

Abstract
Whereas amastigotes of the protozoan parasite Leishmania proliferate inside acidic phagolysosomal vacuoles of the macrophage, vacuoles induced by Leishmania donovani promastigotes during initiation of infection are poorly characterized. Here, evidence is presented that interaction of these parasitophorous vacuoles with endocytic organelles is very limited. In contrast, vacuoles formed around L. donovani mutants lacking the cell surface lipophosphoglycan (LPG) fuse extensively with endosomes and lysosomes. The role of LPG repeating units in the inhibition of phagosome–endosome fusion was demonstrated using two different approaches. First, genetic complementation of the LPG-defective C3PO mutant restored its ability to inhibit phagosome–endosome fusion to a degree similar to that of wild-type promastigotes. Second, opsonization of C3PO mutant cells with purified L. donovani LPG also conferred to this mutant the ability to inhibit phagosome–endosome fusion. Inasmuch as LPG is essential for infecting macrophages, these results suggest that inhibition of phagolysosomal biogenesis by LPG repeating units represents an intramacrophage survival strategy used by promastigotes to establish infection.
During their life cycle, protozoan parasites of the genus Leishmania alternate between the midgut of their insect vector, where they exist as promastigotes, and a macrophage phagolysosomal compartment, where they proliferate as amastigotes. Over the past several years, evidence was provided that Leishmania parasites succeed in avoiding host immune defenses and destruction by expressing specialized stage-specific molecules (1, 2). The dominant cell surface molecule of promastigotes is lipophosphoglycan (LPG)1, a glycosylinositolphospholipid (GPI)-anchored polymer consisting, in L. donovani, of the repeating disaccharide–phosphate unit [Gal(β1,4)Man(α1-PO4→ 6)] (between 16 to 30 units) (1). This molecule, particularly the repeating units moiety, is essential for the interaction of promastigotes with both the insect vector and the mammalian host (1, 3–11). The requirement for LPG in the establishment of infection inside the macrophage was evidenced by the demonstration that LPG repeating units–defective mutants are rapidly destroyed after phagocytosis, and that passive transfer of purified LPG significantly prolonged their survival (7, 8). This role for LPG repeating units for intramacrophage survival has also been genetically proven (11). Thus, without LPG repeating units promastigotes are unable to withstand the conditions prevailing inside the maturing parasitophorous vacuole. This is in contrast with the amastigotes, which proliferate inside acidic, hydrolase-rich vacuoles (12–17), despite the fact that they synthesize little or no detectable LPG (18–20). Thus, the role of this molecule in intramacrophage survival may be restricted to the establishment of infection, during promastigote-to-amastigote conversion (7–9, 11).
Different properties of LPG repeating units are consistent with their protective role during the establishment of infection, including efficient scavenging of toxic oxygen metabolites generated during the oxidative burst (8, 21), and modulation of the inducible nitric oxide synthase expression (22). In addition, the several million copies of LPG at the promastigote surface form a dense glycocalyx that may provide a physical barrier against the action of hydrolytic enzymes (10, 13). Recent in vitro studies demonstrated that incorporation of full-length LPG in lipid bilayers resulted in reduced fusogenic capacities (23). This finding raised the possibility that LPG repeating units may protect freshly phagocytized promastigotes by limiting the delivery of endosomal contents to the parasitophorous vacuoles, a process requiring multiple fusion events (24, 25). Here, we compared the ability of parasitophorous vacuoles induced by either wild-type L. donovani promastigotes or LPG repeating units–defective mutants to interact with endocytic organelles. We show that LPG repeating units enable Leishmania to inhibit the phagosome-endosome fusion process efficiently, thereby suggesting a survival strategy during their differentiation into amastigotes.
Materials and Methods
Cell Culture.
The murine macrophage cell line J774 was passaged in DMEM supplemented with 10% FCS, 1% glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin at 37°C in a 5% CO2 atmosphere. J774 cells were plated in 3-cm dishes for electron microscopy studies. All experiments were done with subconfluent cultures.
Parasites and LPG.
Leishmania donovani wild-type strain 1S (WT), the LPG-deficient mutants C3PO and RT5 (26), C3PO overexpressing LPG2 (C3PO+LPG2) (27), and the LPG repeating unit–defective line created by targeted disruption of LPG2 (termed lpg2 − knockout) (27) were all grown at 26°C in modified M199 medium as described (8). Opsonization of C3PO with purified L. donovani LPG (provided by S.J. Turco, University of Kentucky) was performed as described (8). All lines were used in stationary phase of growth.
Macrophage Infections, Electron Microscopy, and Morphological Studies.
Interactions and fusion between endocytic organelles and either L. donovani–containing phagosomes or latex bead–containing phagosomes were studied using a morphological approach at the electron microscope level. This in vivo fusion assay, based on the transfer of electron-dense tracers from endosomes to phagosomes, was used because of its simplicity over in vitro fusion assays and its sensitivity. Endocytic organelles were loaded by internalization of 5- and 16-nm gold particles coated with BSA (BSA– gold) using standard procedures (28). Phagosomes were formed by the addition of stationary-phase promastigotes at a parasiteto-host cell ratio of 10:1 in culture medium at 37°C for 60 min. Excess parasites were removed by four washes with cold PBS. Infected cells were then further incubated in culture medium for 60 min to allow complete internalization of bound promastigotes. Two types of loading procedures were used. In some cases, BSA– gold was internalized first, followed by incubation in culture medium to allow the marker to fill various endosome populations. Cells containing BSA–gold were then infected with L. donovani. This approach allowed us to study the interaction of early endosomes and lysosomes with newly formed phagosomes. The second procedure consisted in infecting macrophages with L. donovani first for 60 min, followed by incubation in culture medium for either 60 min or 4 h to form early or late phagosomes, respectively. These macrophages were then fed BSA–gold particles for 30 min and were either processed immediately or further incubated in culture medium for 60 min. This approach allowed us to study the interaction of preexisting phagosomes of various ages with maturing endosomes. For some experiments, promastigotes and 0.8-μm latex beads (Sigma, St. Louis, MO; diluted 1:200 in culture medium) were cointernalized, to study the interaction of endosomes with different phagosome populations. After all internalization steps, cells were washed thoroughly with cold PBS three times for 5 min. At the end of each experiment, cells were fixed in 1% glutaraldehyde, postfixed in OsO4, processed for embedding in Epon 812 resin as described previously (24), and observed by electron microscopy.
