

Abstract
The structurally related T cell surface molecules CD28 and CTLA-4 interact with cell surface ligands CD80 (B7-1) and CD86 (B7-2) on antigen-presenting cells (APC) and modulate T cell antigen recognition. Preliminary reports have suggested that CD80 binds CTLA-4 and CD28 with affinities (K d values ∼12 and ∼200 nM, respectively) that are high when compared with other molecular interactions that contribute to T cell–APC recognition. In the present study, we use surface plasmon resonance to measure the affinity and kinetics of CD80 binding to CD28 and CTLA-4. At 37°C, soluble recombinant CD80 bound to CTLA-4 and CD28 with K d values of 0.42 and 4 μM, respectively. Kinetic analysis indicated that these low affinities were the result of very fast dissociation rate constants (koff); sCD80 dissociated from CD28 and CTLA-4 with koff values of ⩾1.6 and ⩾0.43 s−1, respectively. Such rapid binding kinetics have also been reported for the T cell adhesion molecule CD2 and may be necessary to accommodate dynamic T cell–APC contacts and to facilitate scanning of APC for antigen.
CD28 (1, 2) and CTLA-4 (3) are structurally related cell surface molecules, largely restricted to the T cell lineage, which contribute to antigen recognition by T cells (for reviews see references 4–6). They do so by interacting with the structurally related ligands CD80 (B7-1, B7; 7, 8) and CD86 (B7-2, B70; 9–12), which are expressed on APC. Numerous reports (reviewed in references 4–6) have shown that, upon ligation, CD28 transmits activation signals that enhance T cell activation through the TCR. The role of CTLA-4 is more controversial (13), but recent in vivo studies (14–16) suggest that, unlike CD28, CTLA-4 ligation inhibits T cell activation, apparently by inhibiting signaling through the TCR (17).
Antigen recognition by T cells is unusual in that it occurs at the interface between two cells in direct physical contact, and CD28 is one of several T cell surface molecules that participate in this interaction (18, 19). It is becoming evident that T cells are sensitive to quantitative changes in the molecular interactions that contribute to T cell antigen recognition, and that quantitative differences in, for example, binding affinity, kinetics, or surface density, can lead to very different T cell responses (20–24). Clearly, our understanding of antigen recognition would be aided by knowledge of the affinity and binding kinetics of the individual molecular interactions. Recent studies have provided affinity and kinetic data on the interactions of the TCR with peptide–MHC (25) and CD2 with its ligands CD48 (26) and CD58 (27, 28). In contrast, the interactions of CD28 and CTLA-4 with their ligands are less well characterized. Estimates have been made of the relative avidity and kinetics of CD28 and CTLA-4 binding to CD80 and CD86 (8, 29, 30), but no definitive affinity or kinetic data are available. A soluble CD80 Ig fusion protein was reported to bind to CD28 with a dissociation constant (K d) of ∼200 nM (29) and to CTLA-4 with a K d of ∼12 nM (8), but there was evidence that this CD80 Ig was not monomeric in solution (29). More recent kinetic studies have relied entirely on dimeric Ig fusion proteins (30, 31). Although potentially informative for comparative purposes, such studies do not provide accurate affinity and kinetic data of the sort that can be used to develop quantitative models of Ag recognition by T cells.
In the present study, we undertook to obtain precise estimates of the solution affinity and kinetic constants for the interaction between CD80 and both CD28 and CTLA-4. Molecules involved in cell–cell recognition can have very low affinities and fast dissociation rate constants, leading to technical problems in their measurement (32, 33). Recent studies (for reviews see 33, 34) have demonstrated that surface plasmon resonance (SPR)1, as implemented in the BIAcore instrument (35), is well suited to analysis of such interactions. We use SPR to show that CD80 binds CD28 and CTLA-4 with affinities considerably lower than previously reported, and with very fast kinetics.
Materials and Methods
Expression of CTLA-4 Ig.
The CTLA-4 Ig construct was very similar to that previously described (8) and was made by PCR, using as a template a cDNA library prepared from human peripheral blood acute lymphoblastic leukemia cells (Daenke, S., unpublished data). The 116 base 5′ primer CTAGCCACTGAAGCTTCACCAATGGGTGTACTGCTCACACAGAGGACGCTGCTCAGTCTGGTCCTTGCACTCCTGTTTCCAAGCATGGCGAGCATGGCAATGCACGTGGCCCAGCC encoded the oncostatin M leader (36) and 20 bases of CTLA-4 sequence and incorporated a HindIII restriction site. The encoded junction between the oncostatin leader (uppercase) and CTLA-4 (lowercase) was as follows: SMASMamhva. The 3′ primer ACGGATCCTTATCAGAATCTGGGCACGGTTC encoded the CTLA-4 sequence immediately upstream of the transmembrane region followed by a splice donor site and a BamHI restriction site. The amplified product was subcloned into the pIg expression vector (37) using its BamHI and HindIII restriction sites. pIg incorporates, downstream of the cloning site, a genomic sequence encoding the constant region of human IgG γ1 heavy chain. After splicing, the encoded junction between CTLA-4 (uppercase) and the γ1 heavy chain (lowercase) would be as follows: CPDSDepsksc. The sequence was confirmed by dideoxy sequencing. Stable high level expression of CTLA-4 Ig in Chinese hamster ovary (CHO)– K1 cells was achieved by calcium phosphate cotransfection of the CTLA-4 Ig encoding plasmid together with the glutamine synthetase–encoding expression vector pEE14, as previously described (38–40). Clones expressing high levels of CTLA-4 Ig (∼30–40 mg/L) were identified by inhibition ELISA using a mAb specific for human IgG (40) and grown up to confluence in bulk culture before switching to serum-free medium supplemented with 2 mM Na butyrate. CTLA-4 Ig was then purified by affinity chromatography on protein A–sepharose (Pharmacia Biotech AB, Uppsala, Sweden) followed by size-exclusion chromatography on a SUPERDEX S200 HR10/30 column (Pharmacia Biotech).
Expression and Analysis of sCD80.
The soluble CD80 construct was designed to encode the extracellular portion of CD80 up to lysine-209 followed by a carboxy-terminal oligo-histidine tag for purification (sCD80his). It was made by PCR using cDNA from MT-2 cells (HTLV-1-transformed human T cells) as template. The 5′ primer TAGTAGAAGCTTTCCCCATCCGCTCAAGCAGGCCACCATGGGCCACACACGGAGG was complementary to the CD80 leader sequence but added an HindIII site and inserted, immediately upstream of the initiation codon, the 25 bases that precede the rat CD4 initiation codon (41). The 3′ primer TAGTAGTCTAGACTAATGATGATGATGATGATGCTTGGCTGTATTCCAGTTGAAGGT added six histidine residues and a stop codon after lysine 209, mutated threonine 208 to alanine to remove a potential NH2-linked glycosylation site, and added a XbaI site. The 10 carboxy-terminal amino acids of sCD80his were thus NTAKHHHHHH. The resulting PCR fragment was subcloned into the glutamine synthetase expression vector pEE14 (39) using its XbaI and HindIII restriction sites, and the sequence was confirmed by dideoxy sequencing. CHO-K1 cells were transfected as described (38, 39) with the sCD80hisencoding plasmid by calcium phosphate transfection. Clones expressing high levels of sCD80his (∼40 mg/L) were identified by growth in the presence of [35S]methionine/[35S]cysteine (TRANS35SLABEL; ICN Pharmaceuticals, Costa Mesa, CA), purification of labeled protein from the culture supernatant using Ni-NTA spin columns (Qiagen GmbH, Hilden, Federal Republic of Germany), and then SDS-PAGE of the protein followed by autoradiography. The best clone was grown up to confluence in bulk culture before switching to serum-free medium supplemented with 2 mM Na butyrate. sCD80his was purified by affinity chromatography using Ni-NTA resin (Qiagen GmbH) followed by size-exclusion chromatography on a SUPERDEX S200 HR10/30 column. The extinction coefficient (at 280 nm) of sCD80his was determined by amino acid analysis to be 1.41 ml.mg−1. The carboxy-terminal his tag was cleaved off by incubating 2.5 mg of sCD80his in 1.5 ml Tris–saline buffer (140 mM NaCl, 10 mM Tris [pH 7.5]) with 1.2 U of carboxypeptidase A conjugated to agarose beads (Sigma Chemical, Poole, UK) for 16 h at 30°C with agitation. Amino acid analysis confirmed that ⩾90% of the carboxy-terminal histidine residues (5.4 molecules per sCD80his molecule), but no other amino acids, were released during this incubation (data not shown). The carboxypeptidase A was removed by centrifugation and the digested sCD80his (sCD80) was repurified on a SUPERDEX S200 HR10/30 column (Fig. 1 C).
