

Abstract
T lymphocytes recirculate continually through the T cell areas of peripheral lymph nodes. During each passage, the T cells survey the surface of large dendritic cells (DCs), also known as interdigitating cells. However, these DCs have been difficult to release from the lymph node. By emphasizing the use of calcium-free media, as shown by Vremec et al. (Vremec, D., M. Zorbas, R. Scollay, D.J. Saunders, C.F. Ardavin, L. Wu, and K. Shortman. 1992. J. Exp. Med. 176:47–58.), we have been able to release and enrich DCs from the T cell areas. The DCs express the CD11c leukocyte integrin, the DEC-205 multilectin receptor for antigen presentation, the intracellular granule antigens which are recognized by monoclonal antibodies M342, 2A1, and MIDC-8, very high levels of MHC I and MHC II, and abundant accessory molecules such as CD40, CD54, and CD86. When examined with the Y-Ae monoclonal which recognizes complexes formed between I-Ab and a peptide derived from I-Eα, the T cell area DCs expressed the highest levels. The enriched DCs also stimulated a T-T hybridoma specific for this MHC II–peptide complex, and the hybridoma underwent apoptosis. Therefore DCs within the T cell areas can be isolated. Because they present very high levels of self peptides, these DCs should be considered in the regulation of self reactivity in the periphery.
Lymph nodes, the sites where primary immune responses are generated, contain separate regions through which B and T lymphocytes recirculate and gain access to discrete antigen-presenting cells (APCs). In B cell areas, follicular dendritic cells (DCs)1 retain antigens as immune complexes (1, 2). In T cell areas, marrow-derived DCs are thought to present processed antigens as peptides affixed to MHC products (3–5). T cell area DCs are not readily isolated from lymphoid tissue, particularly lymph nodes, thus hampering efforts to characterize antigen-presenting and costimulatory functions directly. We have succeeded in isolating this major depot of DCs from lymph node T cell areas, to directly assess expression of a specific MHC–peptide complex formed between a self I-Eα peptide and I-Ab. We find that lymph node DCs express the highest levels of MHC–peptide complexes, and efficiently induce the proliferation and then apoptosis of reactive T cells. Therefore DCs not only present foreign antigens, but are major reservoirs for self antigens as well.
Materials and Methods
Monoclonals.
Y-Ae antibody was provided by C. Janeway (Yale University, New Haven, CT) and used as a purified Ig. The Y-Ae mAb recognizes a complex formed between I-Ab and an I-Eα peptide (6). Other mAbs were purchased from PharMingen (San Diego, CA) or American Type Culture Collection (Rockville, MD).
Mice.
C57BL6 × DBA/2 and BALB/c × C57BL/6 F1 mice (6–10 wk, both sexes) express MHC–peptide complexes recognized by the Y-Ae mAb, while C57BL/6 (I-Ab only) and BALB/c (I-Eα peptide only) do not.
DC Enrichment.
Lymph nodes and spleens were dissociated with collagenase (collagenase D; Boehringer Mannheim Biochemicals, Indianapolis, IN) at 100 U/ml during the teasing apart of the tissue and then at 400 U/ml for 30 min at 37°C. The suspension was then immediately diluted 1:10 in Ca2+-free Hanks solution, centrifuged, and suspended in dense BSA prepared as described (7). A low density cell fraction, enriched in DCs, was harvested after centrifugation at 900 g for 30 min at 4°C. The cells were washed in Ca2+-free Hanks solution. DCs were also prepared from bone marrow precursors and epidermis as described (8, 9).
Two Color Immunolabeling of Tissue Sections.
10-μm cryostat sections were applied to multiwell slides (No. 111006; Carlson Scientific, Inc., Peotone, IL), air dried, and fixed in acetone for 10 min at room temperature. Biotinylated mAbs (CD8 for T cells; DEC-205, 2A1, MIDC-8, and CD11c for DCs; and Y-Ae for MHC peptide complexes) were applied at 1 μg/ml for 1 h, the sections were washed in PBS, and alkaline phosphatase–coupled, avidin biotin complexes (AK-5000 alkaline phosphatase standard kit; Vector Labs Inc., Burlingame, CA) were applied for 30 min. The sections were washed and the blue alkaline phosphatase reaction product developed with a substrate kit (SK-5300; Vector Labs). Rat mAb was then added as a culture supernatant for 1 h. After washing, horseradish peroxidase–conjugated F(ab)′2 anti–rat Ig (No. 212-036-082; Jackson ImmunoResearch Labs Inc., West Grove, PA) was reconstituted as recommended for extended storage, diluted 1:200, and applied for 1 h. DAB (Stable DAB No. 750118; Research Genetics, Huntsville, AL) was then used to develop a brown reaction product. The slides were mounted in PBS glycerol, followed by a peroxidase avidin complex and brown diaminobenzidine reaction product.
