

Abstract
The establishment of culture conditions that selectively support hematopoietic stem cells is an important goal of hematology. In this study, we investigated the possibility of using for this purpose a defined medium, mSFO2, which was developed for stromal cell–dependent bone marrow cultures. We found that a combination of epidermal growth factor (EGF), the OP9 stromal cell line, which lacks macrophage colony-stimulating factor, recombinant stem cell factor, and the chemically defined medium mSFO2 provides a microenvironment where c-Kit+ Thy-1+/lo Mac-1+/lo B220− TER119− commonβ+ IL-2Rγ + gp130+ cells are selectively propagated from normal, unfractionated bone marrow cells. This cell population produced an in vitro colony at a very high efficiency (50%), whereas it has only limited proliferative ability in the irradiated recipient. Thus, the cells selected in this culture condition might represent colony-forming units in culture (CFU-c) with short-term reconstituting ability. Transferring this cell population into medium containing differentiation signals resulted in the rapid production of mature myelomonocytic and B cell lineages in vitro and in vivo. The fact that a similar culture condition was created by erb-B2–transduced OP9 in the absence of EGF indicated that EGF exerts its effect by acting on OP9 rather than directly on CFU-c. These results suggested that the balance between self-renewal and differentiation of CFU-c can be regulated by extracellular signals.
An important goal of experimental hematology is to manipulate the proliferation and differentiation of the normal hematopoietic stem cells. As an approach to this end, extensive efforts have been made to establish cultures where the growth of hematopoietic stem cells is maintained. The stromal cell–dependent, long-term bone marrow (BM)1 culture developed by Dexter and his colleagues (1) has been the most successful means of maintaining hematopoietic stem cells in vitro. Although the sustained selfrenewal of the multipotent stem cells is supported in this system, concomitant terminal differentiation into the myelomonocytic cell lineage does occur. As a result of this differentiation in culture, a minor population of multipotent stem cells persists, and the production of hematopoietic cells eventually ceases because of the imbalance between self-renewal and differentiation of the stem cells. Thus, any method that alters this balance in favor for self-renewal might result in a culture where hematopoietic progenitors are spontaneously enriched. In fact, this has been attained in cultured embryonic stem cells using leukemia inhibitory factor as a molecular cue (2, 3).
One way to change the balance in favor of self-renewal is to block the intracellular machinery that drives the hematopoietic stem cell to differentiate. However, once the intracellular machinery for cell differentiation is artificially locked, it is difficult to release it physiologically. Alternatively, extensive attempts have been made to determine the growth factor combination that allows the proliferation of stem cells while arresting cell differentiation (4–13), as leukemia inhibitory factor does to the embryonic stem cells. Moreover, a number of untransformed stem cell lines, some of which have been claimed to give rise to the whole range of hematopoietic cell lineage, have been described (14–16). However, those cell lines have not been generally applied for two main reasons. Most, if not all, of these stem cell lines tend to lose sensitivity to differentiation signals, though a small proportion of them may differentiate upon stimulation either in vitro or in vivo. Moreover, it is usually difficult to reproduce the culture conditions described in the original reports because of the presence of chemically undefined ingredients in the media.
In this study, we investigated ways to overcome this problem. We reduced the differentiation factors present in ordinal long-term BM culture using the defined culture medium mSFO2, which contains transferrin and insulin as the only protein components, as well as the macrophage CSF (M-CSF)–negative stromal cell clone OP9.
Materials and Methods
Cell Preparation.
The stomal cell line OP9 was maintained in α-MEM (GIBCO BRL, Gaithersburg, MD) containing 20% FCS (lot No. 1M1250; BioWhittaker, Walkersville, MD), as reported (17). The subline OP9/erbB2 that can survive in serumfree media without external growth factors was generated by means of calcium phosphate transfection performed essentially as reported (18). Briefly, exponentially growing OP9 cells were seeded at a density of 5 × 105 cells on 10-cm tissue culture plates in α-MEM with 20% FCS 1 d before transfection. On the day of transfection, calcium phosphate DNA solution was added dropwise into the medium, and the cells were incubated for 24 h in a 37°C, 5% CO2 incubator. The OP9 cells were transformed by cotransfection with the pSV2erbB2VE plasmid containing the constitutive active phosphorylation site of erbB2 (19–21) in the cytoplasmic domain (a gift from T. Yamamoto, Institute of Medical Science, University of Tokyo, Tokyo, Japan), as well as the pSV2bsr plasmid vector (Funakoshi Co., Tokyo, Japan) containing the resistance gene for blasticidin S hydrochloride (Funakoshi Co.).
BM cells harvested from normal C57BL/6 mice were suspended in RPMI 1640 medium (GIBCO BRL) with 10% FCS and passed through Sephadex G-10 (Pharmacia , Uppsala, Sweden) twice to deplete stromal cells and macrophages (22, 23). Stromal cell–depleted BM cells (105) were seeded onto OP9 stromal cells under various conditions: in 3 ml of RPMI 1640 containing 10% FCS and 0.1% BSA, or in 3 ml of mSFO2 supplemented with either recombinant (r) epidermal growth factor (EGF; 30 ng/ml; Peprotech Inc., Rocky Hill, NJ) or recombinant basic fibroblast growth factor (rbFGF, 10 ng/ml; GIBCO BRL) with or without recombinant stem cell factor (rSCF; 100 ng/ml; reference 24) on PRIMARIA culture plates measuring 3.5 cm in diameter (Becton Dickinson Labware, Lincoln Park, NJ). The concentration of SCF was decided based on a previous report (13). We omitted 2-ME from all conditions. To remove the effect of serum used for maintaining OP9 cells, confluent OP9 cells were cultured in mSFO2 medium containing EGF or bFGF for 5–7 d before seeding BM cells. To initiate BM culture, all the medium in the culture dish was removed, and BM cells were plated in the presence of EGF or bFGF. 7–10 d later, cultured BM cells were harvested by pipetting, and 2 × 104 of them were replated onto fresh OP9 stromal layers that had been prepared under the same conditions. Subsequently, the passage of the growing BM cells was repeated in the same manner when the cells became confluent. BM culture on the erbB2/OP9 stromal cell layer was initiated and maintained in the same manner, except that EGF and bFGF were omitted from the medium.
