

Abstract
To elucidate the function of the mouse TL antigen in the thymus, we have derived two TL transgenic mouse strains by introducing Tla a -3 of A strain origin with its own promoter onto a C3H background with no expression of TL in the thymus. These transgenic mouse strains, both of which express high levels of Tlaa-3-TL antigen in their thymus, were analyzed for their T cell function with emphasis on cytotoxic T lymphocyte (CTL) generation. A T cell response against TL was induced in Tg.Tlaa-3-1, Tg.Tlaa-3-2, and control C3H mice by skin grafts from H-2K b/T3 b transgenic mice, Tg.Con.3-1, expressing T3b-TL ubiquitously. Spleen cells from mice that had rejected the T3b-TL positive skin grafts were restimulated in vitro with Tg.Con.3-1 irradiated spleen cells. In mixed lymphocyte cultures (MLC), approximately 20% and 15% of Thy-1+ T cells derived from Tg.Tlaa-3-1 and Tg.Tlaa-3-2, respectively, expressed TCRγδ, whereas almost all those from C3H expressed TCRαβ. The MLC from Tg.Tlaa-3-2 and C3H demonstrated high CTL activity against TL, while those from Tg.Tlaa-3-1 had little or none. The generation of γδ CTL recognizing TL in Tg.Tlaa-3-2, but not C3H mice, was confirmed by the establishment of CTL clones. A total of 14 γδ CTL clones were established from Tg.Tlaa-3-2, whereas none were obtained from C3H. Of the 14 γδ CTL clones, 8 were CD8+ and 6 were CD4−CD8− double negative. The CTL activity of all these clones was TL specific and inhibited by anti-TL, but not by anti-H-2 antibodies, demonstrating that they recognize TL directly without antigen presentation by H-2. The CTL activity was blocked by antibodies to TCRγδ and CD3, and also by antibodies to CD8α and CD8β in CD8+ clones, showing that the activity was mediated by TCRγδ and coreceptors. The thymic origin of these γδ CTL clones was indicated by the expression of Thy-1 and Ly-1 (CD5), and also CD8αβ heterodimers in CD8+ clones on their surfaces and by the usage of TCR Vγ4 chains in 12 of the 14 clones. Taken together, these results suggest that Tlaa-3-TL antigen expressed in the thymus engages in positive selection of a sizable population of γδ T cells.
Mice have at least 30–40 genes called “nonclassical” MHC class I or MHC class “Ib” genes mainly mapping to three chromosomal regions within the murine MHC: H-2Q, T, and M (1, 2). Recent studies have demonstrated that some of the proteins encoded by these genes are related to the classical MHC class I molecules in structure and in function, presenting antigens to αβ or γδ T cells (reviewed in reference 2), while others have completely different functions such as that acting as an Fc receptor for IgG (FcRn) (3). The idea that some nonclassical class I molecules might function as restriction elements for γδ T cells has been suggested by the distinctive tissue distribution and limited V gene usage of γδ T cells. Indeed, several studies have shown that γδ T cells recognize nonclassical class I molecules encoded by genes located within the Q or T regions (reviewed in references 4, 5).
TL antigens belong to the family of nonclassical MHC class I antigens (6). At least one gene coding for a TL antigen is present in all mouse strains so far tested, and two or more have been found in some strains; T3 b has been found in C57BL/6 (B6)1, T3 k in C3H/He (C3H), T3 d and T18 d in BALB/c and Tla a -1, Tla a -2 and Tla a -3 in A strain. What makes TL distinct from other MHC class I antigens is their unique expression profile (6–10). TL antigens are expressed in epithelial cells of the small and large intestines in all mouse strains. Certain strains such as A and BALB/c also express TL on normal thymocytes, while others such as B6 and C3H do not. Even in B6 and C3H mice, however, malignant transformation of T cell causes TL to be expressed.
The structures of TL genes and products are very similar to those of classical MHC class I genes and products (6). However, until our recent study, TL had not been shown to behave like MHC class I antigens in terms of histocompatibility antigen. We derived H-2K b/T3 b transgenic mice expressing the TL antigen ubiquitously under the action of an H-2K b promoter (11). By transplanting their skin onto C3H mice, the background strain, we demonstrated that TL can serve as a transplantation antigen and elicit TCRαβ CD8+ CTL response (12). Furthermore, we showed that this CTL response is TL restricted so that CTL directly recognizes TL or a complex of TL plus antigenic molecules without any requirement for antigen presentation by classical MHC class I molecules. The results suggested the possibility that TL can serve as an antigen presenting molecule, similar to classical MHC class I antigens.
We have derived another pair of transgenic mouse strains with Tla a -3 of A strain origin. One of these, Tg.Tlaa-3-1, expresses large amounts of TL antigen on thymocytes and has a small thymus consisting mainly of L3T4(CD4)−Lyt2(CD8)− double negative (DN) cells of γδ lineage, and it develops a high incidence of T cell lymphomas, suggesting that TL plays a critical role in T cell development in the thymus, especially of the γδ T cell lineage (13). However, the other Tla a -3 transgenic strain, Tg.Tlaa-3-2, demonstrates no obvious abnormalities in thymic development and does not develop T cell lymphomas (11). In the present study, to define the role of TL antigens expressed in the thymus, we generated and characterized TL-restricted CTL responses in Tg.Tlaa-3-1 and Tg.Tlaa-3-2 mice and made a comparison with those in C3H strain.
Materials and Methods
TL Genes.
Three TL genes, T3 b of B6 (14), Tla a -3 of A strain (15), and T3 k of C3H (EMBL/GenBank/DDBJ accession No. D86082) were involved in this study. The DNA sequences of their coding regions and deduced amino acid sequences are very similar to each other and >90% of their nucleotides and amino acids are identical. Furthermore, all three TL molecules have identical amino acid sequences in the putative peptide binding region (PBR) which determines the binding specificity of MHC class I molecules to antigenic peptides and to TCR. Outside PBR, however, there are differences as shown in Fig. 1.
Figure 1.

Comparison of the predicted amino acid sequences of TL molecules of Tla a -3 of A mice, T3b of B6, and T3k of C3H. Only amino acid residues different among the three TL molecules are shown. Amino acid sequences are given by the single-letter code, and amino acid numbers in the mature peptide are noted above the sequences.
Mice.
The derivation of transgenic mouse strains has been described previously (11). Briefly, a pair of transgenic strains, Tg.Con.3-1 and Tg.Con.3-2, were produced by introducing a chimeric gene consisting of the H-2K b promoter region and exon 1 (encoding the signal peptide) and the T3 b exons 2–6 (encoding the mature protein). These mice express T3b-TL in almost all tissues including their skins and spleens. The other pair, Tg.Tlaa-3-1 and Tg.Tlaa-3-2, having a Tla a -3 transgene with its own promoter from A strain, express Tlaa-3-TL predominantly on thymocytes. All transgenic mice were generated on a C3H background which does not express TL in the thymus. C3H mice were purchased from Japan SLC Inc. (Hamamatsu, Shizuoka, Japan).