FACS® Analyses.
Biotin-modified mAbs were prepared for CD11c, Y-Ae, and MHC II (clone M5/114) and identified with PE-streptavidin. Rat mAbs were used as hybridoma culture supernatants and identified with FITC-mouse anti–rat Ig (Boehringer Mannheim). We purchased FITC-modified mAb to CD40, CD4, CD8, CD54, and CD86 from PharMingen. In most experiments, cells were fixed and permeabilized, since many of the antigens were primarily intracellular in DCs (DEC-205, MIDC-8, and 2A1). Cells were fixed in 4% paraformaldehyde (Fisher Scientific Co., Pittsburgh, PA) for 30 min, washed twice with PBS, permeabilized with 0.5% saponin (Sigma Chemical Co., St. Louis, MO) in PBS with 1% FCS at 4°C for 10 min, and then washed twice with staining medium.
FACS® Purification of Lymph Node DCs and B Cells. Starting with a low density fraction from BSA columns (see above; 3–4% of lymph node cells) CD11c+ B220− DCs were sorted from CD11c− B220+ B cells on a FACStarPLUS instrument and with FITC-CD11c and PE-B220 mAbs from Endogen, Inc. (Boston, MA) and PharMingen, respectively. The sorted fractions were confirmed to be >98% pure.
Antigen-presenting Function.
T-T hybrids, specific for a Y–Ae complex, were provided by L. Ignatowitz (University of Colorado, Denver, CO; reference 10). The response to APCs was assessed by IL-2 release or apoptosis 18 h later. IL-2 release was measured in a bioassay on conA activated T blasts and the CTLL line. Apoptosis was measured as a decrease in 3H-TdR uptake, and reduced or hypodiploid DNA staining with propidium iodide, 50 μg/ml in 0.1% sodium citrate and 0.1% Triton X-100. The APCs were FACS® purified DCs or B cells added in graded doses to 105 T-T hybridoma cells.
Results
T Cell Areas Contain Dendritic Profiles with Very High Levels of an MHC II–Self Peptide Complex.
The Y-Ae monoclonal recognizes a complex of an I-Eα peptide presented on I-Ab MHC class II molecules (6). Tissue sections of C57BL6 × DBA/2 (I-Ab x I-Eα) lymph nodes which were stained with Y-Ae showed stronger reactivity in T cell areas relative to B cell follicles (Fig. 1). The strong Y-Ae staining in T areas was likely on DCs, since mouse T cells do not express MHC class II and since T cell areas have relatively few macrophages and B cells (11, 12). Prior work had shown strong staining with Y-Ae in thymic medulla (13), but this site has numerous medullary epithelial cells in addition to DCs (14).
Figure 1.
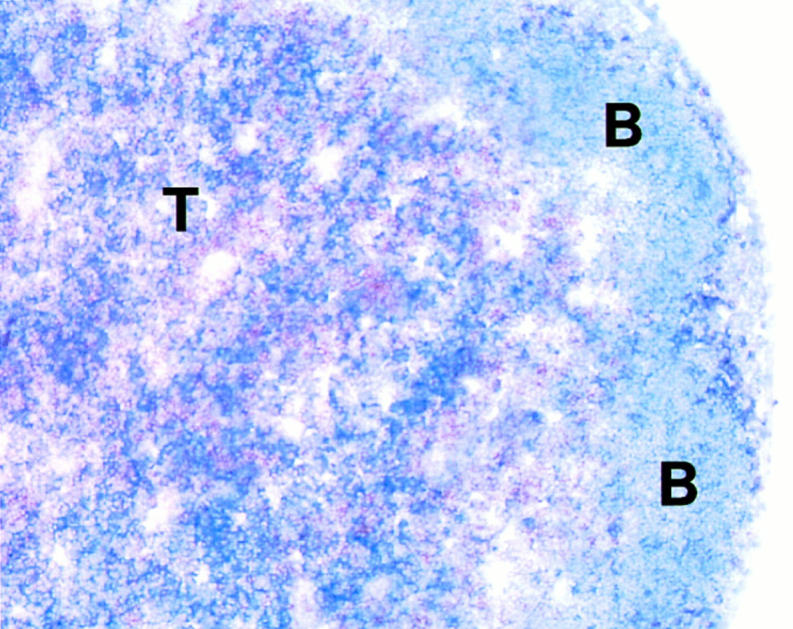
Stronger staining for MHC–self peptide complexes in lymph node T cell (T) vs. B cell areas (B). Double labeling for Y-Ae (blue) and CD8+ T cells (brown).
