

Abstract
The function of the minor subset of T lymphocytes bearing the γδ T cell antigen receptor is uncertain. Although some γδ T cells react to microbial products, responsiveness has only rarely been demonstrated toward a bacterial antigen from a naturally occurring human infection. Synovial fluid lymphocytes from patients with Lyme arthritis contain a large proportion of γδ cells that proliferate in response to the causative spirochete, Borrelia burgdorferi. Furthermore, synovial γδ T cell clones express elevated and sustained levels of the ligand for Fas (APO-1, CD95) compared to αβ T cells, and induce apoptosis of Fashigh CD4+ synovial lymphocytes. The findings suggest that γδ T cells contribute to defense in human infections, as well as manifest an immunoregulatory function at inflammatory sites by a Fas-dependent process.
While most T lymphocytes express a TCR composed of α and β chains, a subpopulation of T cells bearing alternate γ and δ chains exists as a minor subset of peripheral blood lymphocytes (PBL)1 (1). While the function of γδ T cells is uncertain, a clue may lie in their increased proportion at epithelial barriers, during certain infections, and at sites of chronic inflammation such as synovial tissue in rheumatoid arthritis (2–7). Some γδ T cells respond to bacterial products and can be identified after infection of mice with particular bacteria (8–15). However, in humans, leprosy is the only infectious disease to date in which γδ cells from affected individuals have been shown to respond to the causative organism (9).
γδ T cells frequently manifest cytolytic activity toward a broad array of target cells (2, 16). Such a spectrum of cytolysis might occur when a target molecule is widely expressed, such as the Fas antigen (APO-1, CD95) (17). Fas is a 45-kD cell surface molecule that mediates apoptosis and is a member of a family of molecules that includes the type I receptor for TNF. Fas is one of the principle components responsible for T cell–mediated cytotoxicity (18–20). Expression of mRNA for the Fas ligand (FasL) was originally described as being transiently expressed by activated αβ T cells, although higher mRNA levels were noted in γδ T cells (21). More recent findings have noted constitutive expression of FasL by nonlymphoid cells, including Sertoli cells of the testis (22) and certain components of the eye (23). FasL expression by these tissues parallels their ability to suppress immune-mediated inflammation. These collective observations suggested that γδ T cells in Lyme arthritis might respond to Borrelia burgdorferi as well as contribute to regulation of the synovial inflammatory infiltrate.
Materials and Methods
Patients.
Lyme arthritis patients came from areas endemic for Lyme disease and were followed at the Lyme Disease Clinic at the University of Medicine and Dentistry of New Jersey, Robert Wood Johnson Medical School. All patients had histories, exams, and serologies consistent with Lyme arthritis, including Borreliaspecific antibody titers that were higher in synovial fluid relative to serum. Synovial fluid lymphocytes were examined from seven patients with Lyme arthritis of 6-mo to 3.2-yr duration.
Flow Cytometry.
Lymphocytes were isolated from peripheral blood or synovial fluid by Ficoll–Hypaque centrifugation. Cells were stained with the indicated fluorochrome-conjugated antibody at 4°C for 30 min. Antibodies were specific for TCR-αβ (JOVI-1; Ancell Corp., Bayport, MN), TCR-γδ (5A6.E9; T Cell Sciences, Inc., Cambridge, MA), TCR-Vδ1 and TCR-Vδ2 (AB and BB3, respectively, courtesy of Dr. Alessandro Moretta, University of Genoa, Genoa, Italy), CD4 (SFCI12T4D11; Coulter Corp., Hialeah, FL), CD8 (SFCI21Thy2D3; Coulter Corp.), Fas (M38) (24) and FasL (polyclonal C-20; Santa Cruz Biotechnology, Santa Cruz, CA; or monoclonal A11 [25]). Surface staining for FasL was performed by one of three methods. The first approach used a fusion protein composed of the extracellular domain of murine Fas linked to the human Ig Fc portion (Fas-Fc) (26). This was followed by goat anti–human Fc–biotin and then avidin–phycoerythrin. Control staining was accomplished by staining for surface IL4 using an IL4 receptor–Fc fusion protein. Alternatively, surface FasL was measured using either a rabbit antiserum to the extracellular carboxyl-terminal portion of human FasL and purified on a FasL sepharose column (C-20), or monoclonal antibody A11 that recognizes both mouse and human Fas (25). To measure FasL induction, cells were examined 3 h after stimulation with PMA (10 ng/ml) and ionomycin (250 ng/ml), in the absence or presence of metalloprotease inhibition using 5 mM EDTA (27). Samples were analyzed on a Coulter Elite flow cytometer (Coulter Corp.) and at least 2 × 104 events were accumulated for analysis.
Proliferation Assays and Derivation of Lyme Synovial γδ T Cell Clones.
Synovial fluid lymphocytes were cultured in AIM-V serum-free medium (GIBCO BRL, Gaithersburg, MD) in either bulk cultures (106/ml) for phenotyping, or in round-bottomed microtiter wells (105/well) for proliferation assays. Cells were stimulated with 3 μg/ml of a sonicate of B. burgdorferi grown in BSK II medium as previously described (28). Triplicate cultures were pulsed with 3H-TdR during the last 18 h of a 6-d culture, harvested, and counted. From parallel cultures, responding cells were cloned at 0.3 cells/well in AIM-V with 5% FCS in the presence of irradiated PBL (3 × 105/well), human recombinant IL2 (10 U/ml), and 3 μg/ml of B. burgdorferi sonicate. Responding wells were phenotyped and the γδ cells expanded by restimulation at 10-d intervals.
PCR Analysis of Synovial Fluid T Lymphocyte Vδ Repertoire.
