

Abstract
B lymphocytes undergo affinity maturation of their antigen receptors within germinal centers. These anatomical structures develop in secondary lymphoid organs from the clonal expansion of a few antigen-specific founder B cells, whose isolation and characterization are reported here. Human germinal center founder cells express the naive B cell markers surface IgM and IgD as well as the germinal center B cell markers CD10 and CD38. They express low levels of Bcl-2, high levels of Fas, and undergo rapid apoptosis in culture. The smaller nonproliferating sIgM+IgD+CD38+ B cells displayed a lower level of somatic mutation in their immunoglobulin variable region genes compared with the large proliferating ones. Unmutated sIgM+IgD+CD38+ tonsillar B cells may thus represent germinal center founder cells in which the program for apoptotic cell death is triggered before the onset of somatic mutation, allowing the selection of the germline antibody repertoire at an early stage.
The T cell–dependent primary humoral responses are initiated by the activation of sIgM+IgD+ naive B cells in the T cell–rich foci of secondary lymphoid tissues (1–4), allowing the generation of short-lived plasma cells and the recruitment of germinal center (GC)1 founder cells into B cell follicles (1–4). These cells undergo clonal expansion, somatic mutation of their IgV genes (5–11), and antigendriven affinity maturation (12–20). The maturation pathway of peripheral B lymphocytes during T cell–dependent immune response can be traced by changes of surface molecule expression (21). Accordingly, we have previously reported the purification and characterization of five human tonsillar B cell subpopulations: Bm1 and Bm2 are two subsets of follicular mantle B cells which are sIgD+CD38− CD23− and sIgD+CD38−CD23+, respectively; Bm3 and Bm4 represent sIgD−CD38+CD77+ GC centroblasts and sIgD−CD38+CD77− centrocytes, respectively; and Bm5 represents sIgD−CD38− memory B cells (11, 22–25). However, B cells corresponding to the transition stage from naive follicular mantle B cells to GC B cells have not been characterized yet. Recently, we have identified tonsillar B cells that coexpress sIgD and CD38, and that can be further separated into sIgM+ and sIgM− subsets. Sequence analysis of IgV genes shows that the sIgM−IgD+CD38+ subset contains extensively mutated IgV genes, excluding the possibility that they could be GC founder cells (26). Here we present the evidence that the sIgM+IgD+CD38+ subset contains medium sized nonproliferating GC founder cells that acquire the propensity to undergo apoptosis before the onset of somatic mutation.
Materials and Methods
Antibodies.
Antibodies (clone number, isotype, and source) used for phenotyping and immunomagnetic bead depletion are listed in Table 1.
Table 1.
Antibody List
| Anti–human Abs | Clone | Isotype | Source | |||
|---|---|---|---|---|---|---|
| CD 2 (FITC) | 39 C1.5 | IgG2a (rat) | Immunotech (Marseille, France) | |||
| CD 3 (FITC) | UCHT 1 | IgG1 | Immunotech | |||
| CD 10-FITC | W8 E37 | IgG2a | Becton Dickinson (Mountain View, CA) | |||
| CD 14-FITC | RMO 52 | IgG2a | Immunotech | |||
| CD 19-FITC | J4-119 | IgG1 | Immunotech | |||
| CD 20-FITC | IOB 20b | IgG1 | Immunotech | |||
| CD 23-PE | B-G6 | IgG1 | Serotec Ltd. (Oxford, UK) | |||
| CD 38 | T 10 | IgG1 | Immunotech | |||
| CD 38-PE | HB-7 | IgG1 | Becton Dickinson | |||
| CD 39 (Biot.) | Ac-2 | IgG1 | The Binding Site (Birmingham, UK) | |||
| CD 44-FITC | NaM 10-8F4 | IgG1 | Diagast (Lille, France) | |||
| CD 71-FITC | IOA 71 | IgG1 | Immunotech | |||
| CD 77 sup. | 38-13 | IgM (rat) | Immunotech | |||
| Fas-FITC | UB 2 | IgG1 | Immunotech | |||
| FAS | CH 11 | IgM | Immunotech | |||
| Bcl2-FITC | IgG1 | DAKO (Glostrup, Denmark) | ||||
| Ki67-FITC | IgG1 | DAKO | ||||
| IgM-FITC | F(ab′)2 (rabbit) | DAKO | ||||
| Ig D Biot. | IgG (goat) | Amersham Intl. (Buckinghamshire, UK) | ||||
| Ig A pur. | IgG1 | Serotec Ltd. | ||||
| Ig G pur. | IgG1 | Serotec Ltd. | ||||
| Secondary Abs | ||||||
| Goat anti–mouse-FITC | IgG + IgM | Bioart (Neudon, France) | ||||
| Goat anti–rat-FITC | IgM | Nordic Immunological Labs. (Tilburg, The Netherlands) | ||||
| Streptavidin-PE | Becton Dickinson | |||||
| Streptavidin-FITC | Immunotech | |||||
| Streptavidin-Tricolor | CALTAG Labs. (South San Francisco, CA) | |||||
| Ig isotype controls | ||||||
| Mouse IgG1 Biot. | CALTAG Labs. | |||||
| Mouse IgM | Immunotech | |||||
| Rat IgG2a FITC | Immunotech | |||||
| Mouse Ig G1 FITC/PE | Immunotech | |||||
| Mouse Ig G2A FITC/PE | Immunotech | |||||
| Immunochemistry reagents | ||||||
| Ig D Biot. | IgG (goat) | Amersham Intl. | ||||
| IgM | 145-8 | IgGl | Becton Dickinson | |||
| Streptavidin-peroxydase | DAKO | |||||
| APAAP | DAKO |
Isolation of Tonsillar B Cells.