Intermixing of BSA–gold particles and L. donovani was then recorded for each combination of incubations performed. The presence of a single gold particle inside a phagosome was scored as a fusion event. For the analysis of fusion occurrence (Fig. 2), each experiment was done at least twice and a minimum of 25 Leishmania-containing phagosomes per timepoint were recorded, while extreme care was taken to avoid serial sections. For the quantitative analysis of the transfer of BSA–gold particles from endosomes to Leishmania-containing phagosomes (Table 1), the number of 5- and 16-nm gold particles was counted on at least 12–20 of the BSA–gold positive phagosome profiles at the electron microscope.
Figure 2.
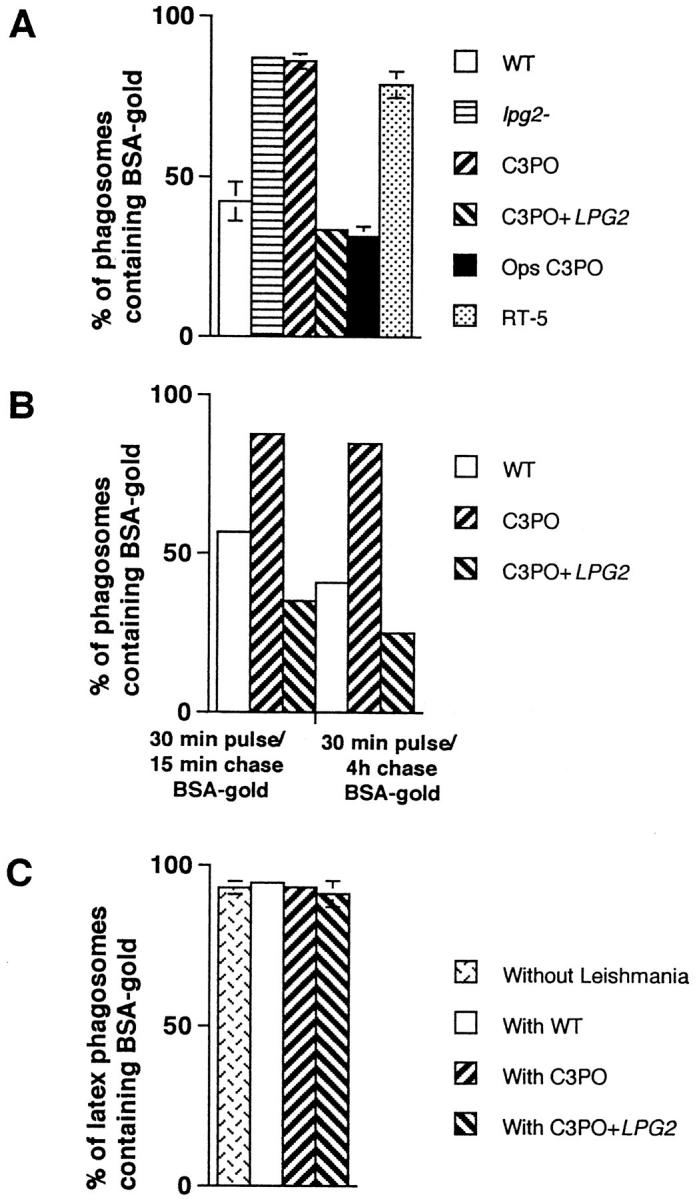
Quantitative analysis of the fusion between phagosomes containing L. donovani parasites and endocytic organelles. (A) Parasites and BSA–gold were internalized as described in Fig. 1. Macrophages were prepared for electron microscopy and quantitative analysis of phagosome– endosome fusion performed on thin sections. The presence of a single gold particle in a parasitophorous vacuole was scored as a fusion event. These results represent the average of three to six experiments. Ops C3PO, LPG-opsonized C3PO. (B) Macrophages were fed a 5- and 16-nm BSA–gold particles mixture for 30 min, followed by either a 15-min or a 4-h incubation period to fill endosomes and lysosomes, respectively. Labeled macrophages were then infected with promastigotes for 60 min and further incubated for 60 min. Fusion was assessed as above. These results represent the average of two experiments. (C) Fusion properties of latex bead–containing phagosomes present in cells coinfected with various parasites (see Fig. 4). Latex bead phagosomes were analyzed only in cells showing the presence of a parasite on the section profile. These results represent the average of three to four experiments.
Table 1.