Antibody and Flow Cytometry.
The mAbs used in immunofluorescence staining for lineage markers were Mac-1 (M1/70), B220 (RA3-6B2), anti-CD4 (GK1.5), anti-CD8 (53-6.72), and TER119 (erythroid lineage marker; 25). THY1 (Thy1.2), ACK4 (anti–c-Kit; 26, 27), A7R (anti–IL-7 receptor; 28), AFS98 (anti– c-Fms; 29), GP130 (anti-gp130, a gift from Dr. T. Taga, Osaka University, Osaka, Japan; references 30–33), AIC2B (anti–common β chain for IL-3, -5, GM-CSF, a gift from Dr. S. Yonehara, Kyoto University, Kyoto, Japan; references 34, 35), and TUGm3 (anti–common γ chain for IL-2, -4, and -7 a gift from Dr. K. Sugamura, Tohoku University, Sendai, Japan; references 36, 37) were also applied. All mAbs were purified and conjugated with FITC or biotin. Biotinylated antibodies were visualized with PEconjugated streptavidin (GIBCO BRL) or FITC-conjugated streptavidin (Sigma Chemical Co., St. Louis, MO). The stained cells were analyzed by EPICS XL. (Coulter Electronics, Hialeah, FL).
In Vitro and In Vivo Colony Assay.
In vitro colony formation in methylcellulose-containing semisolid medium was assayed as described (27). Briefly, cultured or fresh BM cells were incubated in 1 ml of culture medium containing α-MEM (GIBCO BRL), 1.2% methylcellulose (Methocel A-4M; Muromachi Kagaku Kogyo, Tokyo, Japan), 30% FCS (lot No. 1M1137; BioWhittaker), 1% deionized BSA (Sigma Chemical Co.), 50 μM 2-ME, 200 U/ml IL-3, and 2 U/ml erythropoietin (Epo; Chugai Pharmaceutical Co. Ltd., Tokyo, Japan). On the 7th d of culture, aggregates consisting of >40 cells were counted as a colony.
For CFU in spleen (CFU-s) assay, C57BL/6 mice were purchased from Japan SLC Inc. (Shizuoka, Japan) and were used as BM donors and recipients at 8–10 wk of age. Recipient mice were maintained on acidified water (pH 2.5) 1 wk before the irradiation (9.2 Gy) with a 60Co source, and donor cells were injected the next day. 8 or 12 d thereafter, the number of colonies that appeared in the recipient spleen was counted after fixation in Bouin's solution (38, 39).
Results
Distinct Types of Hematopoietic Cells Were Generated on OP9 Stromal Cells Stimulated by EGF or bFGF.
Our previous study showed that the RPMI 1640/DMEM/F12– based defined medium mSFO2 (Sanko Junyaku, Chiba, Japan), which contains transferrin and insulin as the only protein components, supported hematopoiesis on the stromal cell lines ST2 and PA6 (40). To reduce the number of factors present in this BM culture, we initialy examined whether the stromal cell line OP9 (41), which was derived from the M-CSF–deficient op/op mouse (42), can be used for the BM culture with mSFO2 instead of ST2 or PA6, which both express M-CSF. Moreover, because 2-ME is essential for differentiation into B cell lineage, it was removed from the original culture. Although OP9 cells could not survive under serum deprivation, addition of either rEGF or rbFGF solved this problem. While these in vitro microenvironments supported the growth of fresh BM cells, proliferation remained at a low level compared with that of stromal cell–dependent hematopoiesis in the medium-containing serum (see Fig. 2 A and data not shown). To enhance the proliferation of hematopoietic cells, we added the ligand for c-kit (SCF) in the culture. The improvement was striking, as the cells extensively proliferated. More marked was the finding that the cells proliferating in the culture with EGF plus SCF retained an immature morphology, having a large nucleus and azurophilic granules (Fig. 1 C), while those in the culture with bFGF plus SCF were heterogeneous, containing mature and immature myeloid cells (Fig. 1 B). In complete agreement with this morphology, flow cytometry of the same cells showed that 98% of the cultured cells were c-Kit+ in the EGF-containing culture. More than 60% of the cells in the bFGF-containing culture were c-Kit− (Fig. 1 B), while it contained more Mac-1high macrophages than the cultures with FCS (Fig. 1 A).
Figure 2.

Growth kinetics and c-Kit expression of BM cells cultured on OP9 stroma cells with mSFO2 containing EGF and SCF. (A) Unfractionated BM cells (105) from C57BL/6 mice were plated on OP9 stroma cells in mSFO2 containing 30 ng/ml EGF (open circle) or 30 ng/ml EGF plus 100 ng/ml SCF (filled circle) in 60-mm culture dishes. On the indicated days, the cultured cells were harvested, counted, analyzed for c-Kit expression, and replated on the fresh OP9 layer. Each point represents the mean of triplicate cultures ± SD. (B) Expression of c-Kit of the cultured cells harvested on the indicated days. Fresh BM (C57BL/6) and cultured BM cells were stained with biotinylated anti–c-Kit (ACK4) antibody revealed by streptavidin-PE. Each staining profile (solid line) was superimposed onto that of the negative control (dashed line) stained with a biotinylated antibody against an irrelevant antigen. Percentage indicates the population of c-Kit–positive cells.
