

Abstract
The embryonic thymus is colonized by the influx of hemopoietic progenitors in waves. To characterize the T cell progeny of the initial colonization waves, we used intravenous adoptive transfer of bone marrow progenitors into congenic embryos. The experiments were performed in birds because intravenous cell infusions can be performed more efficiently in avian than in mammalian embryos. Progenitor cells, which entered the vascularized thymus via interlobular venules in the capsular region and capillaries located at the corticomedullary junction, homed to the outer cortex to begin thymocyte differentiation. The kinetics of differentiation and emigration of the T cell progeny were analyzed for the first three waves of progenitors. Each progenitor wave gave rise to γ/δ T cells 3 d earlier than α/β T cells. Although the flow of T cell migration from the thymus was uninterrupted, distinct colonization and differentiation kinetics defined three successive waves of γ/δ and α/β T cells that depart sequentially the thymus en route to the periphery. Each wave of precursors rearranged all three TCR Vγ gene families, but displayed a variable repertoire. The data indicate a complex pattern of repertoire diversification by the progeny of founder thymocyte progenitors.
Comparative developmental studies have been informative with regard to the evolution of the immune system in vertebrates. Studies in chickens have contributed to the understanding of the hemopoietic stem cell origin of both myeloid and lymphoid T and B cell lineages (1, 2). This avian model has several advantages for the study of T cell development: (a) T and B cells undergo differentiation in specialized central lymphoid organs, T cells in the thymus, and B cells in the bursa of Fabricius, (b) a large number of precisely staged embryos can be easily obtained, and (c) the embryos are large enough for experimental manipulation. Studies performed in chick–quail chimeras indicate an embryonic paraaortic origin of the stem cell precursors of thymocytes, B cells, and myeloid cells, beginning around the fourth day of embryonic life (E4;1 reference 3). Embryonic stem cells native to the aortic region later migrate via the circulation to colonize the spleen, yolk sac, and, finally, the bone marrow.
The chick–quail model has been used to show homing of the thymocyte progenitors into the embryonic epithelial thymus in three discrete waves (4–8), the first of which begins in chicken embryos on E6.5, the second on E12, and the third around E18. Each wave of progenitor cell influx lasts for 1 or 2 d, and is followed by the transient production of thymocyte progeny (7, 8). The first wave of thymus colonization involves T cell progenitors from the paraaortic region (7, 8), whereas the second and third waves of thymocyte progenitors come from the bone marrow and express the c-kit and the hematopoietic cell adhesion molecules (9, 10). Using congenic chicken strains that differ in the ov alloantigen expressed on hematopoietic progenitors and T lineage cells, H.B19ov+ and H.B19ov−, we have examined chimeras created by grafting thymic lobes from an ov+ donor into thymectomized ov− recipients to show the gradual replacement of donor thymocytes by ov− host thymocytes and their progeny. These experiments indicated that a series of waves or stream of thymocyte progenitors continually enter the thymus after hatching (11–13).
The ontogeny of chick T lineage cells can be monitored with anti-TCR monoclonal antibodies and molecular probes for the different TCR chains (14–16). At E12, ∼ 5 d after the initial influx of thymocyte precursors, a subpopulation of thymocytes begins to express the TCR-γ/δ–CD3 complex on their surface (17). These reach peak numbers on E15, when ∼30% of the thymocytes express TCR-γ/δ (18). TCR-α/β–bearing T cells expressing the Vβ1 variable domain are first detected on thymocytes on E15, and they become the predominant type of thymocytes by E17–18 (19). TCR-α/β–bearing T cells expressing the Vβ2 variable domain emerge around E18 (20). In chick–quail chimeras, the γ/δ and α/β (Vβ1 then Vβ2) T cell subpopulations were generated sequentially in the first wave, but not in the second wave of thymocyte progenitors (8).
Thymocyte transfer experiments in congenic chick strains indicate that the γ/δ and (Vβ1) α/β thymocytes generated by the different thymocyte progenitor waves colonize the peripheral lymphoid organs in discrete waves (11–13, 21). Examination of the TCR Vβ1 repertoire generated by each wave of thymocyte progenitors and expressed by their progeny in the thymus, spleen, and intestines indicates that all of the different Vβ1 gene segments are expressed as early as E17. The thymic Vβ1-Dβ-Jβ repertoire expressed by each of the three waves of hemopoietic progenitors includes the same Vβ1 and Jβ elements, and CDR3 created by the Vβ1-Dβ-Jβ junctions of similar lengths (13). The spleen is colonized both by Vβ1 and Vβ2 T cells, whereas it is difficult to find Vβ2 T cells in the intestine (13, 20).