Antibodies.
The following mAbs and one conventional antibody (cAb) were developed in our laboratory, provided by various scientists, or purchased: Rat mAb to Lyt-2 (53-6.7; Dr. N. Shinohara, Mitsubishi Kasei Institute for Life Science, Machida, Japan [16]), rat mAb to L3T4 (GK1.5; Dr. N. Shinohara [17]), hamster mAb to TCRαβ (H57-597; Dr. R.T. Kubo, National Jewish Center for Immunology and Respiratory Medicine, Denver, CO [18]), hamster mAb to TCRδ (3A10; Dr. S. Tonegawa, Massachusetts Institute of Technology, Cambridge, MA [19]), hamster mAb to TCRγδ (GL3; Cedarlane Labs. Ltd., Hornby, Ontario, Canada), hamster mAb to Vγ4 (UC3-10A6; PharMingen, San Diego, CA), hamster mAb to CD3 (145-2C11; Dr. J.A. Bluestone, The University of Chicago, Chicago, IL [20]), mouse mAb to TL.2 (TT213 [12]), mouse mAb to Ly-1.1, mouse mAb to Lyt-3.2 (ID9P35), rat mAb to Thy-1.2 (Becton Dickinson, Immunocytometry Sys., Mountain View, CA), and cAb to H-2k ([B6 × DBA/2] anti-C3H mammary tumor, MM48).
Flow Cytometric Analysis.
Flow cytometric analysis was performed with a FACScan®. For secondary reagents, PE-conjugated streptavidin (Biomeda Corp., Foster City, CA), FITC-labeled anti– hamster Ig (CALTAG Labs., South San Francisco, CA), anti–rat Ig (Tago Inc., Burlingame, CA), or anti–mouse Ig (DAKOPATTS, Glostrup, Denmark) were used.
Skin Grafts.
Female Tg.Tlaa-3-1, Tg.Tlaa-3-2 and C3H mice (6–10-wk-old) received full-thickness sections of skin (1-cm disks) from the abdomens of Tg.Con.3-1 mice onto their backs. Plaster casts were removed on day 10.
Establishment and Maintenance of CTL Bulk Cultures and Clones.
The methods for induction of CTL and assays for their cytotoxic activity have been described previously (12). Briefly, 4–8 wk after the rejection of grafted skin, spleen cells (5 × 107) from the recipient mice were cultured with 5 × 107 irradiated (2,000 R) Tg.Con.3-1 spleen cells for 5 d. The cells were used as effector cells in CTL assays and maintained as CTL bulk cultures by weekly restimulation with irradiated Tg.Con.3-1 spleen cells in the presence of human recombinant IL-2 (Takeda Chemical Industries Ltd., Osaka, Japan) at a concentration of 5 ng/ml. For CTL assays, 51Cr-labeled target cells (2 × 104) were incubated with various numbers of effector cells. For the blocking test of CTL activity by antibodies, serially diluted antibodies were added to mixtures of effector cells and labeled target cells. After incubation for 3 h at 37°C, the supernatants were harvested using a Supernatant Collection System (Skatron Inc., Sterling, VA) for measurement of 51Cr release. The percent specific lysis was calculated using the following equation: 100 × (a − b/c − b), where a is the radioactivity in the supernatant of target cells mixed with effector cells, b is the radioactivity in the supernatant of target cells incubated without effector cells, and c is the radioactivity in the supernatant after complete lysis of target cells with 2% Triton X-100.
For cloning, cells were diluted to 1–100 cells/well and cultured with irradiated Tg.Con.3-1 spleen cells in the presence of IL-2 in 96-well plates. After 10–14 d, the cytotoxity of clonally growing cells was tested. Established clones were maintained by weekly stimulation and their activity was tested as described above.
Target Cells.
Con A–stimulated spleen cells of Tg.Con.3-1 and C3H were prepared as previously described (12). In addition to Con A–stimulated spleen cells, the following cells were used as targets for CTL assays, with the amounts of TL gene product on cell surfaces were estimated by flow cytometry with TT213 anti-TL mAb: Con A blasts of Tg.Con.3-1 (mean fluorescence intensity [MFI] of stained cells - MFI of unstained cells; 298.6, expressing T3b), C3H (0.1, TL−) and B6 (1.5, TL−), thymidine kinase-negative L-cells ([Ltk−], 4.0, TL−), and H-2K b/T3 b Ltk− transfectants (785.0, T3b), as well as ERLD (derived from B6, 253.5, T3b), RADA1 (derived from A strain, 178.9, Tlaa-1, -2, and -3), and ASL-1 (derived from A strain, 584.1, Tlaa-1, -2, and -3) leukemias maintained in vivo.
RNA Extraction and Reverse Transcription–PCR.
Total RNA was extracted from CTL clones using TRIzol Reagent (GIBCO BRL, Gaitherburg, MD). 10 μg of total RNA were reverse transcribed into first strand cDNA with 3 μg of random primers and 1,000 U of SuperScript II reverse transcriptase (RT) (GIBCO BRL). TCRγ chains were amplified using Vγ-specific primers and a single common Cγ primer by the methods of Takagaki (21). The nomenclature of Vγ is according to Reilly et al. (22). The RT-PCR products were separated by agarose gel electrophoresis.
Nucleotide Sequencing of the Vγ-Jγ Junction.
PCR products were cloned into a T-tailed M13mp18 vector and sequenced by a cycle sequencing method using an automated DNA sequencer (model 373A; Applied Biosystems, Inc., Foster City, CA).
Results
Characteristics of Anti–T3b-TL CTL Generated in Transgenic Mice Expressing Tlaa-3-TL Antigen in the Thymus.
To investigate the role of TL antigen expressed in the thymus, a T cell response against TL was induced by transplanting T3bTL positive skin of Tg.Con.3-1 onto Tg.Tlaa-3-1 and Tg.Tlaa-3-2 mice, two transgenic strains expressing Tlaa3-TL in the thymus, and also onto C3H mice without thymic TL expression (Fig. 2). All three strains rejected the grafted skin within 3 wk, in accordance with our previous data (12). The spleen cells from mice which had rejected Tg.Con.3-1 skin were restimulated in vitro with irradiated T3b-TL+ spleen cells from Tg.Con.3-1. Spleen cells of Tg.Tlaa-3-2 and C3H reacted vigorously and about 80% were found to be blastic cells expressing Thy-1 and CD3 on their surfaces. In contrast, Tg.Tlaa-3-1 reacted weakly and only 25% were Thy-1+CD3+ blastic cells. (Fig. 3 A). Of the Thy-1+ T cells, almost all of those derived from C3H expressed TCRαβ, while ∼15% from Tg.Tlaa-3-2 and >20% from Tg.Tlaa-3-1 expressed TCRγδ. In the γδ T cell population derived from Tg.Tlaa-3-2, two thirds were L3T4−Lyt-2+, while approximately one third were L3T4−Lyt-2− double negative (DN), and very few were L3T4+Lyt-2− (Fig. 3 B). Of the γδ T cells from Tg.Tlaa-3-1, ∼60% were DN, 25% were L3T4−Lyt-2+, and 15% were L3T4+Lyt-2− (data not shown). In the αβ T cell population derived from both Tg.Tlaa-3-2 and C3H, >70% were L3T4−Lyt-2+ and the rest were L3T4+Lyt-2−. In contrast, the αβ T cell population from Tg.Tlaa-3-1 consisted of ∼45% L3T4−Lyt-2+, 45% L3T4+Lyt-2− and 10% DN cells (data not shown).