Figure 1.
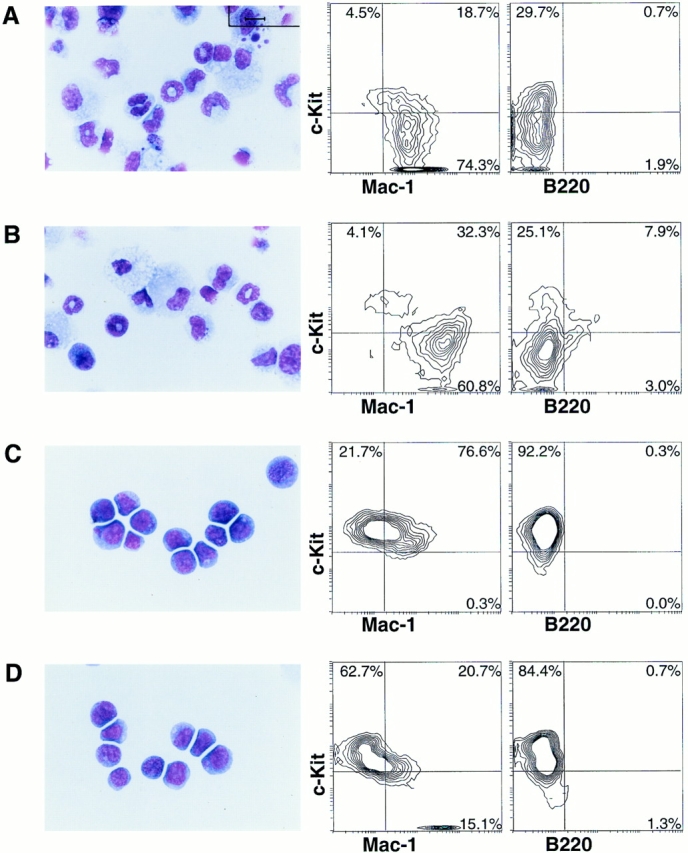
Phenotypes of BM cells cultured under various conditions. Unfractionated BM cells (105) from C57BL/6 mouse were plated onto the OP9 stroma cells and cultured in mSFO2 containing 10% FCS (A), 10 ng/ml bFGF, and 100 ng/ml SCF (B), or 30 ng/ml EGF and 100 ng/ml SCF (C) in 60-mm culture dishes for 14 d. (D) Unfractionated BM cells (105) were plated onto OP9 stromal cells transformed with the v-erbB2 gene (OP9/erbB2) in the mSFO2 plus SCF (100 ng/ml) for 21 d. Cultured BM cells were analyzed by May-Gruenwald-Giemsa staining (left) and by flow cytometry (right) after staining with biotinylated anti–c-Kit (ACK4), FITC-conjugated Mac-1, and anti-B220 antibody. Biotinylated ACK4 was detected with streptavidin-PE. The numbers in each quadrant represent the percentages of gated bone marrow cells. Bar, 10 μm.
Cell Kinetics in the EGF-containing Culture.
The above results indicated that the OP9 stromal cell line with EGF and SCF in the mSFO2-defined medium would provide a culture condition that allows the preferential growth of a subset of hematopoietic stem cells. Next, we investigated how long this selective proliferation of the hematopoietic stem cells lasts under this culture condition. The expression of c-Kit was used as a marker of the immature hematopoietic cells. In this particular experiment, exponential cell proliferation generating > a 1,000-fold increase in the cell number continued up to 40 d (Fig. 2 A). At the initial phase of culture, the number of cultured cells increased ∼100-fold (Fig. 2 A), and the proportion of c-Kit+ cells increased to 80% in the 21-d culture (Fig. 2 B). Since normal BM consists of 14% c-Kit+cells, their net increase over 21 d was ∼600-fold. Thereafter, however, the proportion of c-Kit− cells started to increase concomitantly with a decrease in cell proliferation. These results suggested that this culture condition is indeed selective for the c-Kit+ population during the initial 3 wk, but it cannot block differentiation over a longer period.
EGF Acts on OP9 Stromal Cells to Produce the Microenvironment Selective for Hematopoietic Cells.
Beug et al. have demonstrated that TGF-α acts on chicken erythroid progenitors to maintain self-renewal without differentiation (43). This suggests that EGF, which binds to the same receptor as TGF-α (44), acts directly on murine hematopoietic progenitors to augment their self-renewal. However, we could not detect the direct binding of 125I-labeled EGF to the cells proliferating under this condition (data not shown). An alternative is that EGF acts on OP9 stromal cells to create a microenvironment that is selective for the self renewal of hematopoietic progenitors. To test this notion, we established an OP9 subline (OP9/erbB2) that was transduced with the v-erb-B2 gene containing a constitutive active phosphorylation site in the cytoplasmic domain. This cell line was maintained under serum-deprived conditions without exogenous growth factors.
Unfractionated BM cells (105) were cultured on OP9/ erbB2 in the presence of SCF, but without either EGF or bFGF, and the cells recovered from the culture were counted. As shown in Fig. 3 A, cell growth was exponential under this culture condition, although the growth rate in this culture was slower than that in the culture with EGF. Importantly, the morphological features of the proliferating cells under this condition were similar to those cultured with EGF (Fig. 1 D). This suggested that OP9 stromal cells, when properly stimulated, can create a microenvironment that is selective for c-Kit+ Mac-1dull cells in the absence of exogenous growth factors.
Figure 3.

