

Abstract
T cell costimulation, particularly by the B7 family members B7-1 and B7-2, plays a critical role in regulating T cell–mediated immunity. Two molecules on T cells, CD28 and CTLA-4, are known to bind to B7. It has been suggested that CD28–B7 interaction promotes T cell response, whereas B7–CTLA-4 interaction downregulates T cell clonal expansion. However, the proposed responses of individual receptors to B7 have not been verified directly. Here, we report that B7-1 promotes clonal expansion of CD28-deficient T cells, and that the CD28-independent costimulatory activity is mediated by CTLA-4, as it is completely blocked by intact and Fab of anti–CTLA-4 mAb. In addition, a mutant B7-1 molecule, B7W88 >A, which has lost binding to CD28 but retained significant CTLA-4 binding activity, promotes T cell clonal expansion. Furthermore, while presence of CD28 enhances T cell response to B7-1, such response is also completely blocked by anti–CTLA-4 mAb. Taken together, our results demonstrate that B7–CTLA-4 interaction promotes T cell clonal expansion, and that optimal T cell response to B7 is achieved when both CD28 and CTLA-4 interact with B7. These results establish an important function of CTLA-4 in promoting T cell activation, and suggest an alternative interpretation of the function of CTLA-4 in T cell activation.
Cell surface costimulatory molecules, of which B7 family members are known as prototypes, play an important role in T cell responses (1–4). Two T cell surface molecules, CD28 and CTLA-4, bind B7 with different affinity (5–8). Most T cells can express both receptors, although their expression appears to be differentially regulated (9–11). In addition, both receptors may initiate signal transduction, as they are associated with an overlapping but perhaps distinct set of molecules involved in regulating cell growth (12–17). However, the function of the individual receptors is still unclear.
It has been proposed that B7–CD28 interaction delivers a positive signal, whereas B7–CTLA-4 interaction delivers a negative signal for T cell responses (9, 10). This hypothesis is based on two lines of evidence. First, targeted mutation of CTLA-4 causes lethal lymphoproliferative disease (18, 19). Second, depending on experimental models, anti– CTLA-4 mAbs either enhance or inhibit T cell clonal expansion (9, 10, 20, 21) in vitro; and in vivo, both intact and Fab fragments of anti–CTLA-4 mAbs enhance T cell response and tumor rejection (22–24). However, the proposed inhibitory function of B7–CTLA-4 interaction has not been directly demonstrated.
If B7–CTLA-4 interaction delivers a negative signal for T cell growth, T cell response in CD28-deficient mice should be enhanced when the function of B7 is blocked. However, a study by Green et al. (25) using CD28-deficient T cells has not revealed an enhancement by CTLA-4Ig. In this study, we reevaluated the effect of B7–CTLA-4 interaction on T cell clonal expansion. Our results demonstrate that this interaction is sufficient to costimulate clonal expansion of T cells, and that optimal T cell response to B7-1 requires cooperation of CD28 and CTLA-4.
Materials and Methods
Experimental Animals, mAbs, and Fusion Proteins.
C57BL/6 mice were purchased from the National Cancer Institute (Rockville, MD). CD28-deficient mice (26), backcrossed to C57BL6j for six generations, were provided by Dr. T.W. Mak (University of Toronto, Canada).
Anti–CTLA-4 mAb 4F10 (9) and anti-CD3 mAb 2C11 (27) were provided by Dr. J.A. Bluestone (University of Chicago, Chicago, IL). Anti–B7-1 mAb 16.10A1 (28) was provided by Dr. H. Reiser (Dana-Farber Cancer Institute, Boston, MA). Another anti–B7-1 mAb 3A12 was produced in this laboratory and has been described (29). Anti-CD45RB mAb C363.16A (30) was provided by Dr. K. Bottomly (Yale University, New Haven, CT), anti-CD11c mAb HB224 (31) was a gift from Dr. R. Steinman (Rockefeller University, New York). Intact mAbs were purified by a protein G column, while the Fab fragments were prepared using a Fab kit (catalog no. 44885) from Pierce Chemical Company (Rockford, IL) according to the manufacturer's instructions. The purity of both intact and Fab fragments was verified by 10% SDS-PAGE.
Construction and production of fusion proteins, murine CTLA-4Ig, and CD28Ig have been fully described (32).
Transfection of COS and Chinese Hamster Ovary Cells.
COS cells were transfected with wild-type and mutant B7-1 molecules by DEAE–dextran methods, as has been described (33). The procedure for Chinese hamster ovary (CHO)1 cell transfection has also been described (34).
Flow Cytometry.
Expression of wild-type and mutant B7-1 was determined by flow cytometry using anti–B7-1 mAbs, either 16.10A1, or 3A12, as primary antibody, and FITC-labeled goat anti–hamster IgG was used as second-step reagent, as has been described. Fusion proteins, either CD28Ig or CTLA-4Ig, were used as first-step reagents. The binding to wild-type and mutant B7-1 was determined by using FITC-labeled goat anti–mouse IgG as second-step reagents.
Proliferation Assay.
