

Abstract
Dendritic cells (DCs) have the capacity to initiate immune responses, but it has been postulated that they may also be involved in inducing peripheral tolerance. To examine the function of DCs in the steady state we devised an antigen delivery system targeting these specialized antigen presenting cells in vivo using a monoclonal antibody to a DC-restricted endocytic receptor, DEC-205. Our experiments show that this route of antigen delivery to DCs is several orders of magnitude more efficient than free peptide in complete Freund's adjuvant (CFA) in inducing T cell activation and cell division. However, T cells activated by antigen delivered to DCs are not polarized to produce T helper type 1 cytokine interferon γ and the activation response is not sustained. Within 7 d the number of antigen-specific T cells is severely reduced, and the residual T cells become unresponsive to systemic challenge with antigen in CFA. Coinjection of the DC-targeted antigen and anti-CD40 agonistic antibody changes the outcome from tolerance to prolonged T cell activation and immunity. We conclude that in the absence of additional stimuli DCs induce transient antigen-specific T cell activation followed by T cell deletion and unresponsiveness.
Keywords: antigen delivery, DEC 205, dendritic cells, peripheral T cell tolerance, CD40
Introduction
Dendritic cells (DCs) are uniquely potent inducers of primary immune responses in vitro and in vivo 1 2. In tissue culture experiments, DCs are typically two orders of magnitude more effective as APCs than B cells or macrophages 3 4. In addition, purified antigen-bearing DCs injected into mice or humans migrate to lymphoid tissues and efficiently induce specific immune responses 5 6 7. Likewise, DCs migrate from peripheral tissues to lymphoid organs during contact allergy 8 9 and transplantation 10, two of the most powerful known stimuli of T cell immunity in vivo. Based on these and similar experiments, it has been proposed that the principal function of DCs is to initiate T cell–mediated immunity 1. However, nearly all of these experiments involved DC purification or culture in vitro, or some perturbations in vivo that induce major alterations in DC maturation and function. Thus, the physiologic function of DCs in the steady state has not been determined 6 11.
There is indirect evidence from a number of different laboratories suggesting that DCs may play a role in maintaining peripheral tolerance (summarized in reference 12). For example, injection of mice with 33D1, a rat monoclonal antibody to an unknown DC antigen, appeared to induce T cell unresponsiveness to the rat IgG 13. However, the specificity of antigen delivery was uncertain and the relevant T cell responses could not be analyzed directly. In addition, peripheral tolerance to ovalbumin and hemagglutinin expressed in pancreatic islets was found to be induced by bone marrow–derived APCs 14 15 16, but the identity of these antigen presenting cells has not been determined 17.
Materials and Methods
Mice.
6–8-wk-old females were used in all experiments and were maintained under specific pathogen free conditions. B10.BR, B6.SJL (CD45.1), and B6/MRL (Fas lpr) mice were purchased from The Jackson Laboratory. 3A9 transgenic mice were maintained by crossing with B10.BR mice. To obtain CD45.1 3A9 or 3A9/lpr T cells, B6.SJL or B6/MRL mice were crossed extensively with 3A9 mice and tested for CD45.1 and I-Ak, by flow cytometry. Fas lpr mutation was tested by PCR. Mice were injected subcutaneously with peptide in CFA and subcutaneously or intravenously with chimeric antibodies. All experiments with mice were performed in accordance with National Institutes of Health guidelines.
Flow Cytometry and Antibodies Used for Staining.
CD4- (L3T4), MHC II- (10-3.6), CD11c- (HL3), CD11c- (HL3), B220- (RA3-6B2), or CD3- (145-2C11), CD80(B7-1)-(16-10A1) I-Ak- (10-3.6) CD45.1- (A20), Il-2- (JES6-5H4), IFN-γ- (XMG1.2), CD40- (HM40-3-FITC), CD86(B7-2)- (GL1) specific antibodies were from BD PharMingen. Rat IgG-PE (goat anti–rat IgG) specific antibody was from Serotec. 3A9 T cell receptor (1G12)–specific antibody was a gift from Dr. Emil Unanue, Washington University, St. Louis, MO 18.
For visualization of rat IgGs on surface of mononuclear cells, lymphoid cells were purified from peripheral LNs 14 h after antibody injection and stained with anti–rat IgG-RPE (goat anti–rat IgG-RPE; Serotec) to visualize surface bound NLDC145 and GL117 antibodies. The cells were then incubated in mouse serum to block nonspecific binding and stained with FITC anti-CD11c (HL3), or -B220 (RA3-6B2), or -CD3 (145-2C11).
For intracellular cytokine staining, lymphocytes were stimulated in vitro for 4 h with leukocyte activation cocktail (BD PharMingen) according to the manufacturer's manual. Cells were fixed and permeabilized using cytofix/cytoperm buffer from BD PharMingen.
Immunohistology.
Popliteal LNs were removed from antibody injected mice and 5-μm cryosections (Microm; ZEISS) were prepared. Tissue specimens were fixed in acetone (5 min, room temperature [RT]) air dried, and stained in a moist chamber. The injected antibodies were detected by incubating the sections with streptavidin Cy3 or streptavidin-FITC (Jackson Immunotech). In double labeling experiments, the PE-conjugated antibodies were added for additional 30 min. Specimens were examined using a fluorescence microscope and confocal optical sections of ∼0.3-μm thickness were generated using deconvolution software (Metamorph).
Constructing and Production of Hybrid Antibodies.
Total RNA was prepared from NLDC-145 19 and GLII7 (gift of R.J. Hodes, National Institutes of Health, Bethesda, MD) hybridomas (both rat IgG2a) using Trizol (GIBCO BRL). Full-length Ig cDNAs were produced with 5′-RACE PCR kit (GIBCO BRL) using primers specific for 3′-ends of rat IgG2a and Ig kappa. The V regions were cloned in frame with mouse Ig kappa constant regions and IgG1 constant regions carrying mutations that interfere with FcR binding 20. DNA coding for hen egg lysozyme (HEL) peptide 46–61 with spacing residues on both sides was added to the C terminus of the heavy chain using synthetic oligonucleotides. Gene specific primers for cloning of rat IgG2a and Ig kappa: 3′-ATAGTTTAGCGGCCGCGATATCTCACTAACACTCATTCCTGTTGAAGCT; 3′-ATAGTTTAGCGGCCGCTCACTAGCTAGCTTTACCAGGAGAGTGGGAGAG-ACTCTTCT; HEL peptide fragment construction: 5′-CTAGCGACATGGCCAAGAAGGAGACAGTCTGGAGGCTCGAG-GAGTTCGGTAGGTTCACAAACAGGAAC; 5′-acagacgtagcacagactatggtattctccagattaacagcaggtattatgacggtaggacatgataggc; 3′-gctgtaccggttcttcctctgtcagacctccgagctcctcaa-gccatccaagtgtttgtccttgtgtctg; 3′-CCATCGTGTCTGATACCATAAGAGGTCTAATTGTCGTCCATAATACTGCCATCCTGTACTATCCGCCGG.