Figure 1.

Biochemical analysis of sCD80 and CTLA-4 Ig. (A) sCD80 and CTLA-4 Ig (2.5 μg each) were analyzed by SDS-PAGE on 10% acrylamide under nonreducing and reducing (+βME) conditions. The migration positions of the indicated (in kD) molecular mass markers are shown. (B) Measuring the binding activity of sCD80. Protein A–sepharose beads coated with CTLA-4 Ig or CD22 Ig were incubated with soluble sCD80, pelleted, and the supernatant analyzed for the presence of sCD80 by reducing SDS-PAGE on 12% acrylamide, together with sCD80 not exposed to beads (Control). (C) Analysis of sCD80 by size-exclusion chromatography. sCD80 (2.1 mg in 0.5 ml) was run on a SUPERDEX S200 HR10/30 column (Pharmacia) at 0.5 ml/min. The calibration markers shown (Sigma) were alcohol dehydrogenase (M r 150,000), BSA (M r 66,000), and carbonic anhydrase (M r 29,000). sCD48–CD4 and sCD2 are described in the text.
The proportion of sCD80 that possessed ligand binding activity was determined as follows. Protein A–sepharose beads (100 μl) were incubated with 0.4 mg of CTLA-4 Ig or (as a control) CD22 (d1-3) Ig (42) in 0.33 ml Tris–saline for 1 h at 4°C with rotation and then washed three times with 1.5 ml Tris–saline. CTLA-4 Ig or CD22 Ig beads (40 μl) were then added to 20 μl sCD80 (0.65 μg/μl in Tris–saline) and incubated for 4 h at 4°C with rotation. The beads were then pelleted and 10 μl of each supernant analyzed by SDS-PAGE, together with 3.3 μl of sCD80 that had not been exposed to beads (Fig. 1 B).
SPR Experiments.
Binding experiments were performed by SPR on a BIAcore instrument (35) upgraded with the BIAcore Upgrade Kit (BIAcore AB, St Albans, UK). All experiments were performed at 37°C (except where otherwise indicated) using HBSS buffer (25 mM Hepes [pH 7.4], 150 mM NaCl, 3.4 mM EDTA, and 0.005% surfactant P20) supplied by BIAcore. CTLA-4 Ig and CD28 Ig were covalently coupled by primary amine groups to the carboxymethylated dextran matrix on a research grade CM5 sensor chip (BIAcore) using the Amine Coupling Kit (BIAcore) as directed (43), with the following modifications. After an activation step of 15–240 s, CTLA-4 Ig (30 μg/ml in 10 mM Na acetate [pH 4.7]) or CD28 Ig (15 μg/ml in 10 mM Na acetate [pH 5]) were injected for 7 min. After coupling, CTLA-4 Ig and CD28 Ig were washed with a 3 min injection of NaOH (5 mM). NaOH (5 mM) was also used to regenerate immobilized CD28 Ig and CTLA-4 Ig. The anti-human IgG1 mAb R10Z8E9 was used as previously described (44) to immobilize CD28 Ig and CTLA-4 Ig indirectly to the sensor surface via their Ig fragment.
Analysis of Affinity and Kinetic Data.
Equilibrium binding analysis and analysis of the washout or dissociation phase was performed as previously described (27). Analysis of the injection or association phase was performed using the BIAevaluation software Version 2.1 (BIAcore). The kon was determined by nonlinear curve fitting of the following equation to the injection phase.
(1)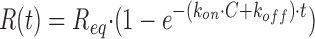
where R(t) is the response (in response units) at time t, Req is the response at equilibrium, and C is the concentration of injected sCD80. The koff value used for this fit was obtained by analysis of the washout phase of the same injection. For the CTLA-4–sCD80 interaction, the kon was also estimated using an approach relying entirely on analysis of the injection phase, as described by Karrlson et al. (43). After injection at several different sCD80 concentrations, plots of dR/dt versus R for each concentration (C) give straight lines with a slope −k s where
(2)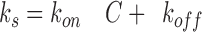
Values for k on and k off were obtained by a linear fit of a plot of k s versus C (see Fig. 4 E).
Figure 4.

Estimating the k on and k off for sCD80 binding CTLA-4 Ig. (A) Example of primary data. sCD80 (265 nM) was injected (solid bar) at 40 μl/min through FCs with nothing immobilized (Control) or CTLA-4 immobilized at low (920 RUs) or high (2500 RUs) levels. (B) Effect of varying the flow rate. sCD80 (265 nM) was injected (solid bar) at 20 (solid line) or 40 (stippled line) μl/min through FCs with high or low levels of CTLA-4. Background responses (following injection through a control FC) have been subtracted. (C) Dissociation of sCD80 from FC with high (•, ○) or low (▴, ▵) levels of CTLA-4 Ig at flow rate of 20 (•, ▴, ▪) or 40 (○, ▵, □) μl/min. Also shown is the fall in response in the same period following injection of sCD80 through control FC (▪, □). The data fitted reasonably well to single exponential decay curves (dotted lines), yielding the following t1/2 values: •, 4.8 s; ○, 4.4 s; ▴, 2.2 s; ▵, 2.35 s; ▪, 0.16 s; □, 0.075 s. (D) Estimating the k on. Equation 1 (see Materials and Methods) was fitted (solid line) to data (•) from (B) (corresponding to binding of sCD80 to CTLA-4 [low level] at 40 μl/min), yielding the indicated residuals (▪) and k on. (E) Independent estimation of k on and k off by analysis of binding at different sCD80 concentrations. The rate at which the binding rate decreases (−k s) during the injection phase was determined for a range of injected sCD80 concentrations (see Materials and Methods [43]) using data from Fig. 2 A. A linear regression fit of equation 2 to a plot of k s versus sCD80 concentration yielded the k on and k off values.
Results and Discussion
Expression and Characterization of Monomeric sCD80.
A soluble form of CD80 with a carboxy-terminal oligo-his tag (sCD80his) was expressed in CHO-K1 cells. Preliminary experiments indicated that the oligo-histidine tag predisposed towards the formation of multimeric aggregates (van der Merwe, P.A., and S.J. Davis, unpublished data) and so it was removed by digestion with carboxypeptidase A (see Materials and Methods) to yield sCD80. Analysis of sCD80 by reducing SDS-PAGE indicated that it migrated as a broad band with a size range of Mr 35–45,000 (Fig. 1 A). This is consistent with the predicted size of the protein backbone (M r 24,747) and variable glycosylation of the seven potential NH2-linked glycosylation sites. sCD80 migrated slightly faster (M r 30–43,000) under nonreducing conditions, indicating that the protein does not form disulphide-linked multimers, and consistent with the existence of two predicted disulphide bonds (45).
