

Abstract
Tip-growing pollen tubes provide a useful model system to study polar growth. Although roles for tip-focused calcium gradient and tip-localized Rho-family GTPase in pollen tube growth is established, the existence and function of tip-localized F-actin have been controversial. Using the green fluorescent protein–tagged actin-binding domain of mouse talin, we found a dynamic form of tip-localized F-actin in tobacco pollen tubes, termed short actin bundles (SABs). The dynamics of SABs during polar growth in pollen tubes is regulated by Rop1At, a Rop GTPase belonging to the Rho family. When overexpressed, Rop1At transformed SAB into a network of fine filaments and induced a transverse actin band behind the tip, leading to depolarized growth. These changes were due to ectopic Rop1At localization to the apical region of the plasma membrane and were suppressed by guanine dissociation inhibitor overexpression, which removed ectopically localized Rop1At. Rop GTPase–activating protein (RopGAP1) overexpression, or Latrunculin B treatments, also recovered normal actin organization and tip growth in Rop1At-overexpressing tubes. Moreover, overexpression of RopGAP1 alone disrupted SABs and inhibited growth. Finally, SAB oscillates and appears at the tip before growth. Together, these results indicate that the dynamics of tip actin are essential for tip growth and provide the first direct evidence to link Rho GTPase to actin organization in controlling cell polarity and polar growth in plants.
Keywords: Rho GTPase, cell polarity, polar growth, actin cytoskeleton, RopGAP
Introduction
The pollen tube has been used as a model system to study tip growth and the actin cytoskeleton for decades because of its uniform cell morphology, rapid growth, and the abundance of actin and its associated proteins (Ren et al. 1997; Taylor and Hepler 1997; Yang 1998). Numerous studies have demonstrated the presence of a tip-focused gradient of cytosolic calcium and its critical role in tip growth (for review see Franklin-Tong 1999). The actin cytoskeleton has also been extensively studied in pollen, but its role is less clear. Extensive axial actin cables are found in the shank of tubes and are believed to be essential for cytoplasmic streaming and to provide a track for organelle movement (for review see Vidali and Hepler 2000).
Unlike in yeast and algal systems, where cortical actin patches are known to establish sites for polar growth (Alessa and Kropf 1999; Kropf et al. 1999; Bahler and Peter 2000; Fowler 2000; Pruyne and Bretscher 2000), the existence and the function of F-actin at the tip of pollen tubes has been controversial. Early studies using chemically fixed cells and fluorescent phalloidin suggested the presence of a dense actin network at the tip (Derksen et al. 1995). However, rapid freezing, freeze substitution, and electron and light microscopy techniques revealed no dense actin network, but instead only occasional sparse fine actin filaments at the tip (Lancelle et al. 1987; Doris and Steer 1996). Similarly, no tip actin network was seen in live lily pollen tubes injected with fluorescence-labeled phalloidin or tobacco pollen tubes expressing a mouse talin (mTalin) tagged with green fluorescence protein (GFP; Miller et al. 1996; Kost et al. 1998). These studies have led to the proposition of an apparent actin-free zone at the tip of pollen tubes (Kost et al. 1998; Vidali and Hepler 2000; Zheng and Yang 2000a). An actin-free tip has also been described in other tip-growing cells, such as root hairs (Miller et al. 1999; Emmons and de Ruijter 2000).
Recent studies instead suggest the presence of F-actin structures just behind the apical dome of pollen tubes. In tobacco, an actin ring was reported in the subapical region of pollen tubes expressing GFP-talin (Kost et al. 1998). A “collar” of F-actin at ∼5 μm behind the tip and a dense meshwork of actin filaments in the collar region were observed in chemically fixed maize and Papaver pollen tubes, respectively (Gibbon et al. 1999; Geitmann et al. 2000). Similarly, fine actin bundles were also observed in the collar of root hair cells (Miller et al. 1999). Finally, a cortical ring of F-actin is present at the base of the dome in rhizoid tips of young algal embryos (Kropf et al. 1998). These subapical actin structures seem to be commonly composed of fine actin filaments distinct from thick axial F-actin cables associated with cytoplasmic streaming back from the tip.
Pharmacological studies suggest that subapical F-actin structures may have a role distinct from that for axial actin cables. Low concentrations of cytochalasin D disrupt subapical fine actin bundles and inhibit root hair elongation, but do not affect axial actin cables and streaming (Miller et al. 1999). Similar effects are seen in maize pollen tubes treated with 5–10 nM Latrunculin B (Gibbon et al. 1999). In both cases, disruption of the subapical actin structures causes extension of axial actin cables to the extreme tip. It was proposed that the subapical actin acts either to target vesicles to the tip or to maintain proper cytoplasmic organization at the tip (Kost et al. 1998; Miller et al. 1999), but its precise role is unknown. Furthermore, it remains possible that there is an additional population of dynamic tip F-actin that is sensitive to low concentrations of Latrunculin B or cytochalasin D, but is not detected by the existing actin-staining techniques, as proposed by Gibbon et al. 1999. Thus, development of sensitive techniques for visualizing F-actin dynamics and studies of the mechanism underlying its dynamics are needed to clarify the issue of a role for F-actin in tip growth.
It is well established that the organization and dynamics of the actin cytoskeleton are controlled by the RHO family of small GTPases in yeast and animal systems (Hall 1998; Li and Yang 2000). In mammals, each of the three major RHO GTPases, Rho, Cdc42, and Rac, is known to regulate specific forms of F-actin and associated cellular processes, e.g., cell morphogenesis and movement (Ridley and Hall 1992; Ridley et al. 1992; Chant and Stowers 1995; Nobes and Hall 1995; Hall 1998). Plants do not possess any of these three subfamilies of Rho GTPases, but have evolved a distinct subfamily, termed Rop (Zheng and Yang 2000b). Rop is encoded by a multigene family in Arabidopsis (Li et al. 1998). Studies using the pollen-specific Rop1At and its homologues show that Rop is localized to the apical region of the tube plasma membrane (PM) and acts as a central switch to control tip growth in pollen tubes (Lin et al. 1996; Lin and Yang 1997; Li et al. 1998, Li et al. 1999; Kost et al. 1999). For example, blocking of Rop signaling by expression of dominant negative (DN)-rop1At mutants caused inhibition of pollen tube elongation, whereas constitutively active (CA)-rop1At mutants induced isotropic growth.
Overexpression of CA-Atrac2/rop5At mutants caused axial actin cables to become spiral cables, whereas DN-Atrac2/rop5At mutants induced apparent reduction in the thickness of actin cables in tobacco pollen tubes (Kost et al. 1999). However, these changes alone cannot explain the dramatic phenotypes induced by the dominant mutants (Kost et al. 1999). Furthermore, Rop is localized to the tip and evidence suggests that Rop is only activated at the apex (Lin et al. 1996). Because the effectors of Rho GTPases have been shown to directly regulate the activity of actin-binding proteins, we suspected that Rop had a role in regulating the organization or the dynamics of tip-localized F-actin, the actin that had not been detected previously.
In this report, we demonstrate the presence of a dynamic form of tip-localized F-actin in living tobacco pollen tubes using the actin-binding domain of mTalin tagged with an enhanced GFP mutant. Importantly, using a combined genetic and chemical approach, we show that the dynamics of tip actin are regulated by Rop signaling and are critical for cell polarity development and tip growth in pollen tubes.
Materials and Methods
DNA Manipulation and Plasmid Constructs
All plasmids used for transient expression in pollen were constructed in a derivative of the pBI221 vector (CLONTECH Laboratories, Inc.), termed pLAT52, in which CaMV 35S::GUS was replaced with the LAT52 promoter (Twell et al. 1991). The enhanced GFP mutant (S65C) gene was cloned downstream of the LAT52 promoter in pLAT52 to create pLAT52:: GFP as described previously (Li et al. 1999). To generate a pLAT52::GFP-mTalin construct, DNA sequence encoding the COOH–terminal actin-binding domain of the mTalin (amino acid residues 2430–2541) was amplified from a mouse EST clone (EMBL/GenBank/DDBJ under accession no. AA615519) using primers 5′-GCAGATCTAACTTTGAGGAACAAATCC and 3′-GCGAGCTCCACAGGGCTTTTTTAGTGC and cloned as translational fusion to the COOH terminus of GFP in pLAT52::GFP. Constructs for transient expression of all other genes, including Rop1At (Li et al. 1999), DN-rop1At (Li et al. 1999), RopGAP1 (Wu et al. 2000), and GDI1 (Bischoff et al. 2000), were also cloned in pLAT52. Rop1At, DN-rop1At, and GDI1 were cloned downstream of the LAT52 promoter using BglII-SstI sites, whereas Nco1-Kpn1 sites were used for the subcloning of Rop GTPase–activating protein 1 (RopGAP1) in pLAT52. For particle bombardment, all plasmids were amplified in Escherichia coli strain top 10 and purified using plasmid Midi or Mini kits according to the manufacturer's instruction (QIAGEN).
Particle Bombardment–mediated Transient Expression in Tobacco Pollen
Nicotiana tabacum plants were grown in growth chambers at 22°C under a light regime of 12 h darkness and 12 h light. Pollen grains collected from these plants were used for transient expression using a particle bombardment procedure (Bate et al. 1996). Pollen grains were collected immediately before each experiment and suspended in a pollen germination medium (GM) containing 5 μM CaCl2, 5 μM Ca(NO3)2, 1 mM Mg(SO4)2, 0.01% H3BO3, and 18% sucrose, pH 6.5–7.0. A drop of 80–100 μl medium containing pollen grains from three flowers was applied to a piece of 2 × 2–cm nylon membrane (Micron Separations, Inc.) placed on top of a piece of 90-mm filter paper (Whatman) in a 100 × 15–mm petri dish. Once liquid was drained from the membrane, an additional 1.5 ml GM was added to the filter paper. Pollen grains were then immediately bombarded with DNA-coated gold particles using a PDS-1000/He particle delivery system (Bio-Rad Laboratories). The microcarrier launch assembly was placed in the second slot from the top (level 2), and the target cells were placed at level 4 or 5 (the distance from the stopping screen to target cells is ∼8 or 11 cm, respectively). Rupture disks of 1,100 psi were chosen to accelerate macrocarriers under a vacuum of 27 inches of mercury.
