

Abstract
Toll-like receptors (TLRs) are ancient microbial pattern recognition receptors highly conserved from Drosophila to humans. To investigate if subsets of human dendritic cell precursors (pre-DC), including monocytes (pre-DC1), plasmacytoid DC precursors (pre-DC2), and CD11c+ immature DCs (imDCs) are developed to recognize different microbes or microbial antigens, we studied their TLR expression and responses to microbial antigens. We demonstrate that whereas monocytes preferentially express TLR 1, 2, 4, 5, and 8, plasmacytoid pre-DC strongly express TLR 7 and 9. In accordance with these TLR expression profiles, monocytes respond to the known microbial ligands for TLR2 (peptidoglycan [PGN], lipoteichoic acid) and TLR4 (lipopolysaccharide), by producing tumor necrosis factor (TNF)-α and interleukin (IL)-6. In contrast, plasmacytoid pre-DCs only respond to the microbial TLR9-ligand, CpG-ODNs (oligodeoxynucleotides [ODNs] containing unmethylated CpG motifs), by producing IFN-α. CD11c+ imDCs preferentially express TLR 1, 2, and 3 and respond to TLR 2-ligand PGN by producing large amounts of TNF-α, and to viral double-stranded RNA-like molecule poly I:C, by producing IFN-α and IL-12. The expression of distinct sets of TLRs and the corresponding difference in reactivity to microbial molecules among subsets of pre-DCs and imDCs support the concept that they have developed through distinct evolutionary pathways to recognize different microbial antigens.
Keywords: immunity,; natural; bacteria; receptors, immunologic; monocytes; cytokines
Introduction
Dendritic cells (DCs) have an outstanding ability to prime naive T cells and to initiate primary T cell–mediated immune responses 1. DCs also dictate the development of T cell–mediated immune responses into either Th1 or Th2 type 2 3 4. Recent studies generated considerable debate as to how DCs regulate the types of T cell responses 3 4 5 6 7. An evolutionary theory suggests that two DC lineages or subsets can induce Th1 or Th2 responses, respectively, under certain circumstances 2 4. In mice, the CD8α+ DC, but not CD8α− DC subset, has the capacity to produce a large amount of IL-12 and induce Th1 versus Th2 responses 5 6. In humans, CD40 ligand (CD40L)-activated monocyte-derived DCs (DC1), but not plasmacytoid pre-DC–derived DCs (DC2), produce a large amount of IL-12 and induce Th1 versus Th2 responses 7. However, several studies have demonstrated that a given DC subset can induce either Th1 or Th2 response depending on the types of stimulation and pathogens 8 9 10. From these studies, an environmental instruction theory was suggested that questioned the existence of distinct DC lineages specialized to the induction of different types of T cell responses 3.
Evolutionary force drove the development of multiple subsets of B and T lymphocytes, which rapidly and efficiently respond to common microbial antigens. Unlike conventional B and T cells, B-1 B cells, γ/δ T cells, and natural killer T cells express restricted and distinct antigen receptors, capable of recognizing common antigens derived from bacteria or damaged host cells 11 12 13 14. Thus, if separate DC lineage/subsets with specialized functions really exist, they may express receptors that recognize different microbial antigens. Toll-like receptors (TLRs) are ancient receptors that are highly conserved from Drosophila to humans and recognize molecular patterns specific to microbial pathogens 15 16 17 18 19. Signaling through TLRs strongly activates DCs to upregulate costimulatory molecules (CD80 and CD86) and to produce proinflammatory cytokines (TNF-α, IL-6, and IL-12) 17 18 19. We examined the expression of all known human TLRs (TLR1–TLR10) on ex vivo subsets of human blood pre-DCs and immature DCs (imDCs) and their responses to microbial antigens with different pattern structures.
Materials and Methods
Isolation and Culture of Cells.