For valid affinity and kinetic analysis, it is critical that the interaction under study is monovalent (32), because increasing the binding valency leads to dramatic (but unpredictable) increases in the strength and stability of an interaction. The use of the term affinity is usually restricted to descriptions of the binding strength of monovalent interactions, whereas avidity is used to describe the binding strength of multivalent interactions. In the present study, monovalency was ensured by using a monomeric form of CD80 (sCD80). To check that sCD80 was indeed monomeric in solution, it was analyzed by size-exclusion chromatography (Fig. 1 C). sCD80 eluted at M r ∼63,000 when using globular unglycosylated proteins to calibrate the column (Fig. 1 C). Although this is slightly higher than might be expected, sCD80 is heavily glycosylated and probably asymmetric. Therefore, we compared sCD80 with two similarly asymmetric glycosylated proteins, namely soluble rat CD2 (sCD2, Mr 26–34,000 on SDS-PAGE [46]) and soluble rat CD48–CD4 (sCD48–CD4, Mr 43–64,000 on SDS-PAGE [26, 47]), both of which are monomeric (26, 47). sCD80 eluted after sCD48–CD4 and ahead of sCD2 (Fig. 1 C), consistent with it existing as a monomer in solution.
sCD80 appeared to be folded correctly as it bound to all four mAbs tested (Table 1) and also bound both CD28 and CTLA-4 (see below). Accurate estimation of affinity and kinetic constants requires knowledge of the exact concentration of the soluble ligand, which in turn requires that the proportion of sCD80 that retains ligand binding activity is known. All of the sCD80 could not be assumed to be active because it had been purified using its oligo-histidine tag, and this would not discriminate between correctly and incorrectly folded forms of sCD80. The activity of sCD80 therefore was estimated directly by depleting the sCD80 with protein A–sepharose beads coated with CTLA-4 Ig (Fig. 1 B). CTLA-4 Ig beads were able to deplete all the sCD80 (Fig. 1 B), whereas CD22 Ig beads did not deplete, indicating that close to 100% of the sCD80 retains CTLA4-binding activity.
Table 1.
mAbs that Bind the Recombinant Proteins Used in This Study
| Protein | mAb | |
|---|---|---|
| sCD80* | 2D10, mAb 104, L307, BB-1 | |
| CD28 Ig‡ | CD28.1, CD28.2, CD28.3, CD28.5, CD28.6, | |
| CLB–CD28/1, KOLT-2 | ||
| CTLA-4 Ig§ | 7F8, 10A8, 11D4 |
2D10 (64) was from Dr. Daniel Olive (Inserm 119, Institut Paoli Calmettes, Marseilles, France). mAb 104 (65) was from Immunotech S.A. (Marseilles, France). L307 (66) was from Becton Dickinson (San Jose, CA). BB-1 (67) was from PharMingen (San Diego, CA).
CD28.1–CD28.6 (68) were from Dr. Daniel Olive. CLB–CD28/ 1(15E8 in [66]) was from Dr. René A. W. van Lier (Netherlands Red Cross Blood Transfusion Service, Amsterdam, the Netherlands). KOLT-2 (69) was from Dr. Kimitaka Sagawa (Department of Immunology, Kurume University School of Medicine, Kurume, Japan).
7F8, 10A8, 11D4 were produced as described (54).
Expression and Analysis of CD28 Ig and CTLA-4 Ig.
The recombinant CD28 used in the present study (CD28 Ig) was a homodimeric fusion protein incorporating the Fc portion of human γ1 Ig heavy chain (29). We expressed and purified a similar CTLA-4 Ig fusion protein (see Materials and Methods). SDS-PAGE under reducing and nonreducing conditions indicated that CTLA-4 Ig was expressed as a disulphide-linked dimer (Fig. 1 A). For kinetic and affinity studies, CTLA-4 Ig and CD28 Ig were immobilized to a dextran matrix on the sensor surface either indirectly, via a mAb specific for human IgG1, or by direct covalent coupling through primary amines (see Materials and Methods). CD28 and CTLA-4 mAbs (Table 1) bound to both directly and indirectly coupled forms of CD28 Ig and CTLA-4 Ig (data not shown). Furthermore, sCD80 bound with a similar affinity to both directly and indirectly coupled CD28 Ig and CTLA-4 Ig (see below; Table 2), indicating that direct covalent coupling did not substantially disrupt the native structure of these proteins. All subsequent measurements were made using directly coupled CD28 Ig and CTLA-4 Ig.
Table 2.
Summary of Affinity Constants Measured by Equilibrium Binding
| Binding of sCD80 to | Immobi- lization* | Temperature | K d (μM)‡ | ||||
|---|---|---|---|---|---|---|---|
| CTLA-4 Ig | Direct | 37°C | 0.42 (± 0.06, n = 3) | ||||
| Direct | 25°C | 0.26 (± 0.06, n = 3) | |||||
| Indirect | 25°C | 0.2 (n = 1) | |||||
| CD28 Ig | Direct | 37°C | 4.0 (± 0.3, n = 4) | ||||
| Direct | 25°C | 2.5 (± 0.4, n = 2) | |||||
| Indirect | 37°C | 5.5 (n = 1) | |||||
Direct immobilization was via primary amines on CTLA-4 Ig or CD28 Ig (see Materials and Methods). Indirect immobilization was via a directly coupled mAb that binds the Ig portion of CTLA-4 Ig and CD28 Ig, as previously described (44).
The values shown are the means of n determinations (± SD for n ⩾3 or ± range for n = 2).
Affinity Measurements.
Affinity and kinetic measurements were performed at 37°C except where otherwise indicated. The affinity of sCD80 binding to CTLA-4 and CD28 was measured directly by equilibrium binding analysis (Figs. 2 and 3), because this avoids the many potential pitfalls associated with kinetic measurements (see below; 48–50). Increasing concentrations of sCD80 were injected over sensor surfaces to which CTLA-4 Ig or CD28 Ig had been immobilized (Figs. 2 A and 3 A, respectively). It is noteworthy that at all sCD80 concentrations binding reached equilibrium very rapidly (95% binding within 20 s; faster for CD28) and that, in the washout phase after the injection, bound sCD80 dissociated within 20 s (faster for CD28). These features are typical of interactions with very fast binding kinetics. The sCD80 samples were also injected over a control sensor surface (with no protein immobilized) to measure the background response (Figs. 2 A and 3 A, right). Larger background responses are seen in Fig. 3 A because a tenfold higher range of sCD80 concentrations was injected over CD28 Ig (Figs. 2 and 3, legends). For each sCD80 concentration the binding response (measured in arbitrary response units [RU]) at equilibrium was calculated by subtracting the response seen in the control flow cell from the response seen in the CTLA-4 (see Fig. 2 B) or CD28 (Fig. 3 B) flow cells. A plot of the binding response indicates saturable binding and direct fitting of a standard Langmuir binding isotherm to the data gave K d values of 0.38 and 4.2 μM for sCD80 binding to CTLA-4 and CD28, respectively (Figs. 2 B and 3 B, left). Scatchard plots of the data gave very similar K d values (Figs. 2 B and 3 B, right). The mean K d (± SD) values from several independent determinations are shown in Table 2.
Figure 2.

Measuring the affinity of sCD80 binding to CTLA-4 Ig by equilibrium binding. (A) A range of sCD80 concentrations (2.65 μM and nine twofold dilutions thereof) were injected sequentially (solid bar) for 2 min at 10 μl/min through a flow cell (FC) with either CTLA-4 Ig (1,500 RUs) or no protein (Control) immobilized. sCD80 (0.26 μM) bound to a very similar level (255 and 250 RUs) when injected at the beginning and end of the experiment (data not shown), indicating that the immobilized CTLA-4 Ig was stable. (B) The equilibrium responses in the CTLA-4 Ig (•) and Control (▴) FCs at each sCD80 concentration and the differences between these responses (representing actual binding, ▪) are plotted. The dotted line represents a nonlinear fit of the Langmuir binding isotherm to the binding data and yields a K d of 0.4 μM and a binding maximum of 774 RUs. A Scatchard plot of the same data is shown on the right. A linear regression fit yields a K d of 0.4 μM and a binding maximum of 750 RUs.
Figure 3.

Measuring the affinity of sCD80 binding to CD28 Ig by equilibrium binding. (A) A range of sCD80 concentrations (26.5 μM and eight twofold dilutions thereof) were injected sequentially (solid bar) for 30 s at 10 μl/min through a FC with either CD28 Ig (immobilized at a level of 7800 RUs) or an irrelevant protein (Control) immobilized. sCD80 (0.82 μM) injected at the beginning and end of the experiment bound to a very similar level (160 and 157 RUs), indicating that the immobilized CD28 Ig was stable. (B) The equilibrium responses in the CD28 Ig (•) and Control (▴) FCs at each sCD80 concentration and the differences between these responses (representing actual binding, ▪) are plotted. The dotted line represents a nonlinear fit of the Langmuir binding isotherm to the binding data and yields a K d of 4.1 μM and a binding maximum of 939 RUs. A Scatchard plot of the same data is shown on the right. A linear regression fit yields a K d of 4.3 μM and a binding maximum of 952 RUs.