>Quantification of BSA–gold Particles Transferred from Endosomes to Leishmania-containing Phagosomes
| 5-nm BSA–gold particles | 16-nm BSA–gold particles | |||
| Wild type | 57.3 ± 28.0 | 4.3 ± 2.7 | ||
| C3PO | 323.6 ± 142.5 | 23.6 ± 11.4 | ||
| C3PO+LPG2 | 86.2 ± 34.4 | 8.2 ± 3.5 |
J774 macrophages were infected as described in the legend to Fig. 1. Cells were then fed a 5-nm and 16-nm BSA–gold particles mixture for 30 min, followed by a chase period of 60 min, and processed for electron microscopy. The 5- and 16-nm gold particles present in the profile section of phagosomes containing the various Leishmania lines were counted. At least 25 phagosomes were analyzed for each cell line. Phagosomes devoid of BSA–gold particles were not included in this quantitative analysis.
Results and Discussion
The transfer of BSA–gold particles from endosomes to phagosomes containing either wild-type promastigotes or the LPG repeating unit–defective C3PO mutant cells (26) was first used to evaluate the interaction occurring between these organelles. Very few gold particles were generally transferred to WT–containing phagosomes despite the close presence of BSA–gold particle–filled endosomes (Fig. 1 A). In contrast, most phagosomes containing the LPG repeating units–defective C3PO mutant were filled with numerous BSA–gold particles (Fig. 1 B). While only ∼40% of WTphagosomes received BSA–gold particles, >90% of phagosomes induced by C3PO mutant cells fused with endocytic organelles (Fig. 2 A). The difference in levels of fusion was more striking when the number of gold particles transferred from endosomes to phagosomes was quantified. Indeed, a sixfold increase in the BSA–gold particle content of C3PO phagosomes over WT-phagosomes was observed (Table 1). High levels of fusion were also observed with phagosomes induced by lpg2 − null mutant cells, which are phenotypically identical to C3PO cells (27), and by RT5, a mutant that accumulates truncated forms of LPG containing 3–5 repeating units (26) (Fig. 2 A). Because assembly of the Gal(β1,4)Man(α1-PO4→ 6) repeating units is defective in these mutants, our observations indicated a role for this LPG moiety in the inhibition of phagosome–endosome fusion. Further, immunofluorescence analysis showed that LPGexpressing promastigotes are contained within a LAMP1negative compartment, whereas LPG-deficient mutants were found in LAMP1-positive phagosomes (data not shown), indicating their respective levels of fusion with endocytic organelles enriched for LAMP1 molecules.
Figure 1.
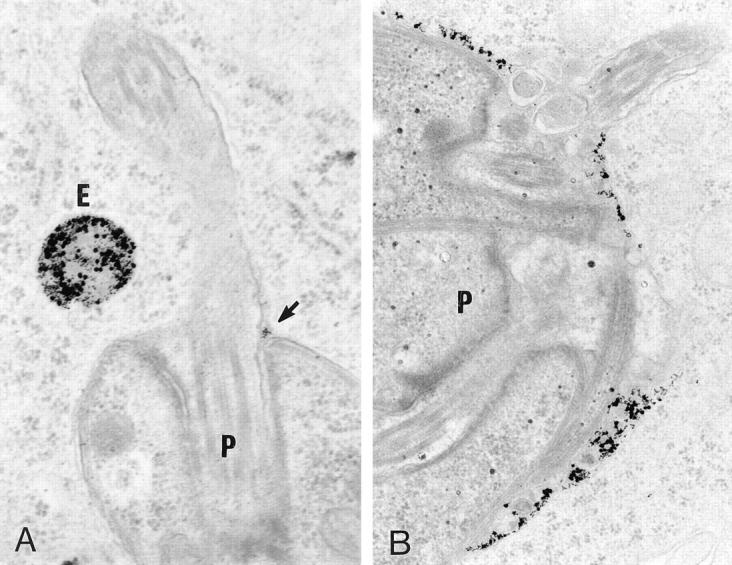
Fusion of L. donovani–containing phagosomes with newly formed endosomes. J774 macrophages were infected with stationary-phase promastigotes for 60 min and further incubated for 60 min. Cells were then fed a 5- and 16-nm BSA–gold particles mixture for 30 min, followed by a chase period of 60 min, and processed for electron microscopy. (A) Phagosomes containing wild-type Leishmania (P) display low fusogenic properties with endocytic organelles (E). When fusion occurred, only few gold particles were observed in the phagosome (arrow). (B) Phagosomes containing the LPG-deficient C3PO mutant (P) were filled with large number of BSA– gold particles, indicating their acute ability to fuse with endosomes.
To demonstrate directly a role for LPG in the inhibition of phagosome–endosome fusion, two distinct approaches were taken. First, we assessed the fusogenic properties of phagosomes induced by C3PO cells transfected with a LPG2 expression construct (C3PO+LPG2) (27). As expected, phagosomes containing C3PO+LPG2 cells behaved similarly to WT-phagosomes with respect to their ability to fuse with endocytic organelles (Fig. 2 A; and Table 1). Thus, restoration of full-length LPG repeating units synthesis by genetic complementation enabled C3PO to inhibit phagosome–endosome fusion. Because LPG2 expression in C3PO also restores repeating units addition on secreted molecules, including the secreted acid phosphatase (27), the possibility existed that such repeating unit addition could have accounted for the ability of C3PO+LPG2 cells to inhibit phagosome–endosome fusion. Therefore, in a second approach, C3PO cells were opsonized with purified L. donovani LPG (8) before infection of J774 macrophages. Effectiveness of the opsonization procedure was confirmed by immunofluorescence (data not shown) with the antirepeating units mAb CA7AE (29). Electron microscopy analysis revealed that phagosomes containing LPG-opsonized C3PO cells displayed low fusion properties similar to that of WT-phagosomes (Fig. 2 A; Fig. 3). This data provided a direct evidence that the sole presence of LPG repeating units at the promastigote surface is sufficient to inhibit phagosome–endosome fusion. Therefore, repeating unit modification of the secreted acid phosphatase, which is not required for both secretion and activity (27, 30), appears to be without apparent effect on phagosomal fusogenic properties.