Hybrid antibodies were transiently expressed in 293 cells after transfection using calcium-phosphate. Cells were grown in serum-free DMEM supplemented with Nutridoma SP (Boehringer). Antibodies were purified on Protein G columns (Amersham Pharmacia Biotech). The concentrations of purified antibodies were determined by ELISA using goat anti–mouse IgG1 (Jackson Immunotech).
Cell Culture and Proliferation Assays.
Pooled axillary, brachial, inguinal, and popliteal LNs were dissociated in 5% FCS RPMI and incubated in presence of collagenase (Boehringer) and EDTA as described 21. For antigen presentation CD19+ and CD11c+ cells were purified using microbeads coupled to anti-mouse CD11c or CD19 IgG (Miltenyi Biotec) and irradiated with 1,500 rad. CD4 T cells were purified by depletion using rat antibodies supernatants specific for mouse: CD8 (TIB 211), B220 (RA3-6B2), MHC II (M5/114, TIB 120), F4/80 (F4/80), and magnetic beads coupled to anti–rat IgG (Dynal). In antigen loading experiments the isolated presenting cells from each experimental group were cultured in 96-well plates with 2 × 105 purified 3A9 CD4+ T cells. Cultures were maintained for 48 h with [3H]thymidine (1 μCi) added for the last 6 h. The results were calculated as a ratio of proliferation in experimental groups to a PBS control group. The proliferation in PBS controls ranged from 500 to 2,000 cpm.
For T cell proliferation assays in adoptive transfer recipients, 9 × 104 of the same irradiated CD11c+ cells isolated from spleens of wild-type B10.BR mice were cultured in 96-well plates with 3 × 105 T cells from each experimental group. Synthetic HEL peptide, at final concentration of 100 μg/ml, was added to half of the cultures. Cultures were maintained for 24 h with [3H]thymidine (1 μCi/ml) added for the last 6 h. Response to HEL peptide was determined by subtracting background (no HEL peptide added) proliferation from proliferation in the presence of HEL peptide. Proliferation index was calculated as the ratio of the response to HEL peptide in a given experimental group to the response to HEL of T cells from a PBS-injected control. Proliferation in PBS groups ranged from 4,000–8,000 cpm in the presence of peptide and the response to HEL peptide in these PBS controls was 1,000–3,000 counts above the background. Synthetic HEL 46-61 peptide was provided by the Howard Hughes Medical Institute Keck Biotechnology Resource Center.
Adoptive Transfer.
CD4 cells from 3A9 mice were enriched by depletion as described above, washed 3× with PBS, and 5 × 106 cells injected intravenously per mouse. Alternatively, before depletion total cells were labeled with 2 μM 5-(6)-carboxyfluorescein diacetate succinimidyl diester (CFSE) in 5% FCS RPMI (Molecular Probes) at 37°C for 20 min and washed twice.
Results
To examine the function of DCs in vivo, we devised a means of delivering antigens to DCs in situ. We used NLDC145 19, a monoclonal antibody specific for DEC-205, an endocytic receptor that is a member of a family of multilectin receptors including the macrophage mannose receptor (MMR) 22 23. Like MMR, DEC-205 displays an NH2-terminal cysteine-rich domain, a fibronectin type II domain, and multiple C-type lectin domains 22. However, the tissue distribution of DEC-205 and the MMR differ in that DEC-205 is highly expressed by DCs within the T cell areas of lymphoid tissues, particularly on CD8+ DCs that have been implicated in cross-priming 24, whereas the MMR is expressed by some tissue macrophages 25 26. We chose DEC-205 for targeting antigens to DCs because the cytoplasmic domain of DEC-205 orchestrates a distinct endocytic pathway that enhances antigen presentation 23. DEC-205 recycles through late endosomes or lysosomes rich in MHC II, and antigens delivered to these compartments by DEC-205 are efficiently processed and presented to T cells 23.
To determine whether the NLDC145 antibody targets DCs in vivo, we injected mice subcutaneously with purified NLDC145 or GL117, a nonspecific isotype-matched rat monoclonal antibody control, and visualized the injected antibody in tissue sections 24 h after injection, NLDC145 was found localized to scattered large dendritic profiles in the T cell areas of LNs and spleen while uptake of control GL117 was undetectable (Fig. 1 A, left and middle). This pattern was similar to the pattern found when the antibody was applied to sections directly (Fig. 1 A, right). The NLDC145-targeted cells were negative for B220 and CD4, markers for B cells and T cells, respectively, but positive for characteristic DC markers including MHC II and CD11c (Fig. 1 B). Thus, subcutaneously injected NLDC145 targets specifically to CD11c+MHC II+ DCs in lymphoid tissues in vivo.
Figure 1.
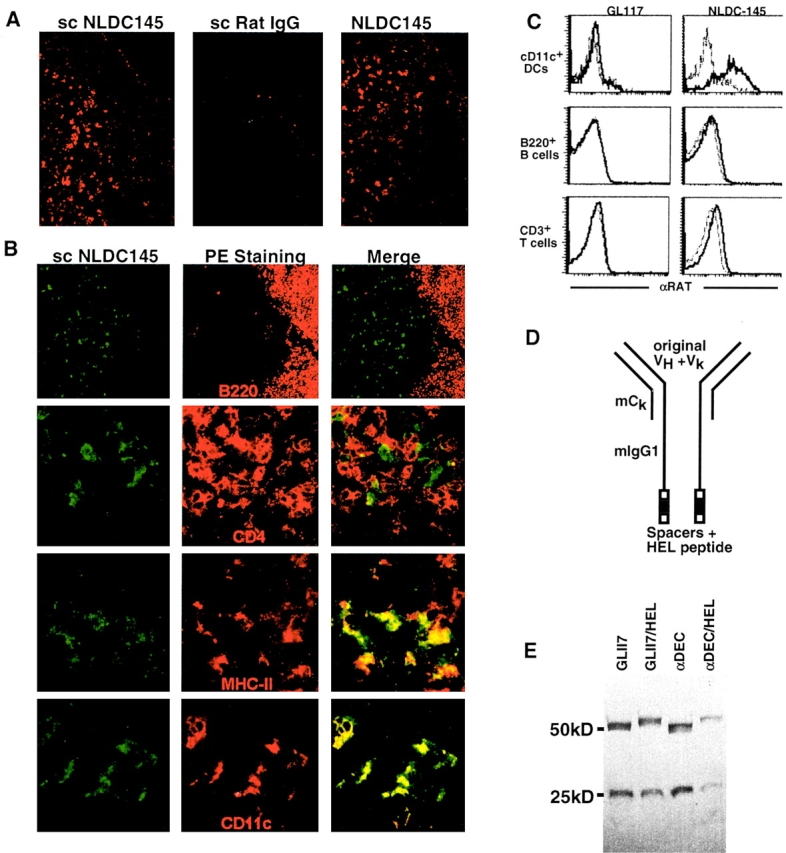
NLDC-145 targets DCs in vivo. (A) Biotinylated NLDC-145 (scNLDC145, left) or rat IgG (scRatIgG, middle) was injected into the hind footpads (50 μg/footpad) and inguinal LNs harvested 24 h later. Sections were stained with Streptavidin Cy3. Control sections from uninjected mice were stained using biotinylated NLDC145 and streptavidin Cy3 (NLDC145, right). (B) Two-color immunofluorescense. Mice were injected with biotinylated NLDC145 as in panel A. Sections were stained with streptavidin FITC (green) and PE-labeled antibodies (red) to B220 as indicated. Specimens were analyzed by deconvolution microscopy. Double labeling is indicated by the yellow color. (C) FACS® analysis of lymphoid cells 14 h after injection with NLDC145 and control GL117 antibody. Histograms show staining with anti–rat IgG on gated populations of CD11c+ DCs, B220+ B cells, and CD3+ T cells. (D) Diagrammatic representation of hybrid antibodies. (E) Hybrid antibodies. GL117, GL117/HEL, αDEC, and αDEC/HEL antibodies analyzed by PAGE under reducing conditions, molecular weights in kD are indicated.