Growth kinetics of BM cells cultured on the OP9/erbB2 stromal layer and surface phenotype of recovered cells. (A) Unfractionated BM cells (105) derived from C57BL/6 were plated on OP9/erbB2 stroma cells in mSFO2 supplemented with 100 ng/ml SCF. On the indicated days, the cultured cells were harvested, counted, and replated onto the fresh OP9/erbB2 stromal layer. (B) Cells harvested on the 21st d of culture were analyzed by flow cytometry after staining with FITCconjugated Mac-1, anti-B220, TER119, CD4, CD8, biotinylated anti–c-Kit (ACK4),Thy-1, anti–IL-7 receptor (A7R), anti–c-Fms (AFS98), antigp130 (GP130), anti–common β (AIC2B), and anti–common γ (TUGm3). Biotinylated antibodies were detected with streptavidin-PE. The vertical lines indicate the mean (left) and the maximal (right) fluorescence intensity of cells stained with an isotype-matched control mAb.
Phenotype of the Cells Cultured with OP9/erbB2.
We analyzed the surface phenotype of the cells proliferating under this culture condition (OP9/erbB2 plus SCF). As shown in Fig. 3 B, >80% of the culture consisted of c-Kit+ immature cells. This c-Kit+ population was positive for common β and common γ chain expression, dull to positive for Mac-1, Thy-1, and gp130 expression, and negative for c-FMS.
The clonogenic activity of this cell population was analyzed by means of in vitro colony-forming cell assays and by transfer into irradiated mice (Table 1). Consistent with the expression of the IL-3 receptor β-chain in this cell population, half of the cultured cells formed colonies in response to IL-3 plus Epo. Since the frequency of CFU in culture (CFU-c) in normal BM is ∼1/140 (Table 1) and the cell number increased 100-fold during 21 d of culture (Fig. 3), the net increase of CFU-c was at least 7,000-fold. In contrast to this high in vitro colony-forming ability, the frequency of CFU-s measured at days 8 and 12 after transfer were 1/140 and 1/250, respectively. Thus, only a part of this cell population maintained CFU-s activity. Since frequencies of day 8 and day 12 CFU-s in normal BM cells were ∼1/5000, their net increases were ∼3,600- and 2,000-fold, respectively. This indicates that while CFU-s could expand under this culture condition, the cells with long-term reconstituting activity were lost more rapidly than the more mature progenitors. These results suggested that the cells that were selected most preferentially under our culture conditions represent short-term reconstituting progenitors, probably CFU-c.
Table 1.
Frequency of Colony-forming Cells
| Fresh BM per 5 × 104 cells | Cultured cells per 103 cells | Control | ||||
|---|---|---|---|---|---|---|
| No. of CFCs | ||||||
| Factor: IL-3 and Epo | 372.5 ± 13.2 | 523.0 ± 34.0 | 0* | |||
| No. of CFU-S | ||||||
| Day 8 | 11.2 ± 2.5 | 7.0 ± 2.0 | 0 | |||
| Day 12 | 10.6 ± 2.6 | 4.0 ± 1.0 | 0.4 ± 0.6 |
This colony number was obtained by counting fresh BM cells (5 × 104) cultured in methylcellulose without IL-3 and Epo. The numbers in the control line indicate colony numbers of those of nuclear cell counts of BM in the control mice that were injected with PBS. The results represent the mean ± SE of triplicate samples.
We next investigated whether this cell population, phenotypically representing CFU-c, preserves reactivity to differentiation signals. In fact, almost all colonies formed in response to IL-3+Epo contained mature myeloid and monocytic cells (data not shown), suggesting that most cells can readily differentiate. Since the culture with OP9 and medium-containing serum provides conditions that favor differentiation into mature hematopoietic cells (Fig. 1), we investigated whether or not this cell population would react to a shift from serum-deprived medium to medium containing serum and 2-ME. As shown in Fig. 4, there was a quick phenotypic shift, as the number of c-Kit+ cells decreased, whereas the number of c-Kit−Mac-1+ cells increased to 45.8% 7 d after the shift. Thus, most cells maintained under our culture conditions were highly reactive to this shift and readily gave rise to mature myelomonocytic cells. At 14 d in culture, B220+ cells were generated, suggesting that this cell population contained the progenitor cells for B cell lineage. A small proportion of B220−IL-7R+ cells appeared before the B220+ cells, which constituted 50% of the cells at the 14th d of culture. This suggested that IL-7 receptor expression precedes the expression of B220 during B lineage differentiation.
Figure 4.

Immediate differentiation of cultured c-Kit+Mac-1dull cell in medium-containing serum. Unfractionated BM cells were cultured on OP9/erbB2, as described in the legend to Fig. 3. 21 d later, the cells were transferred onto a fresh OP9 stroma cell layer with medium-containing serum (RPMI 1640 with 10% FCS and 5 × 10−5 M 2-ME). The recovered cells were analyzed after staining with FITC-conjugated Mac-1 and biotinylated anti–c-Kit (ACK4), or FITC-conjugated anti-B220 and biotinylated anti–IL-7 receptor (A7R) (A) before, (B) 7 d, or (C) 14 d after the transfer. Biotinylated antibody was detected with streptavidin-PE. The percentage of cells in each quadrant is shown in the top right-hand corner of each panel.