Tonsillar B cells were prepared as previously described (25). Briefly, tonsils taken from patients during routine tonsillectomy were finely minced and the resulting cell suspension was subjected to two rounds of depletion of non-B cells: (a) T cells were depleted by rosetting with sheep red blood cells, (b) residual non-B cells were depleted by T cell specific antibodies (CD2, CD3, and CD4), and then by magnetic beads coupled with anti–mouse IgG (Dynabeads; Dynal, Oslo, Norway). The resulting cells from all the experiments contained >98% CD19 positive B cells.
Phenotype Analysis of the Four B Cell Subsets Defined by the Expression of sIgD and sCD38 by Three-color Immunofluorescence Flow Cytometry.
Total tonsillar B cells were incubated with mouse anti–human CD38-PE, goat anti–human IgD-biotin, and a set of FITC-conjugated mouse anti–human IgM, CD23, CD44, CD10, CD71, CD77, and Fas/CD95 for 20 min at 4°C. After washing twice with PBS containing 2% BSA, cells were incubated with streptavidin-tricolor for 30 min and analyzed with a FACScan® flow cytometer. For intracellular Bcl-2 and nuclear Ki67 staining, cells were permeabilized by incubation with 3 g/100 ml saponin for 15 min at 4°C.
Separation of IgD+CD38+ Tonsillar B Cells into IgM+ and IgM−Subsets by Three-color Immunofluorescence FACS® Sorting.
Total tonsillar B cells were incubated with mouse anti–human CD38-PE and goat anti–human IgD-biotin for 20 min at 4°C. After washing twice with PBS containing 2% BSA, the cells were incubated with mouse anti–human IgM-FITC and streptavidin-tricolor for 20 min. Then cells were washed twice and suspended in PBS at a concentration of 3 × 106/ml. IgD+CD38+ B cells were separated into two subsets according to the expression of sIgM on a FACStar plus®. IgM+IgD+CD38+ B cells from one tonsil were further size fractionated according to their forward scatter parameters.
Giemsa Staining and Analysis of Nuclear Antigen Ki67.
105 cells from each of the purified B cell subsets were cytocentrifuged for 5 min at 500 rpm on a microscope slide. Slides were fixed in methanol for 5 min and then stained with Giemsa staining solution (BDH Chemicals Ltd., Poole, England) diluted one in five with distilled water. Some slides were fixed in cold acetone at 4°C for 10 min for immunocytology. Slides were washed in PBS for 5 min and incubated with mouse mAb against Ki67 antigen (DAKO, Glostrup, Denmark) for 45 min. The slides were washed twice in PBS and then incubated with sheep anti–mouse Ig (The Binding Site, Birmingham, England). After 45 min, slides were washed and incubated with mouse mAb against alkaline phosphatase and alkaline phosphatase complexes (APAAP) (DAKO). After an additional 45 min, the slides were washed three times in PBS and the enzyme activity was developed by the FAST RED substrate (DAKO).
Cell Cultures.
Cells were cultured in RPMI 1640 medium containing 10% heat inactivated fetal calf serum, 80 μg/ml gentamicin, and 2 mM glutamine (all from Flow Laboratories, Inc., MacLean, VA) at 37°C. Cells (2.5 × 105/ml) were cultured for 5 d in one of the following conditions: IL-2 (10 U/ml), IL-4 (50 U/ml), or IL-10 (100 ng/ml). 2.5 × 104 CD40-ligand transfected L cells and 2.5 × 103 human fibroblasts from rheumatoid synovium (irradiated with 75Gy) were used for the cultures. DNA synthesis was assessed by an 8 h pulse with 1 μCi [3H]TdR before cell harvesting.
Immunohistology.
Portions of tonsils were snap frozen in liquid nitrogen and stored at −70°C. 5 μm frozen sections were cut and mounted on glass slides. They were thoroughly dried at room temperature for ≥1 h and were fixed in acetone at 4°C for 15 min. Sections were stained by double immunoenzyme technique using biotin-avidin-peroxidase system and alkaline phosphataseanti-alkaline phosphatase system (APAAP technique). Briefly, sections were washed in PBS for 5 min. Then sections were incubated with goat anti–human IgD-biotin and mouse anti–human IgM (IgG1 isotype). After washing for 5 min in PBS, the sections were incubated with streptavidine-peroxidase and sheep anti– mouse IgG1 for 30 min, and then incubated with alkaline phosphatase coupled to mouse antibodies specific for alkaline phosphatase (APAAP complexes). After a final wash, peroxidase was developed by 3-amino-9-ethylcarbazole which gives a red color, and alkaline phosphatase was developed by Fast blue substrate which gives a blue color (27).