Figure 3.
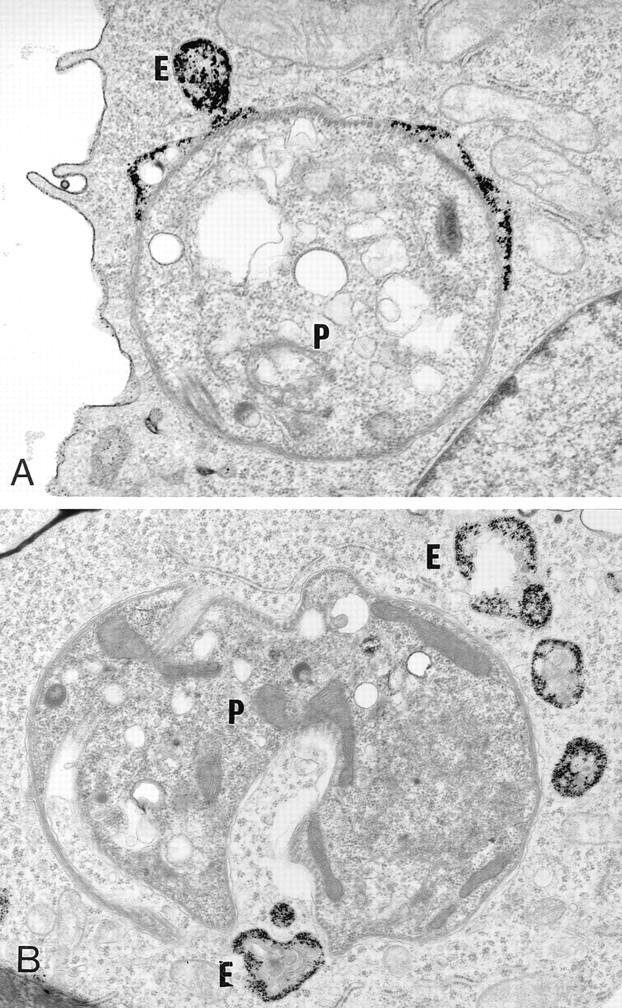
Fusion properties of LPG-opsonized C3PO cells. Cells were infected with the C3PO mutant (A) or with LPG-opsonized C3PO cells (B). Phagosomes containing the C3PO mutant engage in fusion event with endosomes (E) and contain a large number of gold particles. In contrast, phagosomes containing LPG-opsonized C3PO mutants are devoid of gold particles, despite the presence of BSA–gold particles–filled endosomes (E) in the parasitophorous vacuole surrounding.
We also assessed the ability of newly formed phagosomes to fuse with two distinct endosome populations. Here, BSA– gold particles were first internalized for 30 min, followed by either a 15-min or 4-h chase. The first population is considered as early endosomes and the second as lysosomes. At the end of the chase period, macrophages were infected with either WT promastigotes or C3PO mutant cells for 1 h. Increased fusion between early endosomes and C3PO/phagosomes with respect to WT-phagosomes was observed (see Fig. 2 B). Interestingly, WT-phagosomes consistently displayed lower fusogenicity with lysosomes (35% fusion) than with early endosomes (55% fusion), suggesting that, similar to Mycobacterium phagosomes (31–34), fusion with the various endosomal populations might be selective, albeit to a lesser extent.
To evaluate the specificity of the LPG repeating units– induced alteration(s) of phagosome fusion properties, J774 macrophages were coinfected with latex beads and the various Leishmania lines. Latex beads and promastigotes were present in distinct phagosomal compartments (Fig. 4). Regardless of the cointernalized Leishmania line, the fusion rate between endocytic organelles and latex bead phagosomes was around 90% (see Fig. 2 C). Thus, in a given macrophage, fusion can occur between endosomes and latex bead phagosomes but not with WT-phagosomes. This observation indicated that (a) inhibition of fusion by LPG repeating unit is selective, (b) the general host cell fusion machinery remains operational during infection, and (c) LPG repeating units selectively alter local phagosomal fusogenic properties. From a mechanistic point of view, the following models can be considered. First, WT promastigotes and the repeating unit–defective mutants use distinct receptors for attachment and entry, resulting in the formation of phagosomes with different biochemical composition and fusion properties (35–37). However, the observation that promastigote-to-amastigote transformation, which is paralleled by the loss of LPG, restores phagosome–endosome fusion argues against this idea (12–20). In the second model, LPG may inhibit activation of phagosome-associated protein kinase C (PKC) (38–40). Although the precise function of PKC in phagosome–endosome interaction remains speculative, it is known to phosphorylate MARCKS, a membrane protein associated with actin-based motility (41) and with membrane trafficking (42). In this regard, PKC-dependent phosphorylation of phagosome-associated MARCKS results in its displacement from the membrane to Lamp1positive lysosomes (42) and may therefore participate in the movement of both phagosomes and endosomes on microtubules. Inhibition of PKC-dependent MARCKS phosphorylation by LPG (43–44) may block this movement. However, this model is not consistent with the inhibition of PKC-dependent processes by C3PO and amastigotes during infection (43, 45, 46), both of which induce phagosomes that fuse freely with endocytic organelles (see Figs. 1 and 2 A; references 12–17). In a third model, LPG is transferred within minutes from the promastigote surface to the macrophage membrane at the immediate area of internalization (29). Insertion of LPG in lipid-bilayer membranes stabilizes the bilayer against the formation of an inverted hexagonal structure, resulting in reduced fusogenic properties (23). As a consequence, LPG would give rise to an effective steric repulsion between phagosomal and endosomal membranes or reduce the negative curvature strain in bilayers, increasing the energy barrier for forming highly curved fusion intermediates (23), thereby preventing fusion. Interestingly, truncated forms of LPG containing few repeating units are ineffective in modifying the fusogenic properties of membranes (23). This in vitro finding is in agreement with our observation that RT5, a mutant expressing truncated forms of LPG with three to five repeating units (26), is unable to inhibit phagosome–endosome fusion. Alternatively, the possibility exists that LPG inhibits phagosome–endosome fusion indirectly. Indeed, LPG may prevent parasite destruction by scavenging toxic oxygen radicals generated during the oxidative burst (8, 21), allowing for the production of a yet unidentified factor directly responsible for preventing fusion. Regardless of the nature of the exact underlying mechanism, our data have unequivocally demonstrated that expression of full-length LPG enables promastigotes to inhibit phagosome–endosome fusion. The extent to which inhibition of fusion by LPG contributes to promastigote survival remains to be determined.