Gold particles (1.0 μm diameter) were coated with plasmid DNA according to the manufacturer's procedures (Bio-Rad Laboratories) immediately before bombardment. Routinely, 0.5-mg particles were coated with 0.8 μg of pLAT52::GFP-mTalin DNA or a mixture of 0.8 μg of pLAT52::GFP-mTalin DNA with 0.4 μg of a second plasmid DNA containing a gene of interest. As control, the pLAT52 vector DNA was cobombarded with the pLAT52::GFP-mTalin.
Bombarded pollen grains were incubated at room temperature for 30 min before being washed into petri dishes with 0.5 ml GM. The pollen grains were then either treated with Latrunculin B or incubated for an additional 3–6 h before observation under a inverted microscope (model TE300; Nikon) equipped with a cooled CCD camera (model C4742-95; Hamamatsu) or confocal microscope as described below.
Drug Treatments
To determine the effect of caffeine or Latrunculin B on the actin organization in tobacco pollen tubes, pollen grains bombarded with pLAT52::GFP-mTalin were incubated for 3.5 h before the addition of drugs. For caffeine treatment, a stock solution of 0.5 M caffeine (Sigma-Aldrich) was added to germinated pollen to a final concentration of 0.3 mM. For Latrunculin B treatment, a stock solution of 5 mM Latrunculin B (Calbiochem) prepared in anhydrous DMSO (Sigma-Aldrich) was added to germinated pollen for a final concentration of 5 nM. Because the final concentration of DMSO in the medium was 1 ppm after Latrunculin B was added, an equivalent concentration of DMSO was added to the medium of all untreated controls. Pollen tubes were incubated for an additional 30 min before observation.
To study the effects of Latrunculin B treatment on pollen tubes overexpressing Rop1At or DN-rop1At, Latrunculin B was added to germinated pollen 1.5 h after bombardment. Treated pollen tubes were incubated for an additional 3 h before being analyzed for polar growth or actin organization as described below.
Visualization and Analyses of F-actin using GFP-mTalin and Confocal Microscopy
To visualize F-actin in pollen tubes, the GFP-mTalin chimeric gene was transiently expressed in pollen as described above. Tubes expressing GFP-mTalin were identified using epifluorescence microscopy and observed either under an OPTIPHOT upright microscope (Nikon) equipped with an MRC 600 laser scanning device (Bio-Rad Laboratories) or an Axioplan2 microscope equipped with an LSM 510 laser scanning system (ZEISS). In either case, 1-μm optical sections were scanned and captured using Comos or LSM 510 software, respectively. Confocal images were analyzed using the Metamorph v4.5 software (Universal Imaging Corp.) and processed using Adobe Photoshop® v5.5. For three-dimensional reconstruction, serial optical sections were taken for each tube. For a given treatment, 40–70 pollen tubes were observed from several independent experiments and the percentage of tubes with a specific actin-staining pattern was determined. For time–sequence analyses, either midplane sections or cortical sections of the tip were scanned every 15 s.
Analyses of Correlation between Tip F-actin and Elongation Rates
To analyze the correlation between tip-localized F-actin and pollen tube elongation, we used the time serials of midplane optical sections scanned every 15 s from pollen tubes expressing GFP-mTalin. Using Metamorph v4.5, a region of the apical dome (17.5 μm2, ∼4 μm from the extreme tip) was created. Using the measurement function, the average fluorescence intensity in this region was measured and subtracted from background fluorescence, which was determined using background function in a region created from the darkest area near the tip of the tube. The corrected average intensity of GFP fluorescence was used to represent relative amounts of F-actin at the tip. Elongation rates were determined from the same images used for measuring apical F-actin. Using the Metamorph v4.5 measurement function, the distance between the base and the leading edge of the images was determined. Pollen tube elongation rates (μm/min) were calculated as differences in distances between two images over the time interval of 15 s.
Morphological Analyses of Pollen Tubes
To determine the effect of Latrunculin B treatment on changes in polar growth of pollen tubes caused by Rop1At or DN-rop1At overexpression, we measured the width and length of pollen tubes cobombarded with the GFP-mTalin construct after Latrunculin B treatment. Exactly 3 h after pollen tubes were treated with different concentrations of Latrunculin B as described above, images of fluorescent tubes were rapidly recorded through a cooled CCD camera (model C4742-95; Hamamatsu) attached on an Eclipse inverted microscope (model TE300; Nikon). The images were analyzed using the Metamorph v4.5 measurement function. The degree of depolarized growth was determined by measuring the diameter of the widest region of the tube, and the degree of polar growth was determined by measuring the length of pollen tubes. For each treatment, data were collected from three independent experiments (40–80 tubes). Data were subjected to a statistic test using Sigmastat (Hallogram).
Online Supplemental Material
A video (available at http://www.jcb.org/cgi/content/full/152/5/1019/DC1) of images of the tip-localized short actin bundle (SAB) was prepared using QuickTime® v4.0. The images were a time serial (15 s intervals for a period of 12 min) of mid-plane optical sections of a tube expressing GFP-mTalin. Images for the first 2 min 45 s are also shown in Fig. 1 C.
Figure 1.
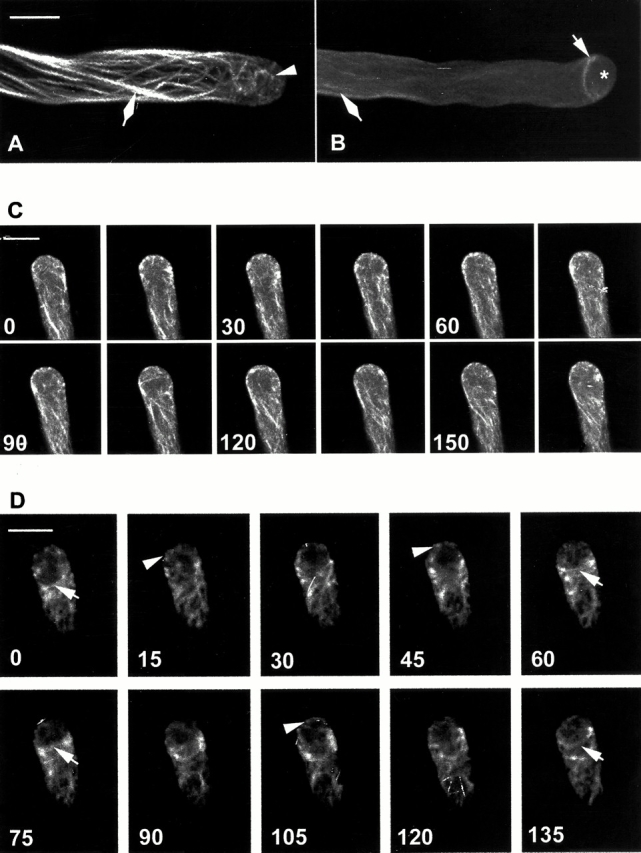
GFP-mTalin reveals dynamic F-actin in the apical and subapical regions of pollen tubes. GFP-mTalin was transiently expressed in tobacco pollen tubes, visualized using confocal microscopy, and analyzed using Metamorph v4.5 as described in Materials and Methods. (A) A projection of 1-μm optical sections showing typical actin organization in pollen tubes. Note SAB in the tip and intense actin cables. (B) A projection of 1-μm optical sections showing a less typical tube. Note an F-actin collar located just behind the apex and less intense actin cables. (C) A time serial of midplane optical sections (1 μm) of a tobacco pollen tube (an extended time serial of the same tube can been seen in the video available at http://www.jcb.org/cgi/content/full/152/5/1019/DC1). Images were collected every 15 s. Note the dynamics of cortical SAB seen as bright dots or patches. Numbers at the lower left corner of each image indicate seconds after the capturing of the first time point (0). (D) A time serial of optical sections (1 μm) through a cortical region near the very tip of a tube. Optical sections were obtained from a region between the midplane and the surface of the tube flank every 15 s. The actin collar behind the tip is generally associated with no or reduced SAB at the tip. Arrowhead, SAB at the tip; arrow, actin collar behind the apical dome; asterisk, apical region lacking F-actin; diamond, axial actin cables. Bars, 10 μm.
Results
F-actin Is Present at the Tip of Pollen Tubes and Is Highly Dynamic
To visualize actin dynamics in pollen tubes, we constructed a chimeric gene encoding an enhanced GFP mutant (Reichel et al. 1996) fused to the actin-binding domain of the mTalin (GFP-mTalin; McCann and Craig 1997). The enhanced GFP (S65C) was used to increase detection sensitivity, which we believed was critical for visualizing fine or dynamic actin filaments. The GFP-mTalin chimeric gene was expressed under the pollen-specific LAT52 promoter using particle bombardment–mediated transient expression (Twell et al. 1991; Bate et al. 1996). As shown previously for a similar construct (Kost et al. 1998), expression of our GFP-mTalin gene did not significantly affect pollen tube growth and morphology. Projection of optical sections revealed several types of microfilaments, including a subapical ring or collar-like actin structure and axial actin cables as described previously (Fig. 1; Kost et al. 1998). The subapical actin, termed actin collar herein, is localized just behind the apical dome and was found in ∼24% of tubes expressing GFP-mTalin (Fig. 1 B). We also observed short bundles or patches apparently composed of short and fine actin filaments in the extreme apex in 85% of tubes examined (Fig. 1 A). We termed this F-actin short actin bundle (SAB).