Analysis of the VH5 Transcripts PCR Amplified from the Human B Cell Subsets.
mRNA was extracted from 25 × 103 B cells (11). cDNA was obtained by reverse transcription using the Superscript Reverse Transcriptase Kit (GIBCO BRL, Gaithersburg, MD), with oligo dT12 –18 primers (Pharmacia, Upsalla, Sweden). Full length VH5 transcripts were amplified with L-VH5 primer (5′CCCGAATTCATGGGGTCAACCGCCATCCT3′) with 3′ primer CHμ (TGGGGCGGATGCACTCCC) with Taq polymerase (Perkin-Elmer Corp., Norwalk, CT) using the reaction buffer provided by the manufacturers and a DNA thermal cycler (Perkin-Elmer Corp.) with 35 cycles of 1 min denaturation at 94°C, 2 min of primer annealing at 60°C, and 3 min extension at 72°C. After the last cycle, the reaction mixtures were incubated for 10 min at 72°C to ensure complete extension of all products. The PCR products were cloned in PCR™II vector, using the TA Cloning Kit (Invitrogen, San Diego, CA). Both DNA strands of plasmids extracted from individual bacterial colonies were sequenced on an automated DNA sequencer (Applied Biosystems Inc., Foster City, CA). Sequencing was done with the −21M13 and M13RP primers flanking the plasmid cloning sites, and with the CHμ primer annealing with the 3′ end of CH1-μ.
Results
Three-color Flow Cytometry Identifies IgM+IgD+CD38+ Tonsillar B Cells.
Human tonsillar B cells that coexpress sIgD and CD38 have been identified, which represent 5–15% of total human tonsillar B cells (Fig. 1 A). Three-color flow cytometry shows that sIgD+CD38+ B cells express low levels of the naive B cell markers CD23, CD44, and IgM, while they express high levels of the germinal center markers CD71, CD10, and CD77 (Fig. 1 B). Unlike the sIgD+CD38− naive B cells, they express low levels of Bcl-2, high levels of Fas, and many of them express the proliferation associated nuclear antigen Ki67 (Fig. 1 B). The sIgD+CD38+ B cells were sorted into a sIgM+ subset (3–9% of total tonsillar B cells) and a sIgM− subset (2–6% of total tonsillar B cells) (Fig. 1 C). Our recent study has shown that sIgM−IgD+CD38+ B cells represent a peculiar population of highly mutated GC centroblasts (28). We have now further characterized the sIgM+IgD+CD38+ B cell subset which displays a phenotype intermediate between follicular mantle naive B cells and GC B cells.
Figure 1.

Subpopulations of tonsillar B cells. (A) Double staining of total tonsil B cells with anti–IgD-PE and anti–CD38-FITC, showing four B cell subsets. The sIgD+CD38+ B cells represent 5–15% of total tonsil B cells from three representative samples. (B) Phenotypic analysis by threecolor flow cytometry. Anti–IgD-tricolor and anti–CD38-PE are used together with other third antibodies directly conjugated with FITC. Anti– IgD-PE and anti–CD38-FITC were used together with Hoechst 33342. For Bcl-2 and Ki67 staining, the cells were permeabilized by saponin (0.33 g/100 ml in PBS, 1% BSA) for 15 min on ice. (C) Cell sorting of sIgD+CD38+ B cells into sIgM+ and sIgM− subsets.
sIgM+IgD+CD38+ B Cells Display GC B Cell Morphology, Propensity for Apoptosis, and Reduced Proliferation Capacity In Vitro.
Unlike small dense sIgM+sIgD+CD38− naive B cells, sIgM+IgD+CD38+ B cells are large- or medium-sized lymphocytes, similar to IgD−CD38+ GC B cells (12, 23). Large IgM+IgD+CD38+ B cells, which account for 20– 30% of the IgM+IgD+CD38+ B cells, express the proliferation associated nuclear antigen Ki67, while medium-sized IgM+IgD+CD38+ B cells are Ki67− (Fig. 2 B). Like IgD−CD38+ GC B cells, >90% of IgM+IgD+CD38+ B cells display apoptotic figures after 16 h of culture (Fig. 2 C and Table 2). The survival and proliferation of IgM+ IgD+CD38+ B cells in vitro depend on the presence of CD40-ligand and T cell cytokines such as IL-2, -4, and -10 (Table 3). The level of DNA synthesis observed under identical culture conditions is always lower in IgM+IgD+CD38+ B cells than in IgD+CD38− naive B cells, but higher than in IgD−CD38+ GC B cells (Table 3).
Figure 2.
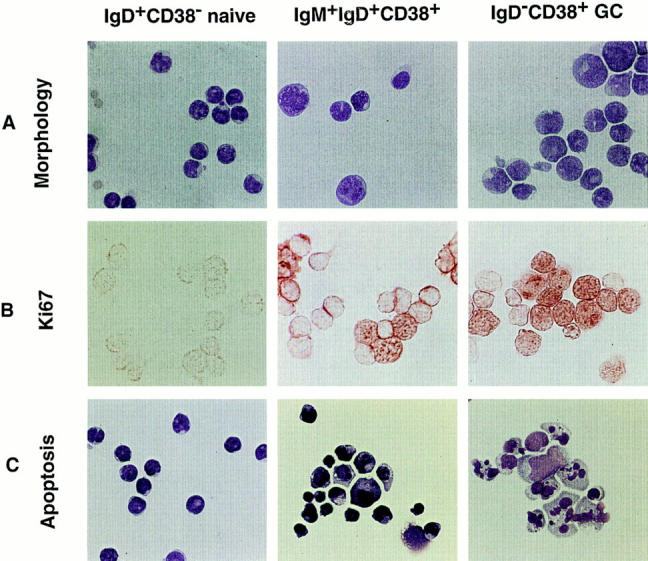
Cytological characterization of slgM+IgD+CD38− naive, slgD−CD38+ GC, and slgM+IgD+CD38+ B cells. (A) Giemsa staining of freshly isolated three B cell subsets (×1,000). (B) Expression of proliferation associated nuclear antigen Ki67 on freshly isolated B cell subsets (×1,000). (C) Giemsa staining of three B cell subsets after a 16 h culture, showing apoptotic figures in slgM+IgD+CD38+ GC founder cells and in slgD−CD38+ GC B cells. The quantitation of apoptotic cells is shown in Table 2.