Figure 4.
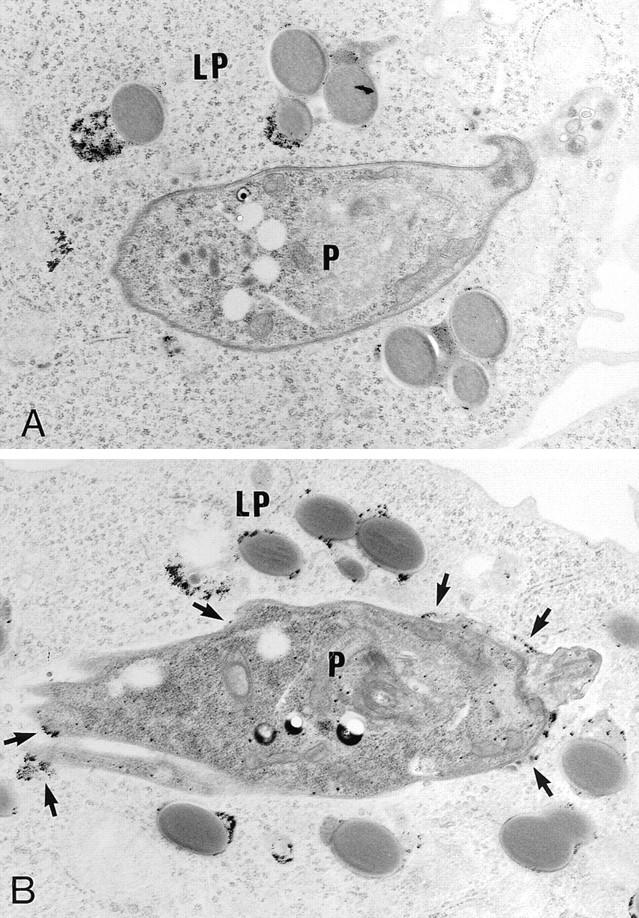
Fusion properties of phagosomes containing Leishmania parasites or latex bead particles. Promastigotes and 0.8-μm latex beads (Sigma, St. Louis, MO; diluted 1:200 in culture medium) were cointernalized. BSA–gold particles were then internalized for 30 min, followed by a 60- min incubation. (A) Latex bead–containing phagosomes (LP) present in macrophages infected with wild type contain large amount of BSA–gold particles, whereas phagosomes containing the parasites (P) are in most cases devoid of gold particles. (B) In contrast, both latex bead– and C3PO-containing phagosomes are filled with gold particles.
In a study aimed at determining the position of L. mexicana parasitophorous vacuoles within the endocytic network of the host macrophage, Russell et al. (16) observed that the transfer of endosomal content to parasitophorous vacuoles induced by promastigotes increased in efficiency with respect to the age of the infection. To explain this phenomenon, the authors suggested that these vacuoles had functionally translocated from a lysosomal to a late endosomal compartment, consistent with their increased content in mannose-6-phosphate receptors. In the light of the findings reported herein, we suggest that the reduced fusogenicity of the parasitophorous vacuoles observed in early infections (day 2) by Russell et al. (16) is caused by the presence of promastigote-derived LPG. Indeed, LPG repeating units epitopes are maximally present in the macrophage membrane 1–2 d after infection, and by 5–6 d, these epitopes are no longer detectable (29). Thus, the transformation of promastigotes into amastigotes, associated with the loss of LPG expression, occurs concomitently with the release of phagosome–endosome fusion inhibition.
Inhibition of phagosome–endosome fusion is an intramacrophage survival strategy used by a variety of intracellular pathogens (35–37). Although the molecular bases of this phenomenon are poorly understood, microbial surface components analogous to the Leishmania LPG may alter the fusion properties of the endocytic system (47). In this regard, in vitro studies have shown that Cord factor (α,α-trehalose 6,6′-dimycolate), a cell wall glycolipid of Mycobacteria, inhibits fusion between phospholipid vesicles (48). Although exclusion of the vacuolar proton–ATPase from mycobacteria–phagosomes appears to be closely related with their reduced fusogenicity, it is not known, however, whether this exclusion is the consequence or the cause of the altered fusion properties (31).