Semi-quantitative PCR was performed on samples using cDNA prepared from oligo-dT–primed RNA and reverse transcriptase (GIBCO BRL) as previously described (29). The 5′ Vδ- and Cδ-specific primers are modifications of published sequences (30) as follows: Vδ1: 5′-AGCAACTTCCCAGCAAAGAG-3′; Vδ2: 5′-AGGAAGACCCAAGGTAACACAA-3′; Vδ3: 5′-CACTGTATATTCAAATCCAGA-3′; Vδ4: 5′-TGACACCAGTGATCCAAGTTA-3′; Vδ5: 5′-CTGTGACTATACTAACAGCATGT-3′; Vδ6: 5′-TATCATGGATTCCCAGCC-3′; 5′Cδ: 5′-CTTGTCTGGTGCAG-3′; 3′Cδ: 5′-CTTCACCAGACAAGCGACAT-3′. A PCR reaction master mix that was common to all samples contained 100 mM Tris HCl, pH 8.3, 500 mM KCl, 2 mM MgCl2, 200 μM dNTPs, with 25 pmoles of 3′ Cδ primer, 2.2 μCi α-32P-dCTP, and 2.5 U Taq polymerase (GIBCO BRL) per tube. The final volume was 100 μl and contained 10 ng cDNA, and 25 pmoles of individual Vδ primer. Samples were run on a thermocycler (model 9600; Perkin-Elmer Corp., Norwalk, CT) for 24 cycles using the parameters: cycle 1: 94°C × 3 min, 50°C × 45 s, 72°C × 1 min; cycles 2–23: 94°C × 30 s, 50°C × 45 s, 72°C × 1 min; cycle 24: 94°C × 30 s, 50°C × 45 s, 72°C × 7 min. Samples were resolved on a 29 cm 10% polyacrylamide gel containing 7 M urea in TBE buffer and electrophoresed at 80 V for 18 h. The gel was dried and developed on an analyzer (Betascope 603; Betagen, Waltham, MA). The percentage expression of each Vδ was assigned by dividing the actual cpm for a specific Vδ by the total cpm for Vδ1–Vδ6 after correction for the total Cδ message in each sample.
Assay of Cytotolytic Activity.
Faslow variants of the wild-type Jurkat T cell line, H7 (3% normal surface Fas levels) and B4 (1% normal Fas levels), were derived through irradiation mutagenesis using five doses of 200 Rads each, delivered at 5-d intervals. After each irradiation, cells were cultured in wells coated with lytic anti-Fas antibody (M2, 3 μg/ml)(24). The Faslow variants and wild-type Jurkat cells were incubated with 51Chromium (51Cr) for 1 h, washed, and then mixed at various effector/target ratios with cloned Vδ1 cells in a total volume of 200 μl. After a 4-h incubation at 37°C, 100 μl of supernatant were removed and counted for γ emission. Spontaneous release was determined from labeled targets in the absence of effector cells. Maximum release was determined by lysing target cells with 1.0 N HCl. The percentage of specific 51Cr release was calculated as:
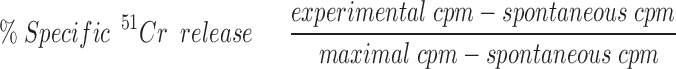 .
.
Blocking studies of cytolysis were performed using either specific antibodies at the concentrations indicated, or Fas-Fc fusion protein (10 μg/ml) preincubated with appropriate cells for 30 min before beginning the cytolysis assay. Antibodies used were specific for TCR-γδ (5A6.E9), HLA class I (W6/32; Accurate Chemical and Science Corp., Westbury, NY), HLA class II (L243; Becton Dickinson & Co., Immunocytometer, Sys., Mountainview, CA), LFA-1 (R7.1; Biosource International, Camarillo, CA), or Fas (M38).
TUNEL Assay for Apoptosis.
Cells were initially stained for expression of surface γδ, CD4, or CD8 and then fixed for 15 min in 1% paraformaldehyde. Cell membranes were then permeabilized for 15 min using 70% ethanol at 4°C. Samples were incubated at 37°C for 1 h in 100 μl containing 10 U terminal deoxyribosyltransferase and 0.5 nM dUTP-biotin (Boehringer Mannheim Biochemicals Corp., Indianapolis, IN) (31, 32). Specimens were washed twice with PBS/1% BSA and incubated with a 1:50 dilution of avidin-tricolor (Caltag Labs., South San Francisco, CA) at 4°C for 30 min. Cells were washed twice and analyzed by flow cytometry.
Results
Reciprocal Changes in Synovial Fluid CD4+ and γδ T Cells with Borrelia Stimulation.
Synovial fluid lymphocytes were examined from seven patients with Lyme arthritis of 6 mo to 3.2-y duration. These contained a predominance of CD4+ over CD8+ αβ T cells in only four of seven cases (Fig. 1 A, Table 1), compared to a consistent CD4 predominance in PBL. Also present in the synovial mononuclear cells was a remarkable percentage of γδ T cells (18.9 ± 6.8%) (Fig. 1 A, Table 1), compared to ∼1–5% in PBL (Reference 1 and see Fig. 3). The synovial γδ population was largely devoid of surface CD4, and only a minor proportion (∼20% on average) expressed low to intermediate levels of CD8 (Fig. 1 B). In addition, whereas γδ T cells from PBL express predominantly the Vδ2 gene product (33), Lyme arthritis synovial fluid γδ cells were primarily of the Vδ1 subset, with lesser proportions of Vδ2 and Vδ3 cells. This was determined by both flow cytometry using Vδ-specific antibodies (Fig. 1 A), and semi-quantitative PCR using specific Vδ primers (Fig. 2).
Figure 1.