Our observations suggest that the subapical actin collar and apical SAB may represent one or more populations of highly dynamic F-actin. Thus, we performed a temporal analysis of GFP-mTalin localization by scanning the midplane or the cortex of the apical region. Because repeated laser scanning caused fluorescence bleaching and growth inhibition, we had to limit our minimal time intervals of laser scanning to 15 s, which allowed visualizing F-actin over a period of 5–10 min without significant fluorescence bleaching or growth inhibition. As shown in Fig. 1C and Fig. D (Fig. 1 C is also shown in a video, available at http://www.jcb.org/cgi/content/full/152/5/1019/DC1), both SAB and the actin collar were very dynamic. The appearance of these structures seems to alternately oscillate every ∼60–75 s in actively growing tubes, i.e., the appearance of SAB was generally associated with reduction or disappearance of the actin collar and vice versa. These results support the notion that these two types of actin structures belong to the same actin population that is subject to both temporal and spatial regulation.
Overexpression of DN-rop1At Mutants Disrupted SAB at the Tip but Induced a Transverse Actin Band behind the Tip
To assess the involvement of Rop in the regulation of tip F-actin, we coexpressed GFP-mTalin with a DN-rop1At mutant in tobacco pollen tubes, which has been shown to inhibit tube growth and cause slight tube expansion when overexpressed in Arabidopsis pollen (Li et al. 1999). DN-rop1At transient expression caused similar phenotypes in tobacco pollen tubes (Fig. 2A and Fig. B). Importantly, DN-rop1At expression also disrupted the formation of apical SAB, suggesting Rop may control SAB formation. Nearly 60% of DN-rop1At tubes contained no SAB, and ∼32% of them had drastically reduced SAB levels; only 8% of pollen tubes were seen with a similar amount of apical actin as wild-type tubes. We did not observe significant changes in axial actin cables in DN-rop1At tubes (Fig. 2 B). Surprisingly, we also observed a transverse actin band composed of cortical actin hoops just behind the apical dome in DN-rop1At cells. Time–sequence analyses showed that the transverse actin band was quite stable and actually widened over time, apparently due to actin polymerization behind the apical dome (data not shown). The DN-rop1At–induced transverse actin band could be caused by DN-rop1At–induced growth inhibition, direct effects of DN-rop1At, or indirect effects of DN-rop1At that result in increased Rop activity behind the tip, because DN-rop1At overexpression causes ectopic localization of wild-type Rop proteins in the subapical region of the PM (Wu, G., Y. Fu, and Z. Yang, unpublished data).
Figure 2.

DN-rop1At overexpression disrupts SAB and induces a transverse actin band behind the tip. Tobacco pollen grains were cobombarded with LAT52::GFP-mTalin and LAT52::DN-rop1At constructs (A and B) or bombarded with LAT52::GFP-mTalin alone before treatment with 5 nM Latrunculin B (C) or 300 nM caffeine (D). GFP-mTalin was analyzed by confocal microscopy as described in the legend to Fig. 1. All images shown are projections of 1-μm confocal sections. Arrow, transverse actin bands behind the apex; asterisk, F-actin free apex; diamond, actin cables. Bar, 10 μm.
To determine whether growth inhibition was responsible for transverse actin band formation, we treated pollen tubes with Latrunculin B or caffeine. As shown in Fig. 2C and Fig. D, neither caffeine (3 mM) nor Latrunculin B (5 nM) induced a transverse actin band, although both drugs inhibited tube elongation dramatically (see Table ), indicating that growth inhibition itself does not cause transverse actin band formation. Latrunculin B treatment disrupted both the actin collar and SAB, leading to the protrusion of actin cables to the extreme tip (Fig. 2 D and 5 D). These results also suggest that the subapical actin collar and apical SAB represent the population of F-actin that is highly sensitive to Latrunculin B, as proposed by Gibbon et al. 1999.
Table 1.
Effect of 5 nM Latrunculin B on Shape and Growth of Pollen Tubes
| 0 nM Latrunculin B (mean ± SD) | 5 nM Latrunculin B (mean ± SD) | |||||
|---|---|---|---|---|---|---|
| Width | Length | Width/Length | Width | Length | Width/Length | |
| μm | μm | μm | μm | |||
| Control | 9.2 ± 0.2 | 352.0 ± 23.5 | 0.03 ± 0.00 | 9.5 ± 0.3 | 201.6 ± 9.2 | 0.06 ± 0.01 |
| DN-rop1AT | 14.2 ± 0.7 | 96.4 ± 9.6 | 0.19 ± 0.02 | 11.7 ± 1.4 | 127.7 ± 17.2 | 0.10 ± 0.01 |
| Rop1A | 18.8 ± 1.9 | 339.1 ± 14.5 | 0.07 ± 0.01 | 11.5 ± 0.9 | 438.4 ± 40.9 | 0.03 ± 0.00 |
Rop1At Overexpression Induced the Formation of an Apical Actin Network and Transverse Actin Band
If the abnormal transverse actin band is caused by Rop proteins ectopically localized to PM behind the apex, we expect that overexpression of wild-type Rop1At would also induce the transverse actin band. Indeed, Rop1At overexpression resulted in an actin band very similar to that induced by DN-rop1At overexpression (Fig. 3). However, the cortical actin band in Rop1At-overexpressing cells was somewhat narrower than seen with DN-rop1At tubes (Fig. 3). More importantly, Rop1At overexpression induced the formation of an apical actin network composed of fine long actin filaments in place of SAB seen in wild-type tubes. This is in sharp contrast with the F-actin–free apex in DN-rop1At–overexpressing tubes.
Figure 3.

Rop1At overexpression induces a network of F-actin filaments at the tip and a transverse actin band behind the tip. GFP-mTalin was transiently expressed alone (A) or coexpressed with Rop1At (B and C) in tobacco pollen tubes, visualized, and analyzed as described in the legend to Fig. 2. Rop1At overexpression causes a network of F-actin composed of fine long filaments (arrowhead) in the tip and a subapical actin band (arrow) composed of thicker transverse hoops of actin filaments. C shows a time serial of the Rop1At-induced transverse actin band formation. The first projection (time 0) was collected 3 h after bombardment and subsequent imaging was performed every 5 min (minutes indicated at lower left corner). Transverse actin band appears to initiate near the tip as aggregates of SABs at time 0 when the shape of the tip is normal. By 5 min, transverse actin band is already formed but no significant expansion of the tip has occurred yet. Transverse actin band becomes wider, accompanied by the swelling of the tip (20 min). Bars, 15 μm.
Because Rop1At overexpression induces tube expansion or tip swelling, we sought to assess the causal relationship between depolarized growth and the transverse actin band. Time–sequence analyses indicate that the transverse actin band appeared before or at the onset of tip swelling in tubes overexpressing Rop1At. As shown in Fig. 3 C, a few transverse actin filaments began to accumulate near the apex at time 0, when the shape of the tube was still normal. At 5 min, an intense transverse actin band was already formed, whereas the tip just started to expand slightly. In the next 15 min, the tip had expanded considerably, whereas the intensity of the transverse actin band did not increase accordingly. These results suggest that transverse actin band formation was not the direct consequence of depolarization of tube growth, but was directly induced by ectopically localized Rop1At as a result of its overexpression (Li et al. 1999). This conclusion is further supported by the fact that DN-rop1At tubes were less expanded than Rop1At tubes but had more intense and wider actin bands (see Fig. 2).
Guanine Nucleotide Dissociation Inhibitor Suppressed Transverse Actin Band Formation Induced by Rop1At or DN-rop1At
To test the hypothesis that the transverse actin band is caused by ectopically localized Rop1At, we investigated the effect of guanine nucleotide dissociation inhibitor (GDI) overexpression on actin organization. In mammalian cells, GDIs act to recycle Rho GTPases from PM, and thus we suspect that Arabidopsis GDIs may have a similar role in pollen tubes. We found that overexpression of an Arabidopsis GDI in tobacco pollen tubes indeed suppressed transverse actin band formation (Fig. 4, B–E). As shown in Fig. 4 H, the transverse actin band was observed in 65% of tubes overexpressing Rop1At alone, but only in 20% of tubes cooverexpressing Rop1At and GDI. Similarly, 59% of tubes overexpressing DN-rop1At had a transverse actin band, whereas cooverexpression with GDI reduced the transverse actin band–containing tubes to 3.7% (Fig. 4 H). The GDI suppression of Rop1At-induced depolarized growth was tightly correlated with that of the transverse actin band (i.e., none of the tubes devoid of the transverse actin band had swollen tips or expanded tubes). Interestingly, a meshwork composed of SABs was observed in tubes cooverexpressing Rop1At and GDI, whereas tubes cooverexpressing DN-rop1At and GDI had an actin-free apex, as in tubes overexpressing DN-rop1At alone. Hence, these results not only provide evidence that ectopically localized Rop is responsible for the formation of abnormal actin bands behind the apex, but also suggest that Rop1At is involved in the regulation of SABs found in the extreme apex.
Figure 4.



GDI and RopGAP1 suppress transverse actin band formation induced by Rop1At or DN-rop1At overexpression. Coexpression of GFP-mTalin with one or more constructs was conducted, and GFP images were captured and analyzed as described in the legend to Fig. 3. (A) A control tube shows SAB at the tip. (B) A Rop1At-overexpressing tube shows a network of apical fine actin and subapical transverse actin band. (C) A tube overexpressing DN-rop1At shows clear apex and subapical transverse actin band. (D) No transverse actin band is observed in a tube cooverexpressing Rop1At with GDI, but extensive SABs are seen. The tube is less expanded than tubes overexpressing Rop1At alone. (E) Neither transverse actin band nor SAB is found in a tube cooverexpressing DN-rop1At and GDI. This tube is less expanded than the one overexpressing DN-rop1At alone. (F) No SABs are observed in tubes overexpressing RopGAP1. (G) A tube cooverexpressing Rop1At and RopGAP1. No transverse actin band is found, but a subapical actin collar (arrowhead) and weak apical SAB are recovered. (H) Frequency of transverse actin bands in tubes overexpressing DN-rop1At or Rop1At alone and in tubes cooverexpressing one of these genes with GDI. (I) Rop1At-induced transverse actin band is suppressed by RopGAP1 in a dose-dependent manner. 800 ng of LAT52::Rop1At plasmid DNA was mixed with indicated ratios of LAT52:RopGAP1 plasmid DNA and used for cobombardment with LAT52::GFP-mTalin. Arrowhead, SAB or actin network at the tip; arrow, actin collar or transverse actin band behind the apex; asterisk, F-actin–free apex; CK, tubes expressing GFP-mTalin alone. Bar, 10 μm.