Figure 2.

Experimental design for testing the characteristics of anti-TL CTL generated in Tla a -3 transgenic and C3H mice. The skins obtained from Tg.Con.3-1 mice expressing T3b-TL antigen were grafted onto Tg.Tlaa-3-1 and Tg.Tlaa-3-2 transgenic and C3H mice. Tg.Tlaa-3-1 and Tg.Tlaa-3-2 mice express Tlaa3-TL encoded by the transgene in the thymus and in the intestines as well as endogenous T3kTL in the intestines, while C3H mice express T3k-TL in the intestines, but not in the thymus. CTL were generated by MLC, in which spleen cells of mice that had rejected the skin graft were restimulated with irradiated Tg.Con.3-1 spleen cells.
Figure 3.

Surface antigen profiles of the TL-reactive cells obtained from bulk MLC. Spleen cells from Tg.Tlaa-3-2, Tg.Tlaa-3-1, and C3H that had rejected Tg.Con.3-1 skin grafts were cultured with irradiated spleen cells from Tg.Con.3-1. After 5 d in culture, the cells were double-stained with various combinations of antibodies, and the blastic cell population was gated and analyzed with a FACScan®. (A) Expression of TCR/CD3 complexes on Thy-1+ cells in bulk MLC. About 80, 25, and 80% of the gated cells derived from Tg.Tlaa-3-2, Tg.Tlaa-3-1, and C3H, respectively, expressed Thy-1 on their surfaces. The percentages of cells expressing CD3, TCRαβ, and TCRγδ among Thy-1+ cells are indicated. (B) Expression of L3T4 and Lyt-2 on TL-reactive γδ T cells obtained from Tg.Tlaa-3-2.
The in vitro stimulated T cells were tested for cytotoxic activity against TL+ targets. T cells from Tg.Tlaa-3-2 and C3H showed high levels of CTL activity against TL+ target cells, but not TL− cells (Fig. 4 A). Tg.Tlaa-3-1, however, yielded little or no CTL activity (data not shown), confirming the previous finding that the T cell function of Tg.Tlaa-3-1 is severely impaired (13). Therefore, in the subsequent studies, only CTL from Tg.Tlaa-3-2 and C3H were characterized. The CTL activity of Tg.Tlaa-3-2 against various target cells was not different from that of C3H which has been previously described (12). As shown in Fig. 4 B, the CTL activity from both was blocked by anti-TL antibody, but not by anti-H-2k, thus confirming that CTL recognizes TL directly without involving antigenic presentation by H-2 (12). The CTL activity of C3H was also blocked by anti-TCRαβ, CD3, and Lyt-2 antibodies but not by anti-TCRγδ or anti-L3T4. In the Tg.Tlaa-3-2 case, the CTL activity was blocked partially by anti-TCRγδ antibody, although not as strongly as shown by anti-TCRαβ antibody, indicating a certain population of γδ T cells. Together with the data from flow cytometric analysis, these results suggested that a small but significant proportion of CTL recognizing T3b-TL was skewed toward the γδ lineage by the presence of Tlaa-3-TL antigen in the thymus.
Figure 4.
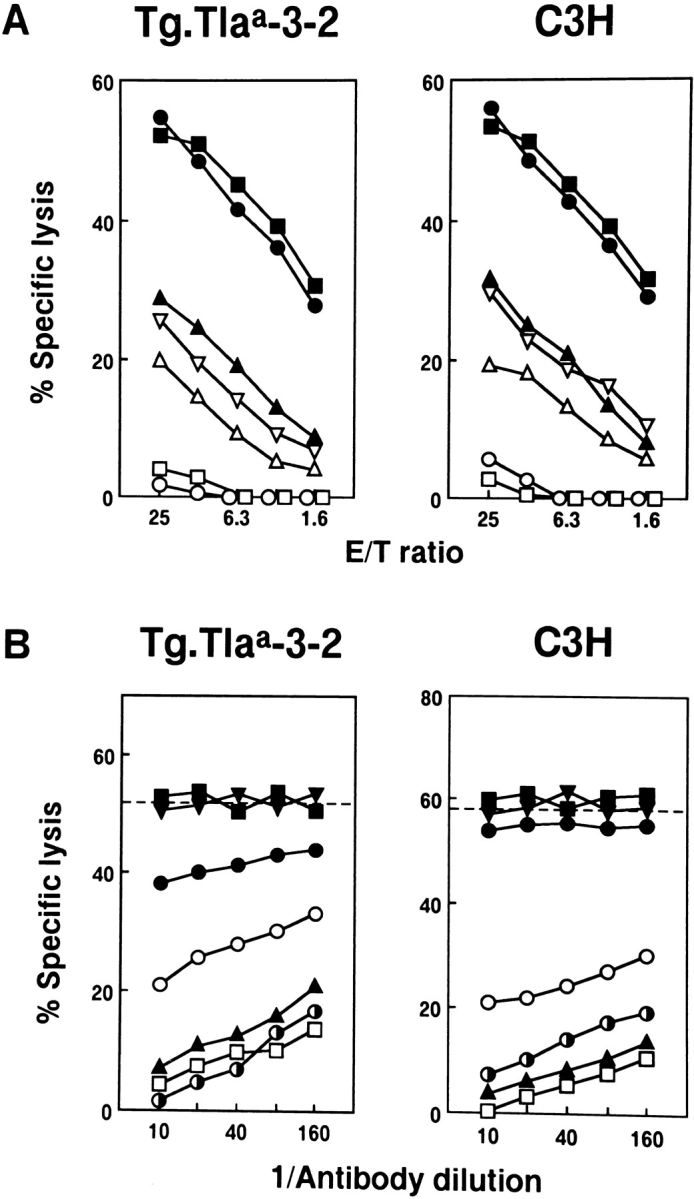
Specificity of the TL-reactive CTL populations in bulk cultures derived from Tg.Tlaa-3-2 and C3H. (A) Generation of CTL against TL antigen in Tg.Tlaa-3-2 and C3H. The effector cells were prepared from Tg.Tlaa-3-2 and C3H mice that had rejected Tg.Con.3-1 skin grafts. Target cells were T3b-TL+ blast cells of Tg.Con.3-1 (•) and TL− blast cells of C3H (○), H-2K b/T3 b Ltk− transfectants (▪), Ltk− (□), ERLD (▴), RADA1 (▵), and ASL1 (▿). For TL expression on these target cells, see Materials and Methods. (B) Antibody blocking for characterization of the CTL populations from Tg.Tlaa-3-2 and C3H. The cytotoxic activity of CTL generated in Tg.Tlaa-3-2 and C3H by grafting Tg.Con.3-1 skin was tested in the presence of mAbs to TL.2 (TT213) (□), TCRαβ (○), TCRγδ (•), CD3 (◑), L3T4 (▾), Lyt-2 (▴), or cAb to H-2k (▪). The broken lines indicate the percentage specific lysis obtained without antibodies. The target cells were Con A blast cells of Tg.Con.3-1 and E/T ratio was 15:1.