Reciprocal shifts in the percentages of γδ versus CD4+ T cells after stimulation by B. burgdorferi of Lyme arthritis synovial fluid T cells. Synovial fluid mononuclear cells, isolated by Ficoll-Hypaque centrifugation, were analyzed either freshly isolated or 6 d after stimulation with a 3 μg/ml sonicate of B. burgdorferi. (A) Flow cytometric analysis of synovial fluid mononuclear cells reveals a prominent population of γδ cells that expresses mostly Vδ1 and expands dramatically following stimulation with B. burgdorferi. The numbers in the histograms indicate the percent of positively stained cells. (B) Synovial fluid γδ cells are predominantly CD4− CD8−. FACS® staining demonstrates that γδ cells are largely devoid of CD4 and only a minor subset expresses low to intermediate levels of surface CD8.
Table 1.
Phenotypic Changes in Lyme Arthritis Synovial Fluid T Cells following Borrelia Stimulation
| Patient No. | Before Borrelia stimulation | After Borrelia stimulation | Proliferation | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| %CD4/%CD8 | %TCR-γδ | %CD4/%CD8 | %TCR-γδ | Medium | + Borrelia | |||||||
| cpm | ||||||||||||
| 1 | 29.7/45.3 | 23.8 | 9.0/39.8 | 56.8 | 5,316 | 178,964 | ||||||
| 2 | 52.6/28.2 | 20.0 | 18.0/29.6 | 45.8 | 526 | 19,122 | ||||||
| 3 | 27.8/45.7 | 15.7 | 29.0/33.2 | 29.9 | 17,369 | 82,440 | ||||||
| 4 | 64.4/16.6 | 11.1 | 55.1/8.8 | 11.8 | 2,791 | 32,418 | ||||||
| 5 | 25.4/39.5 | 13.5 | 7.4/38.3 | 41.5 | 4,516 | 51,278 | ||||||
| 6 | 36.5/18.8 | 30.8 | 51.4/28.9 | 2.9 | 2,937 | 44,474 | ||||||
| 7 | 45.4/35.3 | 11.6 | 14.0/23.6 | 25.7 | 30,230 | 89,533 | ||||||
Synovial fluid lymphocytes were isolated by Ficoll-Hypaque. Specimens were analyzed freshly isolated, or placed in serum-free AIM-V medium (GIBCO BRL) with a sonicate of B. burgdorferi (3 μg/ml) and re-phenotyped after 7 d. Proliferation of 5 × 104 cells/well was measured by 3H-TdR uptake during the last 18 h of culture. cpm, mean of triplicate cultures. Standard deviations were <15%.
Figure 3.

High level expression of surface Fas by synovial CD4+ cells. Mononuclear cells from freshly isolated Lyme arthritis synovial fluid or PBL were immunofluorescently stained for surface expression of Fas and either CD4, CD8, or γδ. Number insets represent percent of positively stained cells. Only the CD4+ subset of synovial fluid cells contains a large proportion of Fashigh cells.
Figure 2.


Semi-quantitative PCR using 5′ primers specific for Vδ1Vδ6 or Cδ, and a 3′ primer specific for Cδ. (A) Actual polyacrylamide gel, imaged on a Betascope, of the Vδ bands for Lyme synovial fluid mononuclear cells either freshly isolated (lane 1) or six days after culture with B. burgdorferi (lane 2). No Vδ product was detected from PBL (lane 3) using the parameters of this assay. At the right side of the gel are the Cδ products from the same three specimens. In addition, a control Cδ product, C , is shown from a Vδ1 clone. (B) Graph of the quantitation of the Betascope results for the synovial fluid lymphocytes freshly isolated (SF 0′) or after culture with B. burgdorferi (SF + Bb). Each Vδ is displayed as a percentage of the total Vδ cpm.
Stimulation of Lyme arthritis synovial fluid mononuclear cells with a sonicate of B. burgdorferi (strain N40) induced vigorous proliferation (Table 1), yielding a two- to threefold increase in cell number over 6 d. During this period, the composition of T cell subsets shifted considerably. Although the percentage of CD8+ cells changed only slightly, there was frequently a striking loss in the proportion of CD4+ cells by as much as threefold. Thus, despite the increase in total lymphocyte number during the 6-d culture, there was frequently little change or even a decrease in the absolute number of CD4+ cells, as illustrated by patient no. 2 in Table 2. This was paralleled by a reciprocal increase in γδ T cells, in some cases to as much as 50% of the cultured synovial lymphocytes (Fig. 1, Table 1). These continued to be mostly Vδ1 cells as determined by both antibody (Fig. 1 A) and PCR (Fig. 2) analysis.
Table 2.
Changes in Absolute Numbers of Synovial T Cell Subsets with Borrelia Stimulation
| Fresh (total absolute No. = 1.1 × 106) | + Borrelia (total absolute No. = 2.3 × 106) | |||||||
|---|---|---|---|---|---|---|---|---|
| Subset | Percent | Absolute No. (× 105) | Percent | Absolute No. (× 105) | ||||
| CD4+ | 52.6 | 5.8 | 18.0 | 4.1 | ||||
| CD8+ | 28.2 | 3.1 | 29.6 | 6.8 | ||||
| γδ+ | 20.0 | 2.2 | 45.8 | 10.5 | ||||
Synovial fluid lymphocytes from patient No. 2 were analyzed for surface phenotype and absolute counts were determined when freshly isolated and after seven days of stimulation with B. burgdorferi.
The loss of CD4+ synovial cells might have resulted from unresponsiveness of this subset to B. burgdorferi, and hence overgrowth by the CD8+ and γδ+ subsets. However, this seems unlikely since we have previously observed that PBL also proliferate strongly to B. burgdorferi with an expansion of predominantly CD4+ cells (28). Alternatively, because PBL contain only a small proportion of γδ cells (1), the γδ subset might be responsible for the loss of CD4+ cells in Borrelia-activated synovial cultures. Consistent with this notion was the one case (patient no. 6) where the percentage of γδ T cells did not increase following stimulation with B. burgdorferi. In this instance, the proportion of CD4+ cells actually increased from 36.5 to 51.4% (Table 1).