To further characterize the lymphoid cells that were targeted by NLDC145 in vivo, we stained lymphoid cell suspensions from antibody injected mice with anti–rat Ig and examined the cells by multiparameter flow cytometry (Fig. 1 C). High levels of injected NLDC145 were found on the surface of most CD11c+ DCs but not on the surface of B220+ B cells or CD3+ T cells (Fig. 1 C). We conclude that when NLDC145 is injected into mice it binds efficiently and directly to DCs but not to other lymphoid cells.
To deliver antigens to DCs in vivo, we produced fusion proteins with amino acids 46–61 of HEL added to the COOH terminus of cloned NLDC145 (αDEC/HEL) and GL117 (GL117/HEL) control antibody (Fig. 1 D). To minimize antibody binding to Fc (FcR) receptors and further ensure the specificity of antigen targeting, the rat IgG2a constant regions of the original antibodies were replaced with mouse IgG1 constant regions that carry point mutations interfering with FcR binding 20. The hybrid antibodies and control Igs without the terminal HEL peptide (αDEC and GL117) were produced by transient transfection in 293 cells (Fig. 1 E).
To determine whether antigens delivered by αDEC/HEL were processed by DCs in vivo, we injected mice with the hybrid antibodies and controls and tested CD11c+ DCs, CD19+ B cells and CD11c−CD19− mononuclear cells for their capacity to present HEL peptide to naive HEL-specific T cells from 3A9 TCR transgenic mice 27. DCs isolated from antibody-injected mice expressed levels of CD80 and MHC II similar to those found on PBS controls and thus showed no signs of increased maturation, in contrast to what occurs when DCs are stimulated with microbial products like bacterial LPS and CpG deoxyoligonucleotides (28 29; Fig. 2 A). Nevertheless DCs from mice injected with αDEC/HEL induced strong T cell proliferative responses, whereas DCs isolated from PBS-injected mice or mice injected with the control antibodies had no effect (Fig. 2 B). DC isolated 3 d after αDEC/HEL injection showed reduced antigen-presenting activity (data not shown). In contrast to DCs, B cells and bulk CD11c−CD19− mononuclear cells purified from the same mice showed little antigen-presenting activity (Fig. 2 B). We conclude that antigens can be selectively and efficiently delivered to DC by αDEC/HEL in vivo, and the targeted DCs successfully process and load the peptides onto MHC II.
Figure 2.

DCs process and present antigen delivered by hybrid antibodies. (A) MHC II and CD80 expression on DCs is not altered by multiple injections of αDEC/HEL and 3A9 T cells. B10.BR mice transferred with 3A9 T cells and controls were injected subcutaneously in the footpads with 0.2 μg αDEC/HEL or PBS either at 8 d (αDEC/HEL) or at 1 and 8 d (αDEC/HELX2) after transfer (similar results were obtained by intravenous injection of chimeric antibodies, data not shown). 24 h after the last αDEC/HEL injection, DCs were purified from peripheral LNs and analyzed by flow cytometry for expression of CD80 and MHC II. Dotted lines in histograms indicate PBS control. (B) αDEC/HEL delivers HEL peptide to DCs in vivo. B10.BR mice were injected subcutaneously into footpads with 0.3 μg of αDEC/HEL or GL117/HEL or αDEC or PBS as indicated. CD11c+, CD19+, and CD11c−CD19− cells were isolated from draining LNs 24 h after antibody injection and assayed for antigen processing and presentation to purified 3A9 T cells in vitro. T cell proliferation was measured by [3H]thymidine incorporation and is expressed as a proliferation index relative to PBS controls. The results are means of triplicate cultures from one of four similar experiments.
As DC isolation leads to activation, we performed adoptive transfer experiments with HEL-specific transgenic T cells to follow the response of these T cells to otherwise unmanipulated, antigen-targeted DCs in vivo. CD4+ 3A9 T cells were transferred into B10.BR recipients and 24 h later hybrid antibodies were injected subcutaneously. To measure T cell responses, CD4+ cells were isolated from the draining LNs of the injected mice and cultured in vitro in the presence or absence of added HEL peptide. T cell responses were measured by [3H]thymidine incorporation and are shown as proliferation indices normalized to the PBS control (this index facilitates comparison between experiments, see Materials and Methods). In addition to αDEC/HEL, GL117/HEL, αDEC, and GL117 antibodies, we included 100 μg of HEL peptide in CFA as a positive control.
As described in previous reports 30 31, CD4+ T cells isolated 2 d after challenge with 100 μg of HEL peptide in CFA showed strong proliferative responses to antigen when compared with PBS controls (Fig. 3 A). Similar responses were obtained from mice injected with as little as 0.2 μg of αDEC/HEL (i.e., ∼4 ng peptide per mouse) but not from mice injected with up to 1 μg of αDEC, GL117, or GL117/HEL controls (Fig. 3 A, and not shown). We conclude that antigen delivered to DCs in vivo by αDEC/HEL efficiently induces activation of specific T cells.
Figure 3.

