

Abstract
A subset of CD4+CD11c−CD3− blood cells was recently shown to develop into dendritic cells when cultured with monocyte conditioned medium. Here, we demonstrate that CD4+ CD11c−CD3− cells, isolated from tonsils, correspond to the so-called plasmacytoid T cells, an obscure cell type that has long been observed by pathologists within secondary lymphoid tissues. They express CD45RA, but not markers specific for known lymphoid- or myeloid-derived cell types. They undergo rapid apoptosis in culture, unless rescued by IL-3. Further addition of CD40-ligand results in their differentiation into dendritic cells that express low levels of myeloid antigens CD13 and CD33.
Band T lymphocytes share many similarities, including their developmental programmes, the structures of their antigen receptors, and the requirement for costimulatory molecules for activation. B lymphocytes will finally differentiate into plasma cells that secrete antibody, the key effector molecule of humoral immune responses. Interestingly, pathologists have long observed the presence of plasma cells that express the T cell marker CD4 and lack intracellular immunoglobulins within the T cell zones of human lymph nodes (1–3). These plasma cells were suggested to represent terminally differentiated T plasma cells of unknown secretory functions. Paradoxically, these so called plasmacytoid T cells were found not to express other T cell markers such as CD3 and TCRs (4, 5). They expressed MHC class II antigens, HLADR and HLADQ, and the invariant chain CD74 (6, 7).
The CD4+ plasmacytoid cells were also found in human spleen and thymus from patients with myasthenia gravis (5). These cells were found in increased number in lymph nodes taken from patients suffering from lymphomas, leukemias (8), patients with breast cancer metastases (4), and lymphadenitis (5). In addition, cases of plasmacytoid T cell lymphomas were reported that paradoxically evolved towards myelomonocytic leukemia (9). These data suggest that the CD4+ plasmacytoid cells represent a neglected but important cell type of the immune system. However, the nature and the fate of these plasmacytoid cells has remained unknown.
Here, we report the isolation of plasmacytoid cells from human tonsils. The phenotypical, anatomical and functional characterization indicates that these cells correspond to the CD4+CD11c− blood dendritic cell (DC)1 presursors, that either undergo rapid apoptosis or differentiate into DC upon culture with IL-3 and CD40-ligand.
Materials and Methods
Immunohistological Localization of CD4+CD11c−CD3− Cells.
Double stainings on human tonsil sections were performed using mouse IgG1 anti-CD3 (Immunotech, Marseille, France) and antiCD11c (Dako, Glostrup, Denmark) together with mouse IgG2a anti-CD4 (Innotest, Besancon, France). The binding of mouse IgG1 antibodies was revealed by sheep anti–mouse IgG1 (The Binding Site, Birmingham, UK), followed by mouse anti-alkaline phosphatase-alkaline phosphatase complexes (Dako), the APAAP technique. The binding of mouse IgG2a antibodies was revealed by sheep anti–mouse IgG2a-biotin (The Binding Site), followed by ExtrAvidin-peroxidase (Sigma Chem. Co., St. Louis, MO). Alkaline phosphatase activity was developed by the Fast Blue substrate, whereas peroxidase activity was developed by 3-aminoethylcarbazole.
Purification of CD4+CD11c− Lin− Cells.
CD4+CD11c− cells were isolated from human tonsils. In brief, tonsils were cut into small pieces and digested for 12 min at 37°C with collagenase IV (1 mg/ml; Sigma) and deoxyribonuclease I (50 KU/ml, Sigma) in RPMI 1640. The cells, pooled from two rounds of tissue digestion, were centrifuged over 50% Percoll (Pharmacia, Uppsala, Sweden) for 20 min at 400 g. CD3+ T cells, CD14+ monocytes, CD19+ and CD20+ B cells, and CD56+ NK cells were depleted from the resulting low density cells by immunomagnetic beads (sheep anti–mouse Ig–coated Dynabeads; Dynal, Oslo, Norway). The resulting cells were stained with mouse anti–CD4-PE-Cy5 (Immunotech), anti–CD11c-PE (Becton Dickinson), and a cocktail of FITC-labeled mAbs anti-CD3 and anti-CD34 (Immunotech), anti-CD20, anti-CD57, anti-CD7, anti-CD14, and antiCD16 (Becton Dickinson), and anti-CD1a (Ortho). Then CD4+ CD11c− Lin− cells were isolated by cell sorting. Reanalysis of the sorted cells confirmed a purity of 98%.
Purification of Naive T Cells from Adult Blood.
CD4+ CD45RA+ T cells were purified from blood mononuclear cells by immunomagnetic bead depletion using a cocktail of mAbs IOM2 (CD14), ION16 (CD16), ION2 (HLA DR) (Immunotech), NKH1 (CD56), OKT8 (CD8) (Ortho), 4G7 (CD19), UCHL1 (CD45RO), and mAb 89 (CD40). After three rounds of bead depletion, purity of CD4+ T cells was routinely ⩾96%.
FACS® Analysis of CD4+CD11c− Cocktail FITC−-sorted Cells.
FITC-labeled mAbs for surface phenotyping of sorted (or cultured) cells were purchased from Immunotech (CD13, CD33, CD34, CD8, CD11b, CD21, CD18, CD54, CD58, and CD38), Becton-Dickinson (CD14, CD2, TCR αβ, MHC class II DR and DQ, CD16, and CD71), Coulter (TCR-γδ), Sigma (CD45, CD44), Dako (CD45RO, CD23, CD11a, and CD11c), PharMingen (CD35), Medarex (CD32). PE-labeled mAbs were purchased from Becton Dickinson (CD80), Immunotech (CD40), and PharMingen (CD86). Cells, incubated with antibody for 15 min at 4°C, were analyzed after one wash with a FACScan® flow cytometer. Negative controls were performed with unrelated murine mAbs (Dako). These negative controls are indicated by filled histograms.