Markers for DCs in the T Cell Areas.
To identify and isolate the cells which expressed high levels of MHC II–self peptide complexes from lymph nodes, we first stained sections with a panel of monoclonals which react with isolated mature DCs but not with T cells and thymocytes. T cell areas contained many profiles which labeled for (a) a panel of mAbs which react with antigens within intracellular granules of mature DCs and are termed 2A1 (8), M342 (15), and MIDC-8 (16); (b) the CD11c integrin which is abundant on DCs (17, 18), and (c) the DEC-205 multilectin receptor expressed by DCs (19–21). The expression of each antigen was weaker (2A1) or not detected (CD11c, DEC-205, M342, and MIDC-8) on B cells. Antigen-presenting (MHC II, invariant chain) and costimulatory (CD54 and CD86) molecules also were prominently displayed on dendritic profiles in the T cell areas (not shown, but for reviews see references 11 and 22).
Fig. 2 illustrates the distribution of the 2A1 staining. This monoclonal stains granules within isolated mature DCs as described (8), but the molecule expressing the 2A1 antigen has yet to be identified. In sections, strong staining is seen on many profiles in the T cell areas, with weaker staining in the B cell areas. However, 2A1 staining is not seen on most DCs which are isolated by earlier methods from spleen and lymph node. Two other mAbs, M342 (15) and MIDC-8 (16), also do not stain freshly isolated less mature DCs but only stain after the cells have been cultured overnight.
Figure 2.

Staining for the 2A1 antigen which is abundant in DCs in lymph node T areas. (a and b) Low and high power views for macrophages (sialoadhesin+) in the subcapsular sinus (arrows) and medulla (brown) and the 2A1 antigen in the deep cortex (T area) and to a less extent B cell follicles (blue). (c) The deep cortex with several venules (*) is stained for the 2A1 antigen (brown) and counterstained with hematoxylin.
Isolation of Cells Carrying the Markers of T Cell Area DCs.
By emphasizing the use of Ca2+-free media (23), we were able to release and enrich (on BSA columns) cells with all the above markers of T cell area DCs from lymph nodes. Standard dissociation procedures with collagenase yielded very few cells that stained for DEC-205, CD11c, or the 2A1, M342, and MIDC-8 granule antigens which are found on T cell area DCs (Fig. 2). However Ca2+-free media increased the yield of CD11c+ cells 25-fold from lymph node (Table 1). The large cells had all the markers of T cell area DCs, including large cell size, irregular shape, abundant MHC II, DEC-205, and granule antigens (Fig. 3). Most other low density cells were smaller, CD11c− B220+ B cells (Fig. 3, right). Few macrophages were present, by cytology or staining for SER-4 (24) and F4/80 (12) antigens.
Table 1.
Calcium-free Media Increase Yields of Lymph Node DCs
| Tissue/medium | n | Total cells (×107) | Percentage of low density | Percentage of CD11c+≳ | ||||
|---|---|---|---|---|---|---|---|---|
| Lymph node, Hanks | 2 | 2.31 ± 0.45 | 2.4 ± 0.02 | 0.04 ± 0.05 | ||||
| Lymph node, Ca-free | 7 | 2.32 ± 0.25 | 3.0 ± 0.59 | 0.84 ± 0.11 | ||||
| Spleen, Hanks | 3 | 19.6 ± 0.9 | 8.8 ± 0.89 | 0.67 ± 0.06 (10.6) | ||||
| Spleen, Ca-free | 7 | 20.0 ± 1.5 | 4.3 ± 0.78 | 0.95 ± 0.19 (22.0) |
In spleen, the percentages of T cell area DCs are in parentheses and correspond to the percentage of DEC-205 or CD8 positive cells in the CD11c+ fraction.
Figure 3.

Enrichment of lymph node cells (see Materials and Methods) with many markers of T cell area DCs: MHC II, DEC-205, M342+ and MIDC-8+ granules, and B220+ B cells. Typical DCs (black arrowheads) and B cells (white arrowhead) are present.