Table 2.
Percentage of Apoptotic Cells in B Cell Subsets After a 16 h Culture
| sIgD+CD38− Naive B | sIgD−CD38+ GC B | sIgM+IgD+CD38+ GC founder | ||||
|---|---|---|---|---|---|---|
| Experiment 1 | 22 | 80 | 98 | |||
| Experiment 2 | 8 | 78 | 92 |
Table 3.
DNA Synthesis in Three B Cell Subsets at Day 5 of Culture
| CD 40-ligand | CD40-ligand IL-2 + IL-10 | CD40-ligand IL-4 + IL-10 | ||||
|---|---|---|---|---|---|---|
| sIgD+CD38− | 18,550 ± 1,590 | 32,870 ± 2,140 | 25,900 ± 2,830 | |||
| sIgM+IgD+ | ||||||
| CD38+ | 570 ± 160 | 8,610 ± 160 | 18,350 ± 2,720 | |||
| sIgD−CD38+ | 1,340 ± 330 | 8,270 ± 730 | 6,220 ± 460 |
[3H]Thymidine uptake is expressed as cpm ± SD. It represents one of six identical experiments.
Somatic Mutation Has Occurred in a Fraction of sIgM+IgD+CD38+ B Cells.
VH5-μ sequence analysis has been successfully used to trace the progression of human tonsillar B cell subsets from the virgin to the memory compartment (11). Therefore, 32 VH5-μ sequences from sIgM+IgD+ CD38+ B cells were compared to 30 VH5-μ sequences from sIgM+IgD+CD38− naive B cells of three tonsil samples (Fig. 3, A and B). In agreement with our previous reports, all 30 sequences from naive B cells contain 0–2 mutations per sequence, with an overall mutation frequency of 2 × 10−3 bp (20 mutations/9,600 bp) that is barely distinguishable from the PCR Taq-error rate (1 × 10−3 bp). Within 32 sequences from sIgM+IgD+CD38+ B cells, while 17 might be considered germline (0–2 mutations/sequence), 15 have accumulated from 3–13 mutations, with an average mutation frequency of 20 × 10−3 bp (88 mutations/4,480 bp). The replacement mutations in 12 less mutated sequences (3.2, 3.4, 3.10, 3.14, 3b.5, 3b.6, 3b.9, 3b.13, 3c.5, 3c.7, 3c.9, and 3c.12) are randomly distributed within the CDRs and FWs, an indication of the absence of antigen-driven selection. In contrast, sequences 3.11, 3b.7, and 3c.6, which have accumulated 10–13 mutations, showed replacement mutations mainly focused in the CDRs, suggesting that these cells have undergone antigen-driven selection. Unlike the VH5-γ sequences from sIgD−CD38+ GC B cells and VH5-δ sequences from sIgM−IgD+CD38+ reported previously (11, 28), no clonal relatedness could be found among the VH5-μ transcripts from tonsillar sIgM+ IgD+CD38+ B cells. To establish whether the mediumsized nonproliferating cells from this population are those with unmutated V regions, sIgM+IgD+CD38+ B cells were isolated from a fourth tonsil and fractionated according to their size. The subset of medium-sized sIgM+IgD+CD38+ B cells was enriched for unmutated B cells (Fig. 3, C and D) as five out of nine sequences from the medium-sized B cells displayed less than two mutations and were therefore considered as nonmutated. In contrast, only one out of nine sequences from the large B cells was unmutated. In those two subsets, the most mutated sequences show features of antigen-driven selection.
Figure 3.


Schematic representation of mutations in rearranged VH5-μ sequences. Each line represents one VH5 sequence. VH5-2 sequences are underlined. Leader region, CDR1, and CDR2 are indicated. Mutations present in the DJH regions are not shown. Mutations are represented by different symbols. •, replacement mutation; ‖, silent mutation; ○, stop codon. (A and B) VH5-μ sequences from slgM+IgD+CD38− B cells and sIgM+IgD+CD38+ B cells, respectively. The upper, middle, and lower groups of sequences represent three different tonsil samples. (C and D) VH5-μ sequences from medium-sized and large sIgM+IgD+CD38+ B cells, respectively, that were isolated from the fourth tonsil.
IgM+IgD+ B Cells Can Be Found Within GC.
It has been previously reported that a small proportion of human GC contain IgD+ B cells (29, 30). Indeed, by double immunohistological staining, we have recently confirmed these observations and further shown that IgD+ B cells within GC display CD38, the majority of them expressing the proliferation associated nuclear antigen Ki67 (28). To distinguish IgM+IgD+CD38+ GC founder cells from highly mutated IgM−IgD+CD38+ GC B cells in situ (26), double staining with anti-IgM and anti-IgD were performed on tonsil sections. While numerous large IgM−IgD+ B blasts were found to be packed within occasional GC dark zones (26), only scattered IgM+IgD+ B cells were found throughout a few GCs (Fig. 4).
Figure 4.