In vivo activation of CD4+ T cells by αDEC/HEL. In all experiments, 3A9 T cells were transferred into B10.BR mice, and the recipients were injected subcutaneously in the footpads with antibodies in PBS or 100 μg of HEL peptide in CFA 24 h after T cell transfer as indicated. T cell proliferation was measured by [3H]thymidine incorporation and is expressed as a proliferation index relative to PBS controls. (A) T cells are efficiently activated by antigen delivered by αDEC/HEL. 48 h after challenge with antigen, CD4 T cells were isolated from peripheral LNs and cultured in vitro with irradiated B10.BR CD11c+ cells in the presence or absence of HEL peptide. (B) CD4+ T cells are only transiently activated by antigen (αDEC/HEL 0.2 μg) delivered to DCs in vivo. CD4+ cells were purified from peripheral LNs 2 or 7 d after challenge with antigen and cultured with irradiated CD11c+ cells in the presence or absence of HEL peptide. (C) Failure to induce persistent T cell activation with multiple injections of αDEC/HEL. 3A9 cells were transferred into B10.BR mice and recipients were injected with αDEC/HEL (0.2 μg/mouse) once (on day 9 or 2 before analysis) or multiple times (days 9, 6, and 2 before analysis). Assay for T cell activation was as above. (D) T cells initially activated by αDEC/HEL show diminished response to rechallenge with HEL peptide in CFA. Recipients were initially injected with either αDEC/HEL (0.2 μg), GL117/HEL(0.2 μg), or PBS and rechallenged 7 or 20 d later with 100 μg of HEL peptide in CFA or with PBS. CD4+ cells were purified from peripheral LNs (or spleens, not shown) 2 d after the rechallenge and cultured with irradiated CD11c+ cells in the presence or absence of HEL peptide. Assay for T cell activation was as above. (E) Antigen loading of DCs with αDEC/HEL. B10.BR mice with or without transferred 3A9 T cells, were injected subcutaneously with 0.2 μg αDEC/HEL or PBS either at 8 d (αDEC/HEL) or at 1 and 8 d (αDEC/HELX2) after transfer. Antigen loading was measured 1 d after the last dose of αDEC/HEL by purifying CD11c+ DCs from peripheral LNs and culturing with purified 3A9 T cells. The results are means of triplicate cultures from one of three similar experiments.
To determine whether antigen delivered to DCs in vivo induces persistent T cell activation, we measured T cell responses to antigen 7 d after the administration of αDEC/HEL. CD4 T cells continued to show heightened responses to antigen when purified from LNs 7 d after injection with 100 μg of HEL peptide in CFA (30 31; Fig. 3 B). In contrast, T cells isolated from mice 7 d after injection with αDEC/HEL were no longer activated when compared with PBS controls (Fig. 3 B). Thus, T cell activation by antigen delivered to DCs by αDEC/HEL in vivo is transient, readily detected at 2 but not 7 d. This transient activation resembles the CD4 T cell response to large doses of peptide in the absence of adjuvant, or the response to self-antigens presented by bone marrow–derived antigen-presenting cells in the periphery 15 16 30 31 32. To determine whether the absence of persistent T cell activation in mice injected with αDEC/HEL is due to clearance of the injected antigen, multiple doses of αDEC/HEL were administered. Repeated injection of αDEC/HEL at 3-d intervals failed to induce prolonged T cell activation (Fig. 3 C). In addition, after 7 or 20 d, T cells initially activated by αDEC/HEL could not be reactivated when the mice were challenged with 100 μg of HEL peptide in CFA (Fig. 3 D). In contrast, comparable numbers of 3A9 T cells found in PBS-injected controls mounted a vigorous response to challenge with HEL peptide in CFA (compare Fig. 3 D and 4 C). Thus, the transient nature of the T cell response in mice injected with αDEC/HEL is not due to a lack of antigen, and T cells initially activated by DCs under physiologic conditions are unresponsive to subsequent challenge with antigen even in the presence of strong adjuvants.
Absence of persistent T cell responses could be due to DC deletion, T cell deletion, or induction of T cell anergy. To assess DC function in mice receiving multiple doses of αDEC/HEL, we isolated DCs from these mice and monitored presentation to 3A9 T cells in vitro (Fig. 3 E). DCs from mice injected with two doses of antibody showed the same T cell stimulatory activity as DCs isolated from mice receiving a single injection of αDEC/HEL (Fig. 3 E). In addition, the transfer of antigen specific T cells into αDEC/HEL recipients did not alter the ability of the isolated DCs to stimulate 3A9 T cells in vitro. Thus, the transient nature of the T cell response to DC-targeted antigens in vivo is not the result of a lack of antigen-bearing DCs.
To examine the fate of 3A9 T cells after exposure to antigen presented by DCs in vivo, we performed adoptive transfer experiments with CD45.1+ 3A9 T cells labeled with CFSE, a reporter dye for cell division. As described previously, T cells challenged with peptide in CFA divide, upregulate CD69 but not CD25, and produce IL-2 and IFN-γ but not IL-4 or IL-10. These cells are therefore considered to be Th1 polarized (30 31; Fig. 4A and Fig. B, and not shown). A burst of cell division and increase of CD69 but not CD25 expression was also seen after injection with 0.2 μg αDEC/HEL but not with GL117/HEL. Only clonotype positive CD4 cells showed these effects (Fig. 4A and Fig. c, and not shown). However, 3A9 cells activated by antigen presented on αDEC/HEL targeted DCs produced only IL-2 but not IFN-γ, IL-4, or IL-10 at the time of the assay and thus were not polarized to Th1 or Th2 phenotype 3 d after antigen challenge. (Fig. 4 B, and not shown). Therefore, 3A9 cells proliferate in response to αDEC/HEL targeted DCs in vivo, but the T cells do not produce a normal effector cell cytokine profile.
Figure 4.


CD4+ T cells divide in response to antigen presented by DCs in vivo, produce IL-2 but not IFN-γ, and are then rapidly deleted. (A) CFSE labeled CD45.1+ 3A9 T cells were transferred into B10.BR and 24 h later, the recipients were injected subcutaneously in the footpads with αDEC/HEL (0.2 μg), GL117/HEL (0.2 μg), HEL peptide in CFA, or PBS. CD4+ T cells were purified by negative selection from regional LNs 3 d after challenge with antigen and analyzed by flow cytometry. The plots show staining with 1G12 anti-3A9 and CFSE intensity on gated populations of CD4+CD45.1+ cells. The numbers indicate the percentage of CFSE high (undivided) and CFSE low (divided) CD4+ T cells. The results are from one of two similar experiments. (B) T cells produce IL-2 but not IFN-γ in response to antigens presented on DCs under physiological conditions. 3A9 cells were transferred into B10.BR mice and 24 h later the recipients were injected subcutaneously in the footpads with αDEC/HEL (0.2 μg), GL117/HEL (0.2 μg), HEL peptide in CFA. CD4+. Histograms show staining with anti–IL-2 and anti–IFN-γ on gated populations of 3A9+CD4+ cells. The thick lines indicate PBS control. (C) Same as in panel A but analysis performed 7 or 20 d after antigen administration.