Blood CD11c + imDCs, plasmacytoid pre-DCs, and CD14 2+CD16− monocytes were isolated from human peripheral blood as described 7 20. The purity of each cell population was >99%. Monocytes were cultured for 5 d in RPMI 1640 (BioWhittaker) supplemented with 10% FCS (BioWhittaker), 2 mM L-glutamine, 10 mM HEPES, 1 mM sodium pyruvate, 55 μM 2-mercaptoethanol, penicillin G, and streptomycin (all from Life Technologies), in the presence of 50 ng/ml GM-CSF (Schering-Plough) and 200 U/ml IL-4 (Schering-Plough). The resulting monocyte-derived imDCs were washed and cultured for 24 h with human CD40L-transfected L cells (irradiated at 5,500 rad) to obtain mature DC1 7. Plasmacytoid pre-DCs were cultured for 5 d with 10 ng/ml IL-3 (R&D Systems). The resulting plasmacytoid pre-DC-derived imDCs were washed and cultured for 24 h with CD40L-transfected L cells to obtain plasmacytoid pre-DC–derived DCs. To induce the maturation of imDCs, they were cultured for 24 h with CD40L-transfected L cells. To induce cytokine production, monocytes, CD11c+ imDCs, or plasmacytoid pre-DCs were cultured for 24 h at 2 × 104/200 μl in round-bottom 96-well culture plates in the presence of 10 μg/ml peptidoglycan (PGN) from Staphylococcus aureus (Fluka), 10 μg/ml of lipoteichoic acid (LTA) from S. aureus (Sigma-Aldrich), 10 μg/ml LPS from S. minnesota serotype Re595 (Sigma-Aldrich), 50 μg/ml poly (I:C) (Sigma-Aldrich), 5 μM (46 μg/ml) phosphodiester CpG-oligodeoxynucleotide (ODN) AAC-30 21 22. AAC-30 was added at 0, 4, and 16 h to compensate for their degradation by DNase activity in medium.
Reverse Transcription PCR for TLRs.
RNA was isolated with the acid guanidinium thiocyanate–phenol-chloroform method 23. Contaminating DNA was removed by digestion with 5 U deoxyribonuclease I (Boehringer) for 30 min at 37°C. Reverse transcription (RT) was performed with random hexamers (Promega) for priming and SuperScript™II (Life Technologies). The PCR reaction volume was 50 μl, containing 0.5 μM of each primer, 40 nM of each deoxynucleoside triphosphate, and 1.25 U AmpliTaq (PerkinElmer). Primers used are as shown in Table . A GeneAmp PCR System 9700 (PerkinElmer/Applied Biosystems) was used with an initial denaturation step of 94°C for 5 min, followed by 35 cycles of 94°C for 30 s, 55°C for 30 s, 72°C for 1 min, and a final elongation step of 72°C for 7 min. PCR products were separated on a 3% agarose gel containing ethidium bromide. A 1-kb DNA ladder standard (Life Technologies) was used as a size marker.
Table 1.
Sequences of PCR Primers
| RT-PCR primers | ||
|---|---|---|
| TLR | Forward primers | Reverse primers |
| 1 | CGTAAAACTGGAAGCTTTGCAAGA | CCTTGGGCCATTCCAAATAAGTCC |
| 2 | GGCCAGCAAATTACCTGTGTG | CCAGGTAGGTCTTGGTGTTCA |
| 3 | ATTGGGTCTGGGAACATTTCTCTTC | GTGAGATTTAAACATTCCTCTTCGC |
| 4 | CTGCAATGGATCAAGGACCA | TCCCACTCCAGGTAAGTGTT |
| 5 | CATTGTATGCACTGTCACTC | CCACCACCATGATGAGAGCA |
| 6 | TAGGTCTCATGACGAAGGAT | GGCCACTGCAAATAACTCCG |
| 7 | AGTGTCTAAAGAACCTGG | CTTGGCCTTACAGAAATG |
| 8 | CAGAATAGCAGGCGTAACACATCA | AATGTCACAGGTGCATTCAAAGGG |
| 9 | TTATGGACTTCCTGCTGGAGGTGC | CTGCGTTTTGTCGAAGACCA |
| 10 | CAATCTAGAGAAGGAAGATGGTTC | GCCCTTATAAACTTGTGAAGGTGT |
| β-actin | ATCTGGCACCACACCTTCTACAATGAGCTGCG | CGTCATACTCCTGCTTGCTGATCCACATCTGC |
| Real-time PCR primers | ||
| TLR | Forward primers | Reverse primers |
| 2 | GGCCAGCAAATTACCTGTGTG | AGGCGGACATCCTGAACCT |
| 4 | CTGCAATGGATCAAGGACCA | TTATCTGAAGGTGTTGCACATTCC |
| 7 | TTACCTGGATGGAAACCAGCTACT | TCAAGGCTGAGAAGCTGTAAGCTA |
| 9 | TGAAGACTTCAGGCCCAACTG | TGCACGGTCACCAGGTTGT |
Real-Time Quantitative RT-PCR.
RNA was isolated with the acid guanidinium thiocyanate–phenol-chloroform method 23. The RT was performed with SuperScript™II (Life Technologies). cDNA was analyzed for the expression of TLR genes by the Fluorogenic 5′-nuclease PCR assay 7 using a PerkinElmer ABI Prism 7700 Sequence Detection System (Applied Biosystems). Reactions were incubated for 2 min at 50°C, denatured for 10 min at 95°C, and subjected to 40 two-step amplification cycles with annealing/extension at 60°C for 1 min followed by denaturation at 95°C for 15 s. Primers for TLRs are shown in Table . Values are expressed as arbitrary units (relative to ubiquitin ×1,000).