Kinetic Measurements.
Measurements of binding kinetics are prone to error, particularly when the kinetics are very fast, as in the present study (48–50). One source of error is that part of the response is a background signal that does not represent true binding. To correct for this, the response obtained when sCD80 was injected through a control flow cell (containing no immobilized protein) was subtracted before analysis. Figs. 4 A and 5 A show typical responses obtained after injection of sCD80 through flow cells with two different levels of CTLA-4 Ig (or CD28 Ig) immobilized, as well as through a control flow cell. Subtraction of the control flow cell response from the responses in the CD28 and CTLA-4 flow cells gives the actual binding response shown in Figs. 4 B and 5 B (solid lines), which was subjected to kinetic analysis. A second potential source of error is that binding and dissociation are limited by the rate at which protein is delivered to and removed from the sensor surface. This was addressed by using higher than normal flow rates (40 or 80 μl/min). These flow rates were judged to be optimal because halving them had little effect on the association (injection) or dissociation (washout) phases when sCD80 was injected over CTLA-4 (see Figs. 4 B and C) or CD28 (Figs. 5, B and C). It should be noted that even at very high flow rates, binding may still be limited by mass-transport within an unstirred layer close to the binding surface (48, 50). The effects of mass-transport limitations within this unstirred layer can be reduced by decreasing the level of immobilized ligand (see below). A third source of error is rebinding of protein during the dissociation (washout) phase, which will lead to underestimation of the dissociation rate. Rebinding can be decreased by immobilizing lower levels of ligand. This is illustrated in Figs. 4 C and 5 C, in which sCD80 dissociates more rapidly when the level of immobilized CTLA-4 or CD28 is decreased. sCD80 dissociated from sensor surfaces with high and low levels of CTLA-4 with apparent dissociation rate constants (k off) of 0.24 s−1 and 0.43 s−1, respectively (see Fig. 4 C; Table 3). Similarly, sCD80 dissociated from high and low levels of CD28 with k off values of 1.1 s−1 and 1.6 s−1, respectively (Fig. 5 C; Table 3). Although it is quite likely that some sCD80 rebinding is still occurring even at these lower CD28 Ig and CTLA-4 Ig levels, it proved difficult to collect data of satisfactory quality when CD28 Ig and CTLA-4 Ig levels were reduced further. Therefore, we assume that the actual k off values for dissociation of sCD80 from CTLA-4 and CD28 are at least 0.43 s−1 and 1.6 s−1, respectively.
Figure 5.

Estimating the k on and k off for sCD80 binding CD28 Ig. (A) Example of primary data. sCD80 (2.65 μM) was injected (solid bar) at 80 μl/min through FCs with nothing immobilized (Control) or CD28 Ig immobilized at low (3400 RUs) or high (6200 RUs) levels. (B) Effect of varying the flow rate. sCD80 (2.65 μM) was injected (solid bar) at 40 (solid line) or 80 (stippled line) μl/min through FCs with high or low levels of CD28 Ig. Background responses (following injection through a control FC) have been subtracted. (C) Dissociation of sCD80 from FC with high (•, ○) or low (▴, ▵) levels of CD28 Ig at flow rate of 40 (•, ▴, ▪) or 80 (○, ▵, □) μl/min. Also shown is the fall in response in the same period following injection of sCD80 through a control FC (▪, □). The data fitted well to single exponential decay curves (dotted lines), yielding the following t1/2 values: •, 0.93 s; ○, 0.84 s; ▴, 0.69 s; Δ, 0.64 s; ▪, 0.075 s; □, 0.04 s. (D) Obtaining the k on by nonlinear curve fitting. Eq. 1 (see Materials and Methods) was fitted (solid line) to data (•) from (B) (corresponding to binding of sCD80 to CD28 Ig [low level] at 80 μl/min), yielding the indicated residuals (▪) and k on.
Table 3.
Summary of Kinetic Measurements
| Binding of sCD80 to | Immobilization level* | k on (× 10−5 M−1s−1) | k off (s−1) | K dcalc (k off/k on, μM) | ||||
|---|---|---|---|---|---|---|---|---|
| CTLA-4 Ig | High | 6.2 ± 1.8§ (n = 3) | 0.24 ± 0.03 (n = 3) | 0.3 | ||||
| High (linear regression)‡ | 3.4 (n = 1) | 0.2 (n = 1) | 0.63 | |||||
| Low | 9.4 ± 0.3 (n = 2) | 0.43 ± 0.03 (n = 3) | 0.46 | |||||
| CD28 Ig | High | 4.7 ± 0.6 (n = 4) | 1.1 ± 0.15 (n = 3) | 2.3 | ||||
| Low | 6.6 ± 0.8 (n = 4) | 1.6 ± 0.1 (n = 4) | 2.4 |
CTLA-4 Ig or CD28 Ig were immobilized directly. For CTLA-4 Ig, high and low levels were 2,500 and 900 RUs, respectively. For CD28 Ig, high and low levels were 6,200 and 3,400 RUs, respectively.
Data from Fig. 4 E.
The values shown are the mean of n determinations (± SD for n ⩾3 or ± range for n = 2).
Association rate constants (k on) for binding were estimated by nonlinear curve fitting to data from the injection phase (Figs. 4 D and 5 D) using k off values obtained from analysis of the corresponding dissociation phase. Reasonably good fits were obtained with a simple one-to-one (Langmuir) binding model (Eq. 1). When CTLA-4 and CD28 were immobilized at the lower levels, sCD80 bound with mean k on values of 9.4 × 105 M−1s−1 and 6.6 × 105 M−1s−1, respectively (Table 3). Slighty slower k on values were obtained with higher levels of immobilized CTLA-4 (6.2 × 105 M−1s−1) and CD28 (4.7 × 105 M−1s−1) (Table 3). As discussed above, this is likely to reflect underlying masstransport limitations within the unstirred layer, which would be noticeable at higher levels of immobilized ligand (49, 50). Because it was not possible to eliminate completely the effect of mass transport limitations, we assume that the actual k on values for sCD80 binding to CTLA-4 and CD28 are at least 9.4 × 105 M−1s−1 and 6.6 × 105 M−1s−1, respectively.
Kinetic constants for CTLA-4 binding were also estimated independently of the dissociation phase by analyzing the association phase after injection of different concentrations of sCD80 (see Materials and Methods [43]). Although less accurate and less stringent than a direct fit of equation 1 (48), this approach is useful for verification since it does not require prior measurement of the k off, but instead provides its own independent estimate (see Fig. 4 E). Such an analysis was not possible for the lower affinity CD80–CD28 interaction because it has faster kinetics (compare Figs. 4 D and 5 D) and requires much higher sample concentrations. The k on (3.2 × 105 M−1s−1) and k off (0.2 s−1) values measured in this manner for sCD80 binding to CTLA-4 Ig were only slightly slower than values obtained by the direct nonlinear curve fitting with similar levels of CTLA-4 (see Table 2).
The accuracy of the measured rate constants can be further assessed by comparing the ratio between the k off and the k on (which provides a calculated K d or K dcalc) with the K d measured directly by equilibrium binding (Tables 2 and 3). For both CTLA-4 and CD28, the K d and K dcalc were very similar (Tables 2 and 3), indicating consistency between the kinetic and equilibrium binding analyses. However, this internal consistency does not exclude the possibility that the true k on and k off values are both faster than the measured values. Therefore, the measured k on and k off values represent lower limits for the true k on and k off.
Comparisons with Other Studies.