Establishment of γδ CTL Clones.
To examine further the influence of TL antigen expressed in the thymus on the CTL population, TL-reactive CTL clones were established from Tg.Tlaa-3-2 and C3H. 23 clones were initially derived from four Tg.Tlaa-3-2 mice and the same number from three C3H mice. By flow cytometric and antibody blocking analyses, 3 out of the 23 CTL clones derived from Tg.Tlaa-3-2 mice were found to express TCRγδ, whereas all C3H CTL clones expressed TCRαβ. Thus, establishment of the CTL clones indicated that γδ CTL are induced characteristically in mice expressing TL in their thymus, confirming the results of bulk cultures.
Characteristics of the γδ CTL Clones.
The three γδ CTL clones described above were all Lyt-2+ (see below), although one third of γδ T cells in bulk culture were DN as shown in Fig. 3 B. To test whether TL-reactive γδ CTL clones with DN phenotype could be established, 11 additional γδ CTL clones were derived from five Tg.Tlaa-3-2 mice. Thus, a total of 14 γδ CTL clones were characterized, as summarized in Table 1. Flow cytometric analysis showed eight to be Lyt-2+ and six to be DN. Lyt-2 is a CD8α chain, and in T cells of thymic origin, it forms a heterodimer with Lyt-3, a CD8β chain, while it forms an αα homodimer in those of intestinal epithelium (23). All Lyt-2+ γδ CTL clones established in this study also expressed Lyt-3, indicating a thymic origin. In addition, a thymic origin of not only Lyt-2+, but also DN γδ CTL clones, was indicated by the fact that all clones expressed Thy-1 and Ly-1 (CD5) on their cell surface (Table 1). Both antigens have been shown to be expressed on γδ T cells of thymic origin, but not on those of extra-thymic origin (24, 25).
Table 1.
Summary of γδ CTL Clones
| Clone* | Vγ usage (RT-PCR) | γδTCR expression§ | Surface antigens (FACS®) | Blocking of the cytotoxicity with antibody¶ | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TCRγδ | TCRδ | Vγ4 | CD4/CD8‖ | Thy-1 | Ly-1 | TL | H-2 | TCRδ≳≳ | Lyt-2 | Lyt-3 | L3T4 | |||||||||||||||
| KC1-3 | 1/2‡ | + | + | − | CD8αβ | + | + | + | − | + | + | + | − | |||||||||||||
| 1-7 | 4‡ | + | + | + | CD8αβ | + | + | + | − | + | + | + | − | |||||||||||||
| 4-4 | 2/4‡ | + | + | + | CD8αβ | + | + | + | − | + | + | + | − | |||||||||||||
| 4-8 | 1/2/4‡ | + | + | + | DN | + | + | + | − | + | − | − | − | |||||||||||||
| 4-9 | 2/4 | + | + | + | CD8αβ | + | + | + | − | + | + | + | − | |||||||||||||
| 5-1 | 2/4 | + | + | + | CD8αβ | + | + | + | − | + | + | + | − | |||||||||||||
| 5-2 | 2/4 | + | + | + | CD8αβ | + | + | + | − | + | + | + | − | |||||||||||||
| 6-1 | 2/4‡ | + | + | + | DN | + | + | + | − | + | − | − | − | |||||||||||||
| 6-2 | 2/4 | + | + | + | CD8αβ | + | + | + | − | + | + | + | − | |||||||||||||
| 7-3 | 1/2/4 | + | + | + | CD8αβ | + | + | + | − | + | + | + | − | |||||||||||||
| 7-4 | 1/2‡ | + | + | − | DN | + | + | + | − | + | − | − | − | |||||||||||||
| 8-1 | 2/4 | + | + | + | DN | + | + | + | − | + | − | − | − | |||||||||||||
| 8-3 | 2/4‡ | + | + | + | DN | + | + | + | − | + | − | − | − | |||||||||||||
| 8-4 | 2/4 | + | + | + | DN | + | + | + | − | + | − | − | − | |||||||||||||
The first digit of the clone name shows the origin of the mouse.
RT-PCR products were sequenced (see Fig. 6).
Expression of TCRγδ, TCRδ, and Vγ4 chains on the cell surface were detected by FACS® using moAb GL3, 3A10, and UC3-10A6, respectively.
Monoclonal antibodies to L3T4, Lyt-2 and Lyt-3 were used.
The target cells were Con A blasts of spleen cells from Tg.Con.3-1 (T3b-TL positive).
Monoclonal antibody 3A10 was used.
The 14 γδ clones were examined for their cytotoxic activity against TL+ and TL− target cells. All lysed TL+ target cells including Con A blasts of Tg.Con.3-1 and RADA1, but not TL− cells including Con A blasts of C3H and B6. The results with CTL clones were essentially identical to those with the bulk culture. The γδ CTL clones were also tested by antibody blocking for their target specificity and the expression of TCR and coreceptor molecules. The results are summarized in Table 1 and representative findings are shown in Fig. 5. Fig. 5, A and C, illustrate results for one of the Lyt-2+ γδ CTL clones, KC4-4, and Fig. 5, B and D, for one of the DN γδ CTL clones, KC8-3. The CTL activity of all these γδ CTL clones was blocked by anti-TL antibodies, indicating the TL-specific CTL activity. Blocking by anti-TL antibodies but not by anti–H-2k antibody further indicated that these γδ CTL clones directly recognize the TL molecule. The CTL activity was also blocked by anti-TCRγδ and CD3 antibodies. All Lyt-2+ γδ CTL clones were blocked by anti–Lyt-2 and anti–Lyt-3 antibodies while all DN clones were not, confirming the results of flow cytometric analysis. Other than the Lyt-2 and Lyt-3 expression, these 14 γδ CTL clones showed no differences in their expression of adhesion, costimulatory, and accessory molecules; all γδ CTL clones were positive for Pgp-1 (CD44), LFA-1 (CD11a), ICAM-1 (CD54), CD2, CD28, CTLA-4, and IL-2Rα/β, and negative for L-selectin (CD62L), VLA-4α (CD49d), and HSA (data not shown).