Although there was persistent expansion of 3A9 T cells in regional LNs and spleen 7 and 20 d after challenge with HEL peptide in CFA (Fig. 4 C, spleen not shown), few 3A9 T cells survived in the LNs or spleen after exposure to antigen delivered by αDEC/HEL. Surviving cells appeared to be anergic as they could not be stimulated in vivo by HEL peptide in CFA (see Fig. 3 D). The loss of 3A9 T cells was Fas independent as it also occurred with 3A9/lpr T cells (Fig. 4 C, and not shown). Thus, the initial expansion of T cells in response to antigen presented by DCs in vivo is not sustained, and most of the initial responding T cells disappear from lymphoid organs by day 7. These cells are either deleted or persist in extravascular sites 33. If they do persist outside lymphoid tissues they must be anergic, because they cannot be activated by further exposure to antigen, including peptide in CFA (Fig. 3 D).
DCs can be stimulated to increase their antigen presenting activity and their immunogenic potential by exposure to bacterial products or CD40L 34 35 36, a TNF family member expressed on activated CD4 T cells, platelets, and mast cells 37. To determine whether the combination of costimulators and antigen delivery to DCs produces persistent T cell activation, mice were injected with αDEC/HEL and the agonistic anti-CD40 antibody FGK 45 38. In contrast to αDEC/HEL, the combination of αDEC/HEL and FGK 45 induced persistent T cell activation (Fig. 5 B). The level of T cell activation seen with αDEC/HEL and FGK 45 at day 7 was comparable to αDEC/HEL at day 2 or HEL peptide in CFA at day 2 and 7 (compare Fig. 3 B and 5 B). To determine whether anti-CD40 treatment altered 3A9 T cell numbers in αDEC/HEL-treated mice, we performed adoptive transfer experiments with CD45.1 allotype-marked T cells and assayed by flow cytometry. Whereas FGK 45 alone showed no effect on the number of 3A9 T cells in LNs at day 7, the combination of FGK 45 and αDEC/HEL induced persistent ∼8–10-fold expansion of 3A9 T cells, an increase similar to that seen with HEL peptide in CFA at day 7 (Fig. 5 A and 4). We conclude that persistent T cell responses can be induced by antigen delivered to DCs in vivo if an additional activation signal such as CD40 ligation is provided.
Figure 5.

CD40 ligation prolongs T cell activation in response to antigens delivered to DCs and induces upregulation of costimulatory molecules on DCs. (A) CD40 ligation induces persistent expansion of 3A9 cells in response to antigens delivered to DCs. CD45.1+ 3A9 T cells were transferred into B10.BR mice and 24 h later the recipients were injected subcutaneously in the footpads with 0.2 μg of αDEC/HEL alone or 90 μg of FGK45 or both or PBS. CD4+ T cells were purified by negative selection from regional LNs 7 d after challenge with antigen and analyzed by flow cytometry using antibodies specific for CD45.1 and CD4. The numbers indicate the percentages of CD4+CD45.1+ cells in LNs. (B) CD40 ligation prolongs T cell activation. 3A9 T cells were transferred into B10.BR mice and 24 h later, recipients were injected subcutaneously in the footpads with 0.2 μg of αDEC/HEL alone or 90 μg of FGK45 or both or PBS. After 2 or 7 d, CD4 T cells were isolated from the draining LNs and cultured in vitro with irradiated B10.BR CD11c+ cells in presence or absence of HEL peptide. T cell proliferation was measured by [3H]thymidine incorporation. The results represent triplicate cultures from two independent experiments. (C) CD40 ligation induces costimulatory molecules on DCs. B10.BR mice with or without 3A9 cell transfer were injected with 90 μg FGK45 plus 0.2 μg αDEC/HEL or αDEC/HEL or PBS. 3 d later DCs were isolated as in Fig. 2 and analyzed by flow cytometry using antibodies specific for CD11c, B220, CD86, and CD40. Histograms show staining with anti-CD40 and anti-CD86 on gated populations of DCs. Thick lines indicate control with PBS, which was same as αDEC/HEL alone.
To determine if CD40 ligation induced detectable phenotypic changes on DCs in our system, we analyzed DCs from mice transferred with 3A9 cells and injected with FGK 45 and αDEC/HEL. Consistent with work by others we found that those DCs upregulated their surface expression of CD40 and CD86 (39; Fig. 5 C). This increase was more pronounced in the presence of antigen-specific T cells suggesting a positive feedback mechanism between activated DCs and T cells (Fig. 5 C).
Discussion
Targeting Antigens to DCs In Situ through DEC-205.
Our results establish that antigens can be selectively delivered to DCs in vivo via the DEC-205 adsorptive endocytosis receptor. DEC-205, originally identified as an antigen recognized by the monoclonal antibody NLDC-145, offers several advantages as a receptor that will mediate antigen targeting to DCs in situ for purposes of antigen presentation. DEC-205 is expressed in abundance on DCs in the T cell area 19 40 and antibodies bound to DEC-205 are efficiently internalized and delivered to antigen processing compartments 22 23. When compared with the MMR, a closely related receptor, DEC-205 was at least 30 times more effective in antigen delivery to processing compartments 23. In vivo, anti–DEC-205 monoclonal antibody targets to DCs very efficiently, a dose of <1 μg of antibody (20 ng of HEL peptide) leading to presentation by DCs that was comparable to 100 μg of peptide in CFA (Fig. 3 A). It thus appears that the use of DEC-205 increases the efficiency of DC presentation by at least 100–1,000-fold relative to current adjuvants.
DCs Induce T Cell Tolerance in the Periphery.
Remarkably, when DEC-205 targeting is used to charge DCs with antigen in the steady state, these MHC II rich cells induce extensive T cell proliferation but do not induce normal Th-subset polarization or prolonged T cell expansion and activation. Instead, the T cells exposed to antigen on DCs in vivo either disappear or become anergic to antigenic restimulation. An initial burst of T cell proliferation followed by deletion was reported when ovalbumin or hemagglutinin were expressed as transgenes in the beta cells of the pancreatic islets 15 16. These, antigens were presented not by pancreatic beta cells but by bone marrow–derived APCs in the draining LNs. Our studies, using direct DC targeting of antigen, suggest that DCs are likely to be the tolerogenic APCs for self-antigens in vivo (for a review, see reference 17). Thus, in the steady state, the primary function of DCs is to maintain peripheral tolerance (Fig. 3C and Fig. D). While it has been proposed that immature antigen-bearing DCs can induce tolerance to transplantation antigens and contact allergens 41 42, the DEC-205 approach reveals the capacity of DCs to induce profound antigen-specific tolerance in vivo and in the steady state.
