

The Ig class switch allows the expression of a V region with new CH regions associated with various effector functions (for reviews, see references 1 and 2). Switch recombination (SR) occurs by an intrachromosomal deletion process in which the intervening genetic material between the switch (S) regions is excised as a circle. The recombination breakpoints are located within S regions but not at consensus recombination signal sequences, suggesting that SR is not site-specific recombination 3. Switch junctions do not contain long stretches of homology, arguing that homologous recombination does not contribute to this process. Thus, SR is unique in that it does not fall neatly into one of the well-defined categories of recombination. Results indicating an important role for mismatch repair (MMR) proteins in Ig class switch, published in a recent issue of EMBO (European Molecular Biology Organization) Journal 4 and in this issue of The Journal of Experimental Medicine 5, serve to significantly augment our understanding of the long-elusive molecular mechanism of Ig SR.
A model for the mechanism of SR was previously proposed in which heteroduplex DNA was located at DNA ends in recombination intermediates 6. Successful completion of the switch reaction required removal of the nonhomologous ends. In a perspicacious search for activities that mediate processing of heteroduplex DNA ends in SR, Schrader et al. 5 analyzed mice deficient for the MMR proteins Msh2, Pms2, Mlh1 and the compound knockout of Pms2/Mlh1 for their ability to support isotype switching in vitro. Extensive analysis of proliferation by tritiated thymidine uptake and of cell cycle by propidium iodide staining and flow cytometry demonstrated no difference between MMR-deficient and normal splenic B cells. MMR-deficient splenic B cells stimulated with LPS and the appropriate lymphokines displayed a 35–75% reduction in isotype switching to IgG3, IgG1, IgG2b, and IgA. This evaluation was based on cell surface staining for switched isotypes and was confirmed by digestion-circularization PCR. It is striking that the degree of isotype switch reduction is partial and somewhat variable among the isotypes.
In a screen for the effects of the MMR protein Msh2 on somatic hypermutation, it was noted that mice deficient in Msh2 could make good IgM but poor IgG responses to T cell–dependent antigen, suggesting that SR might be selectively affected 7. Ehrenstein and Neuberger 4 have now systematically examined this issue in Msh2-deficient mice to find that in both the antigen-specific T cell–dependent response to (4-hydroxy-3-nitrophenyl)acetyl–chicken γ-globulin (NP-CG) and the T cell–independent response to NP-Ficoll, the IgG response was diminished while the IgM response was intact. Analysis of the LPS or the LPS plus IL-4 response in B cells derived from mice deficient for Msh2 showed reduced switching to IgG3 or to IgG1, respectively, whereas proliferative responses were normal 4. Similar to the study by Schrader et al. 5, these findings reveal a partial reduction of isotype switching. The diminished response to T-independent antigen strongly suggests that this effect is intrinsic to B cells. In another recently published study of the immune response in Msh2-deficient mice, it was found that IgG expression was diminished but not abolished in antigen-specific T-dependent and -independent responses, whereas the LPS response was intact 8. Detection of an LPS response in Msh2-deficient B cells does not agree with the results of Schrader et al. 5 and Ehrenstein and Neuberger 4, and may be related to the method used to prepare splenic B cells for in vitro culture.
Classically, MMR maintains genetic stability by correcting mismatched nucleotides that arise from DNA replication errors, DNA damage, and genetic recombination 9. In Escherichia coli, the MutS, MutL, and MutH proteins provide the unique functions of mismatch recognition and DNA incision that are required to initiate bidirectional strand-specific repair 10. Homologues of MutS have been shown to function similarly in Saccharomyces cerevisiae and in humans. Both mice and humans have at least five homologues of MutS, termed Mut2–6. MMR proteins, most notably Msh2, bind to mismatched basepairs 11, heteroduplex loops 11, Holliday junctions 12, and other branched intermediates 13. What possible relationship can there be between isotype switching and MMR pathways? To address this question, we must digress for a moment into consideration of the mechanism of isotype class switch.