Quantitation of Cytokines by ELISA.
ELISA kits from the following companies were used to analyze cytokine production: TNF-α, IL-6 (R&D Systems), IL-12, and IFN-α (Biosource International).
Results
Three populations of cells, monocytes, CD11c + imDCs, and plasmacytoid pre-DCs, were isolated by cell sorting, to a purity >99%. As shown in Fig. 1, monocytes expressed high levels of TLR 1, 2, 4, 5, and 8, low level of TLR6, and undetectable levels of TLR 3, 7, 9, or 10 by RT-PCR. CD11c+ imDCs expressed high levels of TLR 1, 2, and 3, low levels of TLR 5, 6, 8, and 10, and undetectable levels of TLR 4, 7, and 9. In marked contrast to monocytes and CD11c + imDCs, plasmacytoid pre-DCs express high levels of TLR 7 and 9 (Fig. 1). They also expressed low levels of TLR 1, 6, and 10, but not TLR 2, 3, 4, 5, or 8. Thus, monocytes and CD11c + imDCs, and plasmacytoid pre-DCs express distinct sets of TLRs.
Figure 1.
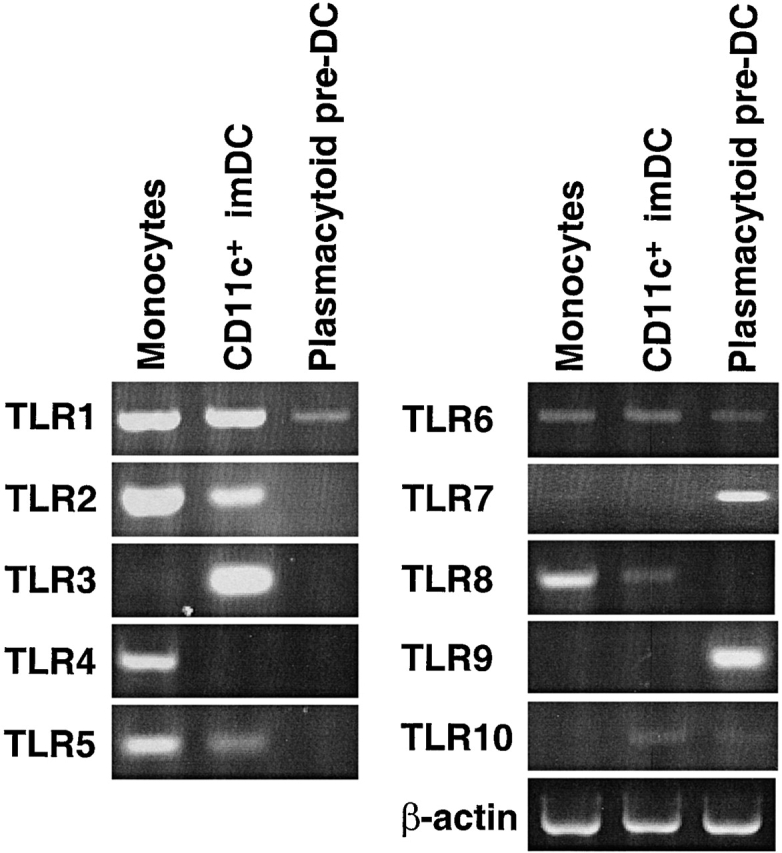
Detection of TLR mRNA by RT-PCR in freshly isolated monocytes, CD11c+ imDCs, and plasmacytoid pre-DCs. The three cell populations were isolated by cell sorting to the purity of >99%. cDNAs were amplified for 35 cycles and were separated on a 3% agarose gel containing ethidium bromide. The data shown are representative of three experiments.
The expression of TLR 2, 4, 7, and 9 was further analyzed by real-time quantitative RT-PCR, because of their selective expression on monocytes or plasmacytoid pre-DCs. As shown in Fig. 2, Monocytes express the highest levels of TLR2 and TLR4 of the cell populations examined. The expression of these two receptors is dramatically downregulated upon differentiation of monocytes into imDCs after 5 d of culture with GM-CSF plus IL-4 and is almost lost after further 24 h activation with CD40L. CD11c+ imDCs express moderate levels of TLR2 and TLR4, similar to those expressed by imDCs generated in culture (Fig. 2). The expression of TLR2 and TLR4 on CD11c+ imDCs is downregulated by CD40L activation. Plasmacytoid pre-DCs do not express TLR2 and TLR4 at any stages of maturation, but express the highest levels of TLR7 and TLR9 (Fig. 2). The expression of TLR7 and TLR9 is progressively down-regulated after differentiation into imDCs with IL-3 and then into mature DCs after CD40L activation. Monocytes and CD11c+ imDCs do not express significant levels of TLR7 and TLR9 at any stages of maturation.