CD4 T cells were purified from spleen cells as described (35). The purity of CD4 T cells isolated was >95%. Given numbers of the CD4 T cells were stimulated with anti-CD3 mAb (0.25 μg/ml) using mitomycin C (100 μg/ml, 37°C for 1 h)–treated CHO cells transfected with either FcR (CHOFcR) or FcR plus murine B7-1 (CHOFcRB7). The cultures were maintained for 42 h, and were pulsed with [3H]TdR (1.25 μCi/ well) for an additional 6 h. The cultures were then harvested and the [3H]TdR incorporated into the genomes was determined by a betaplate counter. The blocking mAbs were added at the beginning of the culture. The data presented are means of duplicates with variation <15% of the mean.
In some experiments, CD4 T cells were separated based on their cell surface expression of CD45RB. In brief, CD4 T cells were incubated with anti-CD45RB mAb C363.16A (5 μg/ml) for 1 h at 4°C; the unbound antibodies were washed away. After adding DYNAL beads coated with goat anti–rat Ig for 1 h, the cells that bound to C363.16A were collected by magnet. The cells bound to the beads were eluted by adding 2 mg/ml of normal rat Ig at 37°C for 1 h and used as naive cells. The cells that did not bind DYNAL beads in the first place were depleted once more with DYNAL beads and used as memory cells. The CD45RB profiles were determined by flow cytometry using either C363.16A (CD45RB) or medium (control) as the first-step reagent and FITC-labeled mouse anti–rat Ig as second-step reagent.
Site-directed Mutagenesis.
Site-directed mutagenesis of murine B7-1 was achieved by PCR using oligonucleotides carrying the desired mutations. B7Y that carries a mutation from Y >A at position 201 has been described (32). Mutant B7W (W >A at position 88) was created as follows. A B7-1 fragment containing the desired mutation was produced by PCR using GCTCGAGAAGCTTATGGCTTGCAATTGTCAG as forward primer, and CTTAGCCTCGGGCGCCAC TTTTAGTTTCCC as reverse primer. After digestion by AvaI, the PCR product was ligated to the AvaI–XbaI fragment of murine B7-1 (32). B7L (L109 >A) was created by a two-step PCR. First, a mutant B7-1 fragment containing the desired mutation (L >A) is generated using GCTCGAGAAGCTTATGGCTTGCAATTGTCAG as forward primer, and CGACGCAGCTGTAGGTGCCCCGGTCGCTAGCGACCAGGC as reverse primer; another fragment covering the remaining sequence of B7-1 open reading frame is amplified using CACCTACAGCTGCGTCGTTCAAAAGAAGGA as forward primer and CGAATTCTAGAACTAAAGGAAGACGGTCT as reverse primer. Second, a mixture of the two B7-1 fragments were used as template to amplify the full-length B7-1 mutant; GGCTCTAGATTCCTGGCTTTCCCCATCATG was used as forward primer and GTCAGCCATCTCGAGTTTTTCCCAGGTGAAGTC was used as reverse primer. The PCR condition has been described (32).
Results
CD28-independent Costimulatory Activity of B7-1.
To examine the function of CD28 and CTLA-4, we compared wild-type and CD28-deficient T cells in their response to costimulation by B7 in vitro. We produced CHO cells transfected with either FcR (CHOFcR), which cross-links anti-CD3 mAb on T cell surface, or FcR in conjunction with murine B7-1 (CHOFcRB7) (23), and tested their costimulatory activity for clonal expansion of CD4 T cells from wild-type and CD28(−/−) mice. As expected, wildtype CD4 T cells respond to costimulation by B7-1 (Fig. 1 a). Surprisingly, T cells from CD28-deficient mice also mount a positive response to B7-1, although the response is approximately fivefold lower than that of wild-type T cells (Fig. 1 b). The critical role of B7-1 is confirmed, as an anti– B7-1 mAb 3A12 (29) significantly blocks responses of wildtype and CD28-deficient T cells. In addition, CD4 T cells from wild-type and CD28-deficient mice bearing naive or memory markers both respond positively to costimulation by B7-1 (Fig. 2). These results demonstrate that B7-1 can costimulate T cell clonal expansion by a CD28-independent mechanism.
Figure 1.

Costimulatory molecule B7-1 significantly enhances antiCD3–induced proliferative responses of CD4 T cells from either wildtype (a) or CD28-deficient (b) mice. Varying numbers of the wild-type (a) and CD28-deficient (b) CD4 T cells were stimulated with anti-CD3 mAb and mitomycin C–treated CHO cells that were transfected with either FcR (CHOFcR) or FcR plus murine B7-1 (CHOFcRB7) (104/well). The cultures were maintained for 42 h, and were pulsed with [3H]TdR (1.25 μCi/well) for an additional 6 h. Either normal hamster Ig or anti-B7 mAb 3A12 was added at the beginning of the culture to evaluate the contribution of B7-1 to T cell proliferation. cpm of [3H]TdR incorporated into mitomycin-treated CHO cells: CHOFcR, 326; CHOFcRB7, 1474. The CD28-independent costimulatory activity of B7-1 has been repeated 10 times.
Figure 2.

CD4 T cells expressing either naive or memory markers are capable of responding to B7-1 by a CD28-independent mechanism. (a) CD45RB profiles of the subsets of CD4 T cells used in the study. (b) Proliferative responses of CD4 T subsets from either wild-type (WT ) or CD28(−/−) mice to soluble anti-CD3 using CHO cells transfected with either FcR or FcR plus murine B7-1 as accessory cells.