Figure 5.
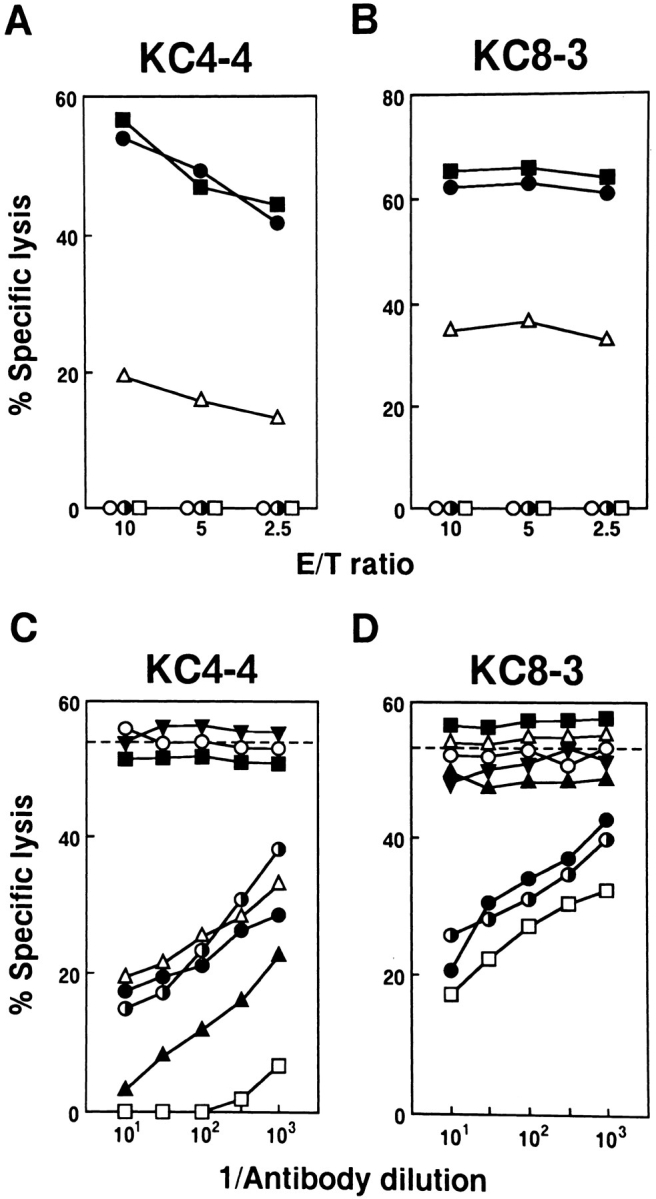
Specificity of two representatives of Lyt-2+ and DN γδ CTL clones. (A and B) Target specificity of Lyt-2+ KC4-4, and DN KC8-3 γδ CTL clones against TL. Target cells were Con A blast cells of Tg.Con.3-1 (•), C3H (○) and B6 (◑), H-2K b/T3 b Ltk− transfectants (▪), Ltk− (□), and RADA1 (▵). For TL expression on these target cells, see Materials and Methods. (C and D) Antibody blocking for characterization of Lyt-2+ KC4-4 and DN KC8-3 γδ CTL clones. The cytotoxic activity of γδ CTL clones was tested in the presence of mAb to TL.2 (TT213)(□), TCRαβ (○), TCRγδ (•) , CD3 (◑), L3T4 (▾), Lyt-2 (▴), Lyt-3 (▵), or cAb to H-2k (▪). The target cells were Con A blast cells of Tg.Con.3-1 and E/T was 5:1.
To further confirm the thymic origin of these γδ CTL clones, their Vγ segment usage was determined. By RTPCR, nine clones yielded Vγ2 and Vγ4 products, two clones Vγ1 and Vγ2, two clones Vγ1, Vγ2 and Vγ4, and one clone Vγ4, while no clones yielded Vγ5, Vγ6, or Vγ7 products (Table 1). There was no distinctive difference in the usage of Vγ by Lyt-2+ clones from that by DN clones. All of 12 clones with RT-PCR Vγ4 products reacted with an antibody specific to Vγ4 in the flow cytometric analysis, indicating that Vγ4 chains were indeed expressed on the cell surface. It has been reported that the Vγ4 chain is preferentially used by thymus-dependent γδ T cells, while Vγ1 and Vγ2 chains are used by both thymus-dependent and -independent γδ T cells (26, 27). Thus, the results indicated that most, if not all, of these clones were derived from adult thymus. To determine the diversity in the CDR3 region of γ chains, RT-PCR products from seven representative clones, three Lyt-2+ and four DN, were sequenced (Fig. 6). One or two in-frame products were identified and they contained N region nucleotides at all the V–J junctions except for Vγ2 and Vγ4 chains in KC1-3 and KC4-8, respectively. The deduced amino acid sequences for the CDR3 were different from one another except for those of the Vγ2 chains in the KC6-1 and KC7-4 clones, suggesting that a diverse set of γδ TCR recognize the TL molecule. In summary, the usage of Vγ4 chains with N sequences in 12 of 14 clones, and the expression of Thy-1, Ly-1, and CD8αβ heterodimer in the case of Lyt-2+ clones, indicated that these γδ CTL clones that recognize TL were matured in adult thymus.
Figure 6.

Nucleotide sequences of the V-J junction of TCRγ-genes expressed in γδ CTL clones. RNA was extracted from γδ CTL clones and RT-PCR was carried out. Sequencing of the RT-PCR products was conducted as described in Materials and Methods. For each sequence, the N nucleotides were deduced from the germline sequence previously reported (26–30). The nomenclature of Vγ segments is according to Reilly et al. (22).
Discussion
The present study on the role of TL antigen expressed in the thymus on T cell selection demonstrated that a sizable population of CTL recognizing T3b-TL induced in Tlaa-3-2 transgenic mice are γδ T cells, in contrast to those in C3H mice which were found to be mostly αβ T cells. Most of the γδ CTL were of adult thymic origin; all of them expressed Thy-1, Ly-1 (CD5) and CD8αβ heterodimers in the cases of Lyt-2+ clones, and 12 of 14 clones used TCR Vγ4 chains with N sequences in the V–J junctions except for Vγ4 of KC4-8. Thus, our results indicate that the presence of Tlaa-3-TL in the thymus promotes the positive selection of γδ T cells recognizing T3b-TL. T3b and Tlaa-3 have identical amino acids in the putative PBR which determine the binding specificity to antigenic peptides and to TCR. Therefore, it is most likely that γδ T cells, selected on the basis of their affinity for Tlaa-3, are able to react with T3b as well, and that such γδ T cells are stimulated to generate CTL against T3b by T3b+ skin grafts. Thus, the presence of TL molecules in the thymus is required for the selection of γδ T cells reactive to TL, and cannot be substituted for by any other MHC class I or CD1 molecules expressed in C3H mice. This notion is supported by the expansion of γδ T cells in the thymus and periphery of the other transgenic mouse strain, Tg.Tlaa-3-1, as we reported previously (13). An association of TL antigen with γδ T cells has also been speculated from the observation that it is expressed in the intestinal epithelium and γδ T cells are present nearby (9, 10), although no direct evidence for any interaction between the intestinal epithelium and γδ T cells has been presented.