RopGAP1 Overexpression Disrupted SABs at the Tip and Suppressed DN-rop1At– or Rop1At-induced Transverse Actin Band Formation
To further understand the role of Rop1At activity in regulating the dynamics of F-actin in pollen tubes, we investigated the effect of overexpressing RopGAPs on actin organization in pollen tubes. Biochemical studies indicate that RopGAP1 is a negative regulator of Rop1At by promoting the conversion of the GTP-bound form of Rop1At to the GDP-bound form (Wu et al. 2000).
Overexpression of RopGAP1 alone reduced or disrupted SAB, but had no effects on axial actin cables (Fig. 4 F), analogous to the effect of DN-rop1At overexpression. However, in contrast to DN-rop1At, RopGAP1 overexpression did not cause transverse actin band formation behind the apex (Fig. 4 F), further confirming that the actin band was not induced by Rop1At inactivation or by tube elongation inhibition as shown above. On the contrary, the actin collar observed in normal tubes was never found in RopGAP1-overexpressing tubes. The disruption of both SAB and the actin collar is tightly associated with the reduction in both elongation and expansion of pollen tubes induced by RopGAP1 overexpression (Fig. 4; Wu, G., Y. Fu, and Z. Yang, unpublished data).
When RopGAP1 was cooverexpressed with Rop1At, the transverse actin band was no longer formed (Fig. 4 G). As shown in Fig. 4 I, increasing the ratio of RopGAP1 to Rop1At led to a decreasing percentage of tubes containing the cortical actin band. The absence of the transverse actin band was consistently associated with the actin pattern seen in wild-type tubes, i.e., the presence of the dynamic SAB and actin collar. Furthermore, the reversal of Rop1At-induced actin pattern by RopGAP1 was also tightly associated with the restoration of wild-type pollen tube morphology and elongation rate. These results strongly suggest that Rop regulates polar growth at least in part through the control of the organization of dynamic F-actin at the tip of pollen tubes. These observations also suggest that the tip-localized activation of Rop most likely turns on actin assembly at the tip.
Acting as a negative control, cooverexpression with a RopGAP mutant (in which Rop interaction domains were removed) or an empty vector did not affect the changes in actin organization and tube morphology induced by Rop1At (data not shown), demonstrating that RopGAP1 or GDI co-overexpression had no effects on the expression of Rop1At or DN-rop1At.
Latrunculin B Reversed the Abnormal Actin Patterns and Depolarized Growth Induced by Rop1At Overexpression
We next sought to test whether the Rop1At-induced transverse actin band is due to stabilization of the actin collar and increased actin assembly at the subapical region by ectopically localized Rop using the actin-depolymerizing drug Latrunculin B. Latrunculin B disassembles F-actin specifically through sequestering G-actin (Gibbon et al. 1999). As shown in maize pollen (Gibbon et al. 1999), 5 nM Latrunculin B significantly inhibited the elongation of tobacco pollen tubes, but did not affect axial actin cables (Table and Fig. 5 D). 30 min after Latrunculin B treatments, SAB and actin collar disappeared, whereas axial actin cables protruded into the tip. The disruption of tip actin was associated with the inhibition of pollen tube elongation; the average length of treated pollen tubes is ∼202 μm, much shorter than untreated tubes (352 μm). Latrunculin B did not significantly affect pollen tube morphology, as both treated and untreated tubes have a tube diameter ∼10 μm.
Figure 5.


Latrunculin B mimics the effect of Rop1At-negative regulators in suppressing Rop1At overexpression-induced actin reorganization. Tobacco pollen was bombarded and treated with Latrunculin B and analyzed using confocal microscopy as described in Materials and Methods. As controls, tubes expressing GFP-mTalin alone (A) were treated with 5 nM Latrunculin B (D). Pollen tubes cooverexpressing Rop1At (B) were treated with either 5 or 30 nM Latrunculin B. The former removed the transverse actin band as well as recovered the apical SAB (E). This treatment also restored the dynamics of tip-localized F-actin as determined by fluorescence intensity of GFP in the extreme apex (H). Treatment with 30 nM Latrunculin B severely disrupted axial actin cables (F). Treatment of tubes expressing DN-rop1At (C) with 5 nM Latrunculin B similarly removed the transverse band of actin hoops. DN-rop1At tubes treated with 5 nM Latrunculin B (G) display a similar actin-staining pattern as tubes overexpressing RopGAP1 (see Fig. 4 F) and those cooverexpressing DN-rop1At and GDI (see Fig. 4 E). The effects of Latrunculin B treatment on the suppression of transverse actin bands were also quantitated. As shown in I, untreated Rop1At or DN-rop1At–overexpressing tubes (black bars) had a dramatically higher percentages of tubes with a transverse actin band (57 and 49%, respectively) than those of tubes treated with 5 nM Latrunculin B (white bars) (17 and 6%, respectively). Creatine kinase (CK), control tubes expressing GFP-mTalin alone. Bar, 10 μm.
Interestingly, 20–30 min after Latrunculin B treatment, the transverse actin band disappeared from the majority of tubes overexpressing DN-rop1At (only 5.6% of tubes with transverse actin band compared with 54.5% in untreated tubes) or Rop1At (17.2% compared with 56.7% in untreated tubes) (Fig. 5E, Fig. G, and Fig. H). Moreover, treatments of Rop1At-overexpressing tubes with 5 nM Latrunculin B restored the dynamic apical F-actin observed in wild-type tubes (Fig. 5 E). Time–sequence analyses of tip-localized F-actin showed that the amount of tip-localized actin fluctuated in a comparable manner between wild-type tubes and Latrunculin B–treated Rop1At-overexpressing tubes (Fig. 5 H). In contrast, the amount of tip-localized F-actin exhibited little fluctuation in untreated Rop1At-overexpressing tubes (Fig. 5 H). Finally, Latrunculin B–treated DN-rop1At tubes showed F-actin localization patterns similar to those of tubes overexpressing RopGAP1 and those cooverexpressing DN-rop1At and GDI (compare Fig. 5 G with Fig. 4E and Fig. F). These results strongly indicate that Rop GTPases play a crucial role in controlling the dynamics of tip F-actin in pollen tubes.
We then determined whether the rescue of actin organization in Rop1At-overexpressing cells by Latrunculin B leads to the recovery of normal tip growth. 3 h after Latrunculin B treatment, the length and the maximal width of pollen tubes were measured. As shown in Table and Fig. 6, Latrunculin B treatments suppressed tube expansion or tip swelling induced by either Rop1At or DN-rop1At overexpression. Latrunculin B treatment reduced the width of Rop1At-overexpressing pollen tubes from 19 to 12 μm. Similarly, this treatment also reduced the width of DN-rop1At tubes from 14 to 12 μm. These results suggest that Rop controls the polarity of pollen tube growth via dynamic tip-localized F-actin.
Figure 6.
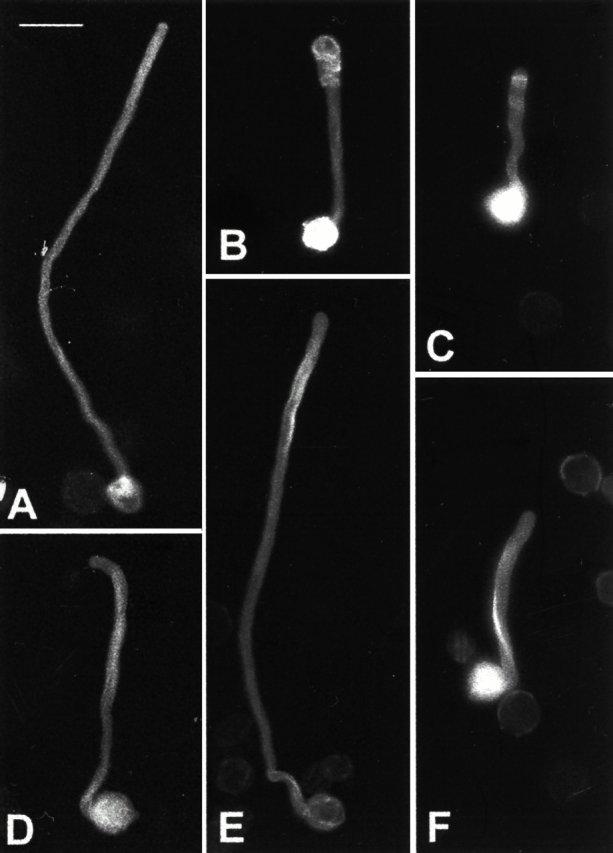
Effects of Latrunculin B on morphological phenotypes of pollen tubes overexpressing Rop1At or DN-rop1At. Pollen was transformed with different constructs and visualized using epifluorescence microscopy and a cooled CCD camera as described in Materials and Methods. A–C show typical morphology of untreated pollen tubes expressing GFP-mTalin alone (A), GFP-mTalin and Rop1At (B), and GFP-mTalin and DN-rop1 tube (C). D–F show typical morphology of pollen tubes treated with 5 nM Latrunculin B. Treated tubes expressing GFP-mTalin alone (D), GFP-mTalin and Rop1At (E), and GFP-mTalin and DN-rop1 tube (F). Bar, 50 μm.