Discussion
Our results demonstrated that the proliferation of a subset of hematopoietic progenitors, most probably CFU-c, can be selectively supported in vitro, whereas that of more mature cells cannot. The optimal conditions for such selection were attained by using either the OP9 stromal cell line, SCF, and EGF, or by using v-erbB2–transduced OP9 cells and SCF. Cells that were c-Kit+ Mac-1dull did not selectively proliferate on the M-CSF–expressing stromal cell lines PA6 or ST2 (40). Moreover, medium-containing serum provided a microenvironment that favored hematopoietic differentiation, even if EGF was added to the culture (data not shown). The addition of 2-ME in mSFO2 promoted the proliferation of pre-B cells (c-Kit+ B220+ Mac-1−) at 21 d of culture, even in the presence EGF and SCF (data not shown). Therefore, the 2-ME–free, chemically defined medium and the M-CSF–deficient OP9 stromal cell line are essential components of these culture conditions. The addition of bFGF instead of EGF, however, produced mature hematopoietic cells. This suggested that an in vitro microenvironment formed by a uniform stromal cell line in the absence of serum could be varied according to the stimulation given to the stromal cells. Thus, the selective proliferation of c-Kit+ Mac-1dull cells described here appeared to be an outcome of a particular combination of stromal cell molecules expressed by EGF-stimulated or v-erbB2–transduced OP9. Nevertheless, these results indicated that hematopoiesis is regulated, not only by molecules directly acting on hematopoietic progenitors, but also by those that stimulate stromal cell components.
It is totally obscure why the stimulation of OP9 cells by EGF created a microenvironment that favored the selfrenewal of immature hematopoietic progenitors, whereas bFGF could not. To examine whether more granulocyte colony stimulating factor (G-CSF) or GM-CSF is produced by OP9 stimulation with bFGF than with EGF, thereby generating mature hematopoietic cells, we tried to inhibit these signals in the culture with OP9 stimulated by bFGF. We inhibited G-CSF and GM-CSF signals, using G-CSF receptor protein fused with human IgG1 protein that saturates G-CSF (our unpublished data) and a neutralizing mAb against GM-CSF (19). Even though we simultaneously inhibited both G-CSF and GM-CSF-signals, we could not block the generation of differentiated cells completely in culture containing bFGF (data not shown). Thus, it is very unlikely that the G-CSF or GM-CSF produced by OP9 is responsible for the difference in the microenvironment created by EGF or bFGF stimulation.
Nevertheless, in complete agreement with the description by Beug et al. (43) of chicken erythroid progenitors, our results suggest that the balance between the self-renewal and differentiation of nontransformed hematopoietic progenitors can be regulated by a combination of the molecules expressed in the microenvironment. This notion challenges the widely held view that differentiated hematopoietic progenitors, including committed progenitors, undergo only a limited number of cell divisions. We are currently investigating which molecules are induced specifically by EGF to support the self-renewal of c-Kit+ Mac-1dull progenitors.
The most important feature of our procedure for enriching CFU-c, as compared with others, lies in the fact that the hematopoietic progenitor cells spontaneously dominate the culture. This is all that is required to simply culture unfractionated BM cells under the conditions described here. However, stem cell clones cannot be established, because the dominance of hematopoietic progenitors does not last more than 30 d because of their eventual differentiation into mature cells that do not proliferate. It is unclear whether this limitation of our method in maintaining the dominance of the immature progenitors is caused by the limited life span of c-Kit+ Mac-1dull cells before terminal differentiation or the inability of our culture conditions to block their terminal differentiation. One line of evidence suggests that this cell population is highly sensitive to the differentiation cue. First, they are likely to represent shortterm reconstituting progenitors rather than the most immature stem cells, because 50% of them formed in vitro colonies, whereas only 0.4% could form day 12 spleen colonies. Second, the expression of Mac-1 in this population agrees with the study of Heimfeld and Weissman (45, 46), who found that Mac-1 is expressed by short-term–reconstituted myeloid stem cells. Although our study and their cell-sorting study did not address the differential capacity of individual cells with this phenotype, Mac-1dull cells in our culture were by no means restricted to myeloid cell lineage, because they also gave rise to B lineage cells (Fig. 4). In fact, a recent study by Morrison et al. (47) has demonstrated that the Thy-1low Mac-1dull population generated cells of multiple lineages. Third, they are highly sensitive to the differentiation cue, because they promptly gave rise to mature blood cells both in vitro and in vivo. Thus, any changes of the environment in the culture that could be induced by stromal cell aging or even by an increase of hematopoietic cells themselves could alter the balance in favor of differentiation of the progenitors. To develop long-term stem cell cultures, it is important to determine how to avoid such changes in the culture conditions.
A number of attempts have been made to establish a stromal cell–dependent culture that selects hematopoietic progenitors. The reports of Dexter and his colleagues have shown that hematopoietic progenitors are spontaneously selected from unfractionated BM cells on a v-src–transduced stromal cell (48, 49). While this method is very similar to ours, it has not become popular, probably because the culture conditions are complex and include a high serum concentration, as well as undefined stromal cell components. In contrast, we used defined medium, only two recombinant growth factors, and a cloned stromal cell line. Palacios and Samaridis (50, 51) took the opposite approach and added a cocktail of growth factors and carefully selected FCS to a cultured stromal cell line to produce stem cell clones. Although they claimed to be able to establish progenitor clones, it has obvious problems in that (a) a large number of cloned cells (>5 × 106) has to be transferred into the irradiated recipient to detect their differentiation; (b) they could not form in vitro colonies, which are important characteristics of normal hematopoietic progenitors; and (c) only sorted cells can be used to initiate the culture. Compared with the reported methods, ours is simple and reproducible, because it uses chemically defined conditions and a cloned stromal cell line. However, it is not effective for isolating cell clones. The c-Kit+ Mac-1dull population enriched in this culture readily differentiated in response to various molecular cues. More than half of it responded to IL-3 to form colonies that contained mature myelomonocytic cells. Moreover, when cultured on OP9 stromal cells together with FCS and 2-ME, they gave rise to both myeloid and B220+ B lymphocytes.