Proliferation Assays and DC Generation in Culture.
CD4+ CD11c− Lin− cells were cultured with IL-3 in the presence or absence of CD40L fibroblasts (10). All cultures were performed in RPMI 1640 supplemented with 10% of FCS, 2 mM l-glutamine and antibiotics. Cells (1.5 × 104) were seeded in 96-well flat-bottom microtiter plates for the DNA synthesis assay. After 3 d, cells were pulsed with 1 μCi [3H]thymidine for 8 h before harvesting and counting. Tests were carried out in triplicate and standard deviations are indicated with bars. For viable cell recovery estimation, cells were counted by Trypan blue dye exclusion. rhIL-3 or GMCSF (Schering-Plough Research Institute, Kenilworth, NJ) were used at a saturating concentration of 10 ng/ml (50 U/ml) and 100 ng/ml (200 U/ml), respectively. rhTNFα (Genzyme, Boston, MA) was used at 2.5 ng/ml (50 U/ml). For phenotypic studies, 3 × 105 to 5 × 105 cells were cultured in 48-well flat-bottom plates (5 CD40L fibroblasts for 1 CD4+CD11c− cocktail FITC− cell).
T Cell Proliferation Assay.
DC (10 to 5,000) were cultured with 5 × 104 allogeneic blood CD4+CD45RA+ T cells in round-bottomed 96-well culture plates in RPMI 1640 containing 10% human AB+ serum. The DC used were generated by culture of (a) CD4+CD11c−Lin− cells for 6 d in the presence of IL-3 with or without CD40L transfected fibroblasts; (b) blood monocytes for 10 d with GM-CSF plus IL-4 (50 U/ml) (11, 12); and (c) CD34+ cells for 12 d with GM-CSF and TNFα (13). After 5 d of DC-T cell cocultures, cells were pulsed with 1 μCi [3H]thymidine for 8 h before harvesting and counting. Tests were carried out in triplicate and standard deviations are indicated with bars.
Electron Microscopic Analysis of CD4+CD11c−Lin− Cells.
For electron microscopy, CD4+CD11c−Lin− cells (freshly isolated or cultured cells) were fixed in 2.5% glutaraldehyde plus 0.8% paraformaldehyde in 0.1 M cacodylate buffer and postfixed with an aqueous solution of 1% OsO4 containing K4Fe(CN)6. For scanning electron microscopy (SEM), cells were allowed to settle on coverslips precoated with poly-l-lysine, dehydrated in an acetone series, dried by the critical point method (CO2), and coated with 10 nm gold in sputter device. Observations were made with a SEM 505 instrument (Philips, Eindhoven, Netherlands). For transmission electron microscopy (TEM), the cells were dehydrated in an alcohol series and embedded into epon. Ultrathin sections (∼50 nm) were stained with lead citrate and uranyl acetate and studied with a CM 100 instrument (Philips).
Results
Localization and Isolation of Tonsillar Plasmacytoid T Cells.
In previous studies, plasmacytoid T cells in situ were shown to express CD4 but to lack lineage markers specific for B cells, T cells, NK cells, monocytes, Langerhans cells, and granulocytes (1–7). They expressed CD11a but not CD11b or CD11c (5). To determine if human tonsils contained CD4+ plasmacytoid T cells, double staining with anti-CD3 (blue) plus anti-CD11c (blue) together with anti-CD4 (red) was performed on tonsil sections. Accordingly, whereas CD4+CD3+ helper T cells, CD4low CD11c+ interdigitating cells and CD4+CD11c+ dendritic cells within germinal centers (GCDC) (14) should be double stained purple, CD4+CD11c−CD3− plasmacytoid T cells should be single stained red (Fig. 1). Indeed, red CD4+ CD11c−CD3− cells were found in the T cell–rich extrafollicular areas (Fig. 1, A, C, D, E) but not within germinal centers (Fig. 1, A and B), confirming the previous observations in lymph nodes (5). Interestingly, many red CD4+ CD11c−CD3− cells were found to be close to or within the high endothelial veinules (HEV) (Fig. 1, C, D, E), suggesting that these cells might be blood derived cells that were crossing the HEV. The CD4+CD11c−CD3− cells within the T cell rich areas indeed displayed a plasmacytoid morphology (Fig. 1, C, D, E).
Figure 1.