The affinities measured in the present study for CD80 binding CD28 (K d 4 μM) and CTLA-4 (K d 0.42 μM) are much lower than previously reported (8, 29). One possible difference is that these earlier measurements were made at 23°C (8, 29). However, we found that decreasing the temperature from 37°C to 25°C increased the affinity less than twofold (Table 2). A more significant difference is likely to be that the soluble CD80 Ig used in the earlier studies was not monomeric. This is supported by the observation that the CD80 Ig fusion protein used in these earlier studies, although monomeric on SDS-PAGE (M r 70,000), eluted at M r ∼350,000 by size-exclusion chromatography (29), suggesting the presence of higher aggregates. Such aggregates could well account for the much higher affinities previously reported. However, this explanation does not account for 100- to 200-fold higher avidity observed for soluble CTLA-4 Ig compared with soluble CD28 Ig (for example see reference 30), a difference much larger than the 10-fold difference in affinity reported in the present study. This discrepancy is not due to differences in the recombinant proteins used in these studies, because we were able to reproduce the large difference in avidity by injecting CD28 Ig and CTLA4 Ig over a sensor surface onto which sCD80 had been immobilized (Fig. 6). CTLA-4 Ig injected at 0.1 and 1 μg/ml bound to higher levels than 100-fold higher concentrations of CD28 Ig (Fig. 6), indicating a >100-fold higher avidity. Although it is unclear why the avidity difference between CTLA-4 Ig and CD28 Ig is so much greater than the affinity difference, these results illustrate a major disadvantage of using multimeric proteins for comparative binding studies.
Figure 6.

Comparison of avidity of CD28 Ig and CTLA-4 Ig binding to immobilized sCD80. CD28 Ig or CTLA-4 Ig were injected at 5 μl/ min (25°C) at the indicated concentration for 6–8 min through FC with either sCD80 (3300 RU) or no protein (Control) immobilized. The responses seen with injection of the three CTLA4 Ig concentrations through the Control FC were indistinguishable when superimposed.
A similar affinity constant for the CTLA-4–CD80 interaction (K d 0.21 μM at 25°C) has recently been obtained by SPR using a monomeric form of CTLA-4 (51) in solution and a CD80 Ig fusion protein (29) covalently immobilized onto the sensor surface (52). No monomeric form of CD28 was analyzed by Greene et al. (52) but, unexpectedly, an apparently dimeric form of CD28 (CD28tp) bound the immobilized CD80 Ig with an avidity (K d 2.1–41 μM at 25°C) similar to the affinity that we obtained for monomeric sCD80 binding to immobilized CD28 (K d 2.5 μM at 25°C). CD28tp also bound immobilized CD80 Ig with an unusually slow apparent k on (7,000 M−1s−1). One explanation for this discrepency, which is consistent with the slow apparent k on, is that only a small proportion of the CD28tp was active. An alternative explanation is that covalent immobilization of the CD80 Ig alters preferentially its interaction with CD28 but not CTLA-4.
Functional Implications.
To understand the functional implications of the solution affinity and kinetic constants reported in the present study, it is necessary to extrapolate these values to interactions between the membrane-tethered forms of these molecules (28, 53). Like CD28, CD2 binds its ligand CD58 in solution with a very low affinity and rapid binding kinetics (27). Dustin et al. (28) have shown that the interaction between membrane-tethered CD2 and CD58 also exhibits rapid binding kinetics. This suggests that the fast solution-binding kinetics observed between these molecules is a reflection of rapid binding kinetics when they are attached to membrane. Theoretical considerations (53) suggest that the difference in the affinity of soluble CD80 for CD28 versus CTLA-4 is likely to be conserved for membrane-attached forms of these molecules. If correct, this implies that CTLA-4 will engage ∼10-fold lower surface densities of CD80 than CD28 because of its ∼10 fold higher affinity. Unlike CD28, CTLA-4 is not expressed on resting T cells (4) and, although expression is induced on T cell activation, CTLA-4 is always present at a ∼30- to 50-fold lower surface density than CD28 (54). Under these circumstances CTLA-4 is unlikely to compete effectively for CD80 and displace it from CD28, despite its higher affinity. Instead, both CD28 and CTLA-4 are likely to engage CD80 when they are expressed simultaneously. In this regard, it is noteworthy that inhibitory signals transmitted through CTLA-4 appear to be dominant over the activation signals transmitted through CD28 (55, 56).
The affinity constants reported in the present study and previous studies of CD2 and CD4 suggest that accessory T cell molecules, including those with signaling functions, have affinities comparable to or lower than the reported affinities of TCRs for their specific peptide–MHC ligands (Table 4). Even more striking are the 10- to 100-fold faster k off values seen for the CD2–CD58 and CD28–CD80 interactions compared with TCR/peptide–MHC interactions (Table 4). While it is possible that future studies will identify TCR/ peptide–MHC interactions with similarly fast k off values, these results suggest that the accessory molecule interactions are generally more unstable than TCR interactions. Indeed, the binding of CD4 and CD8 to MHC class II and MHC class I is likely to stabilize further TCR/peptide– MHC complexes relative to accessory molecule interactions (57).
Table 4.
Comparison of the Binding Constants for the Interactions of the TCR, CD2, CD4, and CD28 with Their Respective Ligands
| Interaction | kon (M−1s−1) | koff (s−1) | K d * (μM) | References | ||||
|---|---|---|---|---|---|---|---|---|
| TCR binding peptide–MHC | ||||||||
| 2B4 TCR–MCC peptide plus I-Ek | 900–1,700 | 0.057–0.09 | 5–50 | (70, 71) | ||||
| 2C TCR–p2C peptide plus Ld | 11,000–210,000 | 0.006–0.026 | 0.1–0.5 | (72, 73) | ||||
| 42.12 TCR–OVA peptide plus Kb | 3,100 | 0.02 | 7–19 | (22) | ||||
| CD2–CD58 | ⩾400,000 | ⩾4 | 9–22 | (27) | ||||
| CD4–MHC class II | − | − | ⩾3 | (74, 75) | ||||
| CD28–CD80 | ⩾660,000 | ⩾1.6 | 4 | Present study |
K d values shown were obtained by equilibrium binding, but the range of K d values shown for TCR interactions includes some values calculated from the koff and kon.
These observations suggest that within the contact area between T cells and APC or target cells (18, 58), TCR/ peptide–MHC complexes will generally be longer lived than CD28–CD80 or CD2–CD58 complexes. This raises the question as to whether the rapid binding kinetics of CD2 and CD28 interactions are functionally significant. One crucial feature of T cell antigen recognition is the capacity of T cells to detect as few as 10 specific peptide–MHC ligands on cells (59–61). Furthermore, Valitutti et al. (62) recently reported that, even when presented with very low levels of peptide–MHC (∼100 complexes per cell), a substantial proportion of TCRs on the T cell surface (∼18,000) appeared to be ligated in the process of T cell activation. Two important implications of these results are the following: (a) many peptide–MHC molecules will need to diffuse through the zone of contact between a T cell and an APC for the TCR to encounter specific peptide–MHC molecules; and (b) each specific peptide–MHC molecule will need to engage serially many TCRs (62). In turn, this implies that peptide–MHC and TCR molecules will need to diffuse rapidly into and out of a stable zone of contact between the T cells and APC and/or that new contacts will continually need to be formed between these cells (63). It seems likely that the very fast binding kinetics of CD2 and CD28 interactions will facilitate these processes by contributing to a highly dynamic contact zone between T cells and APC.
Since CD28 and CTLA-4 are clearly signaling molecules, able, like the TCR, to transmit positive and negative cells to T cells, the very fast k off values reported in the present study indicate that signaling between cells can be transmitted by very transient molecular interactions. Because in the case of the TCR, the nature of the signal (positive or negative) appears to be highly dependent on the kinetics of its interaction (22), it is possible that signaling through CD28 and CTLA-4 will be similarly sensitive to binding kinetics. Preliminary data suggest that CTLA-4 binds CD86 with a lower affinity and faster dissociation rate constant than CD80 (52), which may contribute to the reported functional differences between CD80 and CD86 (6).
In this study, we show that CD28 and CTLA-4 bind CD80 with far lower affinities than previously reported and that these low affinities are the result of very fast dissociation rate constants. These kinetic parameters are similar to those measured for CD2–ligand interactions but very different to reported values for TCR–ligand interactions. We propose that the fast binding kinetics of CD28 (and CD2) serve to optimize T cell antigen recognition by (a) allowing rapid formation and disruption of contacts between T cells and APC and (b) facilitating lateral diffusion of TCRs and peptide–MHC molecules through T cell–APC contact zones.