In conclusion, though we still need to use the stromal cell line of which the activity on the growth and differentiation of CFU-c has not been fully defined, we are the first to define the culture conditions that can enrich CFU-c. In view of increasing interest in the clinical application of the ex vivo expansion of hematopoietic stem cells, our results are encouraging because we have showed that self-renewal and differentiation of hematopoietic progenitors are somewhat controllable under defined culture conditions. Although we determined that the removal of M-CSF and serum was essential, while high levels of soluble SCF should be included in the culture condition, other conditions created by EGF-stimulated OP9 cells remain to be determined so that this technology can be applied to human BM culture.
Acknowledgments
We would like to thank Dr.T. Yamamoto for kindly providing the pSV2erbB2 VE plasmid, Dr. T. Taga for GP130, Dr. S. Yonehara for AIC2B, and Dr. K. Sugamura for TUGm3, Dr. T. Sudo for G-CSFR-hIgG1 chimeric protein and anti–GM-CSF, and Dr. Y. Katsura for technical advice regarding the CFU-S analysis.
Footnotes
This study was supported by grants from the Japanese Ministry of Education, Science and Culture (Nos. 07CE2005, 07457085, and 06277103), a grant from the Ministry of Science and Technology (No. 130732125-14), and a grant from RIKEN.
1 Abbreviations used in this paper: bFGF, basic fibroblast growth factor; BM, bone marrow; CFU-c, CFU in culture; CFU-s; CFU in spleen; EGF, epidermal growth factor; Epo, erythropoietin; G-CSF, granulocyte CSF; M-CSF, macrophage CSF; SCF, stem cell factor.
References
- 1.Dexter TM, Allen TD, Lajtha LG. Conditions controlling the proliferation of haematopoietic stem cells in vitro. J Cell Physiol. 1977;91:335–344. doi: 10.1002/jcp.1040910303. [DOI] [PubMed] [Google Scholar]
- 2.Smith AG, Heath JK, Donaldson DD, Wong GG, Moreau J, Stahl M, Rogers D. Inhibition of pluripotential stem cell differentiation by purified polypeptides. Nature (Lond) 1988;336:688–690. doi: 10.1038/336688a0. [DOI] [PubMed] [Google Scholar]
- 3.Williams RL, Hilton DJ, Pease S, Wilson TA, Stewart CL, Gearing DP, Wagner EF, Metcalf D, Nicola NA, Gough NM. Myeloid leukemia inhibitory factor maintains the developmental potential of embryonic stem cells. Nature (Lond) 1988;336:684–686. doi: 10.1038/336684a0. [DOI] [PubMed] [Google Scholar]
- 4.Tsuji K, Zsebo KM, Ogawa M. Enhancement of murine blast cell colony formation in culture by recombinant rat stem cell factor, ligand for c-kit. Blood. 1991;78:1223–1229. [PubMed] [Google Scholar]
- 5.Mayani H, Little MT, Dragowska W, Thornbury G, Lansdorp PM. Differential effects of the hematopoietic inhibitors MIP-1alpha, TGF-beta, and TNF-alpha on cytokine-induced proliferation of subpopulations of CD34+cells purified from cord blood and fetal liver. Exp Hematol. 1995;23:422–427. [PubMed] [Google Scholar]
- 6.Cicuttini FM, Welch KL, Boyd AW. The effect of cytokines on CD34+ Rh-123high and lowprogenitor cells from human umbilical cord blood. Exp Hematol. 1994;22:1244–1251. [PubMed] [Google Scholar]
- 7.Shiohara M, Koike K, Nakahata T, Komiyama A. Hematopoietic progenitors and synergism of interferongamma and stem cell factor. Leuk Lymphoma. 1994;14:203–211. doi: 10.3109/10428199409049670. [DOI] [PubMed] [Google Scholar]
- 8.Mayani H, Dragowska W, Lansdorp PM. Characterization of functionally distinct subpopulations of CD34+cord blood cells in serum-free long-term cultures supplemented with hematopoietic cytokines. Blood. 1993;82:2664–2672. [PubMed] [Google Scholar]
- 9.Verfaillie CF. Soluble factor(s) produced by human bone marrow stroma increase cytokine-induced proliferation and maturation of primitive hematopoietic progenitors while preventing their terminal differentiation. Blood. 1993;82:2045–2053. [PubMed] [Google Scholar]
- 10.Peters SO, Kittler EL, Ramshaw HS, Quesenberry PJ. Murine marrow cells expanded in culture with IL-3, IL-6, IL-11, and SCF acquire an engraftment defect in normal hosts. Exp Hematol. 1995;23:461–469. [PubMed] [Google Scholar]
- 11.Verfaillie CM, Catanzarro PM, Li WN. Macrophage inflammatory protein 1α, interleukin 3, and diffusible marrow stromal factors maintain human hematopoietic stem cells for at least eight weeks in vitro. J Exp Med. 1994;179:643–649. doi: 10.1084/jem.179.2.643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Migliaccio AR, Visser JW. Proliferation of purified murine hematopoietic stem cells in serum-free cultures stimulated with purified stem-cell-activating factor. Exp Hematol. 1986;14:1043–1048. [PubMed] [Google Scholar]
- 13.Berardi AC, Wang A, Levine JD, Lopez P, Scadden DT. Functional isolation and characterization of human hematopoietic stem cells. Science (Wash DC) 1995;267:104–108. doi: 10.1126/science.7528940. [DOI] [PubMed] [Google Scholar]