It is an intriguing question whether normal mice that express TL in the thymus also produce γδ CTL with specificity for TL. However, they do have at least two, and as many as four, TL genes (6, 31) which are expressed simultaneously in the thymus at levels which differ from strain to strain. Therefore, it will be a very difficult task to determine which TL genes are responsible for T3b-specific responses and for selection of γδ CTL. Although two TL congenic strains, B6-TL+ and A-TL− are available, it is known that they are different from their counterparts, B6 and A, not only in terms of their TL genes but also for as many as 20 other MHC class I genes located in the TL region (32). Some of these class I genes have very similar structures and expression patterns to TL genes (10, 32), and some have been shown to cause skin graft rejection (33). Accordingly, it will be very difficult to rule out their involvement in TL recognition. To unravel the complexity of the TL system, we have generated and analyzed transgenic mice with Tla a -3 gene driven by its own promoter as a first step. A definite answer can only be obtained from the analysis of transgenic mice with other TL genes, such as Tla a-1, Tla a -2 and T18 d, which are currently under construction.
Regarding the specificity, the finding that both bulk and cloned γδ CTL were cytotoxic against Tlaa-3-expressing RADA1 suggested that they may be auto-reactive. RADA1, however, expresses at least two other TL genes, in addition to Tla a -3 (6). Thus, there remains a possibility that the reactivity is directed against antigens encoded by TL genes other than Tla a-3. To clarify this point, target cells expressing only Tla a-3 are needed. Transfection experiments to this end are now underway. Only after they are concluded and modes of recognition of TL molecules by CTL are elucidated (see below), will we be in a position to answer the questions of whether and how the negative selection operates against Tla a-3 in the thymus.
The majority of γδ T cells in mice and humans and most γδ T cell clones or hybridomas reported so far do not express either CD4 or CD8 (4, 5). This suggests that γδ T cells recognize MHC or non-MHC molecules with low or no affinity to CD4 or CD8 coreceptors, or that the maturation and activation of γδ T cells do not require the presence of CD4 or CD8 molecules. Since there have hitherto been only few reports of γδ T cell clones with clear CTL or helper activity, it has been difficult to conduct detailed studies on the requirement of coreceptors for γδ T cells. Our present finding that 8 of 14 TL-restricted γδ CTL clones express CD8αβ, supports the proposal that TL can bind CD8 (34) and further implies that the CD8-p56lck signal pathway can operate in γδ CTL, similarly to the αβ CTL case (35). In addition to CD8+ γδ CTL, DN γδ CTL clones were also established. Analysis of DN γδ CTL clones has so far shown no differences from CD8+ γδ CTL clones in their cytotoxic specificity or activity, or in the expression of costimulatory or adhesion molecules except CD8. The differential expression of CD8 in these γδ CTL may be due to the affinity of TCR for the target, or to the density of antigenic epitopes. Measurement of the affinity of TCRγδ expressed in CD8+ and DN CTL clones, and detailed analysis of the target molecules of these γδ CTL, are now needed to clarify these issues.
Most of our sequenced γδ CTL clones were found to have two in-frame TCR γ chain transcripts. Further studies with transgenic mice are needed to determine which inframe γ chain is responsible for TL recognition, although surface expression of the Vγ2 chain is in doubt (30, 36). It has been speculated that the TL molecule, a member of the nonpolymorphic MHC class I molecules, may be recognized by a limited set of TCR (37). However, this seems to be unlikely since the present analysis showed diversity in the CDR3 region of γ chains, although analysis of δ chains is still necessary. The results suggest heterogeneity in antigenic epitopes and TL recognition by CTL as discussed below.
Tlaa-3, T3k, and T3b have identical amino acid sequences in their PBRs. In their whole proteins, however, Tlaa-3 and T3b differ at over 30 amino acid positions, while T3k and T3b are different at a single amino acid position, 276 in the transmembrane region (Fig. 1). T3k expressed in the intestine of C3H and Tla a -3 transgenic mice may not have any significance for the recognition of T3b, since a previous experiment suggested that T3b expressed in the intestine does not induce tolerance against T3b itself expressed in the skin (12). Identification of the antigenic epitopes recognized by γδ and αβ CTL against TL should reveal antigenic determinant(s) responsible for skin graft rejection. The CTL against TL were all TL-restricted, regardless of the C3H or Tg.Tlaa-3-2 origin and usage of TCRαβ or TCRγδ. There are several possible modes in which CTL can recognize TL molecules. For example, CTL may recognize a complex of TL and endogenous peptides, similar to conventional CTL recognition of classical MHC molecules (38), or a complex of TL and nonpeptide antigens such as CD1 plus lipids (39). CTL may interact with TL molecules without any contribution from bound peptides, possibly from the side of the TL molecule, away from the putative PBR of the α1 and α2 domains (40), or with a complex of TL molecules and peptides derived from TL itself. The CTL may be heterogeneous in TL recognition; some CTL may recognize TL + peptides while others may recognize TL itself. To determine how TL molecules are recognized by CTL, the established CTL clones will be tested further on various TL+ target cells including transfectants of Drosophila melanogaster cells expressing TL molecules devoid of binding endogenous peptides (41). Recent studies using a TAP-2–deficient cell line, RMA-S, have demonstrated that the transport of TL molecules to the cell surface is TAP-2 independent (31, 42), but these findings do not necessarily exclude the possibility that TL molecules bind antigenic peptides by other mechanisms. Our previous structural comparison of TL antigens with classical class I antigens suggested that TL molecules can form peptide-binding clefts (37). There has been a report that they bind to peptides modified at the amino termini (43), although an absence of peptides bound to TL has been also reported (42). The CTL assay is by far the most sensitive method for detecting antigenic peptides bound to MHC molecules (44), and its application should allow more complete elucidation of the function of TL in the immune system.
Acknowledgments
This work was supported in part by a Grant-in-Aid for Scientific Research on Priority Areas and a Grantin-Aid for General Scientific Research from the Ministry of Education, Science, Sports and Culture, Japan, and a Grant-in-Aid from the Ministry of Health and Welfare, Japan. This work was also supported in part by a grant from Naito Foundation and a grant from Imanaga Foundation.