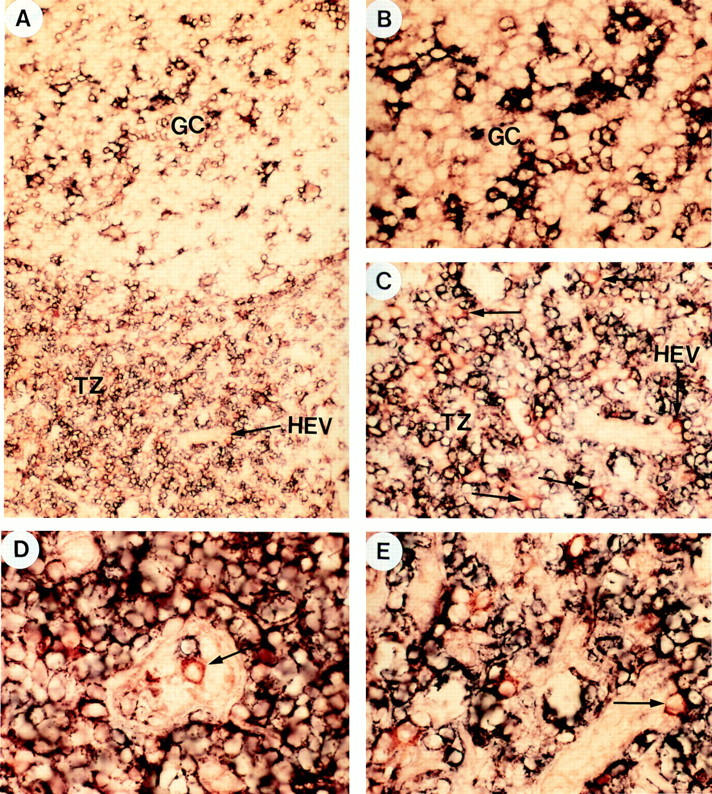
Anatomical localization of CD4+CD11c−CD3− plasmacytoid cells. To distinguish CD4+CD11c−CD3− plasmacytoid cells from CD4+CD3+ T cells and CD4+CD11c+ germinal center dendritic cells, double staining with anti-CD3 (blue), anti-CD11c (blue), and anti-CD4 (red) on tonsillar sections was performed. Accordingly, whereas CD4+CD3+ T cells and CD4+CD11c+ germinal center dendritic cells should be double stained purple, CD4+CD11c−CD3− plasmacytoid cells should be single stained red. (A) shows a germinal center and its adjacent T cell–rich extrafollicular areas containing HEV. Red CD4+CD11c−CD3− plasmacytoid cells can only be found in T cell area but not in germinal center. (B) shows that the same germinal center displayed in A does not contain red CD4+CD11c−CD3− plasmacytoid cells. (C) shows that the same T cell area displayed in A contains many red CD4+CD11c−CD3− plasmacytoid cells, located within or around HEV. (D) shows a red CD4+ CD11c−CD3− plasmacytoid cell within the lumen of HEV. (E) shows many red CD4+CD11c−CD3− plasmacytoid cells, one in particular is within the endothelial wall of HEV. Original magnification: (A) ×100; (B, C) ×200; (D, E) ×400.
According to the in situ phenotype of plasmacytoid T cells, a multistep isolation method was developed including: (a) centrifugation >50% Percoll; (b) magnetic bead depletion of B cells (CD19+CD20+), T cells (CD3+), monocytes (CD14+) and NK cells (CD56+); (c) finally sorting for CD4+CD11c− cells that were negative for CD7, CD57, CD20, CD3, CD14, CD1a (Langerhans cells), CD34, and CD16 (Fig. 2). Two CD4+Lin− populations were identified (Fig. 3 A). The CD4+CD11c+ population that represents GCDCs has been described earlier (14). On Giemsa staining, freshly sorted CD4+CD11c− cells display a plasma cell-like morphology, characterized by an eccentric nucleus, a blue basophilic cytoplasm and pale Golgi zone (Fig. 3 A). By scanning electron microscopy, these cells display a smooth round lymphoid morphology and are 8–10 μm in diameter (Fig. 3 B). By transmission EM, the cells display nuclei with marginal heterochromatin and cytoplasm containing juxtanuclear Golgi apparatus and parallel arrays of rough endoplasmic reticulum (RER) (Fig. 3, C and D). However, unlike immunoglobulin containing CD38++ CD20− plasma cells isolated from human tonsils (Fig. 3 F), these CD4+CD11c−Lin− plasmacytoid cells did not contain intracellular immunoglobulin (Fig. 3 E).
Figure 2.

Cell sorting of CD4+CD11c−CD3− plasmacytoid cells. After preparation of tonsillar cells that are negative for CD3, CD14, CD19, CD20, and CD56 (detailed in Materials and Methods), cells were stained with mouse anti–CD4-PE-Cy5 (Immunotech), anti–CD11c-PE (Becton Dickinson), and a cocktail of FITC-labeled mAbs anti-CD3 and CD34 (Immunotech), anti-CD20, anti-CD57, anti-CD7 and anti-CD16 (Becton Dickinson), and anti-CD1a (Ortho). As shown in the left panel, cells were first selected by their cocktail FITC− (Lin−). This selected population contains CD4+CD11c+ germinal center dendritic cells (14) and CD4+ CD11c− plasmacytoid cells, as shown in the right panel.
Figure 3.

Morphology of freshly isolated CD4+CD11c−CD3− plasmacytoid cells. (A) Giemsa staining; (B) SEM; (C) TEM; (D) TEM. (E) negative antiIg κ+λ light chain staining of CD4+CD11c−Lin− plasmacytoid cells; (F) positive anti-Ig κ+λ light chain staining of CD38++CD20− Ig–secreting plasma cells. Original magnification: (A, F) ×1,000; (B) ×3,500; (C) ×2,000; (D) ×8,000.
The Phenotype of Isolated CD4+CD11c−Lin− Tonsillar Plasmacytoid Cells.
Next, the surface phenotype of isolated plasmacytoid cells was analysed by flow cytometry (Fig. 4). Although CD4+CD11c− cells express CD45RA, indicating their hematopoietic origin, they lack all the lineage markers for B cells (CD19, CD21), T cells (CD2, CD3, TCR, CD8), natural killer cells (CD56 and CD57), myeloid cells (CD13 and CD33), monocytes (CD14 and CD11b), granulocytes (CD16 and CD35), and Langerhans cells (CD1a). However, they express MHC class II antigens and the adhesion molecules CD11a, CD18, CD44, and CD54 (but not CD58). They also express low levels of CD40 and CD38. They do not express detectable levels of B7.1/CD80 and B7.2/CD86. This phenotypic profile fits well with that of the in situ plasmacytoid T cells characterized by immunohistochemistry (5). In addition, the phenotype of these tonsillar CD4+CD11c−Lin− plasmacytoid cells is similar to that of the blood CD4+CD11c−Lin− cells, that differentiate into dendritic cells when cultured with monocyte-conditioned medium (15).