Synovial CD4+ Cells are Fashigh Whereas Synovial γδ T Cell Clones are FasL high.
To more directly address the possibility that synovial γδ cells might be cytolytic toward the CD4+ subset, γδ T cell clones were derived from synovial fluids of two Lyme arthritis patients using a sonicate of B. burgdorferi and irradiated autologous PBL. A panel of 18 Borreliaresponsive γδ clones was established, the majority of which express Vδ1 and lack surface CD4 and CD8. DNA sequencing of the δ chain from seven clones confirmed that they all express Vδ1, but were otherwise each unique and contained varying degrees of N region diversity (Roessner, K., manuscript in preparation).
γδ T cells frequently manifest cytolytic activity toward a broad array of target cells (2, 16). Such a spectrum of cytolysis might occur when a target molecule is widely expressed, as is the case with the apoptosis-inducing molecule, Fas (17). As shown in Fig. 3, Fas expression by fresh CD4+ PBL was low to negligible, but was present on a large proportion of CD4+ synovial lymphocytes. By contrast, the CD8+ and γδ+ subsets of PBL or synovial lymphocytes displayed considerably lower levels of Fas.
Surface expression of FasL protein by B. burgdorferi–reactive γδ and CD4+ αβ T cell clones was examined by flow cytometry using two methods, a Fas-Fc fusion protein as well as a purified anti–human FasL rabbit antiserum. Control staining for Fas-Fc was determined using a human IL4 receptor-Fc (IL4R-Fc) fusion protein (as surface-bound IL4 would not be anticipated for a secreted cytokine). Fig. 4 A (column 3) illustrates results of staining using the FasFc fusion protein, on represetative αβ (114B) and γδ (2.11) synovial T cell clones. By this method, surface FasL protein was expressed on a considerably higher proportion of the γδ cells than on the B. burgdorferi–reactive αβ T cell clones seven days after the last stimulation. Similar findings were seen with an additional two αβ and two γδ synovial T cell clones. In contrast, the levels of surface Fas antigen on the γδ clones were somewhat less than on the αβ clones, (Fig. 4 A, column 4).
Figure 4.


Lyme arthritis synovial γδ clones express high levels of FasL. (A) Surface FasL expression using a Fas-Fc fusion protein (column 3) is shown for a B. burgdorferi–reactive αβ T cell clone (114B) and a synovial γδ clone (2.11). Negative staining controls included either second step fluorescein–anti-Fc antibody alone (column 1), or initial staining with IL4R-Fc followed by second step anti-Fc antibody (column 2). Column 4 indicates the levels of Fas expression by the same clones. (B) Surface FasL expression detected by an anti–human FasL rabbit antiserum (C-20) purified using a FasL-sepharose column. Cell lines included the T cell leukemic line, molt 4, and the αβ and γδ T cell clones used in A. Shown is the control staining using rabbit immunoglobulin ( faint line) superimposed on the staining with the anti-FasL antibody (dark line). T cell clones were examined either seven days after the last stimulation with B. burgdorferi (unstimulated ), or 3 h after activation with PMA and ionomycin (iono). Cells were also analyzed in the absence or presence of the metalloprotease inhibitor EDTA in an effort to block degradation of surface FasL (27). Open numbers in histograms represent the percent of positive cells above background staining. Numbers in parentheses indicate the mean fluorescence intensity of the positively stained cells.
The anti-FasL antibody confirmed the disparity in surface FasL expression between synovial γδ versus αβ T cell clones. Fig. 4 B (column 1) shows that 7 d after antigenic stimulation of the Borrelia-reactive αβ (114B) and γδ (2.11) clones, surface FasL was present on the γδ clone, but was only marginally detectable on the αβ clone. This finding was consistent for three αβ and three γδ clones studied. However, the αβ clones were capable of induction of FasL upon stimulation, as shown after 3 h of activation with PMA and ionomycin. In agreement with a recent report (27), FasL expression on the T cell line, Molt 4, was enhanced by blocking metalloprotease activity with EDTA (Fig. 4 B, column 4). This was less consistently observed for the αβ T cell clones, and was not observed for the γδ clones. It was particularly striking that the levels of FasL on the γδ clones remained detectable for at least 10 d following stimulation with B. burgdorferi (Fig. 4 B, column 1). This is in distinct contrast to αβ T cells which express FasL only transiently after activation (21; Roessner, K., unpublished observations).
Synovial γδ cells induce apoptosis of CD4+ cells in a Fasdependent manner.
To further explore whether the Lyme arthritis synovial fluid γδ T cell clones might be cytolytic toward T lymphocytes expressing high levels of surface Fas, the Jurkat T cell line was initially used as a representative Fashigh target. Fig. 5 A shows that the γδ clones manifested very efficient cytolytic activity toward Jurkat cells, with 50% maximal lysis achieved at an effector/target ratio between 10:1 and 3:1. This finding was remarkably consistent for each of five different Vδ1 clones tested from two patients. In contrast, Borrelia-reactive CD4+ αβ T cell clones manifested little, if any, cytolysis of Jurkat cells (data not shown). Cytolysis by the γδ clones was not inhibited by antibodies to TCR-γδ, HLA class I or II, but was blocked by anti–LFA-1 antibody (Fig. 5 C ), supporting the notion that cytolysis was dependent on cell contact.
Figure 5.