The mechanisms and molecules involved in the regulation of phagosome fusion properties remain largely unknown. While proteins of the rab family were found to associate and dissociate from maturing phagosomes, their role in phagosome fusion is still unclear (24). Phagosome maturation is also accompanied by the acquisition of Nramp1, a molecule conferring resistance to a variety of intracellular microbes, including L. donovani (49), the sequential phosphorylation and dephosphorylation of some of their proteins, as well as modifications of their phospholipid content (50, 51). Although no direct correlation between phagosome fusogenic properties and their lipid composition has been demonstrated, the finding that LPG modifies the molecular structure of lipid bilayers (23) suggests a role for lipids in the fine tuning of membrane fusion events. In this regard, we expect that molecular and biochemical characterization of Leishmania-containing phagosomes will yield novel insights on the mechanistic aspects of membrane fusion.
LPG repeating units are essential for the successful transition of Leishmania parasites from the sandfly midgut to the inside of a macrophage phagolysosome (3–11). In the macrophage, LPG repeating units may contribute to promastigote survival through their capacity to scavenge oxygen radicals (21), to inhibit PKC (40, 44), and to modulate nitric oxide synthase expression (22). Our data suggest a novel function for LPG repeating units during the early phase of macrophage infection, where inhibition of phagolysosomal biogenesis may protect invading promastigotes from hydrolytic degradation and provide an environment propitious for their differentiation into amastigotes. Finally, the anti-fusogenic properties of LPG repeating units provide a powerful probe to investigate the regulation of phagosome–endosome fusion during microbial invasion.
Acknowledgments
We acknowledge the technical assistance of C. Rondeau and J. Léveillé. We are grateful to S.J. Turco (University of Kentucky) and S.M. Beverley (Harvard Medical School) for authorizing the use of C3PO, C3PO+LPG2, and lpg2 −. We thank S.J. Turco for kindly providing RT-5 cells and LPG purified from Leishmania donovani and for suggestions, and R. Nabi, G. Matlashewski, and R. Epand for comments and critical reading of the manuscript.
This work was supported by grants from the Medical Research Council (MRC) of Canada to M. Desjardins (MT-12951) and A. Descoteaux (MT-12933). M. Desjardins is a Scholar from the Fonds de la recherche en santé du Québec and A. Descoteaux is a Scholar from the MRC.
Footnotes
1 Abbreviations used in this paper: LPG, lipophosphogylcan; GPI, glycosylinositolphospholipid; PKC, protein kinase C; WT, wild-type.
References
- 1.Turco SJ, Descoteaux A. The lipophosphoglycan of Leishmaniaparasites. Annu Rev Microbiol. 1992;46:65–94. doi: 10.1146/annurev.mi.46.100192.000433. [DOI] [PubMed] [Google Scholar]
- 2.McConville MJ, Ferguson MAJ. The structure, biosynthesis and function of glycosylated phosphatidylinositols in the parasitic protozoa and higher eukaryotes. Biochem J. 1993;294:305–324. doi: 10.1042/bj2940305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pimenta PFP, Turco SJ, McConville MJ, Lawyer P, Perkins P, Sacks DL. Stage-specific adhesion of Leishmaniapromastigotes to the sand fly midgut. Science (Wash DC) 1992;256:1812–1815. doi: 10.1126/science.1615326. [DOI] [PubMed] [Google Scholar]
- 4.Pimenta PFP, Saraiva EMB, Rowton E, Modi GB, Garraway LA, Beverley SM, Turco SJ, Sacks DL. Evidence that the vectorial competence of phlebotomine sand flies for different species of Leishmaniais controlled by structural polymorphisms in the surface lipophosphoglycan. Proc Natl Acad Sci USA. 1994;91:9155–9159. doi: 10.1073/pnas.91.19.9155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Butcher BA, Turco SJ, Hilty BA, Pimenta PF, Panunzio M, Sacks DL. Deficiency in β1,3-galactosyltransferase of a Leishmania major lipophosphoglycan mutant adversely influences the Leishmania–sand fly interaction. J Biol Chem. 1996;271:20573–20579. doi: 10.1074/jbc.271.34.20573. [DOI] [PubMed] [Google Scholar]
- 6.Culley FJ, Harris RA, Kaye PM, McAdam KPWJ, Raynes JG. C-reactive protein binds to a novel ligand on Leishmania donovaniand increases uptake into human macrophages. J Immunol. 1996;156:4691–1696. [PubMed] [Google Scholar]
- 7.Handman E, Schnur LF, Spithill TW, Mitchell GF. Passive transfer of Leishmanialipopolysaccharide confers parasite survival in macrophages. J Immunol. 1986;137:3608–3613. [PubMed] [Google Scholar]
- 8.McNeely TB, Turco SJ. Requirement of lipophosphoglycan for intracellular survival of Leishmania donovaniwithin human monocytes. J Immunol. 1990;144:2475–2750. [PubMed] [Google Scholar]
- 9.Elhay M, Kelleher M, Bacic A, McConville MJ, Tolson DL, Pearson TW, Handman E. Lipophosphoglycan expression and virulence in ricin-resistant variants of Leishmania major. . Mol Biochem Parasitol. 1990;40:255–268. doi: 10.1016/0166-6851(90)90047-p. [DOI] [PubMed] [Google Scholar]
- 10.Eilam Y, El-On J, Spira DT. Leishmania major: excreted factor, calcium ions, and the survival of amastigotes. Exp Parasitol. 1985;59:161–168. doi: 10.1016/0014-4894(85)90068-2. [DOI] [PubMed] [Google Scholar]