Figure 2.

Quantitation of TLR mRNA by real-time quantitative RT-PCR in freshly isolated monocytes (Mono), CD11c+ imDCs, and plasmacytoid pre-DCs (pDC), and in immature and mature DCs induced from them. Monocytes were cultured with 50 ng/ml GM-CSF and 200 U/ml IL-4 for 5 d to obtain imDC1 and were further stimulated with CD40L-transfected L cells for 24 h to obtain mDC1. CD11c+ imDCs were stimulated with CD40L-transfected L cells for 24 h to obtain mature CD11c+ DCs. Plasmacytoid pre-DCs were cultured with 10 ng/ml IL-3 for 5 d to obtain imDC2 and were further stimulated with CD40L-transfected L cells for 24 h to obtain mature DC2. The amounts of mRNA were quantitated by real-time quantitative RT-PCR, and were shown by arbitrary unit relative to the amount of ubiquitin mRNA. The data shown are representative of three experiments.
Microbial molecules that trigger TLR2, TLR4, and TLR9 signaling have been identified. PGN and LTA from Gram-positive bacteria signal through TLR2 24 25 26 27. LPS from Gram-negative bacteria signals through TLR2 26. Bacterial DNA containing unmethylated CpG motifs signals through TLR9 28 29 30. It is unclear up to now which receptor mediates signaling triggered by poly I:C, a viral double-stranded RNA-like molecule. Based on the TLR-expression data above, we determined whether monocytes, CD11c+ imDCs, and plasmacytoid pre-DCs respond to the different microbial TLR-ligands and poly I:C, by producing the proinflammatory cytokines TNF-α, IL-6, IL-12, and IFN-α.
As shown in Fig. 3, the TLR2-ligand, PGN, stimulated monocytes to produce large amounts of TNF-α and IL-6. PGNs stimulate CD11c+ imDCs to produce large amounts of TNF-α, and small amounts of IL-6 and IL-12. PGN did not stimulate plasmacytoid pre-DCs to produce any of the cytokines tested. Plasmacytoid pre-DCs died in cultures either with medium or with PGN (data not shown). Similar to PGN, the other TLR2-ligand, LTA, stimulated monocytes to produce large amounts of TNF-α and IL-6. However, unlike PGN, LTA did not stimulate CD11c+ imDCs to produce detectable levels of cytokines tested. Plasmacytoid pre-DCs did not respond to LTA.
Figure 3.

TNF-α, IL-6, IFN-α, and IL-12 production by monocytes (Mono), CD11c+ imDCs, and plasmacytoid pre-DCs (pDC) stimulated with microbial molecules that trigger different TLR signaling. The three cell populations were cultured with 10 μg/ml PGN, 10 μg/ml LTA, 10 μg/ml LPS, 50 μg/ml poly I:C, or with 5 μM CpG-ODN AAC-30 for 24 h, and the concentrations of cytokines in the supernatants were measured by ELISA. The data shown are representative of four or five experiments.
The TLR4-ligand, LPS, stimulated monocytes to produce large amounts of TNF-α and IL-6. LPS stimulated CD11c+ imDCs to produce moderate amounts of IL-6. LPS also stimulated CD11c+ imDCs to produce small amounts of IL-12p75 in two out of four donors. LPS did not stimulate plasmacytoid pre-DCs to produce any cytokines tested and cell die in cultures with LPS or medium.
The TLR9-ligand, CpG-ODN AAC-30, selectively stimulated plasmacytoid pre-DCs, but not monocytes and CD11c+ imDCs, to produce IFN-α. Interestingly, poly I:C selectively stimulated CD11c+ imDCs, but not monocytes and plasmacytoid pre-DCs to produce IFN-α and IL-12p75.