Contribution of CTLA-4 to B7-1–mediated Costimulation.
To verify the contribution of CTLA-4, we tested whether an anti–CTLA-4 mAb affects the function of B7-1. We prepared Fab of the mAb by papain digestion and depletion of Fc-containing immunoglobulin. As shown in Fig. 3, in SDS-PAGE under reducing conditions, intact mAb preparation consists of two peptides of ∼50 kD and ∼28 kD, the respective molecular masses of the heavy and light chain of IgG. Fab preparation run as one band of ∼28 kD, which indicate that the Fab used for the study is free of detectable intact Ig. As shown in Fig. 4, the anti–CTLA-4 mAb 4F10 completely blocks the function of B7-1. This blocking does not require cross-linking of CTLA-4, as the Fab fragment of the mAb also blocked the function of T cells. Surprisingly, both intact and Fab fragment of anti–CTLA-4 mAb inhibit response of CD28(+/+) T cells. The ability of both intact and Fab of anti–CTLA-4 mAb 4F10 to block the function of wild-type T cells indicates that CTLA-4 is critically involved in B7-1–mediated costimulation for wildtype T cells.
Figure 3.
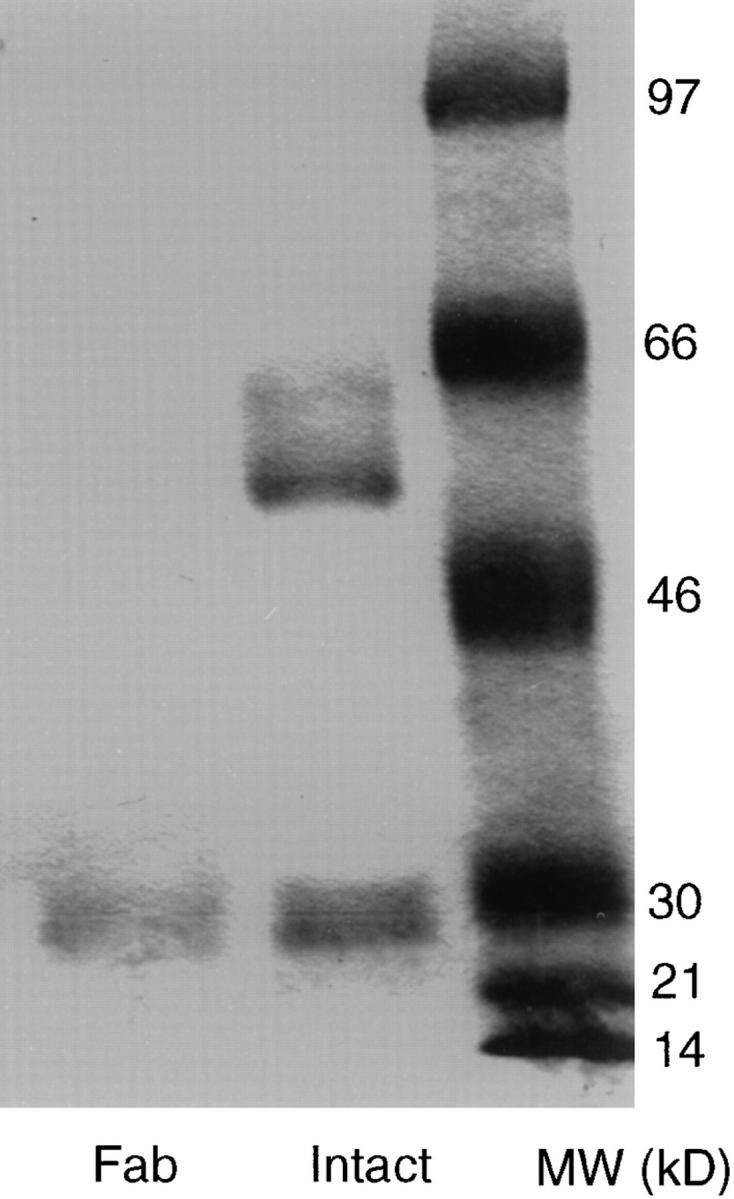
Fab preparation used in the study does not contain intact mAb. Anti–CTLA-4 mAb 4F10 was purified by affinity purification. Purified mAb was digested with papain, and Fc-containing molecules removed using immobilized protein A. 5 μg of intact and 2 μg of Fab were analyzed by 105 SDS-PAGE under reducing conditions.
Figure 4.

Both intact and Fab fragments of anti–CTLA-4 mAb block the costimulatory activity of B7-1. Purified CD4 T cells (105/well) either wild-type (a) and CD28-deficient (b) mice were stimulated with soluble anti-CD3 mAb (0.25 μg/ml) in the presence of mitomycin C–treated CHO cells (3 × 103/well) transfected with FcR plus B7-1 (FcRB7) for 48 h. The proliferation of CD4 T cells was determined by incorporation of [3H]TdR in the last 6 h of culture. Given concentrations of intact or Fab fragment of anti–CTLA-4 mAb 4F10, or control mAb HB224 were added at the beginning of the culture. Data shown represent five independent experiments.