There has been a sneaking suspicion for the last several years that nonhomologous end joining (NHEJ), also called illegitimate recombination, mediates Ig class switching. Illegitimate recombination in mitotic cells is stimulated by DNA damage, leading to the formation of double strand breaks (DSBs). DSB repair is a process characterized by recombination junctions with distinct features 14 15 and the involvement of specific proteins in both yeast and mammalian cells 16 17. In some cases, DSBs repaired by NHEJ undergo blunt end ligation, which uses no homology between the ligated ends. More frequently, the recombination junctions contain microhomologies of one to five nucleotides and short deletions of genetic material. In mammalian cells, 97% of the end-joining events are within 15 nucleotides of the original broken end 15. Repair of a site-specific DSB is associated with introduction of mutations in the flanking DNA 18. In murine cells, DSBs are frequently subjected to extensive single strand 5′→3′ or 3′→5′ exonuclease digestion during NHEJ 19. The gap is thought to be resynthesized, resulting in little net loss of genetic material 19. The resynthesis of DNA by an error-prone DNA polymerase could introduce nucleotide substitutions which would be detected as mutations in the regions flanking the DSB.
The recombination junctions in composite S/S regions conform to the classical features associated with DSB repair by NHEJ. There is either no homology or microhomologies of one to eight nucleotides at the recombination breakpoints for S/S composite genes 3. Mutations flanking the switch junctions have been frequently observed in recombined S regions 3 20 21. It has been difficult to assess the degree of deletion at broken DNA ends since the precise locations of most DNA breaks in S regions are not known. However, if DNA breaks occur at specific sites in the S region tandem repeat, the processing that leads to short deletions at the DNA ends is predicted to result in clustered recombination breakpoints for switch junctions. Clustered recombination breakpoints in switch junctions of Sμ/Sγ3, Sμ/Sγ1, or Sμ/Sγ2b have been reported 21 22. Moreover, a mitogen-inducible DSB found in normal splenic B cells in Sγ3 DNA was located in the region of the clustered breakpoints 23. Taken together, these findings indicate that switch junctions contain the characteristic features associated with DSB repair by NHEJ.
NHEJ is a well-regulated process mediated by DNA-dependent protein kinase (DNA-PK) 16 17. DNA-PK is composed of the DNA-PK catalytic subunit (DNA-PKCS) and the Ku70/Ku80 heterodimer, which has DNA end-binding activity. The finding of DSBs in Sγ3 DNA and the dependency of SR on the DSB repair proteins Ku80 24, Ku70 25, and DNA-PKCS 26 strongly suggest that isotype switching occurs through an NHEJ mechanism. Alternatively, it may be that DSBs in S DNA are not related to the switching process but are normally repaired in a Ku-dependent pathway. Thus, viability of Ku-deficient B cells is poor in the presence of DSBs at S regions, making it appear as though SR is dependent on DNA-PK. Nonetheless, both the distinctive DNA sequence at switch junctions and the identification of a role for DSB repair proteins in SR are consistent with the tentative conclusion that isotype switching is mediated by NHEJ.
The key to understanding a possible role for MMR in SR requires consideration of the physical structure of the DNA ends that are involved in an NHEJ transaction. I begin with the assumption that blunt end DSBs, observed in S DNA 23, are indeed intermediates in the SR reaction (Fig. 1 A). There are two pathways in which a blunt DSB may be processed for use in NHEJ. First, the broken end may be directly ligated (Fig. 1 B). This is presumed to be an inefficient process, since these types of recombination products are relatively infrequent in populations of NHEJ events 15. Second, the broken end may be chewed back along one strand in a 5′→3′ (as shown in Fig. 1 C) or in the 3′→5′ direction (not shown). In some cases, microhomology between the recombinogenic ends will occur precisely at the DNA termini (Fig. 1 D). These events use an error-prone polymerase to fill in the residual gaps (Fig. 1 E). In other cases, short homologies will be located at more internal locations, leaving heteroduplex DNA at the ends (Fig. 1 F). The removal of the heteroduplex ends is postulated by both Schrader et al. 5 and Ehrenstein and Neuberger 4 to be MMR dependent (Fig. 1 G), and must be followed by gap fill-in. Precedent has been established for involvement of the MMR proteins Msh2 and Msh3 in the removal of heteroduplex ends during DSB repair by homologous recombination 13.
Figure 1.