TCR-γ genes have been identified recently in the chicken. These include three Vγ families, three Jγ segments, and one Cγ region (15, 16). Although ontogenetic studies in mice indicate that TCR-γ gene rearrangement proceeds in waves with γ/δ T cells expressing the different Vγ gene segments are generated sequentially (22–25), pilot studies in the chicken suggested that TCR-γ rearrangement may not be so tightly regulated during avian development (16). In the present study, we have examined the potential of each wave of embryonic progenitors to produce TCR-γ/δ and TCR-α/β (Vβ1) thymocytes by adoptive transfer of hemopoietic progenitors into congenic embryos. The ov alloantigen marker was used to purify the thymocyte progeny from the individual waves of progenitors colonizing the thymus. Repertoire analysis demonstrated that although the precursors of each wave rearrange all three TCR Vγ gene families, each wave may display differing Vγ and Jγ usage. The sequential differentiation of γ/δ and α/β T cells was a constant feature of all three developmental waves. Irrespective of the wave of progenitor colonization, the γ/δ T cell progeny differentiated about 3 d faster than the α/β T cell progeny. Likewise, the migration of γ/δ and α/β T cells was found to occur to in an alternating fashion during each migration wave from the thymus to the periphery.
MATERIALS AND METHODS
Animals.
Embryonated eggs from the H.B19 strain of White Leghorn chickens were produced at the Institute Chicken Facility (Gipf-Oberfrick, Switzerland). Fertilized eggs were incubated at 38°C and 80% humidity in a ventilated incubator. The H.B19 strain was subdivided into two congenic lines, H.B19ov+ and H.B19ov−, distinguished by the ov antigen present on T lineage cells in H.B19ov+ animals. The ov antigen, which is also expressed on bone marrow cells and a B cell subset, is recognized by the 11-A-9 monoclonal antibody (9, 26, 27).
Immunolabeling.
The ov, TCR-γ/δ and TCR Vβ1 antigens were detected by the 11-A-9, TCR1, and TCR2 mAbs, respectively (17, 19, 26, 28). 11-A-9 is a mouse IgM and TCR1 and TCR2 are mouse IgG1 antibodies. Second step antibodies were fluorescein labeled, sheep anti–mouse IgM and phycoerythrin- or Texas red–coupled anti–mouse IgG1 antibodies (Southern Biotechnology Assoc., Birmingham, AL). Controls were performed using the second step antibodies alone and regular staining of tissues from noninjected individuals of the H.B19ov− strain. Recent thymocyte emigrants, detected in blood by their FITC staining, were labeled by phycoerythrin-coupled TCR1 or TCR2 antibodies (Southern Biotechnology Assoc.). Frozen sections of embryonic organs were cut to a thickness of 5 μm on a cryostat (Bright, Hunkingdom, UK), fixed with acetone, and rehydrated in PBS containing 1% BSA.
Injection of Lymphoid Cells into Congenic Chickens.
Adoptive transfer between H.B19ov+ and H.B19ov− strains could be performed without complications since these strains do not differ at major histocompatibility antigens and T cell alloreactivity against a different ov antigenic determinant has not been observed in mixed lymphocyte reaction and graft versus host reactions. Bone marrow cells (2.0 × 107) from donor H.B19ov+ embryos were injected into a large vein near the airsac of recipient H.B19ov− embryos (29). These experiments were performed with E13 and E18 age-matched donor and recipient embryos. Control injections of sorted TCR-positive populations of E18 bone marrow cells were performed to determine that differentiated bone marrow lymphocytes were not able to colonize the thymus in this assay. For that purpose, bone marrow cells from 18-d-old H.B19ov+ embryos were suspended in PBS containing 10% FCS, filtered through a nylon sieve (mesh width of 25 μm; Nytal P-25 my; SST, Thal, Switzerland) and centrifuged at 225 g for 7 min. Immunofluorescence staining was performed in 96-well plates, to avoid repeated centrifugation using either the anti–TCR-γ/δ antibody TCR1 or the anti-TCR Vβ1 antibody TCR2 and then fluorescein-coupled anti–mouse Ig antibody (Silenus, Hawthorn, Australia). Stained and unstained bone marrow cells were resuspended in 10% FCS/ PBS and sorted using a FACSTAR Plus® cell sorter (Becton Dickinson, Mountain View, CA). None of the recipients received irradiation or other immunosuppressive treatment. Donor ov+ cells in the thymus were analyzed by flow cytometry and by two-color immunofluorescence staining of frozen tissue sections. For analysis by FACScan®, single thymocyte suspensions were made by physical disruption in PBS and filtration through a nylon sieve.