Latrunculin B–induced reduction in tip expansion in tubes overexpressing Rop1At or DN-rop1At was correlated with an increase in tube elongation. At 4.5 h after bombardment, Rop1At tubes elongated significantly faster, to 438 μm when treated with 5 nM Latrunculin B compared with 339 μm in untreated tubes (Table and Fig. 6). Latrunculin B–treated DN-rop1At tubes (128 μm) were also longer than nontreated tubes (96 μm) on the average, though the difference is not statistically significant. Interestingly, Latrunculin B–treated Rop1At-overexpressing tubes (438 μm) elongated much faster than the control, wild-type tubes (352 μm). This surprising result provides strong evidence for the role of actin dynamics in the regulation of pollen tube growth.
To further assess the role of actin dynamics in pollen tube growth, we tested the effect of various concentrations of Latrunculin B on tubes overexpressing Rop1At. As shown in Table , treatments with Latrunculin B as low as 1 nM somewhat reduced the width of tubes and promoted tube elongation. However, 5 nM Latrunculin B was optimal for promoting elongation, as well as inhibiting expansion. High concentrations of Latrunculin B (e.g., 30 nM) inhibited elongation more severely than expansion (shorter tubes with greater width to length ratio than untreated Rop1At tubes), presumably due to the disruption of actin cables (Fig. 5 F; Gibbon et al. 1999). Taken together, these results clearly indicate that the dynamics of the tip-localized actin play a critical role in the control of pollen tube growth.
Table 2.
Effect of Different Concentrations of Latrunculin B on Shape and Growth of Pollen Tubes Overexpressed Rop1
| Latrunculin B | Length | Width | Width/Length |
|---|---|---|---|
| nM | mean ± SD | mean ± SD | mean ± SD |
| 0 | 278.2 ± 42.9 | 19.2 ± 1.1 | 0.084 ± 0.02 |
| 1 | 297.7 ± 68.3 | 16.8 ± 1.3 | 0.077 ± 0.02 |
| 5 | 412.1 ± 49.5 | 12.4 ± 0.3 | 0.041 ± 0.01 |
| 30 | 110.5 ± 9.1 | 15.1 ± 2.7 | 0.159 ± 0.02 |
Data came from three independent experiments.
The Abundance of Tip-localized SABs Oscillates and Increases before Growth
To understand how the dynamics of the tip-localized actin are involved in the control of tip growth, we determined the relationship between actin dynamics and growth oscillation. Growth oscillation has been shown to occur in pollen tubes from various plant species, including tobacco (Malhó et al. 1994; Pierson et al. 1996; Holdaway-Clarke et al. 1997; Messerli and Robinson 1997). To quantitate apical F-actin, we measured average GFP fluorescence intensity in an extreme apical area of midplane optical sections from laser scanning of pollen tubes expressing GFP-mTalin. Tip GFP fluorescence intensity for a time serial (15-s intervals) obtained from a given tube was plotted against elongation rates. Analyses of eight pollen tubes showed a similar correlation between tip GFP intensity and elongation rates. Fig. 7 shows results from a representative tube. The peak of GFP fluorescence generally preceded the peak of elongation rate by ∼15–30 s, although a more refined time-lapse imaging may allow deciphering of a more accurate phasic relationship. However, the amplitude of a GFP peak correlated with that of the following growth peak. Interestingly, the apex of a GFP peak generally coincided with the base of a growth peak. These results provide evidence that tip F-actin plays a role in events that precede tip growth.
Figure 7.

Tip-localized F-actin oscillates with pollen tube growth. A time series of confocal sections through the midplane of a pollen tube expressing GFP-mTalin was taken and GFP intensity in the extreme apex (♦) and elongation rates (□) of this tube were determined using Metamorph v4.5 software as described in Materials and Methods. Y-axis values are arbitrary units of fluorescence intensity; x-axis indicates times after the first laser section.
Discussion
In this study, we have used chemical genetic analysis in conjunction with live imaging of the actin cytoskeleton to demonstrate that the dynamics of an actin structure localized to the site of growth plays an essential role in the development of cell polarity and polar growth in pollen tubes. Importantly, the dynamics of this actin is dependent on the signaling of a plant-specific Rho-type GTPase, Rop. This study provides the first direct evidence to link F-actin to polarity development through a Rho GTPase in plant cells. This extends the common theme of Rho GTPase–mediated cell polarity control to the plant system (Chant and Stowers 1995; Drubin and Nelson 1996; Mata and Nurse 1998; Bahler and Peter 2000; Fowler 2000).
The Apex of Pollen Tubes Contains a Highly Dynamic Cortical Actin Structure
The actin cytoskeleton has been implicated in many aspects of plant cell growth, division, and development (Hable et al. 1998; Kropf et al. 1998; Li and Yang 2000; Schmit 2000; Staiger et al. 2000). Actin functions in plants involve the dynamics and the reorganization of specific arrays of F-actin in response to external stimuli and internal developmental cues (Emmons and de Ruijter 2000; Geitmann et al. 2000; Li and Yang 2000; Schmit 2000; Staiger 2000). However, we have little knowledge of mechanisms by which actin dynamics and reorganization are regulated in plants, except for the existence of several classes of actin-binding proteins and their roles in actin organization (Huang et al. 1996; Gibbon et al. 1998; Hussey et al. 1998; Smertenko et al. 1998; Staiger 2000).
One of the major challenges in studying the actin cytoskeleton in the plant cell has been difficulties in faithfully visualizing F-actin and its dynamics. Chemical fixation methods frequently fail to detect dynamic and fine actin filaments and may cause dramatic structural alteration (Doris and Steer 1996; Miller et al. 1996), whereas injected labeled phalloidin may induce actin bundling (Cooper 1987). Freezing methods at the transmission electron microscopy level and immunocytochemistry appear to produce a better view of F-actin in pollen tubes with few, and only fine, actin filaments detected in the tip (Lancelle et al. 1987; Miller et al. 1996). Nonetheless, a major advance was the recent development of a GFP-tagged actin-binding domain of the mTalin, which has revealed novel actin structures and has been successfully used to investigate actin dynamics in live plant cells (Kost et al. 1998). However, results from these studies revealed no F-actin in the pollen tube tip.
Our current study, using an enhanced GFP mutant (S65C) to tag mTalin, has uncovered a dynamic population of tip-localized SABs in tobacco pollen tubes. We believe that the use of this improved GFP mutant was critical, because it produces much more intense fluorescence compared with the GFP (S65T) mutant used previously (Reichel et al. 1996; Kost et al. 1998). Consequently, we were able to visualize tip F-actin, which is less abundant but more dynamic than actin cables in the shank of the tube. More importantly, the high sensitivity of the improved GFP made it possible to conduct time–sequence analyses of live pollen tubes, because of reduced laser damage to the cell and reduced GFP bleaching. We found that SABs rapidly appear in and disappear from the cortex of the extreme apex of pollen tubes, the expected site of exocytosis. This novel actin structure is distinct from the dense actin network reported for chemically fixed pollen tubes (Derksen et al. 1995), which was obviously an artifact (Doris and Steer 1996; Miller et al. 1996). However, the spare fine actin filaments that were occasionally observed at the tip of pollen tubes using electron microscopy, rapid freeze, and substitution techniques (Lancelle et al. 1987; Miller et al. 1996) could belong to the dynamic SAB. SAB is probably different from the dense tip F-actin that forms in response to S protein treatments in poppy pollen tubes (Geitmann et al. 2000).
In addition to SAB, GFP-mTalin also revealed another F-actin structure localized just behind the extreme apex, similar to an actin ring or actin collar described previously (Kost et al. 1998; Gibbon et al. 1999), although this structure was only present in less than a quarter of tubes examined. Interestingly, this form of F-actin is also very dynamic and its dynamics are associated with those of SAB. In most cases, the actin collar decreases when SAB increases (Fig. 1), suggesting that the apical SAB and subapical actin collar may be two dynamic components of the same actin population. This is further supported by our results showing that alteration of Rop activity and Latrunculin B treatments changed both SAB and actin collar simultaneously.
Rop Controls the Formation and the Dynamics of F-actin at the Tip of Pollen Tubes
Our current results indicate that the formation and the dynamics of tip F-actin are controlled by Rop GTPase activity localized to the apex of the pollen tube PM. Several lines of evidence strongly support this conclusion. First, RopGAP1 overexpression in tobacco pollen tubes drastically reduced SAB and completely removed the actin collar. Second, Rop1At overexpression, which led to ectopic accumulation of Rop proteins to the apical region of PM (Li et al. 1999), induced transverse actin band formation in the subapical region and conversion of SAB into an actin network at the apex. Third, cooverexpression of RopGAP1 reversed the effect on these actin structures caused by Rop1At overexpression. Fourth, DN-rop1At overexpression, which not only inhibits the activation of native Rop GTPases but also causes their ectopic localization to the subapical region of the tube PM, disrupted the apical SAB, but induced the subapical transverse actin band. Fifth, GDI, which may inhibit Rop activation by recycling it from PM, also suppressed the transverse actin band induced by Rop1At or DN-rop1At overexpression. Finally, and most importantly, Latrunculin B treatment of tubes overexpressing Rop1At removed the transverse actin band and recovered the normal dynamics of SABs, mimicking the effect of RopGAP1 or GDI overexpression on these tubes. In all of the aforementioned experiments, we did not observe detectable changes in the pattern and thickness of axial actin cables, suggesting that Rop1At is not directly involved in the regulation of axial actin cables. This is in contrast to the effect of DN-Atrac-2/rop5At and CA-Atrac2/rop5At (Kost et al. 1999). The reason for this discrepancy remains to be determined, although several possibilities, including possible differences in growth conditions and possible distinct functions for Rop1At versus Rop5At, can explain this discrepancy.