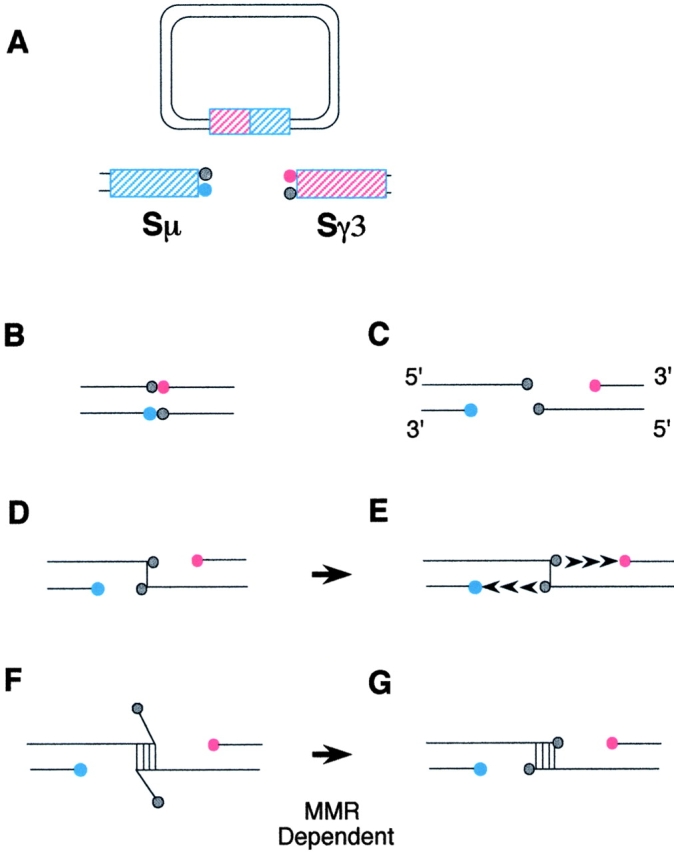
Schematic representation of SR by NHEJ. (A) SR is an intrachromosomal recombination event that occurs by a looping out and deletion of the genetic material between the Sμ (blue striped) and Sγ3 (red striped) DNA. Blunt DSBs in S DNA may be an intermediate in the SR reaction (reference 22). An excision circle is shown in which Sμ and Sγ3 have recombined and DSBs in Sμ (donor) and Sγ3 (acceptor) on the chromosome are formed. The 5′ ends are indicated by the blue (for Sμ) and red (for Sγ3) circles. The 3′ termini are indicated by gray circles. (B) The blunt end DSB may be used directly in a ligation reaction. (C) Single stranded DNA ends may be produced by either 5′→3′ (shown) or 3′→5′ (not shown) exonucleolytic hydrolysis at the donor and the acceptor ends. (D and F) This is followed by a microhomology search in which the donor and acceptor strands are annealed either precisely at DNA termini (D) or at internal locations (F). (E) When the annealing occurs directly at the DNA termini, the gaps are filled in and ligated by an error-prone 5′→3′ DNA synthesis event, as indicated by arrowheads. (G) It is speculated that when homologies are more internal, the heteroduplex ends must be removed by an MMR-dependent process. The gaps at the annealed processed ends are filled in and ligated by an error-prone 5′→3′ DNA synthesis event as in E.
The NHEJ model predicts that isotype switching will be reduced but not abolished by mutations in the MMR pathways, since only those recombination intermediates containing heteroduplex DNA require MMR activity for resolution. Moreover, the frequency at which microhomologies are found at DSB termini may be a function of S sequence. The observations of Schrader et al. 5 and Ehrenstein and Neuberger 4 demonstrating a partial reduction of SR in the absence of MMR and variation in the severity of this reduction as a function of isotype fit remarkably well with the NHEJ model for SR.
The NHEJ model for SR makes a second provocative prediction. Initial DSB formation may be intrinsically biased to consensus sites, as observed previously 23. In MMR-deficient mice, heteroduplex DNA ends cannot be processed, forcing the use of blunt ends, which can be directly abutted, or the pairing of ends that contain microhomologies. Switch junctions arising from these structures will be located at the original site of cleavage. Astoundingly, when Ehrenstein and Neuberger 4 examined Sμ/Sγ3 switch junctions from LPS blasts of MMR-deficient mice, they observed an altered distribution of the Sμ breakpoints that focused to the sequence GAGCT. The distribution of Sγ3 breakpoints was also skewed, and the sequence at which the Sγ3 recombination breakpoints clustered bears homology to the consensus sequence at the DSBs in Sγ3 23. These findings imply that MMR-dependent processing of SR intermediates obscured the originally specific cleavage sites.