Discussion
In this study, we demonstrated that monocytes, CD11c + imDCs, and plasmacytoid pre-DCs express distinct sets of TLRs. Monocytes preferentially express TLR 1, 2, 4, 5, and 8, CD11c+ imDCs selectively express TLR3, and plasmacytoid pre-DCs strongly express TLR 7 and 9. In accordance with the TLR expression patterns, monocytes respond to microbial molecules known to trigger signaling via TLR2 (PGN, LTA) and TLR4 (LPS), but not via TLR9 (CpG-ODN). By contrast, plasmacytoid pre-DCs, but not monocytes and CD11c+ imDCs respond to the TLR9-ligand, CpG-ODN AAC-30. Interestingly, CD11c+ imDCs, but not monocytes and plasmacytoid pre-DCs responded to poly I:C. These remarkable differences among monocytes, CD11c+ imDCs, and plasmacytoid pre-DCs, in their TLR repertoire expression and responsiveness to microbial antigens, suggests that cells have evolved to recognize different pathogens. This study also sheds some light on how different TLRs instruct DC precursors and imDCs to produce different sets of proinflammatory cytokines and dictate the types of adaptive immune responses. While the expression of TLR2 and TLR4 endorse monocytes with the ability to produce large amounts of proinflammatory cytokines TNF-α and IL-6, the expression of TLR9 endorse plasmacytoid pre-DCs with the ability to produce IFN-α during antibacterial immune responses.
Recent studies on DCs have generated considerable debates as to whether there is a functionally distinct lymphoid DC lineage or subset 1 2 3 4 5 6 7 8 9 10. The existence of a lymphoid DC lineage was suggested by the studies showing that DCs can be derived from the earliest T cell precursors within the thymus 31 or from CD19+ pro-B cells in cultures 32. In humans, plasmacytoid pre-DCs were suggested to be of lymphoid origin, because of their expression of lymphoid but not myeloid makers and their shared developmental pathway with T and B cells 20 33 34. The present finding that monocytes and plasmacytoid pre-DCs express distinct sets of TLRs suggests that these two DC precursors may have developed through different evolutionary trails, further supporting the existence of distinct DC lineages.
This study also clarifies the debates on whether plasmacytoid pre-DCs have the ability to produce IL-12 in response to LPS or CD40-ligand, or to produce IFN-α in response to CD40-ligand. Previous studies demonstrated that plasmacytoid pre-DCs did not produce significant amounts of IL-12p75 in respond to activation with CD40-ligand, CpG, LPS, or viruses 7 10 35. However, studies by Cella et al. showed that plasmacytoid pre-DCs isolated according to the expression of two immunoglobulin-like receptors (ILT3+ILT1−) produce large amounts of IL-12p75 in response to LPS (808 pg/ml) and to CD40-ligand (2,100 pg/ml) 36 37. In addition, Cella et al. showed that ILT3+ILT1− plasmacytoid pre-DCs produced IFN-α in response to CD40-ligand 36. These results could not be confirmed by the present and previous studies 7 10 35. The finding that plasmacytoid pre-DCs do not express TLR4 and do not respond to LPS in terms of cell survival and cytokine production, suggests that the observed IL-12 production by ILT3+ILT1− plasmacytoid pre-DCs induced by LPS or CD40-ligand and their ability to induce TH1 responses may be due the contaminating myeloid DCs or macrophages 36 37 38. Indeed, ILT3+ILT1− plasmacytoid pre-DCs appear to contain 10–30% of cells that do not express high IL-3Rα, a key marker for human pre-DC2 37.
Plasmacytoid pre-DCs have been shown to produce an enormous amount of IFN-α in response to viruses 10 36 39 40, indicating that these cells play a critical role in antiviral innate immunity. The present finding that plasmacytoid pre-DCs express high levels of TLR9 and produce IFN-α in response to TLR9-ligand, bacteria CpG, extend the their function into antibacterial innate immunity.
Although CD11c+ imDCs express TLR-2, only PGN, but not LTA, stimulated CD11c+ imDCs to produce large amounts of TNF-α. This suggests that TLR2 is the primary receptor for PGN, but not for LTA. Alternatively, TLR2 may form different heterodimers with different TLRs, and CD11c+ imDCs only express the partner that allows recognition for PGN, but not for LTA 41 42. The progressive downregulation of TLRs during pre-DC differentiation into imDCs and mature DCs, as shown here and in previous studies (18, 42), indicates a functional switch of DC lineage from microbial antigen recognition to antigen presentation, thus priming naive T cells and instructing them to differentiate into appropriate effector T cells. Although monocytes have a strong ability to produce TNF-α and IL-6 in response to microbial stimulation, they acquire the ability to produce large amounts of IL-12 and to present antigen to T cells upon differentiation into DCs. While plasmacytoid pre-DCs produce large amounts of IFN-α/β in response to viruses or CpG ODN, they rapidly loss this ability and acquire the ability to present antigen to T cells upon differentiation into DCs.