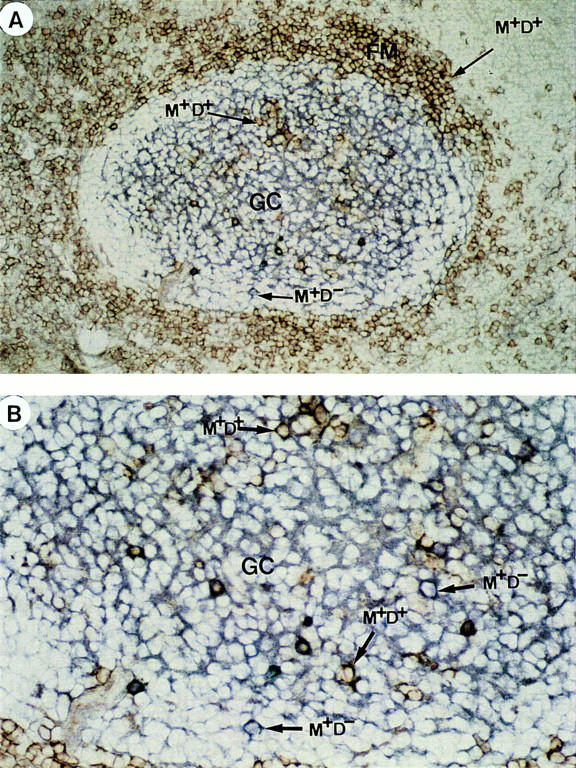
Identification of IgM+IgD+ and IgM+IgD− B cells within GC: Double anti– IgD (red) and anti–IgM (blue) staining of a tonsil section. (A) shows a secondary lymphoid follicle that contains purple IgD+IgM+ follicular mantle B cells. Within the GC, IgM immune complexes (blue) on follicular dendritic cells can be observed. In addition, scattered purple IgM+IgD+ B cells and blue IgM+IgD− B cells can be found (×100). (B) Shows a part of this GC at ×200 magnification.
Discussion
During T cell–dependent primary immune responses, antigen-specific naive B lymphocytes undergo either plasma cell reaction in the T cell zones or enter into the B cell follicles to initiate GC reaction. One of the questions regarding GC development has been the identity of GC precursor cells (16, 31, 32). Progress has been made by analyzing the capacity of B cell subsets to form GC in the host animals after adoptive cell transfer and immunization. SCID mice immunized after repopulation with carrier-primed CD4+ T cells and J11Dlow splenic B cells developed many GC in their spleens. In contrast, when J11Dhigh cells or CD5+ peritoneal B cells were transferred, few, if any, GC were generated (33). However, thoracic duct B cell transfer experiments in rats have generated contradictory conclusions as to whether GC precursor cells are sIgD+ or sIgD− (34, 35). The present identification of tonsillar sIgM+IgD+ CD38+ B cells represents a parallel attempt in the human system to characterize GC founder cells.
The formal identification of GC founder cells would require the demonstration of a precursor–progeny relationship between such cells and GC mutated B cells. However, we could not find clonally related unmutated sIgM+sIgD+ CD38+ B cells and mutated sIgD−CD38+ B cells from the same tonsil within a limited number of sequences (data not shown). In agreement with our results, when GC B cells were individually picked from the same GC on human lymph node tissue sections, no clonal relatedness could be observed between the germline VH/Vκ sequences and the mutated sequences (8). It suggests that it might be difficult to directly illustrate such a precursor-progeny relationship in humans, since kinetic analysis is not easy to perform. However, several important features suggest that mediumsized nonproliferating sIgM+IgD+CD38+ B cells might be enriched for GC founder cells. First, these sIgM+IgD+ B cells coexpress, albeit at a reduced level, markers of naive B cells such as CD23 and CD44, as well as markers of bona fide GC B cells such as CD10, CD38, and CD71. As sIgD+ CD38+ B cells are found within GC exclusively, their intermediate phenotype is suggestive of a transitional stage of maturation between follicular mantle naive B cells and early GC B cells. Second, 29 out of 32 IgVH clonally independent sequences are either in germline configurations (17 sequences) or are poorly mutated (12 sequences) and show no clear evidence for antigen-driven selection. Moreover, when sorted according to their size, >50% of the medium-sized sIgM+IgD+CD38+ B cells are germline or low mutated, a characteristic expected for early GC B cells. Third, in contrast to typical GC centroblasts which give rise, after clonal expansion, to centrocytes, medium-sized sIgM+IgD+CD38+ B cells are not actively dividing as they are Ki67−. This suggests that these cells have not yet undergone clonal expansion, a conclusion that is supported by the lack of clonal relatedness between the IgVH sequences of sIgM+IgD+CD38+ B cells isolated from the same tonsil. In conclusion, medium-sized, nonproliferating sIgM+IgD+ CD38+ B cells are enriched for early GC B cells.
An important finding of the present study is the early triggering of apoptosis program in sIgM+IgD+CD38+ GC founder cells, which happens before the onset of somatic mutation. This early propensity to undergo apoptosis may provide a mechanism for selection of cells bearing high affinity unmutated antigen receptors within GC. Consequently, only the cells bearing antigen receptors with better affinity will have the opportunity to undergo affinity maturation. This hypothesis is supported by the finding that many PNA+ mouse GC B cells, appearing during the first 10 d of primary response to NP (4-hydroxy-3-nitrophenyl) contained selected IgV genes in germ line or low mutated configurations (7, 36). In addition, this early propensity to undergo apoptosis may also allow the immediate selection of mutating cells (37). Consequently, the B cells entering into GC have to mutate rapidly and efficiently in order to survive.