- 11.Beverley, S.M., and S.J. Turco. 1995. Identification of genes mediating lipophosphoglycan biosynthesis by functional complementation of Leishmania donovani mutants. Ann. Trop. Med. Parasitol. 89 (Suppl.):11–17. [DOI] [PubMed]
- 12.Alexander J, Vickerman K. Fusion of host cell secondary lysosomes with the parasitophorous vacuoles of Leishmania mexicana–infected macrophages. J Protozool. 1975;22:502–508. doi: 10.1111/j.1550-7408.1975.tb05219.x. [DOI] [PubMed] [Google Scholar]
- 13.Chang K-P, Dwyer DM. Multiplication of a human parasite (Leishmania donovani)in phagolysosomes of hamster macrophages in vitro. Science (Wash DC) 1976;193:678–680. doi: 10.1126/science.948742. [DOI] [PubMed] [Google Scholar]
- 14.Antoine J-C, Prina E, Jouanne C, Bongrand P. Parasitophorous vacuoles of Leishmania amazonensis–infected macrophages maintain an acidic pH . Infect Immun. 1990;58:779–787. doi: 10.1128/iai.58.3.779-787.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Prina E, Antoine J-C, Wiederanders B, Kirschke H. Localization and activity of various lysosomal proteases in Leishmania amazonensis–infected macrophages. Infect Immun. 1990;58:1730–1737. doi: 10.1128/iai.58.6.1730-1737.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Russell DG, Xu S, Chakraborty P. Intracellular trafficking and the parasitophorous vacuole of Leishmania mexicana–infected macrophages. J Cell Science. 1992;103:1193–1210. doi: 10.1242/jcs.103.4.1193. [DOI] [PubMed] [Google Scholar]
- 17.Veras PST, de Chastellier C, Rabinovitch M. Transfer of zymosan (yeast cell walls) to the parasitophorous vacuoles of macrophages infected with Leishmania amazonensis. . J Exp Med. 1992;176:639–646. doi: 10.1084/jem.176.3.639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.McConville MJ, Blackwell JM. Developmental changes in the glycosylated phosphatidylinositols of Leishmania donovani: characterization of the promastigote and amastigote glycolipids. J Biol Chem. 1991;266:15170–15179. [PubMed] [Google Scholar]
- 19.Turco SJ, Sacks DL. Expression of a stage-specific lipophosphoglycan in Leishmania majoramastigotes. Mol Biochem Parasitol. 1991;45:91–100. doi: 10.1016/0166-6851(91)90030-a. [DOI] [PubMed] [Google Scholar]
- 20.Moody SF, Handman E, McConville MJ, Bacic A. The structure of Leishmania majoramastigote lipophosphoglycan. J Biol Chem. 1993;268:18457–18466. [PubMed] [Google Scholar]
- 21.Chan J, Fujira T, Brennan P, McNeil M, Turco SJ, Sibille J, Snapper M, Aisen P, Bloom BR. Microbial glycolipids: possible virulence factors that scavenge oxygen radicals. Proc Natl Acad Sci USA. 1989;86:2453–2457. doi: 10.1073/pnas.86.7.2453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Proudfoot L, Nikolaev AV, Feng G-J, Wei X-Q, Ferguson MAJ, Brimacombe JS, Liew FY. Regulation of the expression of nitric oxide synthase and leishmanicidal activity by glycoconjugates of Leishmanialipophosphoglycan in murine macrophages. Proc Natl Acad Sci USA. 1996;93:10984–10989. doi: 10.1073/pnas.93.20.10984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Miao L, Stafford A, Nir S, Turco SJ, Flanagan TD, Epand RM. Potent inhibition of viral fusion by the lipophosphoglycan of Leishmania donovani. . Biochemistry. 1995;34:4676–4683. doi: 10.1021/bi00014a022. [DOI] [PubMed] [Google Scholar]
- 24.Desjardins M, Huber LA, Parton RG, Griffiths G. Biogenesis of phagolysosomes proceeds through a sequential series of interactions with endocytic apparatus. J Cell Biol. 1994;124:677–688. doi: 10.1083/jcb.124.5.677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Desjardins M. Biogenesis of phagolysosomes: the ‘kiss and run' hypothesis. Trends Cell Biol. 1995;5:183–186. doi: 10.1016/s0962-8924(00)88989-8. [DOI] [PubMed] [Google Scholar]
- 26.McNeely TB, Tolson DL, Pearson TW, Turco SJ. Characterization of Leishmania donovanivariant clones using anti-lipophosphoglycan monoclonal antibodies. Glycobiology. 1990;1:63–69. doi: 10.1093/glycob/1.1.63. [DOI] [PubMed] [Google Scholar]
- 27.Descoteaux A, Luo Y, Turco SJ, Beverley SM. A specialized pathway affecting virulence glycoconjugates of Leishmania. . Science (Wash DC) 1995;269:1869–1872. doi: 10.1126/science.7569927. [DOI] [PubMed] [Google Scholar]
- 28.Rabinowitz S, Horstmann H, Gordon S, Griffiths G. Immunocytochemical characterization of the endocytic and phagolysosomal compartments in peritoneal macrophages. J Cell Biol. 1992;116:95–112. doi: 10.1083/jcb.116.1.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Tolson DL, Turco SJ, Pearson TW. Expression of a repeating phosphorylated disaccharide lipophosphoglycan epitope on the surface of macrophages infected with Leishmania donovani. . Infect Immun. 1990;58:3500–3507. doi: 10.1128/iai.58.11.3500-3507.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bates PA, Hermes I, Dwyer DM. Golgi-mediated post-translational processing of secretory acid phosphatase by Leishmania donovanipromastigotes. Mol Biochem Parasitol. 1990;39:247–256. doi: 10.1016/0166-6851(90)90063-r. [DOI] [PubMed] [Google Scholar]