Lyme arthritis synovial fluid Vδ1 T cell clones are highly lytic in a Fas-dependent manner. Effector Vδ1 clones were combined at the ratios indicated with 51Crlabeled Jurkat target T cells in a four h cytolytic assay. (A) Comparison of cytolytic activity toward wild-type Fashigh Jurkat T cells (closed squares) compared with two Faslow Jurkat variants, H7 (open squares) and B4 (open circles), which express, respectively, 3 and 1% of surface Fas levels observed on wild-type Jurkat cells. (B) Level of Fas expression on wild-type Jurkat T cells and two variants, H7 and B4, selected by repeated irradiation and culture in the presence of lytic anti-Fas antibody, M2. Number insets indicate the mean fluorescence intensity of the gated area. (C ) Attempts to block γδ cytolytic activity using antibodies to HLA class I (closed squares), HLA class II (open squares), TCR-γδ (open circles), and anti–LFA-1 (closed triangles). The anti–LFA-1 study was part of a separate experiment in which the baseline cytolysis was 42%. (D) Ability of various concentrations of nonlytic anti-Fas antibody M38 to inhibit cytolysis of wild-type Jurkat cells by the Vδ1 clones. Cytolysis assay was also performed in the absence (closed squares) or presence (open squares) of 2.5 mM EGTA, an inhibitor of calcium-dependent perforin activity (18). Lysis in the presence of control IgG antibody (10 μg/ml) is shown by the closed triangle. (E ) Inhibition of Jurkat cytolysis by the Vδ1 clones 16 and 2.11 in the presence of 10 μg/ml of either anti-Fas antibody M38, Fas-Fc fusion protein, both, or IL4R-Fc fusion protein.
The potential contribution of Fas to cytolysis by γδ cells was examined using three approaches. Initially, two Faslow variants of Jurkat cells, H7 and B4, were independently derived by radiation mutagenesis followed by selection with lytic anti-Fas antibody, M2. H7 expresses 3% of the levels of Fas found on wild-type Jurkat cells, whereas B4 displays 1% (Fig. 5 B). Fig. 5 A demonstrates that the efficiency of cytolysis of both Faslow variants was diminished approximately two- to threefold compared to that observed with wildtype Jurkat cells. However, lysis of the Jurkat Faslow variants was not completely eliminated, suggesting that part of the cytolytic activity of the γδ clones was independent of Fas. This was supported by anti-Fas antibody blocking studies.
Inhibition of Jurkat cell cytolysis by the γδ clones was also achieved using a nonlytic anti-Fas antibody, M38 (24). Fig. 5 D shows that the blocking of cytolysis with M38 was partial, achieving 30–50% inhibition at the highest concentration of antibody (10 μg/ml), whereas control mouse Ig did not block cytolysis. In vitro cytolysis consists of a calcium-independent component mediated by Fas and a calcium-dependent component delivered by perforin (18–20). Blocking perforin action by chelation of calcium with EGTA also resulted in partial inhibition of Jurkat cytolysis, which could then be blocked almost completely by the further addition of anti-Fas antibody (Fig. 5 D). A third method of disrupting Fas-FasL interaction used the Fas-Fc fusion protein. Fig. 5 E shows that Fas-Fc, but not IL4RFc, partially blocked cytolysis of Jurkat cells by the γδ clones, though to a slightly lesser extent than did nonlytic anti-Fas antibody.
The above findings show that γδ clones derived from synovial fluid express prolonged and high levels of FasL and suggest that γδ cells preferentially lyse Fashigh cells. To directly assess whether uncloned synovial γδ cells function in a similar manner, FasL expression was determined on synovial lymphocytes after Borrelia stimulation. As shown in Fig. 6 A, 7 d after activation, FasL expression was confined exclusively to a major proportion of the γδ cells. FasL was still expressed by at least 50% of the synovial γδ cells for as long as 11 d after Borrelia stimulation.
Figure 6.


Synovial γδ cells express FasL and their presence correlates with apoptosis of synovial CD4+ cells. (A) Lyme arthritis synovial fluid lymphocytes were stimulated with B. burgdorferi for 5 d and then stained with anti-γδ and monoclonal anti-FasL, A11. (B) Depletion of γδ cells from fresh Lyme arthritis synovial fluid was performed by flow cytometric sorting. Sorted and unsorted synovial fluid lymphocytes from the same specimen were then cultured with B. burgdorferi for 4 d and stained for expression of TCR-γδ, CD4, and CD8. Note the subpopulation of CD4low cells indicated by the white arrow insets, and the percent in that group noted by the number over the arrow. The CD4low cells comprise a larger proportion of the total CD4+ subset in the cultures containing γδ cells (γδ–nondepleted), even though they contained overall a smaller proportion of CD4+ cells compared to the γδ-depleted cultures. These findings were consistent in three experiments. (C) Determination of DNA fragmentation in a second γδ-nondepleted synovial culture using the TUNEL assay combined with surface labeling and flow cytometry. Note that the CD4low subpopulation is the subset undergoing apoptosis.
To further assess the contribution of the γδ cells to the loss of synovial CD4+ cells, the γδ subset was depleted by flow cytometric sorting and compared to a nondepleted sample of the same specimen after five days of stimulation with B. burgdorferi. During this period, the γδ cells in the nondepleted synovial sample expanded from 4.3% to 11% (Fig. 6 B). This was accompanied by a decreased proportion of CD4+ cells, from 35.6 to 25.3%. In striking contrast, the γδ-depleted population contained only 4% γδ cells after 5 d and manifested a predominance of CD4+ cells (40.8%)(Fig. 6 B). In addition, the CD4+ cells in the 4-day cultures contained a subpopulation of CD4low cells which comprised a greater proportion of the total CD4+ cells in the γδ-replete than the γδ-depleted specimen (Fig. 6 B, arrow inset). These CD4low cells represented apoptotic cells, as determined by the TUNEL assay combined with surface staining and analyzed by flow cytometry (Fig. 6 C ). Smaller proportions of apoptotic cells were also observed in the CD8+ and γδ+ subsets. Observations similar to these have been made with γδ depletion of two additional Lyme synovial fluid specimens, as well as by noting a depletion of CD4+ cells when Vδ1 cloned T cells were added to cultures of PBL that have been stimulated with B. burgdorferi (data not shown).