The demonstration of sIgD on naive B cell derived GC founder cells supports the hypothesis of Thorbecke et al. (16) and Roes and Rajewsky (38) suggesting an auxiliary receptor function for sIgD in antigen-mediated recruitment of B cells into GC reaction and memory B cell formation. This hypothesis was based on the following observations. (a) sIgD are expressed on naive B cells 10 times more efficiently than slgM (39) and bind antigen better than sIgM due to their structural flexibility (40). These two characteristics of sIgD may help naive B cells to colonize the network of follicular dendritic cells when antigen become limiting during GC development (38). (b) A subpopulation of helper T cells bearing receptors for sIgD was found to play an important role in enhancing T cell–dependent humoral immune responses (41, 42). (c) In vivo injection of anti– mouse IgD dramatically increases, within 6 d, the volume of GC in the spleen (43). (d) IgD-knock out mice displayed a delayed affinity maturation of T cell–dependent antibody responses (38) and a higher sensitivity to tolerance induction (44).
The present demonstration of sIgD on GC founder cells also sets a limit to the concept of sIgD being an absolute marker for naive resting B cells. This concept was essentially based on the rapid loss of sIgD on naive B cells after activation in vitro and in vivo (45, 46), and the apparent lack of IgD expression in GC (21). However, consistent with our present identification of sIgD+ proliferating GC founder cells in vivo, sIgD+ proliferating B cells were observed in long-term culture of CD40 activated human naive B cells (47). Furthermore, the three VH-5μ sequences with more than 10 mutations showed evidence for antigendriven selection. These three mutated sIgM+IgD+CD38+ B cells may either represent GC founder cells derived from recirculating sIgM+IgD+ memory B cells or correspond to GC centrocytes currently differentiating into sIgM+IgD+ memory B cells. Therefore, our present findings further support the existence of sIgD+ memory B cells, demonstrated in mouse adoptive transfer experiments a decade ago (48, 49).
In conclusion, human GC founder cells have been isolated as medium-sized, nonproliferating sIgM+IgD+CD38+ B cells. Further study of this subpopulation of B cells should lead to better understanding of the early events governing GC development.
Acknowledgments
We thank Drs. F. Brière and P. Garrone for critical reading of the manuscript, Dr. J. Chiller for support, Mrs. Bonnet-Arnaud and Mrs. Vatan for editorial assistance, and Ms. I. Durand for cell sorting.
Footnotes
C. Arpin is recipient of a grant from Fondation Narcel Nérieux (Lyon, France).
1 Abbreviations used in this paper: APAAP, alkaline phosphatase-anti-alkaline phosphatase system; GC, germinal center.
References
- 1.Liu Y-J, Zhang J, Lane PJL, Chan EY-T, MacLennan ICM. Sites of specific B cell activation in primary and secondary responses to T cell–dependent and T cell– independent antigens. Eur J Immunol. 1991;21:2951–2962. doi: 10.1002/eji.1830211209. [DOI] [PubMed] [Google Scholar]
- 2.Jacob J, Kassir R, Kelsoe G. In situ studies of the primary immune response to (4-hydroxy-3-nitrophenyl) acetyl. I. The architecture and dynamics of responding cell populations. J Exp Med. 1991;173:1165–1175. doi: 10.1084/jem.173.5.1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kroese FGM, Wubbena AS, Seijen HG, Nieuwenhuis P. Germinal centers develop oligoclonally. Eur J Immunol. 1987;17:1069–1072. doi: 10.1002/eji.1830170726. [DOI] [PubMed] [Google Scholar]
- 4.Smith KGC, Hewitson TD, Nossal GJV, Tarlinton DM. The phenotype and fate of the antibodyforming cells of the splenic foci. Eur J Immunol. 1996;26:444–448. doi: 10.1002/eji.1830260226. [DOI] [PubMed] [Google Scholar]
- 5.Jacob J, Kelsoe G, Rajewsky K, Weiss U. Intraclonal generation of antibody mutants in germinal centres. Nature (Lond) 1991;354:389–392. doi: 10.1038/354389a0. [DOI] [PubMed] [Google Scholar]
- 6.Berek C, Berger A, Apel M. Maturation of the immune response in germinal centers. Cell. 1991;67:1121–1129. doi: 10.1016/0092-8674(91)90289-b. [DOI] [PubMed] [Google Scholar]
- 7.McHeyzer-Williams MG, McLean MJ, Lalor PA, Nossal GJV. Antigen-driven B cell differentiation in vivo. J Exp Med. 1993;178:295–307. doi: 10.1084/jem.178.1.295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Küppers R, Zhao M, Hansmann M-L, Rajewsky K. Tracing B cell development in human germinal centres by molecular analysis of single cells picked from histological sections. EMBO (Eur Mol Biol Organ) J. 1993;12:4955–4967. doi: 10.1002/j.1460-2075.1993.tb06189.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Leanderson T, Källberg E, Gray D. Expansion, selection and mutation of antigen-specific B cells in germinal centers. Immunol Rev. 1992;126:47–61. doi: 10.1111/j.1600-065x.1992.tb00630.x. [DOI] [PubMed] [Google Scholar]