Footnotes
We are indebted to Dr. L.J. Old for his continuous encouragement and advice thoughout these studies. We thank Y. Matsudaira, S. Ozeki, and H. Tamaki for their excellent technical assistance. We thank Dr. K. Furukawa, Dr. K. Kuribayashi, Dr. M. Miyasaka, Dr. M. Muto, Dr. E. Nakayama, Dr. S. Sakaguchi, and Dr. H. Yagita for their kind gifts of monoclonal antibodies.
1 Abbreviations used in this paper: B6, C57BL/6; C3H, C3H/He; cAb, conventional antibody; DN, CD4−CD8− double negative; Ltk−, thymidine kinase-negative L cell; MFI, mean fluorescence intensity; PBR, peptide binding region; RT, reverse transcription.
References
- 1.Klein, J. 1986. Natural History of the Major Histocompatibility Complex. John Wiley & Sons, Inc., New York. 775 pp.
- 2.Shawar SM, Vyas JM, Rodgers JR, Rich RR. Antigen presentation by major histocompatibility complex class I-bmolecules. Annu Rev Immunol. 1994;12:839–880. doi: 10.1146/annurev.iy.12.040194.004203. [DOI] [PubMed] [Google Scholar]
- 3.Simister NE, Mostov KE. An Fc receptor structurally related to MHC class I antigens. Nature (Lond) 1989;337:184–187. doi: 10.1038/337184a0. [DOI] [PubMed] [Google Scholar]
- 4.Haas W, Pereira P, Tonegawa S. Gamma/delta cells. Annu Rev Immunol. 1993;11:637–685. doi: 10.1146/annurev.iy.11.040193.003225. [DOI] [PubMed] [Google Scholar]
- 5.Bluestone JA, Khattri R, Sciammas R, Sperling AI. TCRγδ cells: a specialized T-cell subset in the immune system. Annu Rev Cell Dev Biol. 1995;11:307–353. doi: 10.1146/annurev.cb.11.110195.001515. [DOI] [PubMed] [Google Scholar]
- 6.Chen Y-T, Obata Y, Stockert E, Takahashi T, Old LJ. Tla-region genes and their products. Immunol Res. 1987;6:30–45. doi: 10.1007/BF02918102. [DOI] [PubMed] [Google Scholar]
- 7.Old LJ, Stockert E. Immunogenetics of cell surface antigens of mouse leukemia. Annu Rev Genet. 1977;11:127–160. doi: 10.1146/annurev.ge.11.120177.001015. [DOI] [PubMed] [Google Scholar]
- 8.Boyse EA, Old LJ, Luell S. Antigenic properties of experimental leukemias. II. Immunological studies in vivowith C57BL/6 radiation–induced leukemias. J Natl Cancer Inst. 1963;31:987–995. [PubMed] [Google Scholar]
- 9.Hershberg R, Eghtesady P, Sydora B, Brorson K, Cheroutre H, Modlin R, Kronenberg M. Expression of the thymus leukemia antigen in mouse intestinal epithelium. Proc Natl Acad Sci USA. 1990;87:9727–9731. doi: 10.1073/pnas.87.24.9727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wu M, Van Kaer L, Itohara S, Tonegawa S. Highly restricted expression of the thymus leukemia antigens on intestinal epithelial cells. J Exp Med. 1991;174:213–218. doi: 10.1084/jem.174.1.213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hamasima N, Takahashi T, Taguchi O, Nishizuka Y, Stockert E, Old LJ, Obata Y. Expression of TL, H-2, and chimeric H-2/TL genes in transgenic mice: abnormal thymic differentiation and T-cell lymphomas in a TL transgenic strain. Proc Natl Acad Sci USA. 1989;86:7995–7999. doi: 10.1073/pnas.86.20.7995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Morita A, Takahashi T, Stockert E, Nakayama E, Tsuji T, Matsudaira Y, Old LJ, Obata Y. TL antigen as a transplantation antigen recognized by TL-restricted cytotoxic T cells. J Exp Med. 1994;179:777–784. doi: 10.1084/jem.179.3.777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Obata Y, Taguchi O, Matsudaira Y, Hasegawa H, Hamasima N, Takahashi T. Abnormal thymic development, impaired immune function and γδ T cell lymphomas in a TL transgenic mouse strain. J Exp Med. 1991;174:351–362. doi: 10.1084/jem.174.2.351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Obata Y, Chen Y-T, Stockert E, Old LJ. Structural analysis of TLgenes of the mouse. Proc Natl Acad Sci USA. 1985;82:5475–5479. doi: 10.1073/pnas.82.16.5475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chen Y-T, Obata Y, Stockert E, Old LJ. Thymus-leukemia (TL) antigens of the mouse. Analysis of TL mRNA and TL cDNA from TL+ and TL−strains. J Exp Med. 1985;162:1134–1148. doi: 10.1084/jem.162.4.1134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ledbetter JA, Herzenberg LA. Xenogeneic monoclonal antibodies to mouse lymphoid differentiation antigens. Immunol Rev. 1979;47:63–90. doi: 10.1111/j.1600-065x.1979.tb00289.x. [DOI] [PubMed] [Google Scholar]
- 17.Dialynas DP, Quan ZS, Wall KA, Pierres A, Quintáns J, Loken MR, Pierres M, Fitch FW. Characterization of the murine T cell surface molecule, designated L3T4, identified by monoclonal antibody GK1.5: similarity of L3T4 to the human Leu-3/T4 molecule. J Immunol. 1983;131:2445–2451. [PubMed] [Google Scholar]
- 18.Kubo RT, Born W, Kappler JW, Marrack P, Pigeon M. Characterization of a monoclonal antibody which detects all murine αβ T cell receptors. J Immunol. 1989;142:2736–2742. [Google Scholar]
- 19.Itohara S, Nakanishi N, Kanagawa O, Kubo R, Tonegawa S. Monoclonal antibodies specific to native murine T-cell receptor γδ: analysis of γδ T cells during thymic ontogeny and in peripheral lymphoid organs. Proc Natl Acad Sci USA. 1989;86:5094–5098. doi: 10.1073/pnas.86.13.5094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Leo O, Foo M, Sachs DH, Samelson LE, Bluestone JA. Identification of a monoclonal antibody specific for a murine T3 polypeptide. Proc Natl Acad Sci USA. 1987;84:1374–1378. doi: 10.1073/pnas.84.5.1374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Takagaki Y, DeCloux A, Bonneville M, Tonegawa S. Diversity of γδ T-cell receptors on murine intestinal intraepithelial lymphocytes. Nature (Lond) 1989;339:712–714. doi: 10.1038/339712a0. [DOI] [PubMed] [Google Scholar]
- 22.Reilly EB, Kranz DM, Tonegawa S, Eisen HN. A functional γ gene formed from known γ-gene segments is not necessary for antigen-specific responses of murine cytotoxic T lymphocytes. Nature (Lond) 1986;321:878–880. doi: 10.1038/321878a0. [DOI] [PubMed] [Google Scholar]
