalian expression vectors. Nucleic Acids Res. 17:7110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Siegelman, M.H., I.C. Cheng, I.L. Weissman, and E.K. Wakeland. 1990. The mouse lymph node homing receptor is identical with the lymphocyte cell surface marker Ly-22: role of the EGF domain in endothelial binding. Cell. 61:611–622. [DOI] [PubMed] [Google Scholar]
- 32.Zhumabekov, T., P. Corbella, M. Tolaini, and D. Kioussis. 1995. Improved version of a human CD2 minigene based vector for T cell-specific expression in transgenic mice. J. Immunol. Methods. 185:133–140. [DOI] [PubMed] [Google Scholar]
- 33.Stibenz, D., and C. Buhrer. 1994. Down-regulation of L-selectin surface expression by various leucocyte isolation procedures. Scand. J. Immunol. 39:59–63. [PubMed] [Google Scholar]
- 34.Tomonari, K., and E. Lovering. 1988. T-cell receptor-specific monoclonal antibodies against a V beta 11-positive mouse T-cell clone. Immunogenetics. 28:445–451. [DOI] [PubMed] [Google Scholar]
- 35.Steeber, D.A., P. Engel, A.S. Miller, M.P. Sheetz, and T.F. Tedder. 1997. Ligation of L-selectin through conserved regions within the lectin domain activates signal transduction pathways and integrin function in human, mouse, and rat leukocytes. J. Immunol. 159:952–963. [PubMed] [Google Scholar]
- 36.Rao, R.M., J.L. Clarke, S. Ortlepp, M.K. Robinson, R.C. Landis, and D.O. Haskard. 2002. The S128R polymorphism of E-selectin mediates neuraminidase-resistant tethering of myeloid cells under shear flow. Eur. J. Immunol. 32:251–260. [DOI] [PubMed] [Google Scholar]
- 37.Berlin-Rufenach, C., F. Otto, M. Mathies, J. Westermann, M.J. Owen, A. Hamann, and N. Hogg. 1999. Lymphocyte migration in lymphocyte function-associated antigen (LFA)-1–deficient mice. J. Exp. Med. 189:1467–1478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Iezzi, G., D. Scheidegger, and A. Lanzavecchia. 2001. Migration and function of antigen-primed nonpolarized T lymphocytes in vivo. J. Exp. Med. 193:987–993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kahn, J., R.H. Ingraham, F. Shirley, G.I. Migaki, and T.K. Kishimoto. 1994. Membrane proximal cleavage of L-selectin: identification of the cleavage site and a 6-kD transmembrane peptide fragment of L-selectin. J. Cell Biol. 125:461–470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Migaki, G.I., J. Kahn, and T.K. Kishimoto. 1995. Mutational analysis of the membrane-proximal cleavage site of L-selectin: relaxed sequence specificity surrounding the cleavage site. J. Exp. Med. 182:549–557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.De Boer, J., A. Williams, G. Skavdis, N. Harker, M. Coles, M. Tolaini, T. Norton, K. Williams, K. Roderick, A. Potocnik, and D. Kioussis. 2003. Transgenic mice with hematopoietic and lymphoid specific expression of Cre. Eur. J. Immunol. 33:314–325. [DOI] [PubMed] [Google Scholar]
- 42.Yednock, T.A., E.C. Butcher, L.M. Stoolman, and S.D. Rosen. 1987. Receptors involved in lymphocyte homing: relationship between a carbohydrate-binding receptor and the MEL-14 antigen. J. Cell Biol. 104:725–731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Allport, J.R., H.T. Ding, A. Ager, D.A. Steeber, T.F. Tedder, and F.W. Luscinskas. 1997. L-selectin shedding does not regulate human neutrophil attachment, rolling, or transmigration across human vascular endothelium in vitro. J. Immunol. 158:4365–4372. [PubMed] [Google Scholar]
- 44.Faveeuw, C., G. Preece, and A. Ager. 2001. Transendothelial migration of lymphocytes across high endothelial venules into lymph nodes is affected by metalloproteinases. Blood. 98:688–695. [DOI] [PubMed] [Google Scholar]
- 45.Amour, A., P.M. Slocombe, A. Webster, M. Butler, C.G. Knight, B.J. Smith, P.E. Stephens, C. Shelley, M. Hutton, V. Knauper, et al. 1998. TNF-alpha converting enzyme (TACE) is inhibited by TIMP-3. FEBS Lett. 435:39–44. [DOI] [PubMed] [Google Scholar]
- 46.Stocker, W., and W. Bode. 1995. Structural features of a superfamily of zinc-endopeptidases: the metzincins. Curr. Opin. Str. Biol. 5:383–390. [DOI] [PubMed] [Google Scholar]
- 47.Grabovsky, V., O. Dwir, and R. Alon. 2002. Endothelial chemokines destabilize L-selectin-mediated lymphocyte rolling without inducing selectin shedding. J. Biol. Chem. 277:20640–20650. [DOI] [PubMed] [Google Scholar]
- 48.Dwir, O., F. Shimron, C. Chen, M.S. Singer, S.D. Rosen, and R. Alon. 1998. GlyCAM-1 supports leukocyte rolling in flow: evidence for a greater dynamic stability of L-selectin rolling of lymphocytes than of neutrophils. Cell Adhes. Commun. 6:349–370. [DOI] [PubMed] [Google Scholar]
- 49.Giblin, P.A., S.T. Hwang, T.R. Katsumoto, and S.D. Rosen. 1997. Ligation of L-selectin on T lymphocytes activates β1 integrins and promotes adhesion to fibronectin. J. Immunol. 159:3948–3507. [PubMed] [Google Scholar]
- 50.Hemmerich, S., A. Bistrup, M.S. Singer, A. van Zante, J.K. Lee, D. Tsay, M. Peters, J.L. Carminati, T.J. Brennan, K. Carver-Moore, et al. 2001. Sulfation of L-selectin ligands by an HEV-restricted sulfotransferase regulates lymphocyte homing to lymph nodes. Immunity. 15:237–247. [DOI] [PubMed] [Google Scholar]
- 51.Hickey, M.J., M. Forster, D. Mitchell, J. Kaur, C. De Caigny, and P. Kubes. 2000. L-selectin facilitates emigration and extravascular locomotion of leukocytes during acute inflammatory responses in vivo. J. Immunol. 165:7164–7170. [DOI] [PubMed] [Google Scholar]
- 52.Tu, L., J.C. Poe, T. Kadono, G.M. Venturi, D.C. Bullard, T.F. Tedder, and D.A. Steeber. 2002. A functional role for circulating mouse L-selectin in regulating leukocyte/endothelial cell interactions in vivo. J. Immunol. 169:2034–2043. [DOI] [PubMed] [Google Scholar]
- 53.Chao, C.C., R. Jensen, and M.O. Dailey. 1997. Mechanisms of L-selectin regulation by activated T cells. J. Immunol. 159:1686–1694. [PubMed] [Google Scholar]
- 54.Miethke, T., C. Wahl, B. Holzmann, K. Heeg, and H. Wagner. 1993. Bacterial superantigens induce rapid and T cell receptor V beta-selective down-regulation of L-selectin (gp90Mel-14) in vivo. J. Immunol. 151:6777–6782. [PubMed] [Google Scholar]