- 10.Klein U, Küppers R, Rajewsky K. Variable region gene analysis of B cell subsets derived from a 4-year-old child: somatically mutated memory B cells accumulate in the peripheral blood already at young age. J Exp Med. 1994;180:1383–1393. doi: 10.1084/jem.180.4.1383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pascual V, Liu YJ, Magalski A, de Bouteiller O, Banchereau J, Capra JD. Analysis of somatic mutation in five B cell subsets of human tonsil. J Exp Med. 1994;180:329–339. doi: 10.1084/jem.180.1.329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Liu YJ, Joshua DE, Williams GT, Smith CA, Gordon J, MacLennan ICM. Mechanisms of antigendriven selection in germinal centers. Nature (Lond) 1989;342:929–931. doi: 10.1038/342929a0. [DOI] [PubMed] [Google Scholar]
- 13.MacLennan ICM, Gray D. Antigen-driven selection of virgin and memory B cells. Immunol Rev. 1986;91:61–85. doi: 10.1111/j.1600-065x.1986.tb01484.x. [DOI] [PubMed] [Google Scholar]
- 14.MacLennan ICM. Germinal centers. Annu Rev Immunol. 1994;12:117–139. doi: 10.1146/annurev.iy.12.040194.001001. [DOI] [PubMed] [Google Scholar]
- 15.Nossal GJV. The molecular and cellular basis of affinity maturation in the antibody response. Cell. 1992;68:1–3. doi: 10.1016/0092-8674(92)90198-l. [DOI] [PubMed] [Google Scholar]
- 16.Thorbecke GJ, Amin AR, Tsiagbe VK. Biology of germinal centers in lymphoid tissues. FASEB J. 1994;8:832–840. doi: 10.1096/fasebj.8.11.8070632. [DOI] [PubMed] [Google Scholar]
- 17.Banchereau J, Rousset F. Human B lymphocytes: phenotype, proliferation and differentiation. Adv Immunol. 1992;52:125–251. doi: 10.1016/s0065-2776(08)60876-7. [DOI] [PubMed] [Google Scholar]
- 18.Clark EA, Ledbetter JA. How B and T cells talk to each other. Nature (Lond) 1994;367:425–428. doi: 10.1038/367425a0. [DOI] [PubMed] [Google Scholar]
- 19.Weissman IL. Developmental switches in the immune system. Cell. 1994;76:207–218. doi: 10.1016/0092-8674(94)90329-8. [DOI] [PubMed] [Google Scholar]
- 20.Rajewsky, K. 1989. Evolutionary and somatic immunological memory. In Progress in Immunology. VII. F. Melchers, editor. Springer-Verlag, Berlin. 397–403.
- 21.Liu YJ, Johnson GD, Gordon J, MacLennan ICM. Germinal centers in T-cell–dependent antibody responses. Immunol Today. 1992;13:17–21. doi: 10.1016/0167-5699(92)90199-H. [DOI] [PubMed] [Google Scholar]
- 22.Liu, Y.J., C. Barthélémy, O. de Bouteiller, and J. Banchereau. 1994. The differences in survival and phenotype between centroblasts and centrocytes. In In Vivo Immunology. E. Heinen, editor. Plenum Press, New York. 213–218. [DOI] [PubMed]
- 23.Liu YJ, Barthélémy C, de Bouteiller O, Arpin C, Durand I, Banchereau J. Memory B cells from human tonsils colonize mucosal epithelium and directly present antigen to T cells by rapid upregulation of B7.1 and B7.2. Immunity. 1995;2:238–248. doi: 10.1016/1074-7613(95)90048-9. [DOI] [PubMed] [Google Scholar]
- 24.Martinez-Valdez H, Guret C, de Bouteiller O, Fugier I, Banchereau J, Liu YJ. Human germinal center B cells express the apoptosis inducing genes Fas, c-myc, p53, and Bax but not the survival gene blc-2. J Exp Med. 1996;183:971–977. doi: 10.1084/jem.183.3.971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Liu, Y.J., and J. Banchereau. 1996. Human peripheral B cell subsets. In Handbook of Experimental Immunology. D. Weir, C. Blackwell, and L. Hersenberg, editors. Blackwell Scientific Publ., Oxford. In press.
- 26.Liu YJ, Malisan F, de Bouteiller O, Guret C, Lebecque S, Banchereau J, Mills FC, Max EE, Martinez-Valdez H. Within germinal centers isotype switching of immunoglobulin genes occurs after onset of somatic mutation. Immunity. 1996;4:241–250. doi: 10.1016/s1074-7613(00)80432-x. [DOI] [PubMed] [Google Scholar]
- 27.Liu YJ, Oldfield S, MacLennan ICM. Memory B cells in T cell–dependent antibody responses colonize the splenic marginal zones. Eur J Immunol. 1988;18:355–362. doi: 10.1002/eji.1830180306. [DOI] [PubMed] [Google Scholar]
- 28.Liu YJ, de Bouteiller O, Arpin C, Brière F, Galibert L, Ho S, Martinez-Valdez H, Banchereau J, Lebecque S. Normal human IgD+IgM−germinal center B cells can express up to 80 mutations in the variable region of their IgD transcripts. Immunity. 1996;4:603–613. doi: 10.1016/s1074-7613(00)80486-0. [DOI] [PubMed] [Google Scholar]
- 29.Hsu SM, Jaffe ES. Phenotypic expression of B-lymphocytes. 2. Immunoglobulin expression of germinal center cells. Am J Pathol. 1994;114:396–402. [PMC free article] [PubMed] [Google Scholar]