Illegitimate recombination is initiated by DSBs formed in response to DNA damage and repaired by NHEJ in a DNA-PK–dependent pathway. There is also a site-specific form of recombination that uses NHEJ. In V(D)J joining, the DSB is formed by recombination activating gene RAG1 and RAG2 proteins and rejoined in a DNA-PK–dependent pathway. Similar to V(D)J joining, SR is likely to be initiated by formation of sequence-specific DSBs in S DNA by a developmentally regulated process and repaired via DNA-PK. The perturbation of SR by MMR deficiency gives us a glimpse of the recombination intermediates used and significantly informs our understanding of the molecular mechanism of the Ig class switch.
Acknowledgments
I wish to thank Drs. R. Wuerffel, A. Shanmugam, and J. Haber for the critical reading of this manuscript, and M. Neuberger and M. Ehrenstein for providing me with their manuscript before publication.
References
- Stavnezer J. Antibody class switching. Adv. Immunol. 1996;61:79–146. doi: 10.1016/s0065-2776(08)60866-4. [DOI] [PubMed] [Google Scholar]
- Lorenz M., Radbruck A. Developmental and molecular regulation of immunoglobulin class switch recombination. Curr. Top. Microbiol. Immunol. 1996;217:151–169. doi: 10.1007/978-3-642-50140-1_11. [DOI] [PubMed] [Google Scholar]
- Dunnick W., Hertz G.Z., Scappino L., Gritzmacher C. DNA sequences at immunoglobulin switch region recombination sites. Nucleic Acids Res. 1993;21:365–372. doi: 10.1093/nar/21.3.365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehrenstein M.R., Neuberger M.S. Deficiency in Msh2 affects the efficiency and local sequence specificity of immunoglobulin class-switch recombinationparallels with somatic hypermutation. EMBO (Eur. Mol. Biol. Organ.) J. 1999;18:3484–3490. doi: 10.1093/emboj/18.12.3484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schrader C.E., Edelmann W., Kucherlapati R., Stavnezer J. Reduced isotype switching in splenic B cells from mice deficient in mismatch repair enzymes. J. Exp. Med. 1999;190:323–330. doi: 10.1084/jem.190.3.323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunnick W., Stavnezer J. Copy choice mechanism of immunoglobulin heavy chain switch recombination. Mol. Cell. Biol. 1990;10:397–400. doi: 10.1128/mcb.10.1.397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rada C., Ehrenstein M.R., Neuberger M.S., Milstein C. Hot spot focusing of somatic hypermutation in Msh2 deficient mice suggests two stages of mutational targeting. Immunity. 1998;9:135–141. doi: 10.1016/s1074-7613(00)80595-6. [DOI] [PubMed] [Google Scholar]
- Vora K., Tumas-Brundage K.M., Lentz V.M., Cranston A., Fishel R., Manser T. Severe attenuation of the B cell immune response in Msh2-deficient mice. J. Exp. Med. 1999;189:471–481. doi: 10.1084/jem.189.3.471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freidberg E.C., Walker G.C., Siede W. DNA repair and mutagenesis. American Society for Microbiology, ; Washington, D.C.: 1995. [Google Scholar]
- Modrich P., Lahue R. Mismatch repair in replication fidelity, genetic recombination and cancer biology. Annu. Rev. Biochem. 1996;65:101–133. doi: 10.1146/annurev.bi.65.070196.000533. [DOI] [PubMed] [Google Scholar]
- Alani E., Chi N.W., Kolodner R.D. The Saccharomyces cerevisiae protein Msh2 binds to duplex oligonucleotides containing mismatched DNA basepairs and insertions. Genes Dev. 1995;9:234–247. doi: 10.1101/gad.9.2.234. [DOI] [PubMed] [Google Scholar]
- Alani E., Lee S., Kane M.F., Griffith J., Kolodner R.D. Saccharomyces cerevisiae MSH2, a mispaired base recognition protein, also recognizes Holliday junctions in DNA. J. Mol. Biol. 1997;265:289–301. doi: 10.1006/jmbi.1996.0743. [DOI] [PubMed] [Google Scholar]