Recent studies have demonstrated a remarkable functional plasticity of a given DC subset to induce different types of T cell responses depending on the type of invading pathogens 3 8 9 10. However, the expression of limited sets of pattern recognition receptors on monocytes and plasmacytoid pre-DCs suggests that neither monocytes nor plasmacytoid pre-DCs can respond to all microbial antigens, and do not have unrestricted functional plasticity. Therefore, the capacity of DCs to regulate the type of T cell–mediated immune responses may ultimately depend on the environmental instruction that they have received, as well as on their evolutionary lineage heritage.
Acknowledgments
We thank Drs. H. Kanzler, A. O'Garra, and P. Viera for critical reading of the manuscript.
DNAX Research Institute of Molecular and Cellular Biology is supported by Schering-Plough Corporation.
Footnotes
N. Kadowaki's present address is Department of Hematology and Oncology, Graduate School of Medicine, Kyoto University, 54 Shogoin Kawara-cho, Sakyo-ku, Kyoto 606-8507, Japan.
References
- Banchereau J., Steinman R.M. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. doi: 10.1038/32588. [DOI] [PubMed] [Google Scholar]
- Moser M., Murphy K.M. Dendritic cell regulation of TH1-TH2 development. Nat. Immunol. 2000;1:199–205. doi: 10.1038/79734. [DOI] [PubMed] [Google Scholar]
- Kalinski P., Hilkens C.M., Wierenga E.A., Kapsenberg M.L. T-cell priming by type-1 and type-2 polarized dendritic cellsthe concept of a third signal. Immunol. Today. 1999;20:561–567. doi: 10.1016/s0167-5699(99)01547-9. [DOI] [PubMed] [Google Scholar]
- Liu Y.J., Kadowaki N., Rissoan M.C., Soumelis V. T cell activation and polarization by DC1 and DC2. Curr. Top. Microbiol. Immunol. 2000;251:149–159. doi: 10.1007/978-3-642-57276-0_19. [DOI] [PubMed] [Google Scholar]
- Maldonado-Lopez R., De Smedt T., Michel P., Godfroid J., Pajak B., Heirman C., Thielemans K., Leo O., Urbain J., Moser M. CD8alpha+ and CD8alpha− subclasses of dendritic cells direct the development of distinct T helper cells in vivo. J. Exp. Med. 1999;189:587–592. doi: 10.1084/jem.189.3.587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pulendran B., Smith J.L., Caspary G., Brasel K., Pettit D., Maraskovsky E., Maliszewski C.R. Distinct dendritic cell subsets differentially regulate the class of immune response in vivo. Proc. Natl. Acad. Sci. USA. 1999;96:1036–1041. doi: 10.1073/pnas.96.3.1036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rissoan M.-C., Soumelis V., Kadowaki N., Grouard G., Briere F., de Waal Malefyt R., Liu Y.-J. Reciprocal control of T helper cell and dendritic cell differentiation. Science. 1999;283:1183–1186. doi: 10.1126/science.283.5405.1183. [DOI] [PubMed] [Google Scholar]
- d'Ostiani C.F., Del Sero G., Bacci A., Montagnoli C., Spreca A., Mencacci A., Ricciardi-Castagnoli P., Romani L. Dendritic cells discriminate between yeasts and hyphae of the fungus Candida albicans. Implications for initiation of T helper cell immunity in vitro and in vivo. J. Exp. Med. 2000;191:1661–1674. doi: 10.1084/jem.191.10.1661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whelan M., Harnett M.M., Houston K.M., Patel V., Harnett W., Rigley K.P. A filarial nematode-secreted product signals dendritic cells to acquire a phenotype that drives development of Th2 cells. J. Immunol. 2000;164:6453–6460. doi: 10.4049/jimmunol.164.12.6453. [DOI] [PubMed] [Google Scholar]
- Kadowaki N., Antonenko S., Lau J.Y., Liu Y.J. Natural interferon alpha/beta-producing cells link innate and adaptive immunity. J. Exp. Med. 2000;192:219–226. doi: 10.1084/jem.192.2.219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitmore A.C., Haughton G., Arnold L.W. Phenotype of B cells responding to the thymus-independent type-2 antigen polyvinyl pyrrolidinone. Int. Immunol. 1996;8:533–542. doi: 10.1093/intimm/8.4.533. [DOI] [PubMed] [Google Scholar]