Figure 4.

Surface phenotype of isolated CD4+CD11c−CD3− plasmacytoid cells analyzed by flow cytometry. Filled histograms represent isotype matched controls, open histograms represent staining with specific antibodies. Data shown are one representative of six experiments.
Plasmacytoid Cells Die Rapidly by Apoptosis when Cultured in Medium unless Rescued by IL-3.
The nature of these plasmacytoid cells was further analyzed by cell culture with different cytokines. Strikingly, these cells undergo rapid spontaneous apoptosis in culture, resulting in 40% of cells displaying apoptotic figures at 4 h (Fig. 5, B and D) and >90% at 16 h (Fig. 5 F). Among a wide range of tested cytokines (IL-1α, IL-1β, IL-2, IL-3, IL-4, IL-6, IL-7, IL-10, M-CSF, GM-CSF, SCF) and CD40-ligand, IL-3 alone was found to completely block the apoptotic cell death of plasmacytoid cells (Fig. 5, A and C). In contrast to cells cultured in medium alone, cells cultured with IL-3 rapidly formed aggregates within 4 h (Fig. 5 A) and few cells displayed apoptotic figures (Fig. 5 C). During the next 6 d of culture with IL-3, >90% of cells remained viable (Fig. 6 A) and displayed: (a) pseudopods after 16 h of culture (Fig. 5 E); (b) blastic nuclei and mitotic figures after 3 d of culture (Fig. 5 G); and (c) veiled morphology after 6 d of culture (Fig. 5 H). The CD4+ plasmacytoid cells cultured for 3 d in the presence of IL-3 incorporated [3H]thymidine (Fig. 6 B), a finding consistent with the appearance of mitotic figures at this stage. Two cytokine combinations (GM-CSF+TNFα and GM-CSF+IL-4) that, respectively, induce CD34+ progenitor cells (13) and blood monocytes (11) to become DC did not have significant survival (not shown) and proliferation (Fig. 6 B) effects on plasmacytoid cells. CD40-ligand, a key survival and activation signal for several cell types (16–18), including B cells (19, 20) and DC (21), did not interfere with the IL-3–dependent survival of plasmacytoid cells, but inhibited the IL-3–dependent [3H]thymidine uptake (Fig. 6 B).
Figure 5.

CD4+CD11c−CD3− plasmacytoid cells undergo rapid apoptosis when cultured in medium, unless rescued by IL-3. (A) Many cell clusters at 4 h of culture with IL-3; (B) Dispersed cells at 4 h of culture in medium alone; (C) Few apoptotic cells within clusters at 4 h of culture with IL-3; (D) Many apoptotic cells at 4 h of culture in medium alone; (E) Cells display pseudopods at 16 h of culture with IL-3; (F). All cells are dead at 16 h of culture in medium; (G) Mitotic figures at 3 d of culture with IL-3; (H) cells with pseudopods at 6 d of culture with IL-3. Data shown are one representative of six experiments. Original magnification: (B) ×200.
Figure 6.

IL-3 maintains the viability of CD4+CD11c−CD3− plasmacytoid cells for up to 6 d. (A) Total numbers of viable cells at day 1, day 3, and day 6 of culture with medium (open circles), IL-3 (open squares), CD40ligand transfected L cells (open triangles), CD40-ligand negative L cells = ORIENT (solid squares) and CD40-ligand transfected L cells plus IL-3 (solid triangles). (B) IL-3 induces [3H]thymidine incorporation by plasmacytoid cells at day 3 of culture. Data shown are one representative of four experiments.
CD40-ligand Together with IL-3 Promotes the Differentiation of Plasmacytoid Cells into DC, that Express Little CD13, CD33, and CD1a.
As described above, CD40-ligand did not affect the survival of plasmacytoid cells and consistently blocked the [3H]thymidine incorporation induced by IL-3. Most strikingly, CD40-ligand induced the development of long dendrites on these plasmacytoid cells in the presence of IL-3. Fig. 7 A shows that after 3 and 6 d of culture, many clusters of cells with long dendrites appeared. This culture morphology was similar to that of dendritic cells generated from CD34+ progenitor cells cultured with GM- CSF+TNFα (13). Indeed, Giemsa staining of cytospin preparation of plasmacytoid cells cultured for 6 d with IL-3 and CD40-ligand revealed a typical mature dendritic cell morphology (Fig. 7 B). These observations were further confirmed by TEM and SEM (Fig. 7, C and D). The cells cultured with CD40-ligand and IL-3 developed large dendrites (like spines of hedgehog) (Fig. 7 C) and did not have a trace of RER (Fig. 7 D).
Figure 7.

CD4+CD11c−CD3− plasmacytoid cells acquire the morphology of mature interdigitating dendritic cells after 6 d of culture with IL-3 and CD40-ligand. (A) Culture morphology; (B) Giemsa staining; (C) SEM; (D) TEM. Original magnification: (A) ×100; (B) ×400; (C) ×3,000; (D) ×6,000.
The phenotype of plasmacytoid cells cultured for 6 d with IL-3 or with IL-3+CD40-ligand (Fig. 8) was subsequently determined by flow cytometry. With IL-3, the adhesion molecule CD58 and the costimulatory molecules B7.1/CD80 and B7.2/CD86 were induced, and the adhesion molecule CD54, the costimulatory molecule CD40 and MHC class II DR were upregulated. CD40-ligand further upregulated the expression of the above molecules. Antigens specifically expressed by myeloid cells (CD13, CD33, CD11b, CD14, CD35), B cells (CD21), Langerhans cells (CD1a), NK cells (CD16), or activated cells (CD25 and CD45RO) were not induced under these two culture conditions. CD11c, a marker highly expressed by different subsets of DC appeared to be induced on a fraction of cells.