Our findings with peripheral T cells are concordant with the proposed function for DCs in the thymus 43 44 which is to establish tolerance in the medulla by inducing the negative selection of self-reactive T cells. Peripheral tolerance mechanisms are necessary, because self-reactive T cells escape thymic selection 45 and some self-antigens do not gain access to the thymus 46. Furthermore, foreign proteins found in the lumens of the airways and intestine do not normally initiate chronic inflammation. Nevertheless, DCs are likely to be continually internalizing potential self-antigens from tissues 47 and from noninfectious environmental proteins 48. We propose that in the steady state this uptake of proteins by DCs leads to peripheral tolerance.
Superficially, the idea that DCs induce tolerance appears to conflict with abundant evidence that DCs initiate immune responses 1. However, all prior work demonstrating the function of DCs as inducers of primary immune responses involved adoptive immunization with DCs cultured in vitro with antigen and then injected 6 11, or strong T cell responses in the setting of contact allergy and transplantation 8 9 10. Tissue disruption and inflammation alter DCs, increasing expression of critical costimulators like B7 49 50 MHC-peptide complexes 35 and chemokine receptors 51 52 53. These altered DCs are referred to as mature 34 53 54 55. The critical role of DC maturation in immunogenicity 1 is consistent with the idea that the immune system must focus on antigens delivered in the context of danger signals, some of which are registered by pattern recognition receptor 46 56 57. Indeed, combined administration of DC-targeted antigen with an agonistic anti-CD40 antibody that upregulates costimulatory molecules like CD86 on the surface of DCs (Fig. 5 C), prevents induction of peripheral tolerance and leads to prolonged T cell activation.
Our experiments are consistent with the notion that self-antigens such as serum components and apoptotic cells captured and presented to T cells by DCs under physiological conditions induce tolerance. In contrast, antigens taken up by DCs in the context of activation stimuli such as those found during inflammation or tissue destruction induce prolonged T cell activation. These two functions of DCs, maintaining tolerance to self and inducing immunity, are not in conflict because they are elicited under distinct circumstances, the steady state versus inflammation and infection. Moreover, the steady state tolerizing function of DCs may be essential for their subsequent role in eliciting immunity. During inflammation or infection, DCs present self-antigens simultaneously with non-self. By establishing tolerance to self and nonpathogenic environmental proteins before challenge with pathogens, DCs can focus the adaptive immune system entirely on the pathogen, thereby avoiding autoimmunity. The ability to target antigens to DCs and control their function in vivo has significant implications for development of vaccines and therapies for autoimmunity.
Acknowledgments
The authors thank Dr. Hitoshi Nagaoka for help with surgical procedures, Dr. Mark Davis for 3A9 mice, Dr. Fritz Melchers for FGK 45 hybridoma, Dr. Emil Unanue for 1G12 hybridoma, and Dr. Richard J. Hodes for the GL117 hybridoma. The authors also thank Dr. Eva Besmer for critical review of the manuscript.
This work was supported in part by Human Frontier Science (HFS) and grants to M.C. Nussenzweig and R.M. Steinman from the National Institute of Allergy and Infectious Diseases to R.M. Steinman and National Institutes of Health to M.C. Nussenzweig and AI13013, DK program project grant to M.C. Nussenzweig, R.M. Steinman, and J.V. Ravetch. M.C. Nussenzweig is a Howard Hughes Medical Institute investigator.
Footnotes
Abbreviations used in this paper: CFSE, 5-(6)-carboxyfluorescein diacetate succinimidyl diester; DC, dendritic cell; HEL, hen egg lysozyme; MMR, macrophage mannose receptor.
References
- Banchereau J., Steinman R.M. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. doi: 10.1038/32588. [DOI] [PubMed] [Google Scholar]
- Thery C., Amigorena S. The cell biology of antigen presentation in dendritic cells. Curr. Opin. Immunol. 2001;13:45–51. doi: 10.1016/s0952-7915(00)00180-1. [DOI] [PubMed] [Google Scholar]
- Inaba K., Steinman R.M., Van Voorhis W.C., Muramatsu S. Dendritic cells are critical accessory cells for thymus-dependent antibody responses in mouse and in man. Proc. Natl. Acad. Sci. USA. 1983;80:6041–6045. doi: 10.1073/pnas.80.19.6041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinman R.M., Gutchinov B., Witmer M.D., Nussenzweig M.C. Dendritic cells are the principal stimulators of the primary mixed leukocyte reaction in mice. J. Exp. Med. 1983;157:613–627. doi: 10.1084/jem.157.2.613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhodapkar M.V., Steinman R.M., Sapp M., Desai H., Fossella C., Krasovsky J., Donahoe S.M., Dunbar P.R., Cerundolo V., Nixon D.F., Bhardwaj N. Rapid generation of broad T-cell immunity in humans after a single injection of mature dendritic cells. J. Clin. Invest. 1999;104:173–180. doi: 10.1172/JCI6909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inaba K., Metlay J.P., Crowley M.T., Steinman R.M. Dendritic cells pulsed with protein antigens in vitro can prime antigen-specific, MHC-restricted T cells in situ. J. Exp. Med. 1990;172:631–640. doi: 10.1084/jem.172.2.631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lechler R.I., Batchelor J.R. Restoration of immunogenicity to passenger cell-depleted kidney allografts by the addition of donor strain dendritic cells. J. Exp. Med. 1982;155:31–41. doi: 10.1084/jem.155.1.31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macatonia S.E., Knight S.C., Edwards A.J., Griffiths S., Fryer P. Localization of antigen on lymph node dendritic cells after exposure to the contact sensitizer fluorescein isothiocyanate. Functional and morphological studies. J. Exp. Med. 1987;166:1654–1667. doi: 10.1084/jem.166.6.1654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moodycliffe A.M., Shreedhar V., Ullrich S.E., Walterscheid J., Bucana C., Kripke M.L., Flores-Romo L. CD40–CD40 ligand interactions in vivo regulate migration of antigen-bearing dendritic cells from the skin to draining lymph nodes. J. Exp. Med. 2000;191:2011–2020. doi: 10.1084/jem.191.11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larsen C.P., Morris P.J., Austyn J.M. Migration of dendritic