A potential caveat of this interpretation is that anti– CTLA-4 may negatively signal T cells and thus prevent T cell clonal expansion regardless of the presence of B7-1. To rule out this possibility, we tested whether anti–CTLA-4 mAb inhibits T cell response to costimulation by CD44H, which we recently identified as a CD40L-induced costimulatory molecule that promotes T cell clonal expansion by a CD28-independent mechansim (36). As shown in Fig. 5, anti–CTLA-4 mAb 4F10 does not inhibit clonal expansion of T cells when CHO cells expressing murine CD44H are used as costimulator. These results confirm that anti– CTLA-4 mAb is a specific inhibitor of T cell response to costimulation by B7-1.
Figure 5.

Specificity of inhibition by anti–CTLA-4 mAb. Wild-type (a–c, 5 × 104/well) or CD28(−/−) (d–f, 105/well) CD4 T cells were stimulated with anti-CD3 mAb and CHO cells (mitomycin C–treated, 3,000/well) transfected with either FcR, FcR plus B7-1 (FcRB7), or FcR plus CD44H (FcRCD44H) for 48 h in the presence of control mAb HB224, anti–B7-1 mAb 10.16A, and anti–CTLA-4 mAb 4F10. (a and d) Proliferation of wildtype and CD28(−/−) T cells to anti-CD3 in the presence of CHOFcR, CHOFcRB7, and CHOFcR CD44H. Note that twice as many CD28(−/−) CD4 T cells were used to ensure a comparable total proliferative response. (b and e) Lack of inhibition by anti–CTLA-4 mAb when CD44H was used as costimulator. (c and f ) Inhibition by anti–B7-1 and anti–CTLA-4 when B7-1 was used as costimulators.
Functional Analysis of B7-1 Mutant Supports CTLA-4 as a Positive Regulator for T Cell Clonal Expansion.
Anti–CTLA-4 mAbs show different effects on T cell proliferation depending whether costimulation is provided by anti-CD28 or by antigen-presenting cells (9, 10, 20–24). To avoid any question that may be associated with antibody blocking studies, we have carried out a systematic site-directed mutagenesis of B7-1 molecules in both IgC-like (32) and IgV-like domains in search of a mutant B7 that retains binding for CTLA-4 but not CD28. As shown in Fig. 6 a, saturating amount of CTLA-4Ig binds B7W, which carries a mutation from W to A at position 88 of B7-1 (number starts at the first methionine encoded by B7-1 cDNA), at levels comparable to those of wild-type B7, although careful titration of CTLA-4Ig reveals that it takes about ninefold more CTLA-4Ig to achieve such saturation for B7W (Fig. 6 c). In contrast, B7W does not bind CD28Ig (Fig. 6 b). Mutant B7 L109 >A (B7L), which binds CTLA-4Ig less well than B7W, binds CD28 even better than wild-type B7-1. On the other hand, mutant B7Y, which has a mutation at position 201 from Y to A, does not show detectable binding to either CD28Ig or CTLA-4Ig, as we have reported (32). These results demonstrate that B7-1 is recognized by the two receptors asymmetrically, and the selective loss of CD28 binding activity in B7W cannot be accounted for simply on the basis of low avidity of CD28–B7 interaction. The preferential effect of B7W >A mutation on CD28 binding is consistent with an earlier mutagenesis analysis involving human B7-1 expressed on fibroblasts (37), although it is apparently at variance with mutagenesis analysis using recombinant human B7-1 fusion protein (38), perhaps owing to differential glycosylation of B7-1.
Figure 6.

Characterization of B7-1 mutants. (a) FACS® profiles for the binding of wild-type and mutant B7 molecules, expressed on COS cells by transient transfection, to saturating amount of CD28Ig (100 μg/ml) and CTLA-4Ig (10 μg/ml) and anti-B7-1 mAb 10.16A (10 μg/ml). Numbers in each panel are mean fluorescences. (b and c) Quantitative analysis of CD28Ig and CTLA-4Ig binding to wild-type and mutant B7-1 (16, 23). COS cells were either mock-transfected or transfected with wild-type and mutant B7 cDNA. The COS cells were used to determine ability of wild-type and mutant B7 to bind varying concentration of either CD28Ig or CTLA-4Ig. The data shown are mean fluorescences normalized for the cell surface expression of B7-1 according to the following formula: Normalized MF for receptor binding = (MFmutant − MFmock) × k, where k = (MFmutant − MFmock) / (MFWT − MFmock) when anti-B7-1 mAb 10.16A is used to measure cell surface expression of wild-type or mutant B7-1. Representative of three independent experiments.
Therefore, we transfected B7-1, B7W, and B7Y into CHOFcR cells, and produced stable cell lines expressing comparable levels of FcR and wild-type or mutant B7-1 (Fig. 7 a). These cell lines were used to determine the receptors responsible for the costimulatory activity of B7-1. As shown in Fig. 7 b, both wild-type and CD28-deficient T cells respond to costimulation by wild-type B7-1 and B7W, but not to that by B7Y. Thus, CTLA-4 binding activity is sufficient to costimulate T cell proliferation. Moreover, wild-type T cells respond to B7-1 significantly better than to B7W, as ∼20-fold more CHO cells expressing B7W are required to achieve a level of T cell proliferation similar to that costimulated by wild-type B7-1. This difference is diminished when CD28(−/−) CD4 T cells are used, thus confirming the inability of B7W to stimulate CD28. The function of B7W is blocked by anti–B7-1 and anti–CTLA-4 mAbs (Fig. 8).