Figure 8.

CD4+CD11c−CD3− plasmacytoid cells express high levels of costimulatory molecules, but few myeloid antigens, after 6 d of culture with IL-3 or IL-3+CD40-ligand. Filled histograms represent isotype matched controls, open histograms represent staining with specific antibodies. Data shown are one representative of six experiments.
Dendritic Cells Generated from CD4+ Plasmacytoid Cells Display Poor Phagocytosis or Endocytosis Capacities but Strongly Stimulate Allogeneic CD4+CD45RA+ Naive T Cells.
To test the functional properties of DCs generated from plasmacytoid cells cultured with IL-3 or IL-3+CD40-ligand, their antigen processing and presentation capacities were analyzed. In contrast to DC generated from monocytes with GM-CSF+IL-4, DC derived from plasmacytoid cells after 6 d of cultures either with IL-3 alone or with IL-3 plus CD40-ligand did not uptake FITC–dextran as analyzed by FACS® (Fig. 9). In addition, they displayed a poor ability to phagocytose FITC-latex beads as analyzed by confocal microscopy and electron microscopy (not shown). Moreover, fresh isolated CD4+ plasmacytoid cells also failed to uptake FITC–dextran during a 40-min culture with FITC– dextran (not shown). However, DC derived from plasmacytoid cells stimulated allogeneic CD4+CD45RA+ naive T cells and were at least as potent as monocyte-derived DC or CD34+ progenitor cell–derived DC (Fig. 10). The cells with pseudopods generated from plasmacytoid cells after 6 d of culture with IL-3 also significantly stimulated the proliferation of CD4+CD45RA+ naive T cells (Fig. 10).
Figure 9.

Dendritic cells derived from CD4+CD11c−CD3− plasmacytoid cells after 6 d of culture with IL-3+CD40-ligand did not uptake FITC-dextran. Methods are as described by Sallusto et al. (11). Time 0 represents cells that were incubated with FITC–dextran for 30 min at 4°C. After three washes, the FITC–dextran uptake by plasmacytoid cells derived DC cultured with IL-3 (open squares) or with IL-3 plus CD40ligand (solid circles) and monocyte-derived DC (solid triangles) at 37°C were analyzed with a FACScan® flow cytometer. MFI, mean fluorescence intensity. Data shown are one representative of three experiments.
Figure 10.

DC derived from CD4+CD11c−CD3− plasmacytoid cells induces strong proliferation of CD4+CD45RA+ naive T cells. Purified allogeneic blood CD4+CD45RA+ naive T cells were cocultured with different numbers of DC generated from: CD14+ monocyte–derived DCs (solid circle), CD34+ progenitor–derived DCs (solid square), plasmacytoid cells after 6 d of culture with IL-3 (open square) and plasmacytoid cells after 6 d of culture with IL-3 and CD40L (open circle). After 5 d of DC-T cell cocultures, cells were pulsed with 1 μCi [3H]thymidine for 8 h before harvesting and counting. Data shown are one representative of six experiments.
Discussion
The experiments described in this article demonstrate that the plasmacytoid T cells, which have long been observed in the T cell areas of peripheral lymphoid tissues, correspond to CD4+CD11c− blood DC precursors (15). These cells display six striking features: (a) they have a plasmacytoid morphology, characterized by well developed RER and Golgi apparatus, which would normally suggest a terminally differentiated cell type with an active secretory function; (b) they die very rapidly by apoptosis in simple culture medium, consistent with the previous report that nuclear pyknosis and tingible body macrophages were found within clusters of plasmacytoid T cells in situ (5) and suggesting a rapid cell turn over rate; (c) they are found either close to the HEV or within the lumen of HEV; this, together with the identification of CD4+CD11c− blood DC precursors (15), indicates that the tissue plasmacytoid DC precursors are blood derived; (d) IL-3 represents the only cytokine tested so far that maintains their survival and promotes their proliferation; (e) CD40-ligand promotes their differentiation into mature DC; ( f ) the plasmacytoid cell derived DC display features similar to a subset of tonsillar interdigitating cells (Björck, P., and Y.-J. Liu, unpublished observation), including low expression of myeloid antigens CD13 and CD33 and poor ability to uptake soluble antigens or to phagocytose particles.
DC represent an heterogeneous population of hematopoietic-derived cells that display potent ability to prime naive T cells and to stimulate memory T cells (22). The past few years have witnessed remarkable advances in the isolation of DC precursors and in the definition of culture conditions allowing the generation of DCs (23, 24). DCs have been generated from early hematopoietic progenitors that can be isolated from human and mouse bone marrow (25– 28), thymus (29–32), or human blood/cord blood (12, 13, 33, 34). Hematopoietic progenitors isolated from bone marrow were shown to have the potential of becoming dendritic cells as well as granulocytes, monocytes, and macrophages (27, 34–36). This together with the fact that monocytes have the potential to differentiate into both macrophages and dendritic cells (11, 12, 37) suggests a myeloid DC developmental pathway. A critical cytokine for myeloid DC development was shown to be GM-CSF. Other factors such as TNFα, IL-4, SCF, and CD40-ligand were shown to enhance the myeloid DC maturation (23, 24). DC have also been generated from the most immature thymic T cell precursors or from bone marrow lymphoid progenitors, which could differentiate into T cells, B cells, and NK cells (28–32). These experiments suggest a lymphoid DC developmental pathway. Interestingly, GM-CSF does not appear to be required for DC generation from lymphoid precursors (31). In contrast, IL-3 seems to be one of the key cytokine required for lymphoid DC development (28–31). A recent experiment by Maraskovsky et al. (38) showed that Flt3L could potently enhance the development of both myeloid and lymphoid DCs in mice. The dependence of CD4+CD11c− plasmacytoid cells on IL-3 but not GM-CSF to differentiate into DCs, which express relatively low levels of CD13 and CD33 myeloid antigens, suggests that these plasmacytoid cells may represent lymphoid-derived immediate DC precursors.