- 14.Greenberger JS, Sakakeeny MA, Humphries RK, Evans CJ, Eckner RJ. Demonstration of permanent factor-dependent multipotential (erythroid/neutrophil/basophil) hematopoietic progenitor cell lines. Proc Natl Acad Sci USA. 1983;80:2931–2935. doi: 10.1073/pnas.80.10.2931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Scolnick EM, Weeks MO, Shih TY, Ruscetti SK, Dexter TM. Markedly elevated levels of an endogenous src protein in a hematopoietic precursor cell line. Mol Cell Biol. 1981;1:66–74. doi: 10.1128/mcb.1.1.66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Palacios R, Karasuyama H, Rolink A. Ly1+PRO-B lymphocyte clones. Phenotype, growth requirements and differentiation in vitro and in vivo. EMBO (Eur Mol Biol Organ) J. 1987;6:3687–3693. doi: 10.1002/j.1460-2075.1987.tb02702.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nakano T, Kodama H, Honjo T. Generation of lymphohematopoietic cells from embryonic stem cells in culture. Science (Wash DC) 1994;265:1098–1101. doi: 10.1126/science.8066449. [DOI] [PubMed] [Google Scholar]
- 18.Chen CA, Okayama H. Calcium phosphatemediated gene transfer: a highly efficient transfection system for stably transforming cells with plasmid DNA. Biotechniques. 1988;6:632–638. [PubMed] [Google Scholar]
- 19.Carraway III, K.L., and L.C. Cantley. A neu acquaintance for ErbB3 and ErbB4: a role for receptor heterodimerization in growth signaling. Cell. 1994;78:5–8. doi: 10.1016/0092-8674(94)90564-9. [DOI] [PubMed] [Google Scholar]
- 20.Kalthoff H, Roeder C, Gieseking J, Humburg I, Schmiegel W. Inverse regulation of human ERBB2 and epidermal growth factor receptors by tumor necrosis factor alpha. Proc Natl Acad Sci USA. 1993;90:8972–8976. doi: 10.1073/pnas.90.19.8972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yamamoto T, Nishida T, Miyajima N, Kawai S, Ooi T, Toyoshima K. The erbB gene of avian erythroblastosis virus is a member of the srcgene family. Cell. 1983;35:71–78. doi: 10.1016/0092-8674(83)90209-x. [DOI] [PubMed] [Google Scholar]
- 22.Kincade PW, Lee G, Scheid MP, Blum MD. Characterization of murine colony-forming B cells II. Limits to in vitro maturation, Lyb-2 expression, resolution of IgD+subsets, and further population analysis. J Immunol. 1980;124:947–953. [PubMed] [Google Scholar]
- 23.Lee G, Namen AE, Gillis S, Ellingsworth LR, Kincade PW. Normal B cell precursors responsive to recombinant murine IL-7 and inhibition of IL-7 activity by transforming growth factor-β. J Immunol. 1989;142:3875–3883. [PubMed] [Google Scholar]
- 24.Yasunaga M, Wang F-H, Kunisada T, Nishikawa S, Nishikawa S-I. Cell cycle control of c-kit + IL-7R + B precursor cells by two distinct signals derived from IL-7 receptor and c-kitin a fully defined medium. J Exp Med. 1995;182:315–323. doi: 10.1084/jem.182.2.315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ikuta K, Kina T, MacNeil I, Uchida N, Peault B, Chein YH, Weissman IL. A developmental switch in thymic lymphocyte maturation potential occurs at the level of hematopoietic stem cells. Cell. 1990;62:863–874. doi: 10.1016/0092-8674(90)90262-d. [DOI] [PubMed] [Google Scholar]
- 26.Nishikawa S, Kusakabe M, Yoshinaga K, Ogawa M, Hayashi SI, Kunisada T, Era T, Sakakura T, Nishikawa S-I. In utero manipulation of coat color formation by a monoclonal anti-c-kit antibody: two distinct waves of c-kit dependency during melanocyte development. EMBO (Eur Mol Biol Organ) J. 1991;10:2111–2118. doi: 10.1002/j.1460-2075.1991.tb07744.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ogawa M, Matsuzaki Y, Nishikawa S, Hayashi SI, Kunisada T, Sudo T, Kina T, Nakauchi H, Nishikawa S-I. Expression and function of c-kitin hemopoietic progenitor cells. J Exp Med. 1991;174:63–71. doi: 10.1084/jem.174.1.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sudo T, Nishikawa S, Ohno N, Akiyama N, Tamakoshi M, Yoshida H, Nishikawa S-I. Expression and function of the interleukin 7 receptor in murine lymphocytes. Proc Natl Acad Sci USA. 1993;90:9125–9129. doi: 10.1073/pnas.90.19.9125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sudo T, Nishikawa S, Ogawa M, Kataoka H, Ohno N, Izawa A, Hayashi S-I, Nishikawa S-I. Functional hierarchy of c-kit and c-fms in intramarrow production of CFU-M. Oncogene. 1995;11:2469–2476. [PubMed] [Google Scholar]
- 30.Taga T, Kishimato T. Signaling mechanisms through cytokine receptors that share signal transducing receptor components. Curr Opin Immunol. 1995;7:17–23. doi: 10.1016/0952-7915(95)80024-7. [DOI] [PubMed] [Google Scholar]
- 31.Kishimoto T, Akira S, Narazaki M, Taga T. Interleukin-6 family of cytokines and gp130. Blood. 1995;86:1243–1254. [PubMed] [Google Scholar]
- 32.Yin T, Taga T, Tsang ML, Yasukawa K, Kishimoto T, Yang YC. Involvement of IL-6 signal transducer gp130 in IL-11–mediated signal transduction. J Immunol. 1993;151:2555–2561. [PubMed] [Google Scholar]
- 33.Saito T, Taga T, Miki D, Futatsugi K, Yawata H, Kishimoto T, Yasukawa K. Preparation of monoclonal antibodies against the IL-6 signal transducer, gp130, that can inhibit IL-6-mediated functions. J Immunol Methods. 1993;163:217–223. doi: 10.1016/0022-1759(93)90125-q. [DOI] [PubMed] [Google Scholar]