Figure 7.

CTLA-4 binding activity is required for CD28-independent costimulation by B7-1. (a) Levels of FcR and B7 on CHO cells transfected with wild-type and mutant B7-1 measured by antiFcR mAb 2.4G2 or anti–B7-1 mAb 3A12. (b and c) Costimulatory activity of wild-type and B7 mutant B7W (88W >A) and B7Y (201Y >A) for either wild-type (b) or CD28(−/−) (c) T cells. Wild-type and CD28-deficient CD4 T cells were stimulated with anti-CD3 mAb and 104/well of mitomycin C–treated CHO cells. Data presented are representatives of three independent experiments.
Figure 8.

Anti–CTLA-4 mAb inhibits B7W-mediated costimulation for clonal expansion of T cells from wild-type and CD28-deficient mice. CD4 T cells (105/well) were stimulated for 48 h by anti-CD3 mAb and mitomycin C–treated CHO cells that have been transfected with both FcR and B7W (104/well). Given concentration of anti–CTLA-4 mAb 4F10, anti–B7-1 mAb 3A12, and control anti-CD11c mAb HB224 was added at the beginning of culture. Data presented are representatives of three independent experiments.
Discussion
CTLA-4 plays an important role in regulating T cell function, as evidence by the phenotypes of CTLA-4–deficient mice (17, 18) and by the effect of anti–CTLA-4 mAbs (9–11). The biological consequences of B7–CTLA-4 interactions have not been studied in detail. Here we seek to measure directly the effect of B7–CTLA-4 interaction on T cell clonal expansion. Our results demonstrate that B7–CTLA-4 interaction is sufficient to induce T cell clonal expansion in vitro.
First, T cells from CD28-deficient mice react to costimulation by B7-1, although the proliferative response is generally 5 to 10-fold lower than wild-type T cells. This response is mediated by CTLA-4 on T cells, because anti-CTLA-4 mAb, either intact or Fab, blocks B7-1–dependent T cell proliferation. Surprisingly, in numerous experiments, anti– CTLA-4 mAb also completely inhibit B7-1–mediated proliferation of wild-type T cells. Because this mAb does not interfere with T cell proliferation when CD44H is used as costimulator (Fig. 5), it is unlikely that this mAb inhibits T cell response to B7-1 by negative signaling. These results suggest that B7–CD28 interaction may not be sufficient to promote T cell proliferation. This is paradoxical, given the fact that anti-CD28 mAb used as the prototypic costimulator for T cell clonal expansion (39–43), although it has not been demonstrated whether B7–CD28 interaction is sufficient to costimulate T cell proliferation. Because anti-CD28 mAb 9.3 has ∼100-fold higher avidity than B7-1Ig for CD28 (7), this paradox can be reconciled by higher avidity of the anti-CD28 mAb. It should be noted that in an earlier report, Green et al. (25) failed to detect CD28-independent function of B7-1 in promoting T cell clonal expansion in the presence of PMA. This difference can be explained by possible lack of expression of CTLA-4 in the CD28(−/−) cells used by Green et al. Alternatively, our assay based on copresentation of TCR ligand and costimulator B7-1 on the same cells (44) is ∼20-fold more sensitive than the earlier study; the difference can be reconciled on the basis of sensitivity of the assays.
Second, mutant B7-1, B7W, which lacks detectable CD28 binding activity, promotes proliferation of both wild-type and CD28-deficient T cells. For better quantitation, we measured CD28 binding using fusion protein CD28Ig. However, our preliminary data indicated that CD28 expressed on CHO cells as a transmembrane protein also fail to mediate adhesion to B7W (data not shown), much like human B7-1 with a mutation of the corresponding amino acid (37). Furthermore, while mutant B7W stimulates wild-type T cells less well than wild-type B7-1, it is comparable to wild-type B7-1 if CD28(−/−) T cells are used as responder. Thus, the major functional difference between B7W and wildtype B7-1 is in their interaction with CD28. Our functional analysis of B7-1 mutant indicates that B7–CTLA-4 binding is sufficient to costimulate wild-type and CD28deficient T cells. The use of B7-1 mutants allows us to dissect the function of CTLA-4 without the caveats of anti– CTLA-4 mAb.
A recent study suggests that another yet unidentified receptor for B7 may exist on NK cells (45). An intriguing question is whether the CD28-independent response to B7-1, as reported here, is mediated by a yet unidentified receptor for B7-1. Although it is impossible to rule out a possible contribution of other unknown receptors, CTLA-4 is at least an essential part of the signaling complex, because the CD28-independent function of B7-1 is completely blocked by anti–CTLA-4 mAb.