- Constant P., Davodeau F., Peyrat M.A., Poquet Y., Puzo G., Bonneville M., Fournie J.J. Stimulation of human gamma delta T cells by nonpeptidic mycobacterial ligands. Science. 1994;264:267–270. doi: 10.1126/science.8146660. [DOI] [PubMed] [Google Scholar]
- Groh V., Steinle A., Bauer S., Spies T. Recognition of stress-induced MHC molecules by intestinal epithelial gammadelta T cells. Science. 1998;279:1737–1740. doi: 10.1126/science.279.5357.1737. [DOI] [PubMed] [Google Scholar]
- Park S.H., Bendelac A. CD1-restricted T-cell responses and microbial infection. Nature. 2000;406:788–792. doi: 10.1038/35021233. [DOI] [PubMed] [Google Scholar]
- Aderem A., Ulevitch R.J. Toll-like receptors in the induction of the innate immune response. Nature. 2000;406:782–787. doi: 10.1038/35021228. [DOI] [PubMed] [Google Scholar]
- Medzhitov R., Janeway C., Jr. Innate immune recognitionmechanisms and pathways. Immunol. Rev. 2000;173:89–97. doi: 10.1034/j.1600-065x.2000.917309.x. [DOI] [PubMed] [Google Scholar]
- Kaisho T., Akira S. Dendritic-cell function in Toll-like receptor- and MyD88-knockout mice. Trends Immunol. 2001;22:78–83. doi: 10.1016/s1471-4906(00)01811-1. [DOI] [PubMed] [Google Scholar]
- Thoma-Uszynski S., Kiertscher S.M., Ochoa M.T., Bouis D.A., Norgard M.V., Miyake K., Godowski P.J., Roth M.D., Modlin R.L. Activation of toll-like receptor 2 on human dendritic cells triggers induction of IL-12, but not IL-10. J. Immunol. 2000;165:3804–3810. doi: 10.4049/jimmunol.165.7.3804. [DOI] [PubMed] [Google Scholar]
- Hertz C., Kiertscher S., Godowski P., Bouis D., Norgard M., Roth M., Modlin R. Microbial lipopeptides stimulate dendritic cell maturation via toll-like receptor 2. J. Immunol. 2001;166:2444–2450. doi: 10.4049/jimmunol.166.4.2444. [DOI] [PubMed] [Google Scholar]
- Grouard G., Rissoan M.C., Filgueira L., Durand I., Banchereau J., Liu Y.J. The enigmatic plasmacytoid T cells develop into dendritic cells with interleukin (IL)-3 and CD40-ligand. J. Exp. Med. 1997;185:1101–1111. doi: 10.1084/jem.185.6.1101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartmann G., Krieg A.M. Mechanism and function of a newly identified CpG DNA motif in human primary B cells. J. Immunol. 2000;164:944–953. doi: 10.4049/jimmunol.164.2.944. [DOI] [PubMed] [Google Scholar]
- Yamamoto T., Yamamoto S., Kataoka T., Komuro K., Kohase M., Tokunaga T. Synthetic oligonucleotides with certain palindromes stimulate interferon production of human peripheral blood lymphocytes in vitro. Jpn. J. Cancer Res. 1994;85:775–779. doi: 10.1111/j.1349-7006.1994.tb02947.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chomczynski P., Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate- phenol-chloroform extraction. Anal. Biochem. 1987;162:156–159. doi: 10.1006/abio.1987.9999. [DOI] [PubMed] [Google Scholar]
- Schwandner R., Dziarski R., Wesche H., Rothe M., Kirschning C.J. Peptidoglycan- and lipoteichoic acid-induced cell activation is mediated by toll-like receptor 2. J. Biol. Chem. 1999;274:17406–17409. doi: 10.1074/jbc.274.25.17406. [DOI] [PubMed] [Google Scholar]
- Yoshimura A., Lien E., Ingalls R.R., Tuomanen E., Dziarski R., Golenbock D. Cutting edgerecognition of Gram-positive bacterial cell wall components by the innate immune system occurs via Toll-like receptor 2. J. Immunol. 1999;163:1–5. [PubMed] [Google Scholar]
- Takeuchi O., Hoshino K., Kawai T., Sanjo H., Takada H., Ogawa T., Takeda K., Akira S. Differential roles of TLR2 and TLR4 in recognition of gram-negative and gram-positive bacterial cell wall components. Immunity. 1999;11:443–451. doi: 10.1016/s1074-7613(00)80119-3. [DOI] [PubMed] [Google Scholar]
- Dziarski R., Wang Q., Miyake K., Kirschning C.J., Gupta D. MD-2 enables Toll-like receptor 2 (TLR2)-mediated responses to lipopolysaccharide and enhances TLR2-mediated responses to Gram- positive and Gram-negative bacteria and their cell wall components. J. Immunol. 2001;166:1938–1944. doi: 10.4049/jimmunol.166.3.1938. [DOI] [PubMed] [Google Scholar]