leukocytes from cardiac allografts into host spleens. A novel pathway for initiation of rejection. J. Exp. Med. 1990;171:307–314. doi: 10.1084/jem.171.1.307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thurner B., Haendle I., Roder C., Dieckmann D., Keikavoussi P., Jonuleit H., Bender A., Maczek C., Schreiner D., von den Driesch P. Vaccination with mage-3A1 peptide-pulsed mature, monocyte-derived dendritic cells expands specific cytotoxic T cells and induces regression of some metastases in advanced stage IV melanoma. J. Exp. Med. 1999;190:1669–1678. doi: 10.1084/jem.190.11.1669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinman R.M., Turley S., Mellman I., Inaba K. The induction of tolerance by dendritic cells that have captured apoptotic cells. J. Exp. Med. 2000;191:411–416. doi: 10.1084/jem.191.3.411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finkelman F.D., Lees A., Birnbaum R., Gause W.C., Morris S.C. Dendritic cells can present antigen in vivo in a tolerogenic or immunogenic fashion. J. Immunol. 1996;157:1406–1414. [PubMed] [Google Scholar]
- Adler A.J., Marsh D.W., Yochum G.S., Guzzo J.L., Nigam A., Nelson W.G., Pardoll D.M. CD4+ T cell tolerance to parenchymal self-antigens requires presentation by bone marrow–derived antigen-presenting cells. J. Exp. Med. 1998;187:1555–1564. doi: 10.1084/jem.187.10.1555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurts C., Kosaka H., Carbone F.R., Miller J.F., Heath W.R. Class I–restricted cross-presentation of exogenous self-antigens leads to deletion of autoreactive CD8+ T cells. J. Exp. Med. 1997;186:239–245. doi: 10.1084/jem.186.2.239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan D.J., Kreuwel H.T., Sherman L.A. Antigen concentration and precursor frequency determine the rate of CD8+ T cell tolerance to peripherally expressed antigens. J. Immunol. 1999;163:723–727. [PubMed] [Google Scholar]
- Heath W.R., Carbone F.R. Cross-presentation, dendritic cells, tolerance and immunity. Annu. Rev. Immunol. 2001;19:47–64. doi: 10.1146/annurev.immunol.19.1.47. [DOI] [PubMed] [Google Scholar]
- Peterson D.A., DiPaolo R.J., Kanagawa O., Unanue E.R. Quantitative analysis of the T cell repertoire that escapes negative selection. Immunity. 1999;11:453–462. doi: 10.1016/s1074-7613(00)80120-x. [DOI] [PubMed] [Google Scholar]
- Kraal G., Breel M., Janse M., Bruin G. Langerhans' cells, veiled cells, and interdigitating cells in the mouse recognized by a monoclonal antibody. J. Exp. Med. 1986;163:981–997. doi: 10.1084/jem.163.4.981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clynes R.A., Towers T.L., Presta L.G., Ravetch J.V. Inhibitory Fc receptors modulate in vivo cytotoxicity against tumor targets. Nat. Med. 2000;6:443–446. doi: 10.1038/74704. [DOI] [PubMed] [Google Scholar]
- Inaba K., Pack M., Inaba M., Sakuta H., Isdell F., Steinman R.M. High levels of a major histocompatibility complex II–self peptide complex on dendritic cells from the T cell areas of lymph nodes. J. Exp. Med. 1997;186:665–672. doi: 10.1084/jem.186.5.665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang W., Swiggard W.J., Heufler C., Peng M., Mirza A., Steinman R.M., Nussenzweig M.C. The receptor DEC-205 expressed by dendritic cells and thymic epithelial cells is involved in antigen processing. Nature. 1995;375:151–155. doi: 10.1038/375151a0. [DOI] [PubMed] [Google Scholar]
- Mahnke K., Guo M., Lee S., Sepulveda H., Swain S.L., Nussenzweig M., Steinman R.M. The dendritic cell receptor for endocytosis, DEC-205, can recycle and enhance antigen presentation via major histocompatibility complex class II–positive lysosomal compartments. J. Cell Biol. 2000;151:673–684. doi: 10.1083/jcb.151.3.673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- den Haan J.M., Lehar S.M., Bevan M.J. CD8+ but not CD8− dendritic cells cross-prime cytotoxic T cells in vivo. J. Exp. Med. 2000;192:1685–1696. doi: 10.1084/jem.192.12.1685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo M., Gong S., Maric S., Misulovin Z., Pack M., Mahnke K., Nussenzweig M.C., Steinman R.M. A monoclonal antibody to the DEC-205 endocytosis receptor on human dendritic cells. Hum. Immunol. 2000;61:729–738. doi: 10.1016/s0198-8859(00)00144-0. [DOI] [PubMed] [Google Scholar]
- Linehan S.A., Martinez-Pomares L., Stahl P.D., Gordon S. Mannose receptor and its putative ligands in normal murine lymphoid and nonlymphoid organsin situ expression of mannose receptor by selected macrophages, endothelial cells, perivascular microglia, and mesangial cells, but not dendritic cells. J. Exp. Med. 1999;189:1961–1972. doi: 10.1084/jem.189.12.1961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ho W.Y., Cooke M.P., Goodnow C.C., Davis M.M. Resting and anergic B cells are defective in CD28-dependent costimulation of naive CD4+ T cells. J. Exp. Med. 1994;179:1539–1549. doi: 10.1084/jem.179.5.1539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Smedt T., Pajak B., Muraille E., Lespagnard L., Heinen E., De Baetselier P., Urbain J., Leo O., Moser M. Regulation of dendritic cell numbers and maturation by lipopolysaccharide in vivo. J. Exp. Med. 1996;184:1413–1424. doi: 10.1084/jem.184.4.1413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sparwasser T., Vabulas R.M., Villmow B., Lipford G.B., Wagner H. Bacterial CpG-DNA activates dendritic cells in vivoT helper cell-independent cytotoxic T cell responses to soluble proteins. Eur. J. Immunol. 2000;30:3591–3597. doi: 10.1002/1521-4141(200012)30:12<3591::AID-IMMU3591>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- Kearney E.R., Pape K.A., Loh D.Y., Jenkins M.K. Visualization of peptide-specific T cell immunity and peripheral tolerance induction in vivo. Immunity. 1994;1:327–339. doi: 10.1016/1074-7613(94)90084-1. [DOI] [PubMed] [Google Scholar]
- Van Parijs L., Peterson D.A., Abbas A.K. The Fas/Fas ligand pathway and Bcl-2 regulate T cell responses to model self and foreign antigens. Immunity. 1998;8:265–274. doi: 10.1016/s1074-7613(00)80478-1. [DOI] [PubMed] [Google Scholar]
- Aichele P., Brduscha-Riem K., Zinkernagel R.M., Hengartner H., Pircher H. T cell priming versus T cell tolerance induced by synthetic peptides. J. Exp. Med. 1995;182:261–266. doi: 10.1084/jem.182.1.261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reinhardt R.L., Khoruts A., Merica R., Zell T., Jenkins M.K. Visualizing the generation of memory CD4 T cells in the whole body. Nature. 2001;410:101–105. doi: 10.1038/35065111. [DOI] [PubMed] [Google Scholar]