- 30.Nahm MH, Takes PA, Bowen MB, Macke KA. Subpopulations of B lymphocytes in germinal centers. II. A germinal center B cell subpopulation expresses sIgD and CD23. Immunol Lett. 1989;21:201–208. doi: 10.1016/0165-2478(89)90105-3. [DOI] [PubMed] [Google Scholar]
- 31.Thorbecke GJ, Flotte TJ, Baine Y. Maturity of precursor cells for germinal centers. Adv Exp Med Biol. 1982;149:845–847. doi: 10.1007/978-1-4684-9066-4_117. [DOI] [PubMed] [Google Scholar]
- 32.Nieuwenhuis P, Opstelten D. Functional anatomy of germinal centres. Am J Anat. 1984;170:421. doi: 10.1002/aja.1001700315. [DOI] [PubMed] [Google Scholar]
- 33.Linton PJ, Lo D, Lai L, Thorbecke GJ, Klinman NR. Among naive precursor cell subpopulations only progenitors of memory B cells originate germinal centers. Eur J Immunol. 1992;22:1293–1297. doi: 10.1002/eji.1830220526. [DOI] [PubMed] [Google Scholar]
- 34.Seijen HG, Bun JCAM, Wubbena AS, Loehlefink KGL. The germinal center precursor cell is surface IgM and IgD positive. Adv Exp Med Biol. 1988;237:233–237. doi: 10.1007/978-1-4684-5535-9_35. [DOI] [PubMed] [Google Scholar]
- 35.Vonderheide RH, Hunt SV. Comparison of IgD+ and IgD−thoracic duct B lymphocytes as germinal center precursor cells in the rat. Int Immunol. 1991;3:1273–1281. doi: 10.1093/intimm/3.12.1273. [DOI] [PubMed] [Google Scholar]
- 36.Jacob J, Przylepa J, Miller C, Kelsoe G. In situ studies of the primary immune response to (4-hydroxyl3-nitrophenyl) acetul. III. The kinetics of V region mutation and selection in germinal center B cells. J Exp Med. 1993;178:1293–1307. doi: 10.1084/jem.178.4.1293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Weiss U, Zoebelein R, Rajewsky K. Accumulation of somatic mutants in the B cell compartment after primary immunization with a T cell–dependent antigen. Eur J Immunol. 1992;22:511–517. doi: 10.1002/eji.1830220233. [DOI] [PubMed] [Google Scholar]
- 38.Roes J, Rajewsky K. Immunoglobulin D (IgD)– deficient mice reveal an auxiliary receptor function for IgD in antigen-mediated recruitment of B cells. J Exp Med. 1993;177:45–55. doi: 10.1084/jem.177.1.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Brink R, Goodnow CC, Basten A. IgD expression on B cells is more efficient than IgM but both receptors are functionally equivalent in up-regulation CD80/CD86 co-stimulatory molecules. Eur J Immunol. 1995;25:1980–1984. doi: 10.1002/eji.1830250727. [DOI] [PubMed] [Google Scholar]
- 40.Blattner FR, Tucker PW. The molecular biology of immunoglobulin D. Nature (Lond) 1984;307:417–422. doi: 10.1038/307417a0. [DOI] [PubMed] [Google Scholar]
- 41.Coico RF, Xue B, Wallace D, Pernis B, Siskind GW, Thorbecke GJ. T cells with receptors for IgD. Nature (Lond) 1985;316:744–746. doi: 10.1038/316744a0. [DOI] [PubMed] [Google Scholar]
- 42.Coico RF, Siskind GW, Thorbecke GJ. Role of IgD and Tδ cells in the regulation of the humoral immune response. Immunol Rev. 1988;105:45–67. doi: 10.1111/j.1600-065x.1988.tb00765.x. [DOI] [PubMed] [Google Scholar]
- 43.Flotte TJ, Finkelman FD, Thorbecke GJ. Polyclonal activation of the murine immune system by antibody to IgD V. Effect on germinal centers. Eur J Immunol. 1984;14:725–728. doi: 10.1002/eji.1830140810. [DOI] [PubMed] [Google Scholar]
- 44.Carsetti R, Köhler G, Lamers MC. A role for immunoglobulin D: interference with tolerance induction. Eur J Immunol. 1993;23:168–178. doi: 10.1002/eji.1830230127. [DOI] [PubMed] [Google Scholar]
- 45.Monroe JG, Havran WL, Cambier JC. B lymphocyte activation: entry into cell cycle is accompanied by decreased expression of IgD but not IgM. Eur J Immunol. 1983;13:208–213. doi: 10.1002/eji.1830130306. [DOI] [PubMed] [Google Scholar]
- 46.Black SJ, Van der Loo W, Loken MR, Herzenberg LA. Expression of IgD by murine lymphocytes. Loss of surface IgD indicates maturation of memory B cells. J Exp Med. 1978;147:984–996. doi: 10.1084/jem.147.4.984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Galibert L, Durand I, Rousset F, Banchereau J. CD40 activated surface IgD positive lymphocytes constitute the long term IL-4 dependent proliferating B cell pool. J Immunol. 1994;152:22–29. [PubMed] [Google Scholar]
- 48.Lafrenz D, Strober S, Vitetta E. The relationship between surface immunoglobulin isotype and the immune function of murine B lymphocytes. V. High affinity secondary antibody responses are transferred by both IgD-positive and IgD-negative memory B cells. J Immunol. 1981;127:867–872. [PubMed] [Google Scholar]
- 49.Herzenberg LA, Black SJ, Tokuhisa T, Herzenberg LA. Memory B cells at successive stages of differentiation. Affinity maturation and the role of IgD receptors. J Exp Med. 1980;151:1071–1087. doi: 10.1084/jem.151.5.1071. [DOI] [PMC free article] [PubMed] [Google Scholar]