The use of Ca2+-free media followed by flotation on dense BSA allowed the isolation of cells which were B220− (not B cells) but had very high levels of Y-Ae MHC–peptide complex, higher than the B220+ B cells (Fig. 4 A, compare populations with black and white arrows). The Y-Ae+ cells were only found in mice which expressed both I-Ab (C57BL/6) and the I-Eα peptide (BALB/c)(Fig. 4 A). If the low density cells from BALB/c × C57BL/6 F1 mice were double labeled for B220 and either CD11c integrin or Y-Ae, it was clear that CD11c high and Y-Ae high populations (Fig. 4 B, black arrow), were B220−. The B220+ cells had little or no CD11c but intermediate levels of Y-Ae. By sorting cells into CD11c+ B220− and CD11c− B220+ fractions, we were able to obtain 98% pure populations of DCs and B cells, respectively.
Figure 4.

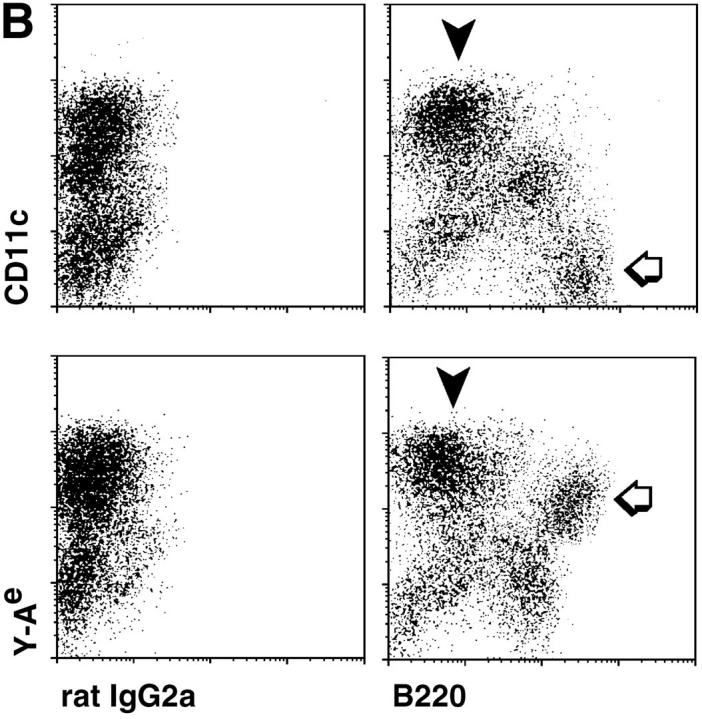
Double labeling of lymph node, low density cells for CD11c integrin, B220 and Y-Ae. (A) In bulk lymph node suspensions, most of the Y-Ae+ cells are B220+ and only found in the appropriate mouse strain, BALB/c × C57BL/6 (white arrow), but by enriching DCs in a fraction with a low buoyant density, a B220− population with very high levels of Y-Ae emerges (black arrow). (B) Double labeling with B220 and either CD11c or Y-Ae to show the DCs (black arrowheads) and B cells (white arrows).
The CD11c+ cells were then labeled with a panel of mAbs to ensure that they carried the markers of T cell area DCs and to look for other cell-specific and costimulatory molecules. CD11c+ lymph node cells carried the same markers as DCs in sections of T cell areas (Fig. 5, top row, black arrows). DEC-205, 2A1, and MIDC-8 (latter not shown) were all abundant on most CD11c+ cells, as was CD86, CD54, CD40, and MHC I (latter not shown). The phagocyte markers, SER-4/sialoadhesin and RB6/GR-1, were not expressed, and the T cell markers, CD8 and CD4, were weak on most profiles (Fig. 5). In mesenteric nodes, or in peripheral lymph nodes stimulated with incomplete Freund's adjuvant, most of the cells with T cell markers had high levels of CD11c, whereas with unstimulated peripheral lymph node (axillary, inguinal, and cervical), many of the DCs had intermediate levels of CD11c (not shown).
Figure 5.

Double labeling of lymph node and spleen for CD11c or Y-Ae on the y axis and a panel of mAbs on the x axis. All cells were fixed in formaldehyde and permeabilized with saponin to reveal predominantly intracellular antigens like DEC-205 and 2A1 (see Fig. 6). In lymph node, many CD11c+ cells and the strongest Y-Ae+ cells express the markers of T cell area DCs (black arrows). In spleen, only a subset of CD11c+ cells, but all the strong Y-Ae+ cells, carry markers of T cell area DCs (black arrows).