- 31.Sturgill-Koszycki S, Schlesinger PH, Chakraborty P, Haddix PL, Collins HL, Fok AK, Allen RD, Gluck SL, Heuser J, Russell DG. Lack of acidification in Mycobacteriumphagosomes produced by exclusion of the vesicular proton–ATPase. Science (Wash DC) 1994;263:678–681. doi: 10.1126/science.8303277. [DOI] [PubMed] [Google Scholar]
- 32.Russell DG, Dante J, Sturgill-Koszycki S. Mycobacterium avium– and Mycobacterium tuberculosis–containing vacuoles are dynamic, fusion competent vesicles that are accessible to glycosphingolipids from the host cell plasmalemma. J Immunol. 1996;156:4764–4773. [PubMed] [Google Scholar]
- 33.Clemens DL, Horwitz MA. The Mycobacterium tuberculosisphagosome interacts with early endosomes and is accessible to exogenously administered transferrin. J Exp Med. 1996;184:1349–1355. doi: 10.1084/jem.184.4.1349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.de Chastellier C, Lang T, Thilo L. Phagocytic processing of the macrophage endoparasite, Mycobacterium avium, in comparison to phagosomes which contain Bacillus subtilisor latex beads. Eur J Cell Biol. 1995;68:167–182. [PubMed] [Google Scholar]
- 35.Joiner KA, Fuhrman SA, Miettinen HM, Kasper LH, Mellman I. Toxoplasma gondii: fusion competence of parasitophorous vacuoles in Fc receptor-transfected fibroblasts. Science (Wash DC) 1990;249:641–646. doi: 10.1126/science.2200126. [DOI] [PubMed] [Google Scholar]
- 36.Small PLC, Ramakrishnan L, Falkow S. Remodeling schemes of intracellular pathogens. Science (Wash DC) 1994;263:637–639. doi: 10.1126/science.8303269. [DOI] [PubMed] [Google Scholar]
- 37.Finlay BB, Falkow S. Common themes in microbial pathogenicity. Microbiol Rev. 1989;53:210–230. doi: 10.1128/mr.53.2.210-230.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Descoteaux A, Turco SJ. The lipophosphoglycan of Leishmaniaand macrophage protein kinase C. Parasitol Today. 1993;9:468–471. doi: 10.1016/0169-4758(93)90105-o. [DOI] [PubMed] [Google Scholar]
- 39.Allen L-AH, Aderem A. A role for MARCKS, the α isoenzyme of protein kinase C and myosin I in zymosan phagocytosis by macrophages. J Exp Med. 1995;182:829–840. doi: 10.1084/jem.182.3.829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Giorgione JR, Turco SJ, Epand RM. Transbilayer inhibition of protein kinase C by the lipophosphoglycan from Leishmania donovani. . Proc Natl Acad Sci USA. 1996;93:11634–11639. doi: 10.1073/pnas.93.21.11634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rosen A, Keenan KF, Thelen M, Nairn AC, Aderem AA. Activation of protein kinase C results in the displacement of its myristoylated, alanine-rich substrate from punctate structures in macrophage filopodia. J Exp Med. 1990;172:1211–1215. doi: 10.1084/jem.172.4.1211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Allen L-AH, Aderem A. Protein kinase C regulates MARCKS cycling between the plasma membrane and lysosomes in fibroblasts. EMBO (Eur Mol Biol Organ) J. 1995;14:1109–1121. doi: 10.1002/j.1460-2075.1995.tb07094.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Descoteaux A, Matlashewski G, Turco SJ. Inhibition of macrophage protein kinase C–mediated protein phosphorylation by Leishmania donovanilipophosphoglycan. J Immunol. 1992;149:3008–3015. [PubMed] [Google Scholar]
- 44.Descoteaux A, Turco SJ, Sacks DL, Matlashewski G. Leishmania donovanilipophosphoglycan selectively inhibits signal transduction in macrophages. J Immunol. 1991;146:2747–2753. [PubMed] [Google Scholar]
- 45.Descoteaux A, Matlashewski G. C-fos and TNF gene expression in Leishmania donovani–infected macrophages. Mol Cell Biol. 1989;9:5223–5227. doi: 10.1128/mcb.9.11.5223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Olivier M, Brownsey RW, Reiner NE. Defective stimulus–response coupling in human monocytes infected with Leishmania donovaniis associated with altered activation and translocation of protein kinase C. Proc Natl Acad Sci USA. 1992;89:7481–7485. doi: 10.1073/pnas.89.16.7481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Goren MB, D'Arcy P, Hart, Young MR, Armstrong JA. Prevention of phagosome–lysosome fusion in cultured macrophages by sulfatides of Mycobacterium tuberculosis. . Proc Natl Acad Sci USA. 1976;73:2510–2514. doi: 10.1073/pnas.73.7.2510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Spargo BJ, Crowe LM, Ioneda T, Beaman BL, Crowe JH. Cord factor (α,α-trehalose 6,6′-dimycolate) inhibits fusion between phospholipid vesicles. Proc Natl Acad Sci USA. 1991;88:737–740. doi: 10.1073/pnas.88.3.737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gruenheid S, Pinner E, Desjardins M, Gros P. Natural resistance to infection with intracellular parasites: the Nramp1 protein is recruited to the membrane of the phagosome. J Exp Med. 1997;185:717–730. doi: 10.1084/jem.185.4.717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Desjardins M, Celis JE, van Meer G, Dieplinger H, Jahraus A, Griffiths G, Huber LA. Molecular characterization of phagosomes. J Biol Chem. 1994;269:32194–32200. [PubMed] [Google Scholar]
- 51.Emans N, Nzala NN, Desjardins M. Protein phosphorylation during phagosome maturation. FEBS Lett. 1996;398:37–42. doi: 10.1016/s0014-5793(96)01213-6. [DOI] [PubMed] [Google Scholar]