- 34.Gorman DM, Itoh N, Kitamura T, Schreurs J, Yonehara S, Yahara I, Arai K, Miyajima A. Cloning and expression of a gene encoding an interleukin 3 receptor-like protein: identification of another member of the cytokine receptor gene family. Proc Natl Acad Sci USA. 1990;87:5459–5463. doi: 10.1073/pnas.87.14.5459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Takaki S, Mita S, Kitamura T, Yonehara S, Yamaguchi N, Tominaga A, Miyajima A, Takatsu K. Identification of the second subunit of the murine interleukin-5 receptor: interleukin-3 receptor-like protein AIC2B is a component of the high affinity interleukin-5 receptor. EMBO (Eur Mol Biol Organ) J. 1991;10:2833–2838. doi: 10.1002/j.1460-2075.1991.tb07832.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kondo M, Takeshita T, Ishii N, Nakamura M, Watanabe S, Arai K, Sugamura K. Sharing of the interleukin-2 (IL-2) receptor gamma chain between receptors for IL-2 and IL-4. Science (Wash DC) 1993;262:1874–1877. doi: 10.1126/science.8266076. [DOI] [PubMed] [Google Scholar]
- 37.Kimura Y, Takeshita T, Kondo M, Ishii N, Nakamura M, Van-Snick J, Sugamura K. Sharing of the IL-2 receptor gamma chain with the functional IL-9 receptor complex. Int Immunol. 1995;7:115–120. doi: 10.1093/intimm/7.1.115. [DOI] [PubMed] [Google Scholar]
- 38.Magli MC, Iscove NN, Odartchenko N. Transient nature of early haematopoietic spleen colonies. Nature (Lond) 1982;295:527–529. doi: 10.1038/295527a0. [DOI] [PubMed] [Google Scholar]
- 39.Spangrude GJ, Heimfeld S, Weissman IL. Purification and characterization of mouse hematopoietic stem cells. Science (Wash DC) 1988;241:58–62. doi: 10.1126/science.2898810. [DOI] [PubMed] [Google Scholar]
- 40.Nishikawa S, Nakasato M, Takakura N, Ogawa M, Kodama H, Nishikawa S-I. Stromal cell-dependent bone marrow culture with a nearly protein-free defined medium. Immunol Lett. 1994;40:163–169. doi: 10.1016/0165-2478(94)90188-0. [DOI] [PubMed] [Google Scholar]
- 41.Kodama H, Nose M, Niida S, Nishikawa S, Nishikawa S-I. Involvement of the c-kit receptor in the adhesion of hematopoietic stem cells to stromal cells. Exp Hematol. 1994;22:979–984. [PubMed] [Google Scholar]
- 42.Yoshida H, Hayashi S-I, Kunisada T, Ogawa M, Nishikawa S, Okamura H, Sudo T, Shultz LD, Nishikawa S-I. The murine mutation osteopetrosis is in the coding region of the macrophage colony stimulating factor gene. Nature (Lond) 1990;345:442–444. doi: 10.1038/345442a0. [DOI] [PubMed] [Google Scholar]
- 43.Hayman MJ, Meyer S, Martin F, Steinlein P, Beug H. Self-renewal and differentiation of normal avian erythroid progenitor cells: regulatory roles of the TGFα/cErbB and SCF/c-kit receptors. Cell. 1993;74:157–169. doi: 10.1016/0092-8674(93)90303-8. [DOI] [PubMed] [Google Scholar]
- 44.Pain B, Woods CM, Saez J, Flickinger T, Raines M, Peyrol S, Moscovici C, Moscovici MG, Kung H-J, Jurdic P, et al. EGF-R as a hematopoietic growth factor receptor: the c-erbB product is present in chicken erythrocytic progenitors and controls their self-renewal. Cell. 1991;65:37–46. doi: 10.1016/0092-8674(91)90405-n. [DOI] [PubMed] [Google Scholar]
- 45.Heimfeld S, Weissman IL. Characterization of several classes of mouse hematopoietic progenitor cells. Curr Top Microbiol Immunol. 1992;177:95–105. doi: 10.1007/978-3-642-76912-2_8. [DOI] [PubMed] [Google Scholar]
- 46.Heimfeld S, Weissman IL. Development of mouse hematopoietic lineages. Curr Top Dev Biol. 1991;25:155–175. doi: 10.1016/s0070-2153(08)60415-9. [DOI] [PubMed] [Google Scholar]
- 47.Morrison SJ, Lagasse E, Weissman IL. Demonstration that THYlosubsets of mouse bone marrow that express high levels of lineage markers are not significant hematopoietic progenitors. Blood. 1994;83:3480–3490. [PubMed] [Google Scholar]
- 48.Boettiger, D., S. Anderson, and T.M. Dexter. Effect of src infection on long-term marrow cultures: increased selfrenewal of hemopoietic progenitor cells without leukemia. Cell. 36:763–773. [DOI] [PubMed]
- 49.Spooncer E, Heyworth CM, Dunn A, Dexter TM. Self-renewal and differentiation of interleukin-3-dependent multipotent stem cells are modulated by stromal cells and serum factors. Differentiation. 1986;31:111–118. doi: 10.1111/j.1432-0436.1986.tb00391.x. [DOI] [PubMed] [Google Scholar]
- 50.Palacios R, Samaridis J. Fetal liver pro-B and pre-B lymphocyte clones: expression of lymphoid-specific genes, surface markers, growth requirements, colonization of the bone marrow, and generation of B lymphocytes in vivo and in vitro. Mol Cell Biol. 1992;12:518–530. doi: 10.1128/mcb.12.2.518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Palacios R, Samaridis J. Bone marrow clones representing an intermediate stage of development between hematopoietic stem cells and pro-T-lymphocyte or pro-B-lymphocyte progenitors. Blood. 1993;81:1222–1238. [PubMed] [Google Scholar]