To assess whether the appearance of the apoptotic CD4low subset in the γδ-replete cultures was in part Fas-mediated, FasL was blocked using the Fas-Fc fusion protein. Synovial fluid lymphocytes were stimulated with B. burgdorferi in the presence of either no additives, Fas-Fc, or control mouse IgG. As shown in Fig. 7, the appearance of apoptotic CD4low cells occurred beginning about five days after Borrelia stimulation. The proportion of this subset increased dramatically thereafter in all cultures except that containing Fas-Fc. The findings support the view that the γδ subset induces apoptosis of synovial CD4+ cells at least partly through Fas/FasL interactions.
Figure 7.

FasL inhibition prevents appearance of CD4low apoptotic subset. Lyme arthritis synovial fluid mononuclear cells were stimulated with B. burgdorferi (B.b.) in the presence of no additives (closed circles), or 10 μg/ml of either Fas-Fc (closed squares), or murine IgG (open circles). After the indicated day of stimulation, cultures were analyzed for expression of CD4, CD8, and γδ. Shown is the expression of apoptotic CD4low cells (as identified by TUNEL in Fig. 6 C ) as a percentage of the total CD4+ cells. The results are representative of two experiments.
Discussion
The collective observations suggest an immunoregulatory circuit whereby synovial Vδ1 T cells bearing high levels of FasL selectively restrict the expansion of infiltrating inflammatory Fashigh CD4+ lymphocytes through cytolysis in a Fas-dependent manner. The findings are in agreement with recent studies showing that FasL mRNA expression by T cells is highest in the γδ subset (21). Not only were levels of surface FasL high on the Vδ1 clones, they remained elevated for considerably longer periods than similarly activated αβ T cells. This may serve to explain the broad spectrum of cytolytic activity that has frequently been observed for many γδ cells (2, 16). The results parallel other recent descriptions of immunosuppression resulting from constitutive expression of FasL by Sertoli cells in the testis (22), and by components of the eye (23).
The current findings may also bear on observations that collagen-induced arthritis in mice (34) and adjuvant arthritis in rats (35) are both more severe following administration of anti-γδ antibody. Collagen-induced arthritis is also more aggressive in mice bearing a genetic deletion of the δ locus (Lefrancois, L., personal communication). Similar results have been observed in a model of orchitis in which γδ depletion accelerated the inflammatory response (36). γδ T cells have also been observed to modulate the functional profile of CD4+ cells. In certain instances this has manifested as selectively inhibiting TH2-dependent cytokine responses, such as IgE production in an allergy model (37) and Coxsackievirus-induced myocarditis (38). The resulting TH1 bias may be due solely to the production of the TH1-type cytokine, IFNγ, by γδ cells (37), but may also reflect a greater sensitivity of TH2 cells to Fas-mediated apoptosis. In this regard, it is noteworthy that B. burgdorferi– reactive CD4+ T cells from Lyme arthritis patients express a TH1 cytokine phenotype (39). Studies are in progress to determine whether a TH1 enrichment results in the residual CD4+ synovial T cells following stimulation with B. burgdorferi.
Lyme arthritis synovial γδ T cells also represent a rare instance where γδ T cell clones obtained from a human infectious disease manifest a proliferative response in the presence of the causative agent. This does not establish that Lyme arthritis synovial γδ cells are responding directly to a Borrelial component. It is entirely possible that B. burgdorferi induces the appearance of surface molecules to which Vδ1 cells respond secondarily. Cutaneous lesions in leprosy also contain γδ T cells that react to the causative agent, Mycobacterium leprae (9). The repertoire of γδ cells that react to mycobacterial products is restricted in both humans and mice (11, 40), and in some instances involves recognition of nonpeptide components such as prenyl pyrophosphates (15, 41). Conceivably, γδ cells in Lyme arthritis may also recognize nonprotein components of B. burgdorferi. On balance, the current findings are consistent with the concept that γδ cells participate in the defense against infectious agents while modulating the immune response through Fas-mediated apoptosis.
Acknowledgments
We thank Colette Charland (The University of Vermont College of Medicine, Burlington, VT) for assistance with flow cytometry and Roberta Christie (The University of Vermont College of Medicine, Burlington, VT) for preparation of the manuscript.
Footnotes
This work was supported by National Institutes of Health grant AR43520 and the Arthritis Foundation.