Our results suggest that Rop controls the dynamics of tip actin by promoting actin assembly, as do Rop-related Cdc42 and Rac GTPases in mammalian cells. First, Rop1At overexpression induced the formation of long actin filaments in the apex of pollen tubes, suggesting that increased actin polymerization allows SAB to become long filaments. Second, treatments of Rop1At-overexpressing tubes with the G-actin sequestering drug Latrunculin B completely recovered normal actin dynamics by suppressing the formation of the apical long actin filaments and subapical transverse actin band. Finally, SAB was disrupted by either deactivation of Rop or by Latrunculin B treatments in wild-type tubes. A role for Rop in promoting actin polymerization is also supported by a recent study suggesting that a phosphatidylinositol 4,5-bisphosphate (PIP) kinase and PIP2 may be a Rop effector in tobacco pollen (Kost et al. 1999). PIP2 has been shown to promote actin polymerization in animal cells (Moreau and Way 1998; Chen et al. 2000).
Both transient deactivation of Rop and actin disassembly at the tip could contribute to the dynamics of the tip actin. Potential factors that control actin disassembly at the tip include profilin, actin-depolymerizing factor, gelsolin, and calcium (Smertenko et al. 1998; Gibbon and Staiger 2000; Kovar and Staiger 2000; Vidali and Hepler 2000). In this regard, it is particularly interesting to note that profilins sequester pollen G-actin in a calcium-dependent manner and are believed to contribute to actin dynamics in pollen tubes (Kovar et al. 2000). A tip-focused gradient of cytosolic calcium oscillates in phase with pollen tube growth (Pierson et al. 1996; Holdaway-Clarke et al. 1997; Messerli and Robinson 1997). The peak of tip calcium is estimated to reach as high as 3–10 μM, falling within the range of calcium concentration (5 μM) optimal for profilin-mediated G-actin sequestration (Kovar and Staiger 2000). Thus, the potential oscillation of profilin-dependent G-actin sequestration could well contribute to actin dynamics at the tip. This hypothesis is consistent with our observation that tip F-actin oscillates in the opposite phase with growth (and presumably the tip-focused calcium gradient) in tobacco pollen tubes (Fig. 7 and Fig. 8).
Figure 8.
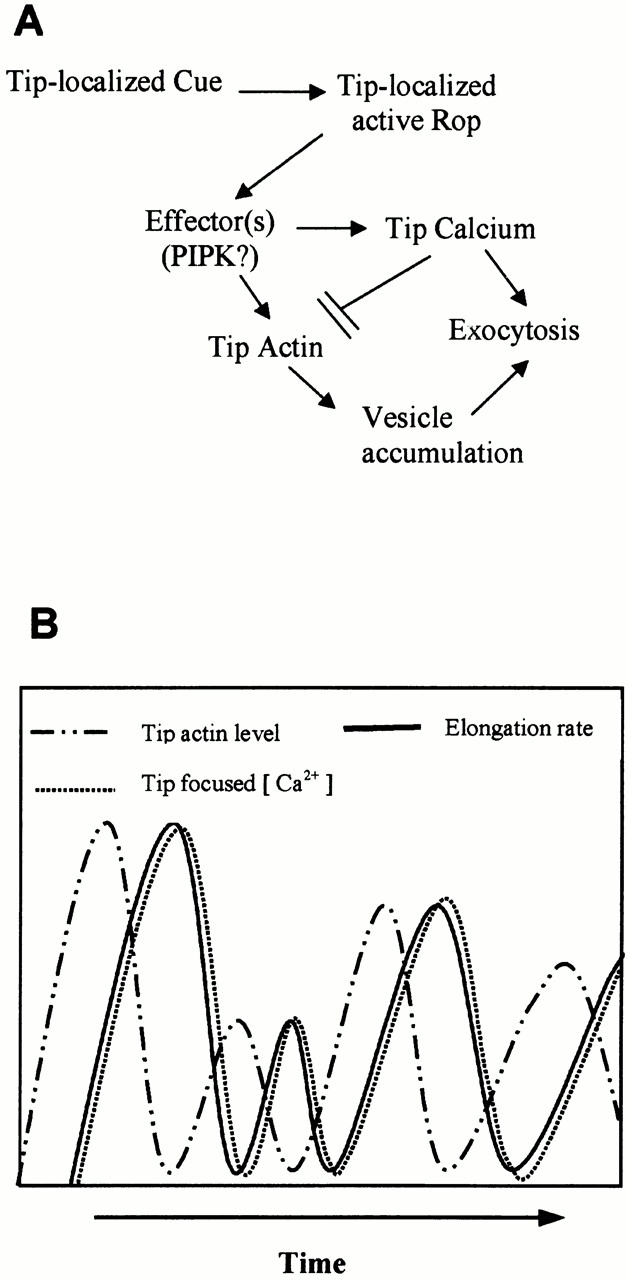
A speculative model for functional interrelationship between Rop, tip-focused calcium gradient, and tip F-actin in the control of polar growth in pollen tubes. Our previous and current data together indicate that Rop acts to turn on tip growth through both the tip-focused calcium (Li et al. 1999) and the tip-localized F-actin (this paper). These data are also consistent with the hypothesis that actin and calcium act independently downstream of Rop to promote polar exocytosis possibly by targeting vesicles to the extreme apex and by regulating vesicle fusion, respectively (A). This hypothesis can explain the temporal relationship between the dynamics of tip actin, calcium, and growth, as schematically illustrated in B. Growth oscillates closely in phase with the tip-focused calcium gradient (Holdaway-Clarke et al. 1997; Messerli and Robinson 1997), whereas this study indicates that the tip-localized actin oscillates in the opposite phase with growth and calcium. Thus, the accumulation of vesicles at the extreme apex would be synchronous with that of calcium, both of which are necessary for the growth burst. High calcium at the tip could also cause tip actin to disassemble (see above) and/or negatively feedback-regulating Rop activity (Li et al. 1999). Utilization of calcium by exocytosis and/or Rop-mediated feedback regulation of calcium would allow Rop to be reactivated and/or tip F-actin to be reassembled.
The calcium-dependent actin disassembly at the tip may also explain the formation of the actin collar in normal tubes and the transverse actin band in Rop1At-overexpressing tubes. Although actin disassembly at the tip is activated by high calcium, polymerization just behind the tip may continue, contributing to the formation of the actin collar. Moreover, as discussed below, Rop1At overexpression-induced transverse actin band probably results from the stabilization of the collar actin by ectopically localized Rop in the subapical region, where calcium-dependent actin disassembly is absent or minimal.
The Rop-dependent Tip F-actin Controls Cell Polarity Development in Pollen Tubes
Our previous studies indicate that localized Rop activity specifies the site for tip growth (Li et al. 1999; Wu, G., Y. Fu, and Z. Yang, unpublished data). Rop1At overexpression causes increased Rop localization and activation in the subapical region of the tube PM, leading to tube expansion or tip swelling. Time–sequence analysis showed that Rop1At-induced formation of the transverse actin band is initiated at the tip before (or at the onset of) tip swelling, suggesting that ectopically localized Rop causes depolarized growth at least in part through the subapical transverse actin band. This hypothesis is further supported by our results showing that the suppression of Rop1At-induced actin changes by RopGAP1 or GDI1 is associated with recovery of normal tip growth. Direct evidence for this hypothesis came from the experiments showing that 5 nM Latrunculin B suppressed Rop1At-induced transverse actin band formation, leading to the recovery of SAB and normal polar growth in Rop1At-overexpressing tubes. Hence, we conclude that the polarization of SAB to the tip is required for Rop-dependent cell polarity development in pollen tubes. In this regard, SAB appears analogous to the polarized cortical actin patches that establish cell polarity in algal zygotes and in yeast (Alessa and Kropf 1999; Kropf et al. 1999; Bahler and Peter 2000; Fowler 2000).
The Dynamics of Apical SABs Couple Cell Polarity Control with Growth Control
Our study suggests that SAB is also critical for pollen tube growth. The actin cable–independent effect of Latrunculin B treatments suggests that a population of F-actin is required for pollen tube growth but not cytoplasmic streaming (Gibbon et al. 1999). Our data indicate that this actin population is SAB. First, Latrunculin B treatments disrupted SAB and inhibited tube growth, as did overexpression of GDI, GAP, or DN-rop1At. Second, the localization of SAB to the tip oscillates with growth and precedes growth. Third, Latrunculin B treatments of Rop1At-overexpressing tubes recovered normal tip growth associated with normal oscillation of SAB. Interestingly, these tubes grew even faster than wild-type tubes, suggesting that the greater dynamics of tip actin resulting from Rop activation of actin assembly and Latrunculin B promotion of actin disassembly may allow a more efficient polar growth.
The requirement of Rop-mediated SAB for both growth and polarity controls is consistent with our previous finding that Rop couples growth and polarity control during pollen tube tip growth. Together, our studies provide strong evidence for the model of Rop-mediated tip growth: a localized tip growth cue activates Rop at the extreme apex of the tube and activated Rop then controls polarized organization of SAB to the site of growth. It remains to be determined how polarized SAB regulates tip growth. An attractive model is that SAB targets vesicles to the site of growth. A high density of vesicles accumulates at the extreme apex of the tube (Lancelle and Hepler 1992). A driving force must be required for the vesicle accumulation and SAB is the most likely candidate. However, it is possible that SABs stabilize growth sites, regulate tip-localized putative calcium channels, or regulate endocytosis.
Regardless of the mode of action for SAB, it is intriguing that the function of Rop and SAB in pollen tubes is very analogous to that of Cdc42 and cortical actin patches in the tip-growing fission yeast: Cdc42 and actin patches are involved in both cell polarity establishment and growth control (Miller and Johnson 1994; Bahler and Peter 2000). In contrast, Cdc42 is only required for the polarization of actin patches to the site of growth, but not for their formation in budding yeast (Johnson and Pringle 1990; Drubin and Nelson 1996). The formation of actin patches appears to be regulated by other types of Rho GTPases such as Rho1p in budding yeast (Bahler and Peter 2000). Thus, the coupling of polarity establishment with growth control via Rho regulation of polar actin formation could be a common mechanism for polar growth that is unique to strictly tip-growing cells, such as fission yeast, neuronal cells, and pollen tubes requiring continuous unidirectional propagation of growth sites.