- 23.Guy-Grand D, Cerf-Bensussan N, Malissen B, Malassis-Seris M, Briottet C, Vassalli P. Two gut intraepithelial CD8+lymphocyte populations with different T cell receptors: a role for the gut epithelium in T cell differentiation. J Exp Med. 1991;173:471–481. doi: 10.1084/jem.173.2.471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lefrancois L. Phenotypic complexity of intraepithelial lymphocytes of the small intestine. J Immunol. 1991;147:1746–1751. [PubMed] [Google Scholar]
- 25.Bandeira A, Itohara S, Bonneville M, Burlen-Defranoux O, Mota-Santos T, Coutinho A, Tonegawa S. Extrathymic origin of intestinal intraepithelial lymphocytes bearing T-cell antigen receptor γδ. Proc Natl Acad Sci USA. 1991;88:43–47. doi: 10.1073/pnas.88.1.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pereira P, Gerber D, Huang SY, Tonegawa S. Ontogenic development and tissue distribution of Vγ1expressing γ/δ T lymphocytes in normal mice. J Exp Med. 1995;182:1921–1930. doi: 10.1084/jem.182.6.1921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Whetsell M, Mosley RL, Whetsell L, Schaefer FV, Miller KS, Klein JR. Rearrangement and junctionalsite sequence analyses of T-cell receptor gamma genes in intestinal intraepithelial lymphocytes from murine athymic chimeras. Mol Cell Biol. 1991;11:5902–5909. doi: 10.1128/mcb.11.12.5902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Iwamoto A, Rupp F, Ohashi PS, Walker CL, Pircher H, Joho R, Hengartner H, Mak TW. T cell-specific γ genes in C57BL/10 mice. Sequence and expression of new constant and variable region genes. J Exp Med. 1986;163:1203–1212. doi: 10.1084/jem.163.5.1203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lafaille JJ, DeCloux A, Bonneville M, Takagaki Y, Tonegawa S. Junctional sequences of T cell receptor γδ genes: implications for γδ T cell lineages and for a novel intermediate of V-(D)-J joining. Cell. 1989;59:859–870. doi: 10.1016/0092-8674(89)90609-0. [DOI] [PubMed] [Google Scholar]
- 30.Pereira P, Gerber D, Regnault A, Huang SY, Hermitte V, Coutinho A, Tonegawa S. Rearrangement and expression of Vγ1, Vγ2 and Vγ3 TCR γ genes in C57BL/6 mice. Int Immunol. 1996;8:83–90. doi: 10.1093/intimm/8.1.83. [DOI] [PubMed] [Google Scholar]
- 31.Rodgers JR, Mehta V, Cook RG. Surface expression of β2-microglobulin–associated thymus-leukemia antigen is independent of TAP2. Eur J Immunol. 1995;25:1001–1007. doi: 10.1002/eji.1830250421. [DOI] [PubMed] [Google Scholar]
- 32.Steinmetz M, Winoto A, Minard K, Hood L. Clusters of genes encoding mouse transplantation antigens. Cell. 1982;28:489–498. doi: 10.1016/0092-8674(82)90203-3. [DOI] [PubMed] [Google Scholar]
- 33.Boyse EA, Flaherty L, Stockert E, Old LJ. Histocompatibility attributable to genes near H-2that are not revealed by hemagglutination or cytotoxicity tests. Transplantation (Baltimore) 1972;13:431–432. doi: 10.1097/00007890-197204000-00013. [DOI] [PubMed] [Google Scholar]
- 34.Teitell M, Mescher MF, Olson CA, Littman DR, Kronenberg M. The thymus leukemia antigen binds human and mouse CD8. J Exp Med. 1991;174:1131–1138. doi: 10.1084/jem.174.5.1131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Anderson SJ, Levin SD, Perlmutter RM. Involvement of the protein tyrosine kinase p56lckin T cell signaling and thymocyte development. Adv Immunol. 1994;56:151–178. doi: 10.1016/s0065-2776(08)60451-4. [DOI] [PubMed] [Google Scholar]
- 36.Takagaki Y, Nakanishi N, Ishida I, Kanagawa O, Tonegawa S. T cell receptor-γ and -δ genes preferentially utilized by adult thymocytes for the surface expression. J Immunol. 1989;142:2112–2121. [PubMed] [Google Scholar]
- 37.Obata Y, Satta Y, Moriwaki K, Shiroishi T, Hasegawa H, Takahashi T, Takahata N. Structure, function, and evolution of mouse TLgenes, nonclassical class I genes of the major histocompatibility complex. Proc Natl Acad Sci USA. 1994;91:6589–6593. doi: 10.1073/pnas.91.14.6589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Townsend A, Bodmer H. Antigen recognition by class I–restricted T lymphocytes. Annu Rev Immunol. 1989;7:601–624. doi: 10.1146/annurev.iy.07.040189.003125. [DOI] [PubMed] [Google Scholar]
- 39.Beckman EM, Porcelli SA, Morita CT, Behar SM, Furlong ST, Brenner MB. Recognition of a lipid antigen by CD1-restricted αβ+T cells. Nature (Lond) 1994;372:691–694. doi: 10.1038/372691a0. [DOI] [PubMed] [Google Scholar]
- 40.Schild H, Mavaddat N, Litzenberger C, Ehrich EW, Davis MM, Bluestone JA, Matis L, Draper RK, Chien Y-H. The nature of major histocompatibility complex recognition by γδ T cells. Cell. 1994;76:29–37. doi: 10.1016/0092-8674(94)90170-8. [DOI] [PubMed] [Google Scholar]
- 41.Jackson MR, Song ES, Yang Y, Peterson PA. Empty and peptide-containing conformers of class I major histocompatibility complex molecules expressed in Drosophila melanogastercells. Proc Natl Acad Sci USA. 1992;89:12117–12121. doi: 10.1073/pnas.89.24.12117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Holcombe HR, Castaño AR, Cheroutre H, Teitell M, Maher JK, Peterson PA, Kronenberg M. Nonclassical behavior of the thymus leukemia antigen: peptide transporter–independent expression of a nonclassical class I molecule. J Exp Med. 1995;181:1433–1443. doi: 10.1084/jem.181.4.1433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sharma P, Joyce S, Chorney KA, Griffith JW, Bonneau RH, Wilson FD, Johnson CA, Flavell RA, Chorney MJ. Thymus-leukemia antigen interacts with T cells and self-peptides. J Immunol. 1996;156:987–996. [PubMed] [Google Scholar]
- 44.Joyce S, Nathenson SG. Methods to study peptides associated with MHC class I molecules. Curr Opin Immunol. 1994;6:24–31. doi: 10.1016/0952-7915(94)90029-9. [DOI] [PubMed] [Google Scholar]