In the spleen (Fig. 5), two populations of CD11c+ cells were evident. About 50% expressed high levels of CD8 and DEC-205, but only a fraction had high levels of 2A1. The CD11c-rich cells had the highest levels of the CD86, CD54, and CD40 accessory molecules (Fig. 5). We conclude that the majority of CD11c+ cells that were released under Ca2+-free conditions from lymph node, and to a lesser extent spleen, carried the DC markers which were found in sections through the T cell areas.
Expression of the MHC II–Peptide Complex Recognized by Y-Ae mAb.
Semiquantitative FACS® data on the markers of cells expressing the I-Ab/I-Eα, MHC II-peptide complexes were then obtained. The cells with the highest labeling for Y-Ae in lymph node (Fig. 5, black arrows) carried all the markers of T cell area DCs (CD11c, high MIDC-8, DEC-205, and 2A1), as well as the highest levels of MHC I, CD40, CD54, CD86, and heat stable antigen. This was especially impressive in spleen where the cells with the most abundant Y-Ae were exclusively those with the DEC-205 and 2A1 DC markers (Fig. 5, black arrows).
To identify cells with strong staining for DEC-205, granule antigens like 2A1 and MIDC-8, and the lysosomal markers CD107a (LAMP-1) and CD68, it was necessary to permeabilize the cells with saponin before staining. The different staining results with fixed and fixed permeabilized cells are illustrated in Fig. 6 (compare F with FP).
Figure 6.

Markers of T cell area DCs are identified after saponin permeabilization. Low density, lymph node cells, fixed with HCHO (F) or fixed and permeabilized with saponin (FP), were stained with biotin Y-Ae (y axis) and mAbs (x axis). DEC-205, CD107a (LAMP-1), and CD68 are all found in the endocytic system.
To rule out acquisition of I-Eα peptide from B cells, we stained DC suspensions which lacked B cells, i.e., DCs from epidermis (9) and from bone marrow–progenitors (8). Both epidermal (Fig. 7 A) and bone marrow–derived (Fig. 7 B) DCs expressed high levels of Y-Ae and the CD86 costimulator. Again only the cells from the appropriate mouse strain (C57BL/6 × DBA/2 F1 vs. C57BL/6) expressed the Y-Ae epitope.
Figure 7.


DCs which have not been exposed to B cells express high levels of the Y–Ae, MHC–peptide complex. DCs were isolated from cultures which lack B cells, epidermal cells (A) and bone marrow stimulated with GM-CSF (B). In each instance, the DCs have high levels of CD86 and MHC II or MHC II–self peptide complexes.
Presentation of the Y-Ae Epitope to T-T Hybridomas.
FACS®-purified, CD11c+ B220− DCs and CD11c− B220+ B cells were used to present the Y-Ae epitope to two T-T hybrids which recognize I-Ab/I-Eα peptide complexes (10). The DCs were >30-fold more potent in inducing IL-2 release and inducing the hybridomas to stop synthesizing DNA (Fig. 8 A), an indication of apoptosis (25). To verify that apoptosis had taken place, the cells were stained with propidium iodide and shown to have high levels of hypodiploid profiles 24 h after stimulation with DCs from the appropriate strain of mice (Fig. 8 B, compare B6D2 with B6 DCs).
Figure 8.


Sorted lymph node DCs are potent APCs for IL-2 release and death of Y-Ae-specific, T-T hybridomas. Function is shown for sorted DCs and B cells from C57BL/6 and B6 × D2 F1 mice. (A) The response was measured as IL-2 release (bottom, proliferation of conA blasts) or 3H-TdR uptake (top) by the T-T hybrid, the latter being reduced by apoptosis. ▪, C57BL/6 B cells; □, B6D2F1 B cells; --•--, C57BL/6 DCs; --○-- B6D2F1 DCs; ○, None. (B) Apoptosis was measured by identifying hypodiploid profiles using propidium iodide staining. Extensive apoptosis occurs when the DCs are from B6D2 mice which express the Y-Ae epitope.
Discussion
An obstacle to the study of antigen presentation in vivo is the difficulty in isolating DCs from lymph nodes, the major organs in which immune responses are generated. This obstacle is paradoxical since sections of T cell areas contain many cells with features of DCs (Fig. 1 and 2; for reviews see references 3–5). In contrast, spleen yields many DCs, yet T cell areas represent a much smaller proportion of spleen than lymph node. Many of the DCs which are released from spleen emanate from the periphery of the white pulp (17, 18) and must be cultured to develop the properties of T cell area DCs, such as high expression of MIDC-8, 2A1, and M342 positive granules, DEC-205, and MHC II and CD86 (15, 22). A subset of spleen DCs which carries CD8 and has a regulatory role, dampening T cell responses, was identified using Ca2+-free media (23, 26, 27). By emphasizing Ca2+-free media, lymph node cells which have all the properties of T cell area DCs can be isolated, although CD8 expression is weak.