- Hemmi H., Takeuchi O., Kawai T., Kaisho T., Sato S., Sanjo H., Matsumoto M., Hoshino K., Wagner H., Takeda K., Akira S. A Toll-like receptor recognizes bacterial DNA. Nature. 2000;408:740–745. doi: 10.1038/35047123. [DOI] [PubMed] [Google Scholar]
- Modlin R.L. Immunology. A Toll for DNA vaccines. Nature. 2000;408:659–660. doi: 10.1038/35047207. [DOI] [PubMed] [Google Scholar]
- Wagner H. Toll meets bacterial CpG-DNA. Immunity. 2001;14:499–502. doi: 10.1016/s1074-7613(01)00144-3. [DOI] [PubMed] [Google Scholar]
- Ardavin C., Wu L., Li C.L., Shortman K. Thymic dendritic cells and T cells develop simultaneously in the thymus from a common precursor population. Nature. 1993;362:761–763. doi: 10.1038/362761a0. [DOI] [PubMed] [Google Scholar]
- Bjorck P., Kincade P.W. CD19+ pro-B cells can give rise to dendritic cells in vitro. J. Immunol. 1998;161:5795–5799. [PubMed] [Google Scholar]
- Spits H., Couwenberg F., Bakker A.Q., Weijer K., Uittenbogaart C.H. Id2 and Id3 inhibit development of CD34+ stem cells into predendritic cell (pre-DC)2 but not into pre-DC1. Evidence for a lymphoid origin of pre-DC2. J. Exp. Med. 2000;192:1775–1784. doi: 10.1084/jem.192.12.1775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bendriss-Vermare N., Barthelemy C., Durand I., Bruand C., Dezutter-Dambuyant C., Moulian N., Berrih-Aknin S., Caux C., Trinchieri G., Briere F. Human thymus contains IFN-alpha-producing CD11c−, myeloid CD11c+, and mature interdigitating dendritic cells. J. Clin. Invest. 2001;107:835–844. doi: 10.1172/JCI11734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bauer M., Redecke V., Ellwart J.W., Scherer B., Kremer J.P., Wagner H., Lipford G.B. Bacterial cpg-dna triggers activation and maturation of human cd11c−, cd123+ dendritic cells. J. Immunol. 2001;166:5000–5007. doi: 10.4049/jimmunol.166.8.5000. [DOI] [PubMed] [Google Scholar]
- Cella M., Jarrossay D., Facchetti F., Alebardi O., Nakajima H., Lanzavecchia A., Colonna M. Plasmacytoid monocytes migrate to inflamed lymph nodes and produce large amounts of type I interferon. Nat. Med. 1999;5:919–923. doi: 10.1038/11360. [DOI] [PubMed] [Google Scholar]
- Cella M., Facchetti F., Lanzavecchia A., Colonna M. Plasmacytoid dendritic cells activated by influenza virus and CD40-ligand drive a potent TH1 polarization. Nat. Immunol. 2001;5:919–923. doi: 10.1038/79747. [DOI] [PubMed] [Google Scholar]
- Langenkamp A., Messi M., Lanzavecchia A., Sallusto F. Kinetics of dendritic cell activationimpact on priming TH1, TH2 and nonpolarized T cells. Nat. Immunol. 2001;1:311–316. doi: 10.1038/79758. [DOI] [PubMed] [Google Scholar]
- Siegal F.P., Kadowaki N., Shodell M., Fitzgerald-Bocarsly P.A., Shah K., Ho S., Antonenko S., Liu Y.J. The nature of the principal type 1 interferon-producing cells in human blood. Science. 1999;284:1835–1837. doi: 10.1126/science.284.5421.1835. [DOI] [PubMed] [Google Scholar]
- Lehner M.D., Morath S., Michelsen K.S., Schumann R.R., Hartung T. Induction of cross-tolerance by lipopolysaccharide and highly purified lipoteichoic acid via different toll-like receptors independent of paracrine mediators. J. Immunol. 2001;166:5161–5167. doi: 10.4049/jimmunol.166.8.5161. [DOI] [PubMed] [Google Scholar]
- Ozinsky A., Underhill D.M., Fontenot J.D., Hajjar A.M., Smith K.D., Wilson C.B., Schroeder L., Aderem A. The repertoire for pattern recognition of pathogens by the innate immune system is defined by cooperation between toll-like receptors. Proc. Natl. Acad. Sci. USA. 2000;97:13766–13771. doi: 10.1073/pnas.250476497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Visintin A., Mazzoni A., Spitzer J.H., Wyllie D.H., Dower S.K., Segal D.M. Regulation of Toll-like receptors in human monocytes and dendritic cells. J. Immunol. 2001;166:249–255. doi: 10.4049/jimmunol.166.1.249. [DOI] [PubMed] [Google Scholar]