- Caux C., Massacrier C., Vanbervliet B., Dubois B., Van Kooten C., Durand I., Banchereau J. Activation of human dendritic cells through CD40 cross-linking. J. Exp. Med. 1994;180:1263–1272. doi: 10.1084/jem.180.4.1263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inaba K., Turley S., Iyoda T., Yamaide F., Shimoyama S., Reis e Sousa C., Germain R.N., Mellman I., Steinman R.M. The formation of immunogenic major histocompatibility complex class II–peptide ligands in lysosomal compartments of dendritic cells is regulated by inflammatory stimuli. J. Exp. Med. 2000;191:927–936. doi: 10.1084/jem.191.6.927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sallusto F., Lanzavecchia A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony–stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J. Exp. Med. 1994;179:1109–1118. doi: 10.1084/jem.179.4.1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foy T.M., Aruffo A., Bajorath J., Buhlmann J.E., Noelle R.J. Immune regulation by CD40 and its ligand GP39. Annu. Rev. Immunol. 1996;14:591–617. doi: 10.1146/annurev.immunol.14.1.591. [DOI] [PubMed] [Google Scholar]
- Rolink A., Melchers F., Andersson J. The SCID but not the RAG-2 gene product is required for S mu-S epsilon heavy chain class switching. Immunity. 1996;5:319–330. doi: 10.1016/s1074-7613(00)80258-7. [DOI] [PubMed] [Google Scholar]
- Koch F., Stanzl U., Jennewein P., Janke K., Heufler C., Kampgen E., Romani N., Schuler G. High level IL-12 production by murine dendritic cellsupregulation via MHC class II and CD40 molecules and downregulation by IL-4 and IL-10 J. Exp. Med. 184 1996. 741 746[published erratum at 184:1590] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Witmer-Pack M.D., Swiggard W.J., Mirza A., Inaba K., Steinman R.M. Tissue distribution of the DEC-205 protein that is detected by the monoclonal antibody NLDC-145. II. Expression in situ in lymphoid and nonlymphoid tissues. Cell. Immunol. 1995;163:157–162. doi: 10.1006/cimm.1995.1110. [DOI] [PubMed] [Google Scholar]
- Fu F., Li Y., Qian S., Lu L., Chambers F., Starzl T.E., Fung J.J., Thomson A.W. Costimulatory molecule-deficient dendritic cell progenitors (MHC class II+, CD80dim, CD86−) prolong cardiac allograft survival in nonimmunosuppressed recipients. Transplantation. 1996;62:659–665. doi: 10.1097/00007890-199609150-00021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinbrink K., Wolfl M., Jonuleit H., Knop J., Enk A.H. Induction of tolerance by IL-10-treated dendritic cells. J. Immunol. 1997;159:4772–4780. [PubMed] [Google Scholar]
- Matzinger P., Guerder S. Does T-cell tolerance require a dedicated antigen-presenting cell? Nature. 1989;338:74–76. doi: 10.1038/338074a0. [DOI] [PubMed] [Google Scholar]
- Zal T., Volkmann A., Stockinger B. Mechanisms of tolerance induction in major histocompatibility complex class II-restricted T cells specific for a blood-borne self-antigen. J. Exp. Med. 1994;180:2089–2099. doi: 10.1084/jem.180.6.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouneaud C., Kourilsky P., Bousso P. Impact of negative selection on the T cell repertoire reactive to a self-peptidea large fraction of T cell clones escapes clonal deletion. Immunity. 2000;13:829–840. doi: 10.1016/s1074-7613(00)00080-7. [DOI] [PubMed] [Google Scholar]
- Matzinger P. Tolerance, danger, and the extended family. Annu. Rev. Immunol. 1994;12:991–1045. doi: 10.1146/annurev.iy.12.040194.005015. [DOI] [PubMed] [Google Scholar]
- Huang F.P., Platt N., Wykes M., Major J.R., Powell T.J., Jenkins C.D., MacPherson G.G. A discrete subpopulation of dendritic cells transports apoptotic intestinal epithelial cells to T cell areas of mesenteric lymph nodes. J. Exp. Med. 2000;191:435–444. doi: 10.1084/jem.191.3.435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vermaelen K.Y., Carro-Muino I., Lambrecht B.N., Pauwels R.A. Specific migratory dendritic cells rapidly transport antigen from the airways to the thoracic lymph nodes. J. Exp. Med. 2001;193:51–60. doi: 10.1084/jem.193.1.51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caux C., Vanbervliet B., Massacrier C., Azuma M., Okumura K., Lanier L.L., Banchereau J. B70/B7-2 is identical to CD86 and is the major functional ligand for CD28 expressed on human dendritic cells. J. Exp. Med. 1994;180:1841–1847. doi: 10.1084/jem.180.5.1841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inaba K., Witmer-Pack M., Inaba M., Hathcock K.S., Sakuta H., Azuma M., Yagita H., Okumura K., Linsley P.S., Ikehara S. The tissue distribution of the B7-2 costimulator in miceabundant expression on dendritic cells in situ and during maturation in vitro. J. Exp. Med. 1994;180:1849–1860. doi: 10.1084/jem.180.5.1849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ngo V.N., Tang H.L., Cyster J.G. Epstein-Barr virus-induced molecule 1 ligand chemokine is expressed by dendritic cells in lymphoid tissues and strongly attracts naive T cells and activated B cells. J. Exp. Med. 1998;188:181–191. doi: 10.1084/jem.188.1.181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schoenberger S.P., Toes R.E., van der Voort E.I., Offringa R., Melief C.J. T-cell help for cytotoxic T lymphocytes is mediated by CD40-CD40L interactions. Nature. 1998;393:480–483. doi: 10.1038/31002. [DOI] [PubMed] [Google Scholar]
- Sallusto F., Schaerli P., Loetscher P., Schaniel C., Lenig D., Mackay C.R., Qin S., Lanzavecchia A. Rapid and coordinated switch in chemokine receptor expression during dendritic cell maturation. Eur. J. Immunol. 1998;28:2760–2769. doi: 10.1002/(SICI)1521-4141(199809)28:09<2760::AID-IMMU2760>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- Pierre P., Turley S.J., Gatti E., Hull M., Meltzer J., Mirza A., Inaba K., Steinman R.M., Mellman I. Developmental regulation of MHC class II transport in mouse dendritic cells. Nature. 1997;388:787–792. doi: 10.1038/42039. [DOI] [PubMed] [Google Scholar]
- Schuler G., Steinman R.M. Murine epidermal Langerhans cells mature into potent immunostimulatory dendritic cells in vitro. J. Exp. Med. 1985;161:526–546. doi: 10.1084/jem.161.3.526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janeway C. Immunogenicity signals 1,2,3 … and 0. Immunol. Today. 1989;10:283–286. doi: 10.1016/0167-5699(89)90081-9. [DOI] [PubMed] [Google Scholar]
- Medzhitov R., Janeway C., Jr. Innate immune recognitionmechanisms and pathways. Immunol. Rev. 2000;173:89–97. doi: 10.1034/j.1600-065x.2000.917309.x. [DOI] [PubMed] [Google Scholar]