Functional assays with cells from different tissues have shown that DCs are the main cell type which carries foreign (28–31) and self (32) antigens. However, strong APC function may reflect either high levels of MHC–peptide complexes (“signal one”) and/or an abundance of T cell adhesion and costimulatory molecules (“signal two”), e.g., ICAM-1/CD54 and B7-2/CD86. Our data indicate that both signal one and two are abundant on T cell area DCs. The Y-Ae monoclonal directly shows that peripheral DCs are presenting MHC II–self peptide complexes in high amounts, and that these DCs can induce cell death in T-T hybridomas which are specific for the MHC II–self peptide complex. Death of T cell hybridomas occurs when DCs present foreign antigens to T-T hybrids as well (our unpublished observation).
Kurts et al. (33) recently reported functional data that an MHC I–self peptide complex from pancreatic islet β cells is expressed on bone marrow–derived APCs in the draining lymph node. They have since shown that these APCs mediate peripheral tolerance (34). DCs have long been considered as adjuvants for responses to foreign antigens, but it is possible that some T cell area DCs can have a regulatory or tolerizing role, perhaps by the expression of fas-ligand, as suggested by the data from Suss and Shortman (26). The reason to consider a role for DCs in peripheral tolerance relates to two well studied systems in vivo, presentation of H-Y antigens and superantigens. These ligands elicit strong immune responses when presented on DCs which have been allowed to mature in vitro (35, 36), yet the same antigens can be tolerogenic in vivo (37–39). Therefore DCs should be assessed in the regulation of autoreactivity and not just immunogenicity to foreign antigens.
Acknowledgments
This work was supported by grants from the Japanese Ministry of Education, Science, and Culture (08282104 and 08044271), the Yamanouchi Fund, the Juvenile Diabetes Foundation, the Human Science Frontiers Program, and National Institutes of Health grants AI-13013 and DK-39672.
Abbreviation used in this paper
- DC
dendritic cell
References
- 1.Liu Y-J, Grouard G, de Bouteiller O, Banchereau J. Follicular dendritic cells and germinal centers. Int Rev Cytol. 1996;166:139–179. doi: 10.1016/s0074-7696(08)62508-5. [DOI] [PubMed] [Google Scholar]
- 2.Szakal AK, Kosco MH, Tew JG. Microanatomy of lymphoid tissue during humoral immune responses: structure function relationships. Annu Rev Immunol. 1989;7:91–109. doi: 10.1146/annurev.iy.07.040189.000515. [DOI] [PubMed] [Google Scholar]
- 3.Steinman RM. The dendritic cell system and its role in immunogenicity. Annu Rev Immunol. 1991;9:271–296. doi: 10.1146/annurev.iy.09.040191.001415. [DOI] [PubMed] [Google Scholar]
- 4.Williams LA, Egner W, Hart DN. Isolation and function of human dendritic cells. Int Rev Cytol. 1994;153:41–103. doi: 10.1016/s0074-7696(08)62188-9. [DOI] [PubMed] [Google Scholar]
- 5.Steinman RM, Pack M, Inaba K. Dendritic cells in the T cell areas of lymphoid organs. Immunol Rev. 1997;156:25–37. doi: 10.1111/j.1600-065x.1997.tb00956.x. [DOI] [PubMed] [Google Scholar]
- 6.Rudensky AY, Rath S, Preston-Hurlburt P, Murphy DB, Janeway CA., Jr On the complexity of self. Nature (Lond) 1991;353:660–662. doi: 10.1038/353660a0. [DOI] [PubMed] [Google Scholar]
- 7.Swiggard, W.J., R.M. Nonacs, M.D. Witmer-Pack, K. Inaba, and R.M. Steinman. 1992. Enrichment of dendritic cells by plastic adherence and EA rosetting. In Current Protocols in Immunology. J.E. Coligan, A.M. Kruisbeek, D.H. Margulies, E.M. Shevach, and W. Strober, editors. Greene Publishing Associates and Wiley-Interscience, New York. 3.7.1–3.7.11.