1 Abbreviation used in this paper: PBL, peripheral blood lymphocytes.
References
- 1.Groh V, Porcelli S, Fabbi M, Lanier LL, Picker LJ, Anderson T, Warnke RA, Bhan AT, Strominger JL, Brenner MB. Human lymphocytes bearing T cell receptor γδ are phenotypically diverse and evenly distributed throughout the lymphoid system. J Exp Med. 1989;169:1277–1294. doi: 10.1084/jem.169.4.1277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Goodman T, Lefrancois L. Expression of the γδ T-cell receptor on intestinal CD8+ intraepithelial lymphocytes. Nature (Lond) 1988;333:855–857. doi: 10.1038/333855a0. [DOI] [PubMed] [Google Scholar]
- 3.Spencer J, Isaacson PG, Diss TC, MacDonald TT. Expression of disulfide-linked and non-disulfide-linked forms of the T cell receptor γδ heterodimer in human intestinal intraepithelial lymphocytes. Eur J Immunol. 1989;19:1335–1344. doi: 10.1002/eji.1830190728. [DOI] [PubMed] [Google Scholar]
- 4.Augustin A, Kubo RT, Sim G-K. Resident pulmonary lymphocytes expressing the γδ T-cell receptor. Nature (Lond) 1989;340:239–241. doi: 10.1038/340239a0. [DOI] [PubMed] [Google Scholar]
- 5.Carding SR, Allan W, Kyes S, Hayday A, Bottomly K, Doherty PC. Late dominance of the inflammatory process in murine influenza by γδ+T cells. J Exp Med. 1990;172:1225–1231. doi: 10.1084/jem.172.4.1225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Brennan F, Londei M, Jackson AM, Hercend T, Brenner MB, Maini RN, Feldman M. T-cell expressing γ/δ chain receptors in rheumatoid arthritis. J Autoimmun. 1988;1:319–326. doi: 10.1016/0896-8411(88)90002-9. [DOI] [PubMed] [Google Scholar]
- 7.Holoshitz J, Koning F, Coligan JE, De Bruyn J, Strober S. Isolation of CD4-CD8- mycobacteria-reactive T lymphocyte clones from rheumatoid arthritis synovial fluid. Nature (Lond) 1989;339:226–229. doi: 10.1038/339226a0. [DOI] [PubMed] [Google Scholar]
- 8.Emoto M, Danbara H, Yoshikai Y. Induction of γδ T cells in murine salmonellosis by an avirulent but not by a virulent strain of Salmonella choleraesuis. . J Exp Med. 1992;176:363–372. doi: 10.1084/jem.176.2.363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Modlin RL, Pirmez C, Hofman FM, Turigion V, Uyemura K, Rea TH, Bloom BR, Brenner MB. Lymphocytes bearing antigen-specific γδ T cell receptors accumulate in human infectious disease lesions. Nature (Lond) 1989;339:544–548. doi: 10.1038/339544a0. [DOI] [PubMed] [Google Scholar]
- 10.Haregewoi A, Soman G, Hom RC, Finberg RW. Human γδ+T cells respond to mycobacterial heatshock protein. Nature (Lond) 1989;340:309–312. doi: 10.1038/340309a0. [DOI] [PubMed] [Google Scholar]
- 11.O'Brien RL, Happ MP, Dallas A, Palmer E, Kubo R, Born WK. Stimulation of a major subset of lymphocytes expressing T cell receptor γδ by an antigen derived from Mycobacterium tuberculosis. . Cell. 1989;57:667–674. doi: 10.1016/0092-8674(89)90135-9. [DOI] [PubMed] [Google Scholar]
- 12.Janis EM, Kaufmann SHE, Schwartz RH, Pardoll DM. Activation of γδ T cells in the primary immune response to Mycobacterium tuberculosis. . Science (Wash DC) 1989;244:713–716. doi: 10.1126/science.2524098. [DOI] [PubMed] [Google Scholar]
- 13.Born WK, Hall L, Dallas A, Boymel J, Shinnick T, Young D, Brennan P, O'Brien R. Recognition of a peptide antigen by heat shock-reactive γδ T lymphocytes. Science (Wash DC) 1990;249:67–69. doi: 10.1126/science.1695022. [DOI] [PubMed] [Google Scholar]
- 14.Rust CJJ, Verreck F, Vietor H, Konig F. Specific recognition of staphylococcal enterotoxin A by human T cell bearing γδ receptors. Nature (Lond) 1990;346:572–574. doi: 10.1038/346572a0. [DOI] [PubMed] [Google Scholar]
- 15.Tanaka Y, Morita CT, Tanaka Y, Nieves E, Brenner MB, Bloom BR. Natural and synthetic non-peptide antigens recognized by human γδ T cells. Nature (Lond) 1995;375:155–158. doi: 10.1038/375155a0. [DOI] [PubMed] [Google Scholar]
- 16.Wright A, Lee JE, Link MP, Smith SD, Carroll W, Levy R, Clayberger C, Krensky A M. Cytotoxic T lymphocytes specific for self tumor immunoglobulin express T cell receptor δ chains. J Exp Med. 1990;169:1557–1564. doi: 10.1084/jem.169.5.1557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nagata S. Fas and Fas ligand: a death factor and its receptor. Adv Immunol. 1994;57:129–144. doi: 10.1016/s0065-2776(08)60672-0. [DOI] [PubMed] [Google Scholar]
- 18.Rouvier E, Luciani M-F, Golstein P. Fas involvement in Ca2+-independent T cell-mediated cytotoxicity. J Exp Med. 1993;177:195–200. doi: 10.1084/jem.177.1.195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kagi D, Vignaux F, Lederman B, Burki K, Depraetere V, Nagata S, Hengartner H, Golstein P. Fas and perforin pathways as major mechanisms of T cell-mediated cytotoxicity. Science (Wash DC) 1994;265:528–530. doi: 10.1126/science.7518614. [DOI] [PubMed] [Google Scholar]
- 20.Lowin B, Hahne M, Mattmann C, Tschopp J. Cytolytic T cell cytotoxicity is mediated through perforin and Fas lytic pathways. Nature (Lond) 1994;370:650–652. doi: 10.1038/370650a0. [DOI] [PubMed] [Google Scholar]
- 21.Suda T, Okazaki T, Naito Y, Yokota T, Arai N, Ozaki S, Nakao K, Nagata S. Expression of the Fas ligand in cells of T cell lineage. J Immunol. 1995;154:3806–3813. [PubMed] [Google Scholar]