Potential Cross-Talk between Actin and Calcium via Rop in Pollen Tube Growth
Rop also regulates the formation of the tip-focused calcium gradient (Li et al. 1999). Like SAB, the calcium gradient also couples the control of growth sites with growth (Zheng and Yang 2000a). What is the functional relationship between Rop, tip actin, and calcium gradient? Several possible models could explain this relationship. The three components could operate in a linear pathway. Rop could first regulate calcium accumulation at the tip, which then might promote SAB assembly. However, current evidence argues against this model, because calcium accumulation at the tip most likely leads to actin disassembly rather than assembly as discussed above. Alternatively, SAB could regulate the formation of calcium gradients. This model would predict that the dynamic tip actin is sufficient for Rop-dependent tip growth, because ectopic localization of Rop to the base of the apical dome is sufficient to induce depolarized growth (Li et al. 1999). However, accumulation of intense actin bands does not lead to complete loss of cell polarity (Fig. 3 and Fig. 4), suggesting that additional factors are required for Rop-dependent growth.
We favor the model that Rop regulates actin and calcium independently (Fig. 8 A). Tip-localized Rop could activate two distinct Rop effectors that are involved in the control of calcium gradient formation and tip actin dynamics, respectively. Alternatively, Rop could activate a single effector, and the calcium and actin pathways bifurcate downstream of this effector. A potential Rop effector of this sort is a phosphotidylinositol phosphate kinase and its product, PIP2 (Kost et al. 1999). On one hand, PIP2 could regulate SAB formation, which may allow vesicles to accumulate at the tip. As mentioned above, PIP2 has been shown to promote actin assembly in animal cells. PIP2 is known to bind profilin and thus inhibit the G-actin sequestering capacity of profilin (Kovar et al. 2000). Thus, increase in tip PIP2 levels may immediately promote actin assembly. On the other hand, PIP2 could be used to generate inositol 1,4,5-trisphosphate, which could then activate calcium release from the vesicles or the ER (Zheng and Yang 2000a). These signaling events ultimately could contribute to the formation of the tip-focused calcium gradient, which would be expected to be slower than PIP2-mediated actin assembly and thus could account for the phase shift between actin assembly and calcium accumulation (Fig. 8 B). The tip calcium may then simultaneously promote exocytosis and actin disassembly, leading to tube elongation and actin dynamics, respectively. Though speculative, this model can explain the oscillation phenomena and the phase shift between tip SAB and calcium during pollen tube growth. Identification of Rop effectors, combined with simultaneous imaging of actin and calcium in live cells, would be useful for testing this model.
Supplemental Material
Acknowledgments
We would like to thank Elizabeth Lord and Eugene Nothnagel for critical reading and comments on this manuscript, and Veronica Franklin-Tong, Chris Staiger, Ming Yuan, and members of Z. Yang's laboratory for helpful discussion.
This work is supported by a grant to Z. Yang from the National Science Foundation (MCB-0096026).
Footnotes
The online version of this article contains supplemental material.
Abbreviations used in this paper: CA, constitutively active; DN, dominant negative; GAP, GTPase-activating protein; GDI, guanine nucleotide dissociation inhibitor; GFP, green fluorescent protein; GM, germination medium; mTalin, mouse talin; PIP, phosphatidylinositol 4,5-bisphosphate; PM, plasma membrane; SAB, short actin bundle.
References
- Alessa L., Kropf D.L. F-actin marks the rhizoid pole in living Pelvetia compressa zygotes. Development. 1999;126:201–209. doi: 10.1242/dev.126.1.201. [DOI] [PubMed] [Google Scholar]
- Bahler J., Peter M. Cell polarity in yeast. In: Drubin D.G., editor. Cell Polarity. Oxford University Press; Oxford: 2000. pp. 21–77. [Google Scholar]
- Bate N., Spurr C., Foster G.D., Twell D. Maturation-specific translational enhancement mediated by the 5′-UTR of a late pollen transcript. Plant J. 1996;10:613–623. doi: 10.1046/j.1365-313x.1996.10040613.x. [DOI] [PubMed] [Google Scholar]
- Bischoff F., Vahlkamp L., Molendijk A., Palme K. Localization of AtROP4 and AtROP6 and interaction with the guanine nucleotide dissociation inhibitor AtRhoGDI1 from Arabidopsis . Plant Mol. Biol. 2000;42:515–530. doi: 10.1023/a:1006341210147. [DOI] [PubMed] [Google Scholar]
- Chant J., Stowers L. GTPase cascades choreographing cellular behaviormovement, morphogenesis, and more. Cell. 1995;81:1–4. doi: 10.1016/0092-8674(95)90363-1. [DOI] [PubMed] [Google Scholar]
- Chen F., Ma L., Parrini M.C., Mao X., Lopez M., Wu C., Marks P.W., Davidson L., Kwiatkowski D.J., Kirchhausen T. Cdc42 is required for PIP(2)-induced actin polymerization and early development but not for cell viability. Curr. Biol. 2000;10:758–765. doi: 10.1016/s0960-9822(00)00571-6. [DOI] [PubMed] [Google Scholar]
- Cooper J.A. Effects of cytochalasin and phalloidin on actin. J. Cell Biol. 1987;105:1473–1478. doi: 10.1083/jcb.105.4.1473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Derksen J., Rutten T., Van Amstel T., de Win A., Doris F., Steer M. Regulation of pollen tube growth. Acta Bot. Neerl. 1995;44:93–119. [Google Scholar]
- Doris F.P., Steer M.W. Effects of fixatives and permeabilisation buffers on pollen tubesimplications for localisation of actin microfilaments using phalloidin staining. Protoplasma. 1996;195:25–36. [Google Scholar]
- Drubin D.G., Nelson W.J. Origins of cell polarity. Cell. 1996;84:335–344. doi: 10.1016/s0092-8674(00)81278-7. [DOI] [PubMed] [Google Scholar]
- Emmons A.M., de Ruijter N. Actina target of signal transduction in root hairs. In: Staiger C.J., Baluska F., Volkmann D., Barlow P.W., editors. ActinA Dynamic Framework for Multiple Plant Cell Functions. Kluwer Academic Publishers; Dordrecht, The Netherlands: 2000. pp. 373–390. [Google Scholar]
- Fowler J.E. Cell polarity in algae and vascular plants. In: Drubin D.G., editor. Cell Polarity. Oxford University Press; Oxford: 2000. pp. 141–180. [Google Scholar]
- Franklin-Tong V.E. Signaling and the modulation of pollen tube growth. Plant Cell. 1999;11:727–738. doi: 10.1105/tpc.11.4.727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geitmann A., Snowman B.N., Emons A.M.C., Franklin-Tong V.E. Alterations in the actin cytoskeleton of pollen tubes are induced by the self-incompatibility reaction in Papaver rhoeas . Plant Cell. 2000;12:1239–1252. doi: 10.1105/tpc.12.7.1239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibbon B.C., Staiger C.J. Profilin. In: Staiger C.J., Baluska F., Volkmann D., Barlow P.W., editors. ActinA Dynamic Framework for Multiple Plant Cell Functions. Kluwer Academic Publishers; Dordrecht, The Netherlands: 2000. pp. 45–66. [Google Scholar]
- Gibbon B.C., Zonia L.E., Kovar D.R., Hussey P.J., Staiger C.J. Pollen profilin function depends on interaction with profilin rich motifs. Plant Cell. 1998;10:981–993. doi: 10.1105/tpc.10.6.981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibbon B.C., Kovar D.R., Staiger C.J. Latrunculin B has different effects on pollen germination and tube growth. Plant Cell. 1999;11:2349–2364. doi: 10.1105/tpc.11.12.2349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hable W.E., Bisgrove S.R., Kropf D.L. To shape a plant-The cytoskeleton in plant morphogenesis. Plant Cell. 1998;10:1772–1774. doi: 10.1105/tpc.10.11.1772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall A. Rho GTPases and the actin cytoskeleton. Science. 1998;279:509–514. doi: 10.1126/science.279.5350.509. [DOI] [PubMed] [Google Scholar]
- Holdaway-Clarke T.L., Feió J.A., Hackett G.R., Kunkel J.G., Hepler P.K. Pollen tube growth and the intracellular cytosolic calcium gradient oscillate in phase while extracellular calcium influx is delayed. Plant Cell. 1997;9:1999–2010. doi: 10.1105/tpc.9.11.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang S., McDowell J.M., Weise M.J., Meagher R.B. The Arabidopsis profilin gene family. Plant Physiol. 1996;111:115–126. doi: 10.1104/pp.111.1.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hussey P.J., Yuan M., Calder G., Khan S., Lloyd C.W. Microinjection of pollen-specific actin-depolymerizing factor, ZmADF1, reorientates F-actin strands in Tradescantia stamen hair cells. Plant J. 1998;14:353–357. [Google Scholar]
- Johnson D.I., Pringle J.R. Molecular characterization of CDC42, a Saccharomyces cerevisiae gene involved in the development of cell polarity. J. Cell Biol. 1990;111:143–152. doi: 10.1083/jcb.111.1.143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kost B., Spielhofer P., Chua N.H. A GFP-mouse talin fusion protein labels plant actin filaments in vivo and visualizes the actin cytoskeleton in growing pollen tubes. Plant J. 1998;16:393–401. doi: 10.1046/j.1365-313x.1998.00304.x. [DOI] [PubMed] [Google Scholar]