- 8.Inaba K, Inaba M, Romani N, Aya H, Deguchi M, Ikehara S, Muramatsu S, Steinman RM. Generation of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony–stimulating factor. J Exp Med. 1992;176:1693–1702. doi: 10.1084/jem.176.6.1693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Schuler G, Steinman RM. Murine epidermal Langerhans cells mature into potent immunostimulatory dendritic cells in vitro. J Exp Med. 1985;161:526–546. doi: 10.1084/jem.161.3.526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ignatowicz L, Kappler J, Marrack P. The repertoire of T cells shaped by a single MHC/peptide ligand. Cell. 1996;84:521–529. doi: 10.1016/s0092-8674(00)81028-4. [DOI] [PubMed] [Google Scholar]
- 11.Witmer MD, Steinman RM. The anatomy of peripheral lymphoid organs with emphasis on accessory cells: light microscopic, immunocytochemical studies of mouse spleen, lymph node and Peyer's patch. Am J Anat. 1984;170:465–481. doi: 10.1002/aja.1001700318. [DOI] [PubMed] [Google Scholar]
- 12.Hume DA, Robinson AP, MacPherson GG, Gordon S. The mononuclear phagocyte system of the mouse defined by immunohistochemical localization of antigen F4/ 80. Relationship between macrophages, Langerhans cells, reticular cells, and dendritic cells in lymphoid and hematopoietic organs. J Exp Med. 1983;158:1522–1536. doi: 10.1084/jem.158.5.1522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Murphy DB, Lo D, Rath S, Brinster RL, Flavell RA, Slanetz A, Janeway CA., Jr A novel MHC class II epitope expressed in thymic medulla but not cortex. Nature (Lond) 1989;338:765–768. doi: 10.1038/338765a0. [DOI] [PubMed] [Google Scholar]
- 14.Surh CD, Gao E-K, Kosaka H, Lo D, Ahn C, Murphy DB, Karlsson L, Peterson P, Sprent J. Two subsets of epithelial cells in the thymic medulla. J Exp Med. 1992;176:495–505. doi: 10.1084/jem.176.2.495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Agger R, Witmer-Pack M, Romani N, Stossel H, Swiggard WJ, Metlay JP, Storozynsky E, Freimuth P, Steinman RM. Two populations of splenic dendritic cells detected with M342, a new monoclonal to an intracellular antigen of interdigitating dendritic cells and some B lymphocytes. J Leukocyte Biol. 1992;52:34–42. doi: 10.1002/jlb.52.1.34. [DOI] [PubMed] [Google Scholar]
- 16.Breel M, Mebius RE, Kraal G. Dendritic cells of the mouse recognized by two monoclonal antibodies. Eur J Immunol. 1987;17:1555–1559. doi: 10.1002/eji.1830171105. [DOI] [PubMed] [Google Scholar]
- 17.Metlay JP, Witmer-Pack MD, Agger R, Crowley MT, Lawless D, Steinman RM. The distinct leukocyte integrins of mouse spleen dendritic cells as identified with new hamster monoclonal antibodies. J Exp Med. 1990;171:1753–1771. doi: 10.1084/jem.171.5.1753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Witmer-Pack MD, Crowley MT, Inaba K, Steinman RM. Macrophages, but not dendritic cells, accumulate colloidal carbon following administration in situ. J Cell Sci. 1993;105:965–973. doi: 10.1242/jcs.105.4.965. [DOI] [PubMed] [Google Scholar]
- 19.Kraal G, Breel M, Janse M, Bruin G. Langerhans cells, veiled cells, and interdigitating cells in the mouse recognized by a monoclonal antibody. J Exp Med. 1986;163:981–997. doi: 10.1084/jem.163.4.981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Witmer-Pack MD, Swiggard WJ, Mirza A, Steinman RM. Tissue distribution of the DEC-205 protein that is detected by the monoclonal antibody NLDC-145. II. Expression in situ in lymphoid and nonlymphoid tissues. Cell Immunol. 1995;163:157–162. doi: 10.1006/cimm.1995.1110. [DOI] [PubMed] [Google Scholar]
- 21.Jiang W, Swiggard WJ, Heufler C, Peng M, Mirza A, Steinman RM, Nussenzweig MC. The receptor DEC-205 expressed by dendritic cells and thymic epithelial cells is involved in antigen processing. Nature (Lond) 1995;375:151–155. doi: 10.1038/375151a0. [DOI] [PubMed] [Google Scholar]