The capacity of CD40-ligand to promote the differentiation of CD4+CD11c−Lin− plasmacytoid cells into DC is consistent with the previous reports on the diverse effects of CD40-ligand on the maturation and function of DCs. Ligation of CD40 on dendritic cells was shown to trigger production of high levels of IL-12 (39, 40), enhance T cell stimulatory capacity (21, 41–43) and improve DC survival (21, 42, 43). A recent experiment showed that ligation of CD40 induced a subset of human CD34+ hematopoietic progenitors to proliferate and differentiate into DC (44). Taken together, these data suggest that T cells play important roles in DC generation and maturation.
Tonsillar CD4+CD11c− plasmacytoid cells share many similar features with the blood CD4+CD11c− (15) or CD33low CD14low DC precursors (45). However, blood CD4+CD11c− cells appear to survive better than tonsillar CD4+CD11c− cells in culture medium (15, our unpublished observation), suggesting that the apoptosis program of these cells may be further triggered upon their arrival within tissues. Blood CD4+CD11c− cells were shown earlier to differentiate into DCs in the presence of undefined factors within monocyte-conditioned medium (15). In our hands, these blood CD4+CD11c− cells were found to become DCs in the presence of IL-3 and CD40-ligand (data not shown), as tonsillar CD4+CD11c− cells. This strongly suggests that the tonsillar CD4+CD11c− plasmacytoid cells are derived from blood CD4+CD11c− cells.
The increased accumulation of CD4+CD11c− plasmacytoid cells in the draining lymph nodes of tumor patients suggests important immunological functions in these pathological conditions. They may be constantly produced in bone marrow and may migrate through blood to the T cell areas of the spleen by crossing the marginal zone sinuses or the T cell areas of lymph nodes or tonsils by crossing the HEV. There, they would undergo rapid apoptosis, unless rescued by T cell signals (IL-3+CD40-ligand) generated by on going antigen-driven T cell–dependent immune responses. Thus, the IL-3 and CD40-ligand–dependent DC development from plasmacytoid precursors may represent a unique peripheral regulatory pathway that can rapidly recruit large numbers of DC into immune responses.
Acknowledgments
We thank C. Caux and K. Shortman for helpful discussions and critically reading the manuscript; M. Balzer and M. Erni for technical help; J. Chiller for support; S. Bonnet-Arnaud and M. Vatan for editorial help.
Footnotes
G. Grouard was supported by the Foundation Marcel Merieux, Lyon, France.
1 Abbreviations used in this paper: DC, dendritic cell; GCDC, dendritic cells within germinal centers; HEV, high endothelial veinules; RER, rough endoplasmic reticulum; SEM, scanning electron microscopy; TEM, transmission electron microscopy.
References
- 1.Lennert K, Kaiserling E, Müller-Hermelink HK. T-associated plasma cells. Lancet. 1975;1:1031–1032. doi: 10.1016/s0140-6736(75)91974-1. [DOI] [PubMed] [Google Scholar]
- 2.Lennert, K., H. Stein, H.K. Müller-Hermelink, and R. Vollenweider. 1984. T cells with plasmacytoid features. In Leucocyte Typing. Human Leucocyte Differentiation Antigens Detected by Monoclonal Antibodies. A. Bernard, L. Boumsell, J. Dausset, C. Milstein and S.F. Schlossman, editors. Springer-Verlag, Berlin.