Footnotes
We gratefully acknowledge Neil Barclay, Marion Brown, and Don Mason for providing valuable advice and critically reading this manuscript; Liz Davies for constructing the sCD80his-encoding plasmid; Marion Brown and Fabiana Horn for expression and purification of CTLA-4 Ig; and Tony Willis for performing the amino acid analyses. We thank Drs. D. Olive, R.A.W. van Lier, and K. Sagawa for kindly providing us with monoclonal antibodies listed in Table 1.
This work was supported by the United Kingdom Medical Research Council and the Wellcome Trust. D.L. Bodian was supported by the Cancer Research Fund of the Damon Runyon–Walter Winchell Foundation, Fellowship DRG-1246.
1 Abbreviations used in this paper: CHO, Chinese hamster ovary; FC, flow cell; k off, dissociation rate constant; k on, association rate constant; RU, response unit; sCD80his, soluble CD80 with carboxy-terminal oligo-histidine tag; SPR, surface plasmon resonance.
References
- 1.Hansen JA, Martin PJ, Nowinski RC. Monoclonal antibodies identifying a novel T-cell antigen and Ia antigen of human lymphocytes. Immunogenetics. 1980;10:247–260. [Google Scholar]
- 2.Aruffo A, Seed B. Molecular cloning of a CD28 cDNA by a high-efficiency COS cell expression system. Proc Natl Acad Sci USA. 1987;84:8573–8577. doi: 10.1073/pnas.84.23.8573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Brunet JF, Denizot F, Luciani MF, Roux-Dosseto M, Suzan M, Mattei MG, Golstein P. A new member of the immunoglobulin superfamily-CTLA-4. Nature (Lond) 1987;328:267–270. doi: 10.1038/328267a0. [DOI] [PubMed] [Google Scholar]
- 4.Linsley PS, Ledbetter JA. The role of the CD28 receptor during T cell responses to antigen. Annu Rev Immunol. 1993;11:191–211. doi: 10.1146/annurev.iy.11.040193.001203. [DOI] [PubMed] [Google Scholar]
- 5.June CH, Bluestone JA, Nadler LM, Thompson CB. The B7 and CD28 receptor families. Immunol Today. 1994;15:321–331. doi: 10.1016/0167-5699(94)90080-9. [DOI] [PubMed] [Google Scholar]
- 6.Lenschow DJ, Walunas TL, Bluestone JA. CD28/B7 system of T cell costimulation. Annu Rev Immunol. 1996;14:233–258. doi: 10.1146/annurev.immunol.14.1.233. [DOI] [PubMed] [Google Scholar]
- 7.Linsley PS, Clark EA, Ledbetter JA. T-cell antigen CD28 mediates adhesion with B cells by interacting with activation antigen B7/BB-1. Proc Natl Acad Sci USA. 1990;87:5031–5035. doi: 10.1073/pnas.87.13.5031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Linsley PS, Brady W, Urnes M, Grosmaire LS, Damle NK, Ledbetter JA. CTLA-4 is a second receptor for the B cell activation antigen B7. J Exp Med. 1991;174:561–569. doi: 10.1084/jem.174.3.561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Azumo M, Ito D, Yagita H, Okumura K, Phillips JH, Lanier LL, Somoza C. B70 antigen is a second ligand for CTLA-4 and CD28. Nature (Lond) 1993;366:76–79. doi: 10.1038/366076a0. [DOI] [PubMed] [Google Scholar]
- 10.Freeman GJ, Gribben JG, Boussiotis VA, Ng JW, Restivo VA, Jr, Lombard LA, Gray GS, Nadler LM. Cloning of B7-2: a CTLA-4 counter-receptor that costimulates human T cell proliferation. Science (Wash DC) 1993;262:909–911. doi: 10.1126/science.7694363. [DOI] [PubMed] [Google Scholar]
- 11.Freeman GJ, Borriello F, Hodes RJ, Reiser H, Hathcock KS, Laszlo G, McKnight AJ, Kim J, Du L, Lombard DB, et al. Uncovering of functional alternative CTLA-4 counter-receptor in B7-deficient mice. Science (Wash DC) 1993;262:907–909. doi: 10.1126/science.7694362. [DOI] [PubMed] [Google Scholar]
- 12.Hathcock KS, Laszlo G, Dickler HB, Bradshaw J, Linsley P, Hodes RJ. Identification of an alternative CTLA-4 ligand costimulatory for T cell activation. Science (Wash DC) 1993;262:905–907. doi: 10.1126/science.7694361. [DOI] [PubMed] [Google Scholar]
- 13.Linsley PS. Distinct roles for CD28 and cytotoxic T lymphocyte–associated molecule-4 receptors during T cell activation? . J Exp Med. 1995;182:289–292. doi: 10.1084/jem.182.2.289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kearney ER, Walunas TL, Karr RW, Morton PA, Loh DY, Bluestone JA, Jenkins MK. Antigendependent clonal expansion of a trace population of antigen-specific CD4+ T cells in vivo is dependent on CD28 costimulation and inhibited by CTLA-4. J Immunol. 1995;155:1032–1036. [PubMed] [Google Scholar]
- 15.Tivol EA, Borriello F, Schweitzer AN, Lynch WP, Bluestone JA, Sharpe AH. Loss of CTLA-4 leads to massive lymphoproliferation and fatal multiorgan tissue destruction, revealing a critical negative regulatory role of CTLA-4. Immunity. 1995;3:541–547. doi: 10.1016/1074-7613(95)90125-6. [DOI] [PubMed] [Google Scholar]
- 16.Waterhouse P, Penninger JM, Timms E, Wakeham A, Shahinian A, Lee KP, Thompson CB, Griesser H, Mak TW. Lymphoproliferative disorders with early lethality in mice deficient in CTLA-4. Science (Wash DC) 1995;270:985–988. doi: 10.1126/science.270.5238.985. [DOI] [PubMed] [Google Scholar]
- 17.Marengére LEM, Waterhouse HW, Duncan GS, Mittrücker HW, Feng GS, Mak TW. Regulation of T cell receptor signalling by tyrosine phosphatase SYP association with CTLA-4. Science (Wash DC) 1996;272:1170–1173. doi: 10.1126/science.272.5265.1170. [DOI] [PubMed] [Google Scholar]
- 18.Springer TA. Adhesion receptors of the immune system. Nature (Lond) 1990;346:425–434. doi: 10.1038/346425a0. [DOI] [PubMed] [Google Scholar]
- 19.Clark EA, Ledbetter JA. How T and B cells talk to each other. Nature (Lond) 1994;367:425–428. doi: 10.1038/367425a0. [DOI] [PubMed] [Google Scholar]
- 20.Jameson SC, Bevan MJ. T cell receptor antagonists and partial agonists. Immunity. 1995;2:1–11. doi: 10.1016/1074-7613(95)90074-8. [DOI] [PubMed] [Google Scholar]
- 21.Sykulev Y, Cohen RJ, Eisen HN. The law of mass action governs antigen-stimulated cytolytic activity of CD8+ cytotoxic T lymphocytes. Proc Natl Acad Sci USA. 1995;92:11990–11992. doi: 10.1073/pnas.92.26.11990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Alam SM, Travers PJ, Wung JL, Nasholds W, Redpath S, Jameson SC, Gascoigne NRJ. T-cell-receptor affinity and thymocyte positive selection. Nature (Lond) 1996;381:616–620. doi: 10.1038/381616a0. [DOI] [PubMed] [Google Scholar]
- 23.Valitutti S, Müller S, Dessing M, Lanzavecchia A. Different responses are elicited in cytotoxic T lymphocytes by different levels of T cell receptor occupancy. J Exp Med. 1996;183:1917–1921. doi: 10.1084/jem.183.4.1917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Viola A, Lanzavecchia A. T cell activation determined by T cell receptor number and tunable thresholds. Science (Wash DC) 1996;273:104–106. doi: 10.1126/science.273.5271.104. [DOI] [PubMed] [Google Scholar]