Taken together, our results presented in this report demonstrate that B7–CTLA-4 interaction promotes T cell clonal expansion. Moreover, optimal costimulation by B7 requires both CD28 and CTLA-4, as has been proposed by Linsley et al. (11). The mechanism of the cooperation between CD28 and CTLA-4 is still unclear at the present. B7–CD28 interaction may be critical for enhancing IL-2 production. Thus, it has been reported that CD28 deficiency leads to a drastic reduction in IL-2 production (25, 26, 37). Our analysis of IL-2 production in CD28-deficient T cells (data not shown) also supports this concept. In addition, mutant B7W, which bind CTLA-4 but not CD28, promotes T cell clonal expansion but barely enhances IL-2 production (data not shown). Therefore, unlike CD28-B7 interaction, CTLA-4–B7 interaction may not enhance T cell response by increasing the production of IL-2. Our results are inconsistent with the notion that CTLA-4 is a negative regulator for T cell activation, although it is conceivable that CTLA-4 may downregulate T cell activation on other circumstances. Several important factors, such as expression and cellular localization of CTLA-4 may influence the function of CTLA-4. Although it is clear that the expression and cellular expression of CTLA-4 is under stringent control (47), the regulatory mechanisms are not well understood. This lack of knowledge has made it difficult to predict the function of CTLA-4 under physiological conditions.
Several recent studies have shown that anti–CTLA-4 mAbs can be potent enhancers of immune response to defined foreign antigen, self-antigens, and tumors (22–24). Although these results were interpreted in the light of blocking negative signal from CTLA-4, it is also possible that these results are achieved by enhancing positive signal transduction from CTLA-4. The mutant B7-1 molecules described in this study may help to resolve the function of CTLA-4 in vivo. The answer to this question will be critical for immune intervention targeted at CTLA-4, as it will determine whether signal transduction initiated by CTLA-4 should be amplified or inhibited.
Acknowledgments
We thank Dr. Tak Mak for CD28-deficient mice, Dr. Jeff Bluestone for anti–CTLA-4 monoclonal antibody, Dr. Stan Vukmanovic for critical reading of the manuscript, and John Hirst for assistance in flow cytometry.
Footnotes
This study is supported by grants from the National Institutes of Health (CA58033 and CA69016) and the Council for Tobacco Research, USA.
1 Abbreviation used in this paper: CHO, Chinese hamster ovary.
References
- 1.Brestcher P, Cohn M. A theory of self–nonself discrimination. Science (Wash DC) 1970;169:1042–1049. doi: 10.1126/science.169.3950.1042. [DOI] [PubMed] [Google Scholar]
- 2.Lafferty KJ, Prowse SJ, Simeonovich CJ, Warren HS. Immunobiology of tissue transplantation: a return to passenger leukocyte concept. Annu Rev Immunol. 1983;1:143–173. doi: 10.1146/annurev.iy.01.040183.001043. [DOI] [PubMed] [Google Scholar]
- 3.Mueller DL, Jenkins MK, Schwartz RH. Clonal expansion vs functional clonal inactivation: a costimulatory signalling pathway determines the outcome of T cell antigen occupancy. Annu Rev Immunol. 1989;7:445–480. doi: 10.1146/annurev.iy.07.040189.002305. [DOI] [PubMed] [Google Scholar]
- 4.Liu Y, Linsley PS. T cell costimulation. Curr Opin Immunol. 1992;4:265–270. doi: 10.1016/0952-7915(92)90075-p. [DOI] [PubMed] [Google Scholar]
- 5.Linsley PS, Clark EA, Ledbetter JA. The T cell antigen, CD28, mediates adhesion with B cells by interacting with the activation antigen B7. Proc Natl Acad Sci USA. 1990;87:5031–5035. doi: 10.1073/pnas.87.13.5031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Linsley PS, Brady W, Grosmaire L, Aruffo A, Damle NK, Ledbetter JA. Binding of the B cell activation antigen B7 to CD28 costimulates T cell proliferation and interleukin-2 mRNA accumulation. J Exp Med. 1991;173:721–730. doi: 10.1084/jem.173.3.721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Linsley PS, Brady W, Grosmaire L, Damle NK, Ledbetter JA. CTLA-4 is a second receptor for B cell activation antigen B7. J Exp Med. 1991;174:561–569. doi: 10.1084/jem.174.3.561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Linsley PS, Green JL, Brady W, Bajorath J, Ledbetter JA, Peach R. Human B7-1 (CD80) and B7-2 (CD86) binds with similar avidity but distinct kinetics to CD28 and CTLA-4 receptors. Immunity. 1994;1:793–801. doi: 10.1016/s1074-7613(94)80021-9. [DOI] [PubMed] [Google Scholar]
- 9.Walunas TL, Lenchow DJ, Bakker CY, Linsley PS, Freeman GJ, Green JM, Thompson CB, Bluestone JA. CTLA-4 can function as a negative regulator of T cell activation. Immunity. 1995;1:405–414. doi: 10.1016/1074-7613(94)90071-x. [DOI] [PubMed] [Google Scholar]