- 22.Inaba K, Witmer-Pack M, Inaba M, Hathcock KS, Sakuta H, Azuma M, Yagita H, Okumura K, Linsley PS, Ikehara S, et al. The tissue distribution of the B7-2 costimulator in mice: abundant expression on dendritic cells in situ and during maturation in vitro. J Exp Med. 1994;180:1849–1860. doi: 10.1084/jem.180.5.1849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Vremec D, Zorbas M, Scollay R, Saunders DJ, Ardavin CF, Wu L, Shortman K. The surface phenotype of dendritic cells purified from mouse thymus and spleen: investigation of the CD8 expression by a subpopulation of dendritic cells. J Exp Med. 1992;176:47–58. doi: 10.1084/jem.176.1.47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Crocker PR, Gordon S. Mouse macrophage hemagglutinin (sheep erythrocyte receptor) with specificity for sialylated glycoconjugates characterized by a monoclonal antibody. J Exp Med. 1989;169:1333–1346. doi: 10.1084/jem.169.4.1333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yang Y, Mercep M, Ware CF, Ashwell JD. Fas and activation-induced Fas ligand mediate apoptosis of T cell hybridomas: inhibition of Fas ligand expression by retinoic acid and glucocorticoids. J Exp Med. 1995;181:1673–1682. doi: 10.1084/jem.181.5.1673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Suss G, Shortman K. A subclass of dendritic cells kills CD4 T cells via Fas/Fas-ligand induced apoptosis. J Exp Med. 1996;183:1789–1796. doi: 10.1084/jem.183.4.1789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kronin V, Winkel K, Suss G, Kelso A, Heath W, Kirberg J, von Boehmer H, Shortman K. A subclass of dendritic cells regulates the response of naive CD8 T cells by limiting their IL-2 production. J Immunol. 1996;157:3819–3827. [PubMed] [Google Scholar]
- 28.Bujdoso R, Hopkins J, Dutia BM, Young P, McConnell I. Characterization of sheep afferent lymph dendritic cells and their role in antigen carriage. J Exp Med. 1989;170:1285–1302. doi: 10.1084/jem.170.4.1285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Crowley M, Inaba K, Steinman RM. Dendritic cells are the principal cells in mouse spleen bearing immunogenic fragments of foreign proteins. J Exp Med. 1990;172:383–386. doi: 10.1084/jem.172.1.383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Liu LM, MacPherson GG. Antigen acquisition by dendritic cells: intestinal dendritic cells acquire antigen administered orally and can prime naive T cells in vivo. J Exp Med. 1993;177:1299–1307. doi: 10.1084/jem.177.5.1299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Guery J-C, Ria F, Adorini L. Dendritic cells but not B cells present antigenic complexes to class II–restricted T cells after administration of protein in adjuvant. J Exp Med. 1996;183:751–757. doi: 10.1084/jem.183.3.751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Guery J-C, Adorini L. Dendritic cells are the most efficient in presenting endogenous naturally processed self-epitopes to class II-restricted T cells. J Immunol. 1995;154:536–544. [PubMed] [Google Scholar]
- 33.Kurts C, Heath WR, Carbone FR, Allison J, Miller JFAP, Kosaka H. Constitutive class I–restricted exogenous presentation of self antigens in vivo. J Exp Med. 1996;184:923–930. doi: 10.1084/jem.184.3.923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kurts C, Kosaka H, Carbone FR, Miller JFAP, Heath WR. Class I–restricted cross-presentation of exogenous self antigens leads to deletion of autoreactive CD8+ T cells. J Exp Med. 1997;186:239–245. doi: 10.1084/jem.186.2.239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Boog CJP, Kast WM, Timmers HTM, Boes J, De Waal LP, Melief CJM. Abolition of specific immune response defect by immunization with dendritic cells. Nature (Lond) 1985;318:59–62. doi: 10.1038/318059a0. [DOI] [PubMed] [Google Scholar]
- 36.Bhardwaj N, Friedman SM, Cole BC, Nisanian AJ. Dendritic cells are potent antigen-presenting cells for microbial superantigens. J Exp Med. 1992;175:267–273. doi: 10.1084/jem.175.1.267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Rocha B, von Boehmer H. Peripheral selection of the T cell repertoire. Science (Wash DC) 1991;251:1225–1228. doi: 10.1126/science.1900951. [DOI] [PubMed] [Google Scholar]
- 38.Renno T, Hahne M, MacDonald HR. Proliferation is a prerequisite for bacterial superantigen-induced T cell apoptosis in vivo. J Exp Med. 1995;181:2283–2287. doi: 10.1084/jem.181.6.2283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mittrucker H-W, Shahinian A, Bouchard D, Kundig TM, Mak TW. Induction of unresponsiveness and impaired T cells expansion by staphylococcal enterotoxin B in CD28-deficient mice. J Exp Med. 1996;183:2481–2488. doi: 10.1084/jem.183.6.2481. [DOI] [PMC free article] [PubMed] [Google Scholar]