- 25.Fremont DH, Rees WA, Kozono H. Biophysical studies of T-cell receptors and their ligands. Curr Opin Immunol. 1996;8:93–100. doi: 10.1016/s0952-7915(96)80111-7. [DOI] [PubMed] [Google Scholar]
- 26.van der Merwe PA, Brown MH, Davis SJ, Barclay AN. Affinity and kinetic analysis of the interaction of the cell-adhesion molecules rat CD2 and CD48. EMBO (Eur Mol Biol Organ) J. 1993;12:4945–4954. doi: 10.1002/j.1460-2075.1993.tb06188.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.van der Merwe PA, Barclay AN, Mason DW, Davies EA, Morgan BP, Tone M, Krishnam AKC, Ianelli C, Davis SJ. The human cell-adhesion molecule CD2 binds CD58 with a very low affinity and an extremely fast dissociation rate but does not bind CD48 or CD59. Biochemistry. 1994;33:10149–10160. doi: 10.1021/bi00199a043. [DOI] [PubMed] [Google Scholar]
- 28.Dustin ML, Ferguson LM, Chan P, Springer TA, Golan DE. Visualization of the CD2 interaction with LFA-3 and determination of the two-dimensional dissociation constant for adhesion receptors in a contact area. J Cell Biol. 1996;132:465–474. doi: 10.1083/jcb.132.3.465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Linsley PS, Brady W, Grosmaire L, Aruffo A, Damle NK, Ledbetter JA. Binding of the B cell activation antigen B7 to CD28 costimulates T cell proliferation and interleukin 2 mRNA accumulation. J Exp Med. 1991;173:721–730. doi: 10.1084/jem.173.3.721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Linsley PS, Greene JL, Brady W, Bajorath J, Ledbetter JA, Peach R. Human B7-1 (CD80) and B7-2 (CD86) bind with similar avidities but distinct kinetics to CD28 and CTLA-4 receptors. Immunity. 1994;1:793–801. doi: 10.1016/s1074-7613(94)80021-9. [DOI] [PubMed] [Google Scholar]
- 31.Morton PA, Fu XT, Stewart JA, Giacoletto KS, White SL, Leysath CE, Evans RJ, Shieh JJ, Karr RW. Differential effects of CTLA-4 substitutions on binding of human CD80 (B7-1) and CD86 (B7-2) J Immunol. 1996;156:1047–1054. [PubMed] [Google Scholar]
- 32.van der Merwe PA, Barclay AN. Transient intercellular adhesion: the importance of weak protein–protein interactions. Trends Biochem Sci. 1994;19:354–358. doi: 10.1016/0968-0004(94)90109-0. [DOI] [PubMed] [Google Scholar]
- 33.van der Merwe PA, Barclay AN. Analysis of cell adhesion molecule interactions using surface plasmon resonance. Curr Opin Immunol. 1996;8:257–261. doi: 10.1016/s0952-7915(96)80065-3. [DOI] [PubMed] [Google Scholar]
- 34.Margulies DH, Plaksin D, Khilko SN, Jelonek MT. Studying interactions involving the T-cell antigen receptor by surface plasmon resonance. Curr Opin Immunol. 1996;8:262–270. doi: 10.1016/s0952-7915(96)80066-5. [DOI] [PubMed] [Google Scholar]
- 35.Jönsson U, Fägerstam L, Ivarsson B, Johnsson B, Karlsson R, Lundh K, Löfås S, Persson B, Roos H, Rönnberg I, et al. Real-time biospecific interaction analysis using surface plasmon resonance and sensor chip technology. BioTechniques. 1991;11:620–627. [PubMed] [Google Scholar]
- 36.Malik N, Kallestad JC, Gunderson NL, Austin SD, Neubauer MG, Ochs V, Marquardt H, Zarling JM, Shoyab M, Wei C, et al. Molecular cloning, sequence analysis, and functional expression of a novel growth regulator, oncostatin M. Mol Cell Biol. 1989;9:2847–2853. doi: 10.1128/mcb.9.7.2847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Simmons, D.L. 1993. Cloning cell surface molecules by transient expression in mammalian cells. In Cellular interactions in development: a practical approach. D.A. Hartley, editor. Oxford, Oxford University Press. 93–127.
- 38.Davis SJ, Ward HA, Puklavec MJ, Willis AC, Williams AF, Barclay AN. High level expression in Chinese hamster ovary cells of soluble forms of CD4 T lymphocyte glycoprotein including glycosylation variants. J Biol Chem. 1990;265:10410–10418. [PubMed] [Google Scholar]
- 39.Bebbington, C.R. 1991. Expression of antibody genes in nonlymphoid mammalian cells. Methods Enzymol. 2(Suppl): 136–145.
- 40.Brown MH, Preston S, Barclay AN. A sensitive assay for detecting low affinity interactions at the cell surface reveals no additional ligands for the adhesion pair rat CD2 and CD48. Eur J Immunol. 1995;25:3222–3228. doi: 10.1002/eji.1830251204. [DOI] [PubMed] [Google Scholar]
- 41.Clark SJ, Jefferies WA, Barclay AN, Gagnon J, Williams AF. Peptide and nucleotide sequences of rat CD4 (W3/25) antigen: evidence for derivation from a structure with four immunoglobulin-related domains. Proc Natl Acad Sci USA. 1987;84:1649–1653. doi: 10.1073/pnas.84.6.1649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kelm S, Pelz A, Schauer R, Filbin MT, Tang S, de Bellard M, Schnaar RL, Mahoney JA, Hartnell A, Bradfield P, Crocker PR. Sialoadhesin, myelin-associated glycoprotein and CD22 define a new family of sialic acid–dependent adhesion molecules of the immunoglobulin superfamily. Curr Biol. 1994;4:965–972. doi: 10.1016/s0960-9822(00)00220-7. [DOI] [PubMed] [Google Scholar]
- 43.Karlsson R, Michaelsson A, Mattsson L. Kinetic analysis of monoclonal antibody–antigen interactions with a new biosensor based analytical system. J Immunol Methods. 1991;145:229–240. doi: 10.1016/0022-1759(91)90331-9. [DOI] [PubMed] [Google Scholar]
- 44.Nath D, van der Merwe PA, Kelm S, Bradfield P, Crocker PR. The amino-terminal immunoglobulinlike domain of sialoadhesin contains the sialic acid binding site: comparison with CD22. J Biol Chem. 1995;270:26184–26191. doi: 10.1074/jbc.270.44.26184. [DOI] [PubMed] [Google Scholar]
- 45.Freeman GJ, Freedman AS, Segil JM, Lee G, Whitman JF, Nadler LM. B7, a new member of the Ig superfamily with unique expression on activated and neoplastic B cells. J Immunol. 1989;143:2714–2722. [PubMed] [Google Scholar]
- 46.Gray F, Cyster JG, Willis AC, Barclay AN, Williams AF. Structural analysis of the CD2 T lymphocyte antigen by site-directed mutagenesis to introduce a disulphide bond into domain 1. Prot Engin. 1993;6:965–970. doi: 10.1093/protein/6.8.965. [DOI] [PubMed] [Google Scholar]
- 47.Brown MH, Barclay AN. Expression of immunoglobulin and scavenger receptor superfamily domains as chimeric proteins with domains 3 and 4 of CD4 for ligand analysis. Prot Engin. 1994;7:515–521. doi: 10.1093/protein/7.4.515. [DOI] [PubMed] [Google Scholar]
- 48.O'Shannessy DJ. Determination of kinetic rate and equilibrium binding constants for macromolecular interactions: a critique of the surface plasmon resonance literature. Curr Opin Biotechnol. 1994;5:65–71. doi: 10.1016/s0958-1669(05)80072-2. [DOI] [PubMed] [Google Scholar]
- 49.Nieba L, Krebber A, Plückthun A. Competition BIAcore for measuring true affinities: large differences in values determined from binding kinetics. Anal Biochem. 1996;234:155–165. doi: 10.1006/abio.1996.0067. [DOI] [PubMed] [Google Scholar]