- 10.Krummel MF, Allison JP. CD28 and CTLA-4 have opposing effects on the response of T cells to stimulation. J Exp Med. 1995;182:459–466. doi: 10.1084/jem.182.2.459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Linsley PS, Green JA, Tan P, Bradshaw J, Ledbetter JA, Anasetti C, Damle NK. Co-expression and functional co-operation of CTLA-4 and CD28 on activated T lymphocyte. J Exp Med. 1992;176:1595–1604. doi: 10.1084/jem.176.6.1595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Pages F, Ragueneau M, Rottapel R, Truneh A, Nunes J, Imbert J, Olive D. Binding of phosphatidylinositol-3-OH kinase to CD28 is required for T cell signalling. Nature (Lond) 1994;369:327–329. doi: 10.1038/369327a0. [DOI] [PubMed] [Google Scholar]
- 13.August A, DuPont B. CD28 of T lymphocytes associates with the phosphatidylinositol-3-OH kinase. Int Immunol. 1994;6:769–774. doi: 10.1093/intimm/6.5.769. [DOI] [PubMed] [Google Scholar]
- 14.Prasad KV, Cai YC, Raab M, Dukeworth B, Cantley L, Shoelson SE, Rudd CE. T cell antigen CD28 interacts with the lipid kinase phosphatidylinositol-3-OH kinase by a cytoplasmic Tyr (p)-Met-Xaa-Met motif. Proc Natl Acad Sci USA. 1994;91:2834–2838. doi: 10.1073/pnas.91.7.2834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Truitt KE, Hicks CM, Imboden JB. Stimulation of CD28 triggers an association of between CD28 and phosphatidylinositol-3-OH kinase in Jurkat T cells. J Exp Med. 1994;179:1071–1076. doi: 10.1084/jem.179.3.1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Schneider H, Cai YC, Pradad KVS, Shoelson SE, Rudd CE. CTLA-4 binding to the lipid kinase phosphatidylinositol 3-kinase in T cells. J Exp Med. 1995;181:351–355. doi: 10.1084/jem.181.1.351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Marengere LE, Waterhouse P, Duncan GS, Mittrucker HW, Feng GS, Mak TW. Regulation of T cell receptor signaling by tyrosine phosphatase SYP association with CTLA-4. Science (Wash DC) 1996;272:1170–1173. doi: 10.1126/science.272.5265.1170. [DOI] [PubMed] [Google Scholar]
- 18.Waterhouse P, Penninger JM, Yimms E, Wakeham A, Shahinian A, Lee KP, Thompson CB, Griesser H, Mak TW. Lymphoproliferative disorders with early lethality in mice deficient in CTLA-4. Science (Wash DC) 1995;270:985–987. doi: 10.1126/science.270.5238.985. [DOI] [PubMed] [Google Scholar]
- 19.Tivol EA, Borriello F, Schweitzer AN, Lynch WP, Bluestone JA, Sharpe AH. Loss of CTLA-4 leads to massive lymphoproliferation and fatal multiorgan tissue destruction, revealing a critical negative regulatory role of CTLA-4. Immunity. 1995;3:541–547. doi: 10.1016/1074-7613(95)90125-6. [DOI] [PubMed] [Google Scholar]
- 20.Krummel MF, Allison JP. CTLA-4 engagement inhibits IL-2 accumulation and cell cycle progression upon activation of resting T cells. J Exp Med. 1996;183:2533–2540. doi: 10.1084/jem.183.6.2533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Walnunas TL, Bakker CY, Bluestone JA. CTLA-4-ligation blocks CD28-dependent T cell activation. J Exp Med. 1996;183:2541–2550. doi: 10.1084/jem.183.6.2541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kearney ER, Walunas TL, Karr RW, Morton RA, Loh DY, Bluestone JA, Jenkins MK. Antigen-dependent clonal expansion of a trace population of antigen-specific CD4+T cells in vivo is dependent on CD28 costimulation and inhibited by CTLA-4. J Immunol. 1995;155:1032–1036. [PubMed] [Google Scholar]
- 23.Karandikar NJ, Vanderlugt CL, Walunas TL, Miller SD, Bluestone JA. CTLA-4: a negative regulator of autoimmune disease. J Exp Med. 1996;184:783–788. doi: 10.1084/jem.184.2.783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Leach DR, Krummel MF, Allison JP. Enhancement of antitumor immunity by CTLA-4 blockade. Science (Wash DC) 1996;271:1734–1736. doi: 10.1126/science.271.5256.1734. [DOI] [PubMed] [Google Scholar]
- 25.Green JM, Noel PJ, Sperling AI, Walunas TL, Gray GS, Bluestone JA, Thompson CB. Absence of B7-dependent responses in CD28-deficient mice. Immunity. 1994;1:501–508. doi: 10.1016/1074-7613(94)90092-2. [DOI] [PubMed] [Google Scholar]
- 26.Shahinian A, Pfeffer K, Lee KP, Kundig KP, Kishihara TM, Wakeham A, Kawai K, Ohashi PS, Thompson CB, Mak TW. Differential T cell costimulatory requirements in CD28-deficient mice. Science (Wash DC) 1993;261:609–612. doi: 10.1126/science.7688139. [DOI] [PubMed] [Google Scholar]