- 22.Bellgrau D, Gold D, Selawry H, Moore J, Franzusoff A, Duke RC. A role for CD95 ligand in preventing graft rejection. Nature (Lond) 1995;377:630–632. doi: 10.1038/377630a0. [DOI] [PubMed] [Google Scholar]
- 23.Griffith TS, Brunner T, Fletcher SM, Green DG, Ferguson TA. Fas ligand-induced apoptosis as a mechanism of immune privilege. Science (Wash DC) 1995;270:1189–1192. doi: 10.1126/science.270.5239.1189. [DOI] [PubMed] [Google Scholar]
- 24.Alderson MR, Armitage RJ, Marakosky E, Tough TW, Roux E, Schooley K, Ramsdell F, Lynch DH. Fas transduces activation signals in normal human T lymphocytes. J Exp Med. 1993;178:2231–2235. doi: 10.1084/jem.178.6.2231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.French LE, Hahne M, Viard I, Radlgruber G, Zanone R, Becker K, Muller C, Tschopp J. Fas and Fas ligand in embryos and adult mice: ligand expression in several immune-privileged tissues and coexpression in adult tissues characterized by apoptotic cell turnover. J Cell Biol. 1996;133:335–343. doi: 10.1083/jcb.133.2.335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Alderson MR, Tough TW, Braddy S, Davis-Smith T, Roux E, Schooley K, Miller RE, Lynch DH. Regulation of apoptosis and T cell activation by Fas-specific mAb. Int Immunol. 1994;6:1799–1806. doi: 10.1093/intimm/6.11.1799. [DOI] [PubMed] [Google Scholar]
- 27.Mariani SM, Matiba B, Baumler C, Krammer PH. Regulation of cell surface APO-1/Fas (CD95) ligand expression by metalloproteases. Eur J Immunol. 1995;25:2303–2307. doi: 10.1002/eji.1830250828. [DOI] [PubMed] [Google Scholar]
- 28.Roessner K, Fikrig E, Russell JQ, Cooper SM, Flavell RA, Budd RC. Prominent T lymphocyte response to Borellia burgdorferifrom peripheral blood of unexposed donors. Eur J Immunol. 1994;24:320–324. doi: 10.1002/eji.1830240207. [DOI] [PubMed] [Google Scholar]
- 29.Cooper SM, Roessner KD, Naito-Hoopes M, Howard DB, Gaur LK, Budd RC. Increased usage of Vβ2 and Vβ6 in rheumatoid synovial fluid T cells. Arthritis Rheum. 1994;37:1627–1636. doi: 10.1002/art.1780371112. [DOI] [PubMed] [Google Scholar]
- 30.De Libero G, Rocci MP, Casorati G, Giachino C, Oderda G, Tavassoli K, Migone N. T cell receptor heterogeneity in γδ T cell clones from intestinal biopsies of patients with celiac disease. Eur J Immunol. 1993;23:499–504. doi: 10.1002/eji.1830230230. [DOI] [PubMed] [Google Scholar]
- 31.Gavrieli Y, Sherman Y, Ben-Sasson SA. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J Cell Biol. 1992;119:493–498. doi: 10.1083/jcb.119.3.493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sgorc R, Boeck G, Dietrich H, Gruber J, Recheis H, Wick G. Simultaneous determination of cell surface antigens and apoptosis. Trends Genet. 1994;10:41–42. doi: 10.1016/0168-9525(94)90140-6. [DOI] [PubMed] [Google Scholar]
- 33.Parker CM, Groh V, Band H, Porcelli SA, Morita C, Fabbi M, Glass D, Strominger JL, Brenner MB. Evidence for extrathymic changes in the T cell receptor γδ repertoire. J Exp Med. 1990;171:1597–1612. doi: 10.1084/jem.171.5.1597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Peterman GM, Spencer C, Sperling AI, Bluestone JA. Role of γδ T cells in murine collagen-induced arthritis. J Immunol. 1993;151:6546–6558. [PubMed] [Google Scholar]
- 35.Pelegri C, Kuhlein P, Buchner E, Schmidt CB, Franch A, Castell M, Hunig T, Emmrich F, Kinne RW. Depletion of γ/δ T cells does not prevent or ameliorate, but rather aggravates, rat adjuvant arthritis. Arthritis Rheum. 1996;38:204–215. doi: 10.1002/art.1780390206. [DOI] [PubMed] [Google Scholar]
- 36.Mukasa A, Hiromatsu K, Matsuzaki G, O'Brien R, Born W, Nomoto K. Bacterial infection of the testis leading to autoaggressive immunity triggers apparently opposed responses of αβ and γδ T cells. J Immunol. 1995;155:2047–2056. [PubMed] [Google Scholar]
- 37.McMenamin C, Pimm C, McKersey M, Holt PG. Regulation of IgE responses to inhaled antigen in mice by antigen-specific γδ T cells. Science (Wash DC) 1994;265:1869–1871. doi: 10.1126/science.7916481. [DOI] [PubMed] [Google Scholar]
- 38.Huber SA, Mortensen A, Moulton G. Modulation of cytokine expression by CD4+ T cells during Coxsackievirus B3 infection of Balb/c mice initiated by cells expressing the γδ+ T cell receptor. J Virol. 1996;70:3039–3044. doi: 10.1128/jvi.70.5.3039-3044.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yssel H, Shanafelt M-C, Soderberg C, Schneidet PV, Anzola J, Peltz G. Borrelia burgdorferiactivates T helper type 1-like T cell subset in Lyme arthritis. J Exp Med. 1991;174:593–603. doi: 10.1084/jem.174.3.593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Haas V, Pereira P, Tonegawa S. Gamma/delta cells. Annu Rev Immunol. 1993;11:637–685. doi: 10.1146/annurev.iy.11.040193.003225. [DOI] [PubMed] [Google Scholar]
- 41.Morita CT, Beckman EM, Bakowski JF, Tamaka Y, Band H, Bloom BR, Golan DE, Brenner MB. Direct presentation of nonpeptide prenyl pyrophosphate antigens to human γδ T cells. Immunity. 1995;3:495–507. doi: 10.1016/1074-7613(95)90178-7. [DOI] [PubMed] [Google Scholar]