- 3.Papadimitriou CS, Stephanaki-Nikou SN, Malamou-Mitsi VD. Comparative immunostaining of T-associated plasma cells and other lymph-node cells in paraffin section. Virchows Arch (Cell Pathol) 1983;43:31–36. doi: 10.1007/BF02932940. [DOI] [PubMed] [Google Scholar]
- 4.Horny HP, Feller AC, Horst HA, Lennert K. Immunocytology of plasmacytoid T cells: marker analysis indicates a unique phenotype of this enigmatic cell. Hum Pathol. 1986;18:28–32. doi: 10.1016/s0046-8177(87)80189-2. [DOI] [PubMed] [Google Scholar]
- 5.Facchetti F, de Wolf-Peeters C. The plasmacytoid monocyte (the so called plasmacytoid T-cell). An enigmatic cell of the human lymphoid tissue. Biotest Bulletin. 1991;4:225–240. [Google Scholar]
- 6.Beiske K, Langholm R, Godal T, Marton PF. T-zone lymphoma with predominance of plasmacytoid T-cells associated with myelomonocytic leukemia-A distinct clinicopathological entity. J Pathol. 1986;150:247–255. doi: 10.1002/path.1711500404. [DOI] [PubMed] [Google Scholar]
- 7.Facchetti F, de Wolf-Peeters C, Mason DY, Pulford K, van den Oord JJ, Desmet VJ. Plasmacytoid T cells. Immunohistochemical evidence for their monocyte/macrophage origin. Am J Pathol. 1988;133:15–21. [PMC free article] [PubMed] [Google Scholar]
- 8.Müller-Hermelink HK, Steinman G, Stein H, Lennert K. Malignant lymphoma of plasmacytoid T-cells. Morphologic and immunologic studies characterizing a special type of T-cell. Am J Surg Pathol. 1983;7:849–862. [PubMed] [Google Scholar]
- 9.Beiske K, Munthe-Daas A, Davies CDL, Marton PF, Godal T. Single cell studies on the immunological marker profile of plasmacytoid T-zone cells. Lab Invest. 1987;56:381–393. [PubMed] [Google Scholar]
- 10.Arpin C, Déchanet J, van Kooten C, Merville P, Grouard G, Brière F, Banchereau J, Liu YJ. Generation of memory B cells and plasma cells in vitro. Science (Wash DC) 1995;268:720–722. doi: 10.1126/science.7537388. [DOI] [PubMed] [Google Scholar]
- 11.Sallusto F, Lanzavecchia A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J Exp Med. 1994;179:1109–1118. doi: 10.1084/jem.179.4.1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Romani N, Gruner S, Brang D, Kämpgen E, Lenz A, Trockenbacher B, Konwalinka G, Fritsch PO, Steinman RM, Schuler G. Proliferating dendritic cell progenitors in human blood. J Exp Med. 1994;180:83–93. doi: 10.1084/jem.180.1.83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Caux C, Dezutter-Dambuyant C, Schmitt D, Banchereau J. GM-CSF and TNF-α cooperate in the generation of dendritic Langerhans cells. Nature (Lond) 1992;360:258–261. doi: 10.1038/360258a0. [DOI] [PubMed] [Google Scholar]
- 14.Grouard G, Durand I, Filgueira L, Banchereau J, Liu YJ. Dendritic cells capable of stimulating T cells in germinal centers. Nature (Lond) 1996;384:364–367. doi: 10.1038/384364a0. [DOI] [PubMed] [Google Scholar]
- 15.O'Doherty U, Peng M, Gezelter S, Swiggard WJ, Betjes M, Bhardwaj N, Steinman RM. Human blood contains two subsets of dendritic cells, one immunologically mature and the other immature. Immunology. 1994;82:487–493. [PMC free article] [PubMed] [Google Scholar]
- 16.Banchereau J, Bazan F, Blanchard D, Brière F, Galizzi JP, van Kooten C, Liu YJ, Rousset F, Saeland S. The CD40 antigen and its Ligand. Annu Rev Immunol. 1994;12:881–922. doi: 10.1146/annurev.iy.12.040194.004313. [DOI] [PubMed] [Google Scholar]
- 17.Clark EA, Ledbetter JA. How B and T cells talk to each other. Nature (Lond) 1994;367:425–428. doi: 10.1038/367425a0. [DOI] [PubMed] [Google Scholar]
- 18.Foy TM, Aruffo A, Bajorath J, Buhlmann JE, Noelle RJ. Immune regulation by CD40 and its ligand GP39. Ann Rev Immunol. 1996;14:591–617. doi: 10.1146/annurev.immunol.14.1.591. [DOI] [PubMed] [Google Scholar]
- 19.Liu YJ, Joshua DE, Williams GT, Smith CA, Gordon J, MacLennan ICM. Mechanisms of antigendriven selection in germinal centers. Nature (Lond) 1989;342:929–931. doi: 10.1038/342929a0. [DOI] [PubMed] [Google Scholar]
- 20.Tsubata T, Wu J, Honjo T. B cell apoptosis induced by antigen receptor cross-linking is blocked by T cell signal through CD40. Nature (Lond) 1993;364:645–648. doi: 10.1038/364645a0. [DOI] [PubMed] [Google Scholar]
- 21.Caux C, Massacrier C, Vanbervliet B, Dubois B, van Kooten C, Durand I, Banchereau J. Activation of human dendritic cells through CD40 cross-linking. J Exp Med. 1994;180:1263–1272. doi: 10.1084/jem.180.4.1263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Steinman RM. The dendritic cell system and its role in immunogenicity. Annu Rev Immunol. 1991;9:271–296. doi: 10.1146/annurev.iy.09.040191.001415. [DOI] [PubMed] [Google Scholar]
- 23.Caux, C., and J. Banchereau. 1996. In vitro regulation of dendritic cell development and function. In Blood Cell Biochemistry. T. Whetton and J. Gordon. Plenum Press. London. 263–301.