- Sugawara N., Paques F., Colaiacovo M., Haber J. Role of Msh2 and Msh3 repair proteins in double-strand break-induced recombination. Proc. Natl. Acad. Sci. USA. 1997;94:9214–9219. doi: 10.1073/pnas.94.17.9214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kramer K.M., Brock J.A., Bloom K., Moore J.K., Haber J.E. Two different types of double strand breaks in Saccharomyces cerevisiae are repaired by similar RAD52 independent nonhomologous recombination events. Mol. Cell. Biol. 1994;14:1293–1301. doi: 10.1128/mcb.14.2.1293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roth D.B., Wilson J.H. Illegitimate recombination in mammalian cells. In: Kucherlapati R., Smith G.R., editors. Genetic Recombination. American Society for Microbiology; Washington, D.C.: 1988. pp. 621–653. [Google Scholar]
- Jeggo P., Taccioli G., Jackson S. Menage a troisdouble strand break repair, V(D)J recombination and DNA-PK. Bioessays. 1995;17:949–957. doi: 10.1002/bies.950171108. [DOI] [PubMed] [Google Scholar]
- Milne G.T., Jin S., Shannon K.B., Weaver D. Mutations in two Ku homologues define a DNA end-joining repair pathway in Saccharomyces cerevisiae . Mol. Cell. Biol. 1996;16:4189–4198. doi: 10.1128/mcb.16.8.4189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strathern J.N., Shafer B.K., McGill C.B. DNA synthesis errors associated with double strand break repair. Genetics. 1995;140:965–972. doi: 10.1093/genetics/140.3.965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henderson G.A., Simons P.J. Processing of DNA prior to illegitimate recombination in mouse cells. Mol. Cell. Biol. 1997;17:3779–3785. doi: 10.1128/mcb.17.7.3779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunnick W., Wilson M., Stavnezer J. Mutations, duplication, and deletion of recombined switch regions suggest a role for DNA replication in the immunoglobulin heavy-chain switch. Mol. Cell. Biol. 1989;9:1850–1856. doi: 10.1128/mcb.9.5.1850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du J., Zu Y., Shanmugam A., Kenter A.L. Analysis of immunoglobulin Sγ3 recombination breakpoints by PCRimplications for the mechanism of isotype switching. Nucleic Acids Res. 1997;25:3066–3073. doi: 10.1093/nar/25.15.3066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kenter A.L., Wuerffel R., Sen R., Jamieson C.E., Merkulov G.V. Switch recombination breakpoints occur at nonrandom positions in the Sγ tandem repeat. J. Immunol. 1993;151:4718–4731. [PubMed] [Google Scholar]
- Wuerffel R.A., Du J., Thompson R.J., Kenter A.L. Ig Sγ3 DNA-specific double strand breaks are induced in mitogen-activated B cells and are implicated in switch recombination. J. Immunol. 1997;159:4139–4144. [PubMed] [Google Scholar]
- Casellas R., Nussenzweig A., Wuerffel R., Pelanda R., Reichlin A., Suh H., Qin X.-F., Besmer E., Kenter A., Rajewsky K., Nussenzweig M.C. Ku80 is required for immunoglobulin isotype switching. EMBO (Eur. Mol. Biol. Organ.) J. 1998;17:2404–2411. doi: 10.1093/emboj/17.8.2404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manis J., Gu Y., Lansford R., Sonoda E., Ferrini R., Davidson L., Rajewsky K., Alt F. Ku70 is required for late B cell development and immunoglobulin heavy chain class switching. J. Exp. Med. 1998;187:2081–2089. doi: 10.1084/jem.187.12.2081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rolink A., Melchers F., Andersson J. The SCID but not the RAG-2 gene product is required for Sμ-S∈ heavy chain class switching. Immunity. 1996;5:319–330. doi: 10.1016/s1074-7613(00)80258-7. [DOI] [PubMed] [Google Scholar]