- Kost B., Lemichez E., Spielhofer P., Hong Y., Tolias K., Carpenter C., Chua N.-H. Rac homologues and compartmentalized phosphatidylinositol 4,5-bisphosphate act in a common pathway to regulate polar pollen tube growth. J. Cell Biol. 1999;145:317–330. doi: 10.1083/jcb.145.2.317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kovar D.R., Staiger C.J. Actin depolymerizing factor. In: Staiger C.J., Baluska F., Volkmann D., Barlow P.W., editors. ActinA Dynamic Framework for Multiple Plant Cell Functions. Kluwer Academic Publishers; Dordrecht, The Netherlands: 2000. pp. 67–86. [Google Scholar]
- Kovar D.R., Drobak B.K., Staiger C.J. Maize profilin isoforms are functionally distinct. Plant Cell. 2000;12:583–598. doi: 10.1105/tpc.12.4.583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kropf D.L., Bisgrove S.R., Hable W.E. Cytoskeletal control of polar growth in plant cells. Curr. Opin. Cell Biol. 1998;10:117–122. doi: 10.1016/s0955-0674(98)80094-x. [DOI] [PubMed] [Google Scholar]
- Kropf D.L., Bisgrove S.R., Hable W.E. Establishing a growth axis in fucoid algae. Trends Plant Sci. 1999;4:490–494. doi: 10.1016/s1360-1385(99)01509-5. [DOI] [PubMed] [Google Scholar]
- Lancelle S.A., Hepler P.K. Ultrastructure of freeze-substituted pollen tubes of Lilium longiflorum . Protoplasma. 1992;167:215–230. [Google Scholar]
- Lancelle S.A., Cresti M., Hepler P.K. Ultrastructure of the cytoskeleton in freeze-substituted pollen tubes of Nicotiana altata . Protoplasma. 1987;140:141–150. [Google Scholar]
- Li H., Yang Z. Rho GTPase and the actin cytoskeleton. In: Staiger C.J., Baluska F., Volkmann D., Barlow P., editors. ActinA Dynamic Framework for Multiple Plant Cell Functions. Kluwer Academic Press; Dordrecht, The Netherlands: 2000. pp. 301–321. [Google Scholar]
- Li H., Wu G., Ware D., Davis K.R., Yang Z. Arabidopsis Rho-related GTPasesdifferential gene expression in pollen and polar localization in fission yeast. Plant Physiol. 1998;118:407–417. doi: 10.1104/pp.118.2.407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H., Lin Y., Heath R.M., Zhu M.X., Yang Z. Control of pollen tube tip growth by a Rop GTPase-dependent pathway that leads to the tip-localized calcium influx. Plant Cell. 1999;11:1731–1742. doi: 10.1105/tpc.11.9.1731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Y., Yang Z. Inhibition of pollen tube elongation by microinjected anti-Rop1Ps antibodies suggests a crucial role for Rho-type GTPases in the control of tip growth. Plant Cell. 1997;9:1647–1659. doi: 10.1105/tpc.9.9.1647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Y., Wang Y., Zhu J., Yang Z. Localization of a rho GTPase implies a role in tip growth and movement of the generative cell in pollen tubes. Plant Cell. 1996;8:293–303. doi: 10.1105/tpc.8.2.293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malhó R., Read N.D., Pais M.S., Trewavas A.J. Role of cytosolic free calcium in the reorientation of pollen tube growth. Plant J. 1994;5:331–341. [Google Scholar]
- Mata J., Nurse P. Discovering the poles in yeast. Trends Cell Biol. 1998;8:163–167. doi: 10.1016/s0962-8924(98)01224-0. [DOI] [PubMed] [Google Scholar]
- McCann R.O., Craig S.W. The I/LWEQ modulea conserved sequence that signifies F-actin binding in functionally diverse proteins from yeast to mammals. Proc. Natl. Acad. Sci. USA. 1997;94:5679–5684. doi: 10.1073/pnas.94.11.5679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Messerli M., Robinson K.R. Tip localized Ca2+ pulses are coincident with peak pulsatile growth rates in pollen tubes of Lilium longiflorum . J. Cell Sci. 1997;110:1269–1278. doi: 10.1242/jcs.110.11.1269. [DOI] [PubMed] [Google Scholar]
- Miller D.D., Lancelle S.A., Hepler P.K. Actin microfilaments do not form a dense meshwork in Lilium longiflorum pollen tube tips. Protoplasma. 1996;195:123–132. [Google Scholar]
- Miller D.D., de Ruijter N.C.A., Bisseling T., Emons A.M.C. The role of actin in root hair morphogenesisstudies with lipochito-oligosaccharide as a growth stimulator and cytochalasin as an actin perturbing drug. Plant J. 1999;17:141–154. [Google Scholar]
- Miller P.J., Johnson D.I. Cdc42p GTPase is involved in controlling polarized cell growth in Schizosaccharomyces pombe . Mol. Cell. Biol. 1994;14:1075–1083. doi: 10.1128/mcb.14.2.1075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moreau V., Way M. Cdc42 is required for membrane dependent actin polymerization in vitro. FEBS Lett. 1998;427:353–356. doi: 10.1016/s0014-5793(98)00443-8. [DOI] [PubMed] [Google Scholar]
- Nobes C.D., Hall A. Rho, Rac, and Cdc42 GTPases regulate the assembly of multimolecular focal complexes associated with actin stress fiber, lamellipodia, and filopodia. Cell. 1995;81:53–62. doi: 10.1016/0092-8674(95)90370-4. [DOI] [PubMed] [Google Scholar]
- Pierson E.S., Miller D.D., Gallaham D.A., van Aken J., Hackett G., Hepler P.K. Tip-localized calcium entry fluctuates during pollen tube growth. Dev. Biol. 1996;174:160–173. doi: 10.1006/dbio.1996.0060. [DOI] [PubMed] [Google Scholar]
- Pruyne D., Bretscher A. Polarization of cell growth in yeast II. The role of the cortical actin cytoskeleton. J. Cell Sci. 2000;113:571–585. doi: 10.1242/jcs.113.4.571. [DOI] [PubMed] [Google Scholar]
- Reichel C., Mathur J., Eckes P., Langenkemper K., Koncz C., Schell J., Reiss B., Maas C. Enhanced green fluorescence by the expression of an Aequorea victoria green fluorescent protein mutant in mono- and dicotyledonous plant cells. Proc. Natl. Acad. Sci. USA. 1996;93:5888–5893. doi: 10.1073/pnas.93.12.5888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ren H., Gibbon B.C., Ashworth S.L., Sherman D.M., Yuan M., Staiger C. Actin purified from maize pollen functions in living plant cells. Plant Cell. 1997;9:1445–1457. doi: 10.1105/tpc.9.8.1445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ridley A.J., Hall A. The small GTP-binding protein rho regulates the assembly of focal adhesions and actin stress fibers in response to growth factors. Cell. 1992;70:389–399. doi: 10.1016/0092-8674(92)90163-7. [DOI] [PubMed] [Google Scholar]
- Ridley A.J., Paterson H.F., Johnston C.L., Diekmann D., Hall A. The small GTP-binding protein rac regulates growth factor-induced membrane ruffling. Cell. 1992;70:401–410. doi: 10.1016/0092-8674(92)90164-8. [DOI] [PubMed] [Google Scholar]
- Schmit A.-C. Actin during mitosis and cytokinesis. In: Staiger C.J., Baluska F., Volkmann D., Barlow P.W., editors. ActinA Dynamic Framework for Multiple Plant Cell Functions. Kluwer Academic Publishers; Dordrecht, The Netherlands: 2000. pp. 437–456. [Google Scholar]
- Smertenko A.P., Jiang C.J., Simmons N.J., Weeds A.G., Davies D.R., Hussey P.J. Ser6 in the maize actin-depolymerizing factor, ZmADF3, is phosphorylated by a calcium-stimulated protein kinase and is essential for the control of functional activity. Plant J. 1998;14:187–193. doi: 10.1046/j.1365-313x.1998.00107.x. [DOI] [PubMed] [Google Scholar]
- Staiger C.J. Signaling to the actin cytoskeleton. Ann. Plant Physiol. Plant Mol. Biol. 2000;51:257–288. doi: 10.1146/annurev.arplant.51.1.257. [DOI] [PubMed] [Google Scholar]
- Staiger C.J., Baluska F., Volkmann D., Barlow P.W. ActinA Dynamic Framework for Multiple Plant Cell Functions 2000. Kluwer Academic Publishers; Dordrecht, The Netherlands: pp. 663 pp [Google Scholar]
- Taylor L.P., Hepler P.K. Pollen germination and tube growth. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1997;48:461–491. doi: 10.1146/annurev.arplant.48.1.461. [DOI] [PubMed] [Google Scholar]
- Twell D., Yamaguchi J., Wing R.A., Ushiba J., McCormick S. Promoter analysis of genes that are coordinately expressed during pollen development reveals pollen-specific enhancer sequences and shared regulatory elements. Genes Dev. 1991;5:496–507. doi: 10.1101/gad.5.3.496. [DOI] [PubMed] [Google Scholar]
- Vidali L., Hepler P.K. Actin in pollen and pollen tubes. In: Staiger C.J., Baluska F., Volkmann D., Barlow P.W., editors. ActinA Dynamic Framework for Multiple Plant Cell Functions. Kluwer Academic Publishers; Dordrecht, The Netherlands: 2000. pp. 323–345. [Google Scholar]
- Wu G., Li H., Yang Z. Arabidopsis RopGAPs are a novel family of Rho GTPase-activating proteins that require the Cdc42/Rac-interactive binding motif for Rop-specific GTPase stimulation. Plant Physiol. 2000;124:1625–1636. doi: 10.1104/pp.124.4.1625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Z. Signaling tip growth in plants. Curr. Opin. Plant Biol. 1998;1:525–530. doi: 10.1016/s1369-5266(98)80046-0. [DOI] [PubMed] [Google Scholar]
- Zheng Z.-L., Yang Z. The Rop GTPase switch turns on polar growth in pollen Trends Plant Sci 5 2000. 298 303a [DOI] [PubMed] [Google Scholar]
- Zheng Z.-L., Yang Z. The Rop GTPasean emerging signaling switch in plants Plant Mol. Biol 44 2000. 1 9b [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.