- 24.Thomas R, Lipsky PE. Dendritic cells: origin and differentiation. Stem Cells. 1996;14:196–206. doi: 10.1002/stem.140196. [DOI] [PubMed] [Google Scholar]
- 25.Inaba K, Steinman RM, Pack MW, Aya H, Inaba M, Sudo T, Wolpe S, Schuler G. Identification of proliferating dendritic cell precursors in mouse blood. J Exp Med. 1992;175:1157–1167. doi: 10.1084/jem.175.5.1157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Reid CDL, Stackpoole A, Meager A, Tikerpae J. Interactions of tumor necrosis factor with granulocytemacrophage colony-stimulating factor and other cytokines in the regulation of dendritic cell growth in vitro from early bipotent CD34+progenitors in human bone marrow. J Immunol. 1992;149:2681–2688. [PubMed] [Google Scholar]
- 27.Szabolcs P, Moore MAS, Young JW. Expansion of immunostimulatory dendritic cells among the myeloid progeny of human CD34+bone marrow precursors cultured with c-kit-ligand, GM-CSF, and TNFα. J Immunol. 1995;154:5851–5861. [PubMed] [Google Scholar]
- 28.Galy A, Travis M, Cen D, Chen B. Human T, B, natural killer, and dendritic cells arise from a common bone marrow progenitor cell subset. Immunity. 1995;3:459–473. doi: 10.1016/1074-7613(95)90175-2. [DOI] [PubMed] [Google Scholar]
- 29.Ardavin C, Wu L, Li CL, Shortman K. Thymic dendritic cells and T cells develop simultaneously in the thymus from a common precursor population. Nature (Lond) 1993;362:761–763. doi: 10.1038/362761a0. [DOI] [PubMed] [Google Scholar]
- 30.Wu L, Vremec D, Ardavin C, Winkel K, Süss G, Georgious H, Maraskovsky E, Cook E, Shortman K. Mouse thymus dendritic cells: kinetics of development and changes in surface markers during maturation. Eur J Immunol. 1995;25:418–425. doi: 10.1002/eji.1830250217. [DOI] [PubMed] [Google Scholar]
- 31.Saunders D, Lucas K, Ismaili J, Wu L, Maraskovsky E, Dunn D, Shortman K. Dendritic cell development in culture from thymic precursor cells in the absence of granulocyte-macrophage colony stimulating factor. J Exp Med. 1996;184:2185–2196. doi: 10.1084/jem.184.6.2185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Reis P, Martinez-Caceres E, Jaleco AC, Staal F, Noteboom E, Weijer K, Spits H. CD34+ CD38dimcells in the human thymus can differentiate into T, natural killer, and dendritic cells but are distinct from pluripotent stem cells. Blood. 1996;87:5196–5206. [PubMed] [Google Scholar]
- 33.Rosenzwajg M, Canque B, Gluckman JC. Human dendritic cell differentiation pathway from CD34+hematopoietic precursor cells. Blood. 1996;87:535–544. [PubMed] [Google Scholar]
- 34.Santiago-Schwarz F, Belilos E, Diamond B, Carsons SE. TNF in combination with GM-CSF enhances the differentiation of neonatal cord blood stem cells into dendritic cells and macrophages. J Leukocyte Biol. 1992;52:274–281. [PubMed] [Google Scholar]
- 35.Inaba K, Inaba M, Deguchi M, Hagi K, Yasumizu R, Ikehara S, Muramatsu S, Steinman RM. Granulocytes, macrophages, and dendritic cells arise from a common major histocompatibility complex class II-negative progenitor in mouse bone marrow. Proc Natl Acad Sci USA. 1993;90:3038–3042. doi: 10.1073/pnas.90.7.3038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Caux C, Vanbervliet B, Massacrier C, Dezutter-Dambuyant C, de Saint-Vis B, Jacquet C, Yoneda K, Imamura S, Schmitt D, Banchereau J. CD34+hematopoietic progenitors from human cord blood differentiate along two independent dendritic cell pathways in response to GM-CSF+ TNFα. J Exp Med. 1996;184:695–706. doi: 10.1084/jem.184.2.695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zhou L-J, Tedder TF. CD14+ blood monocytes can differentiate into functionally mature CD83+dendritic cells. Proc Natl Acad Sci USA. 1996;93:2588–2592. doi: 10.1073/pnas.93.6.2588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Maraskovsky E, Brasel K, Teepe M, Roux ER, Lyman SD, Shortman K, McKenna HJ. Dramatic increase in the numbers of functionally mature dendritic cells in Flit3 ligand-treated mice: multiple dendritic cell subpopulations identified. J Exp Med. 1996;184:1953–1962. doi: 10.1084/jem.184.5.1953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Cella M, Scheidegger D, Palmer-Lehmann K, Lane P, Lanzavecchia A, Alber G. Ligation of CD40 on dendritic cells triggers production of high levels of interleukine12 and enhances T cell stimulatory capacity: T-T help via APC activation. J Exp Med. 1996;184:747–752. doi: 10.1084/jem.184.2.747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kock F, Stanzl U, Jennewein P, Janke K, Heufler C, Kampgen E, Romani N, Schuler G. High level IL-12 production by murine dendritic cells: upregulation via MHC class II and CD40 molecule and downregulation by IL-4 and IL-10. J Exp Med. 1996;184:741–746. doi: 10.1084/jem.184.2.741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.McLellan AD, Sorg RV, Williams LA, Hart DN. Human dendritic cells activate T lymphocytes via a CD40: CD40 ligand dependent pathway. Eur J Immunol. 1996;26:1204–1210. doi: 10.1002/eji.1830260603. [DOI] [PubMed] [Google Scholar]
- 42.Peguet-Navarro J, Dalbiez-Gauthier C, Rattis FM, Van-Kooten C, Banchereau J, Schmitt D. Functional expression of CD40 antigen on human epidermal Langerhans cells. J Immunol. 1995;155:4241–4247. [PubMed] [Google Scholar]
- 43.Ludewig B, Graf D, Gelderblom HR, Becker Y, Kroczek RA, Pauli G. Spontaneous apoptosis of dendritic cells is efficiently inhibited by TRAP (CD40-ligand) and TNF-alpha, but strongly enhanced by interleukin-10. Eur J Immunol. 1995;25:1943–1950. doi: 10.1002/eji.1830250722. [DOI] [PubMed] [Google Scholar]
- 44.Flores-Romo L, Björck P, Duvert V, Van-Kooten C, Saeland S, Banchereau J. CD40 ligation on human cord blood CD34+hematopoietic progenitors induces their proliferation and differentiation. J Exp Med. 1997;185:341–349. doi: 10.1084/jem.185.2.341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Thomas R, Lipsky PE. Human peripheral blood dendritic cell subsets. Isolation and characterization of precursor and mature antigen-presenting cells. J Immunol. 1994;153:4016–4028. [PubMed] [Google Scholar]