- 50.Schuck P. Kinetics of ligand-binding to receptor immobilized in a polymer matrix, as detected with an evanescent wave biosensor. 1. A computer simulation of the influence of mass-transport. Biophysical J. 1996;70:1230–1249. doi: 10.1016/S0006-3495(96)79681-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Linsley PS, Nadler SG, Bajorath J, Peach R, Leung HT, Rogers J, Bradshaw J, Stebbins M, Leytze G, Brady W, et al. Binding stoichiometry of cytotoxic T lymphocyte–associated molecule-4 (CTLA-4). A disulphidelinked homodimer binds two CD86 molecules. J Biol Chem. 1995;270:15417–15424. doi: 10.1074/jbc.270.25.15417. [DOI] [PubMed] [Google Scholar]
- 52.Greene JL, Leytze GM, Emswiler J, Peach R, Bajorath J, Cosand W, Linsley PS. Covalent dimerization of CD28/CTLA-4 and oligomerization of CD80/ CD86 regulate T cell costimulatory interactions. J Biol Chem. 1996;271:26762–26771. doi: 10.1074/jbc.271.43.26762. [DOI] [PubMed] [Google Scholar]
- 53.Bell GI. Models for specific adhesion of cells to cells. Science (Wash DC) 1978;200:618–627. doi: 10.1126/science.347575. [DOI] [PubMed] [Google Scholar]
- 54.Linsley PS, Greene JL, Tan P, Bradshaw J, Ledbetter JA, Anasetti C, Damle NK. Coexpression and functional cooperation of CTLA-4 and CD28 on activated T lymphocytes. J Exp Med. 1992;176:1595–1604. doi: 10.1084/jem.176.6.1595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Krummel MF, Allison JP. CD28 and CTLA-4 have opposing effects on the response of T cells to stimulation. J Exp Med. 1995;182:459–465. doi: 10.1084/jem.182.2.459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Walunas TL, Bakker CY, Bluestone JA. CTLA-4 ligation blocks CD28-dependent T cell activation. J Exp Med. 1996;183:2541–2550. doi: 10.1084/jem.183.6.2541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Luescher IF, Vivier E, Layer A, Mahiou J, Godeau F, Malissen B, Romero P. CD8 modulation of T-cell antigen receptor–ligand interactions on living cytotoxic T lymphocytes. Nature (Lond) 1995;373:353–356. doi: 10.1038/373353a0. [DOI] [PubMed] [Google Scholar]
- 58.Davis SJ, van der Merwe PA. Structure and ligand interactions of CD2: implications for T-cell function. Immunol Today. 1996;17:177–187. doi: 10.1016/0167-5699(96)80617-7. [DOI] [PubMed] [Google Scholar]
- 59.Demotz S, Grey HM, Sette A. The minimal number of class II MHC–antigen complexes needed for T cell activation. Science (Wash DC) 1990;249:1028–1030. doi: 10.1126/science.2118680. [DOI] [PubMed] [Google Scholar]
- 60.Christinck ER, Luscher MA, Barber BH, Williams DB. Peptide binding to class I MHC on living cells and quantitation of complexes required for CTL lysis. Nature (Lond) 1991;352:67–70. doi: 10.1038/352067a0. [DOI] [PubMed] [Google Scholar]
- 61.Kageyama S, Tsomides TJ, Sykulev Y, Eisen HN. Variation in the number of peptide–MHC class I complexes required to activate cytotoxic T cell responses. J Immunol. 1995;154:567–576. [PubMed] [Google Scholar]
- 62.Valitutti S, Müller S, Cella M, Padovan E, Lanzavecchia A. Serial triggering of many T-cell receptors by a few peptide–MHC complexes. Nature (Lond) 1995;375:148–151. doi: 10.1038/375148a0. [DOI] [PubMed] [Google Scholar]
- 63.Valitutti S, Dessing M, Aktories K, Gallati H, Lanzavecchia A. Sustained signalling leading to T cell activation results from prolonged T cell receptor occupancy. Role of the T cell actin cytoskeleton. J Exp Med. 1995;181:577–584. doi: 10.1084/jem.181.2.577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ghiotto-Ragueneau M, Battifora M, Truneh A, Waterfield MD, Olive D. Comparison of CD28–B7.1 and B7.2 functional interaction in resting human T cells: phosphatidylinositol 3-kinase association to CD28 and cytokine production. Eur J Immunol. 1996;26:34–41. doi: 10.1002/eji.1830260106. [DOI] [PubMed] [Google Scholar]
- 65.Valle A, Garrone P, Bonnefoy JY, Freedman AS, Freeman G, Nadler LM, Banchereau J. mAb 104, a new monoclonal antibody, recognizes the B7 antigen that is expressed on activated B cells and HTLV-1 transformed cells. Immunology. 1990;69:531–535. [PMC free article] [PubMed] [Google Scholar]
- 66.Schlossman, S.F., L. Boumsell, W. Gilks, J.M. Harlan, T. Kishimoto, C. Morimoto, J. Ritz, S. Shaw, R. Silverstein, T.A. Springer, T.F. Tedder, and R.F. Todd, editors. 1995. Leucocyte Typing V: White Cell Differentiation Antigens. Oxford, Oxford University Press.
- 67.Yokochi T, Holly RD, Clark EA. B lymphoblast antigen (BB-1) expressed on Epstein–Barr virus–activated B cell blasts, B lymphoblastoid cell lines, and Burkitt's lymphoma. J Immunol. 1982;128:823–827. [PubMed] [Google Scholar]
- 68.Nunés J, Klasen S, Ragueneau M, Pavon C, Couez D, Mawas C, Bagnasco M, Olive D. CD28 mAbs with distinct binding properties differ in their ability to induce T cell activation: analysis of early and late activation events. Int Immunol. 1993;5:311–316. doi: 10.1093/intimm/5.3.311. [DOI] [PubMed] [Google Scholar]
- 69.Okubo K, Hagiwara S, Shiotsuki K, Sagawa K, Yokoyama MM. A new monoclonal antibody (KOLT-2) specific to the cell surface antigen on activated human T lymphocytes. Acta Haematol Jpn. 1984;47:1433–1444. [PubMed] [Google Scholar]
- 70.Matsui K, Boniface JJ, Reay PA, Schild H, Fazekas de St B, Groth, Davis MM. Low affinity interaction of peptide-MHC complexes with T cell receptors. Science (Wash DC) 1991;254:1788–1791. doi: 10.1126/science.1763329. [DOI] [PubMed] [Google Scholar]
- 71.Matsui K, Boniface JJ, Steffner P, Reay PA, Davis MM. Kinetics of T-cell receptor binding to peptide/I-Ek complexes: correlation of the dissociation rate with T-cell responsiveness. Proc Natl Acad Sci USA. 1994;91:12862–12866. doi: 10.1073/pnas.91.26.12862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sykulev Y, Brunmark A, Jackson M, Cohen RJ, Peterson PA, Eisen HN. Kinetics and affinity of reactions between an antigen-specific T cell receptor and peptide–MHC complexes. Immunity. 1994;1:15–22. doi: 10.1016/1074-7613(94)90005-1. [DOI] [PubMed] [Google Scholar]
- 73.Corr M, Slanetz AE, Boyd LF, Jelonek MT, Khilko S, Al-Ramadi BK, Kim YS, Maher SE, Bothwell ALM, Margulies DH. T cell receptor–MHC class I peptide interactions: affinity, kinetics, and specificity. Science (Wash DC) 1994;265:946–949. doi: 10.1126/science.8052850. [DOI] [PubMed] [Google Scholar]
- 74.Cammarota G, Scheirle A, Takacs B, Doran DM, Knorr R, Bannwarth W, Guardiola J, Sinigaglia F. Identification of the CD4 binding site on the β2 domain of HLA-DR molecules. Nature (Lond) 1992;356:799–801. doi: 10.1038/356799a0. [DOI] [PubMed] [Google Scholar]
- 75.Weber S, Karjalainen K. Mouse CD4 binds MHC class II with extremely low affinity. Int Immunol. 1993;5:695–698. doi: 10.1093/intimm/5.6.695. [DOI] [PubMed] [Google Scholar]