- 27.Leo O, Foo M, Sachs DH, Samelson LE, Bluestone JA. Identification of a monoclonal antibody against murine T3 polypeptide. Proc Natl Acad Sci USA. 1987;84:1037–1041. doi: 10.1073/pnas.84.5.1374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Razi-Wolf Z, Freeman GJ, Galvin F, Benacerraf B, Nadler LM, Reiser H. Expression and function of the murine B7 antigen, the major costimulatory molecule expressed by peritoneal exudate cell. Proc Natl Acad Sci USA. 1992;89:4210–4214. doi: 10.1073/pnas.89.9.4210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wu Y, Guo Y, Liu Y. A major co-stimulatory molecule, CTLA-4 ligand A, is distinct from B7. J Exp Med. 1993;178:1789–1793. doi: 10.1084/jem.178.5.1789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bottomly K, Luqman M, Greenbaum L, Carding S, West J, Pasqualini T, Murphy D. A monoclonal antibody to murine CD45R distinguish CD4 T cell population that produce different cytokines. Eur J Immunol. 1989;19:617–623. doi: 10.1002/eji.1830190407. [DOI] [PubMed] [Google Scholar]
- 31.Metlay JP, Witmer-Pack MD, Agger R, Crowley MT, Lawless DS, Steinman RM. The distinct leukocyte integrins of mouse spleen dendritic cells as identified with new hamster monoclonal antibodies. J Exp Med. 1990;171:1753–1763. doi: 10.1084/jem.171.5.1753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Guo Y, Wu Y, Zhao M, Kong XP, Liu Y. Mutational analysis of an alternatively spliced product of B7 defines its CD28/CTLA-4–binding site on Immunoglobulin C–like domain. J Exp Med. 1995;181:1345–1355. doi: 10.1084/jem.181.4.1345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Aruffo A, Seed B. Molecular cloning of CD28 cDNA through a high efficiency COS cell expression system. Proc Natl Acad Sci USA. 1987;84:8573–8577. doi: 10.1073/pnas.84.23.8573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Liu Y, Jones B, Aruffo A, Sullivan KM, Linsley PS, Janeway CA., Jr Heat-stable antigen is a costimulatory molecule for CD4 T cell growth. J Exp Med. 1992;175:437–445. doi: 10.1084/jem.175.2.437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wu Y, Xu J, Shinde S, Grewal I, Henderson T, Flavell RA, Liu Y. Rapid induction of a novel costimulatory activity on B cells by CD40 ligand. Curr Biol. 1995;5:1303–1311. doi: 10.1016/s0960-9822(95)00257-0. [DOI] [PubMed] [Google Scholar]
- 36.Guo Y, Wu Y, Shinde S, Sy M-S, Aruffo A, Liu Y. Identification of a costimulatory molecule rapidly induced by CD40L as CD44H. J Exp Med. 1996;184:955–961. doi: 10.1084/jem.184.3.955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Fargeas CA, Truneh A, Reddy M, Hurle M, Sweet R, Sekaly R-P. Identification of residues in V-domain of CD80 (B7-1) implicated in functional interaction with CD28 and CTLA-4. J Exp Med. 1995;182:667–676. doi: 10.1084/jem.182.3.667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Peach R, Bajorath J, Naemura J, Leytze G, Green J-A, Aruffo A, Linsley PS. Both extracellular immunoglobulin-like domains of CD80 contains residues critical for binding T cell surface receptor CTLA-4 and CD28. J Biol Chem. 1995;270:21181–21187. doi: 10.1074/jbc.270.36.21181. [DOI] [PubMed] [Google Scholar]
- 39.Hara T, Fu SM, Hansen JA. Human T cell activation II. A new activation pathway used by a major T cell population via a disulfide-bonded dimer of a 44-kilodalton peptide (9.3 antigen) J Exp Med. 1985;161:1513–1524. doi: 10.1084/jem.161.6.1513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Moretta A, Pantaleo G, Lopez-Botet M, Moratta L. Involvement of T44 molecule in an antigen-independent pathway of T cell activation. J Exp Med. 1985;162:823–838. doi: 10.1084/jem.162.3.823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.June CH, Ledbetter JA, Gillispie MM, Linsten T, Thompson CB. T cell proliferation involving the CD28 pathway is associated with cyclosporine-resistant IL-2 gene expression. Mol Cell Biol. 1987;7:4472–4481. doi: 10.1128/mcb.7.12.4472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Harding F, McAuther J, Gross JA, Allison JP. CD28-mediated signalling costimulates murine T cells and prevents induction of anergy in T cells. Nature (Lond) 1992;356:607–609. doi: 10.1038/356607a0. [DOI] [PubMed] [Google Scholar]
- 43.Jenkins MK, Taylor PS K.B.Urdahl, and S.D. Norton. CD28 delivers costimulatory signals involved in antigen-specific IL-2 production by human T cells and prevents anergy. J Immunol. 1991;147:2461–2466. [PubMed] [Google Scholar]
- 44.Liu Y, Janeway CA., Jr Cells that present both specific ligand and costimulatory activity are the most efficient inducer of clonal expansion of normal CD4 T cells. Proc Natl Acad Sci USA. 1992;89:3845–3849. doi: 10.1073/pnas.89.9.3845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Chambers BJ, Salcedo M, Ljunggren H-G. Triggering of natural killer cells by the costimulatory molecule CD80 (B7-1) Immunity. 1996;5:311–317. doi: 10.1016/s1074-7613(00)80257-5. [DOI] [PubMed] [Google Scholar]
- 46.Linsley PS, Bradshaw J, Greene J, Peach R, Bennett KL, Mittler RS. Intracellular trafficking of CTLA-4 and focal localization towards sites of TCR engagement. Immunity. 1996;4:535–543. doi: 10.1016/s1074-7613(00)80480-x. [DOI] [PubMed] [Google Scholar]