To analyze the TCR-γ repertoires specifically generated by E13 and E18 bone marrow precursors, γ/δ thymocytes of the donor type were sorted 9 d after injection of the precursors. Thymocytes were submitted to a two-color immunofluorescence staining using the anti–TCR-γ/δ antibody TCR1 and the anti-ov antibody 11-A-9 and then FITC-coupled anti–mouse IgM and phycoerythrin-coupled anti–mouse IgG1 antibodies (Southern Biotechnology Assoc.). The cells were sorted using a FACSTAR Plus® cell sorter.
Analysis of Recent Thymocyte Emigrants.
Emigration of the thymocytes into the circulation was examined after in situ FITC labeling of thymocytes. Young chicks were anesthesized by intramuscular injection of 0.4 ml ketamin solution (Imalgene 500; Rhone Mérieux, Lyon, France; diluted 1:10 in PBS) followed by a short inhalation of Halothane (Hoechst, Frankfurt, Germany). The skin of the neck region was opened with scissors and each thymus lobe was injected with 10 μl of an FITC solution at 1 mM in DMSO. The skin was then closed using tissue clamps (Autoclip; Clay Adams, Becton Dickinson Primary Care, Sparks, MD) and the chickens were kept warm under an infrared lamp until they were fully conscious. Chickens were bled 12 h after injection and FITC-labeled lymphocytes were analyzed by flow cytometry.
cDNA Synthesis.
Total cellular RNA from thymus was isolated using the guanidium isothiocyanate method (30). About 5 μg was used as template for the synthesis of randomly primed single-stranded cDNA using mouse mammary leukemia virus reverse transcriptase (Life Technologies, Inc., Gaithersburg, MD) in a reaction volume of 20 μl according to the supplier's instructions. This cDNA was subsequently diluted to 100 μl with water and heated to 94°C for 2 min to inactivate the mouse leukemia virus reverse transcriptase.
PCR and Semiquantitative PCR of Vγ Transcripts.
A PCR technique was used to amplify the TCR-γ transcripts. Transcripts deriving from rearranged TCR Vγ1, Vγ2, and Vγ3 genes were amplified independently using oligonucleotide primers specifics for each Vγ family and a primer located in the Cγ region. Oligonucleotide primers CKVG1UP2, CKVG2UP3, and CKVG3UP1 were specific for the Vγ1, Vγ2, and Vγ3 regions, respectively. The CKCG1DO1 oligonucleotide primer was located 230 nucleotides downstream the 5′ end of the Cγ region. The procedures used for semiquantitative PCR followed the detailed description given by Keller et al. (31). The amount of cDNA synthesized was calibrated by using the relative expression level of β actin as a standard. The two actin oligonucleotide primers, 4611 and 4612, generated a band of 283 and 648 bp on cDNA and genomic DNA respectively (32). The following are CKVG1UP2 (Vγ1 region): GCTACCAGAGAGAGATCC; CKVG2UP3 (Vγ2 region): CATACAGAGCCCTGTATC; CKVG3UP1 (Vγ3 region): GATACTGTACATGTCTGG; CKCG1DO1 (Cγ region, antisense): GACTCGAGCTCTCCAGTGGTACAGATAAC; CGAMMA1DD (5′ of Cγ, antisense): TTTCATGTTCCTCCTGC; 4611 (5′ of actin, from nucleotide 2057, see reference 32): TACCACAATGTACCCTGGC; 4612 (3′ of actin, from nucleotide 2704, antisense, see reference 32): CTCGTCTTGTTTTATGCGC.
PCR reactions were in 30 μl using 1 U of Taq polymerase (Perkin-Elmer Cetus, Norwalk, CT). PCR buffer was prepared as suggested by Perkin-Elmer, but with the addition of 10 mM β mercaptoethanol. Reaction mixtures were denatured by heating to 96°C for 5 min, and then subjected to 32 rounds of amplification using a Biometra Trio-thermoblock thermocycler under the following conditions: 96°C for 15 s, 50°C for 40 s, and 72°C for 1 min for cDNA amplification. Final extension was done at 72°C for 10 min.
Cloning and Sequencing of TCR-γ Transcripts.
The TCR-γ V-J-C regions were specifically amplified by PCR. Amplified DNA fragments were gel purified and cloned into pCR™II vector (Invitrogen, San Diego, CA). Sequences were determined from denatured double-stranded recombinant plasmid DNA (33) using SequenaseTM (Amersham Corp., Arlington Heights, IL) in the chain termination reaction (34) and the oligonucleotide primer CGAMMA1DD starting 60 nucleotides downstream the 5′ end of Cγ segment in the antisense orientation. In a number of cases where ambiguities remained, several additional nucleotide primers were used. Sequences were analyzed and assembled with the software package of the CITI2 (Paris, France). TCR-γ cDNA sequences have been submitted to the EMBL/GenBank/DDBJ database under accession numbers z97216 to z97332.
RESULTS
Pathways of Thymocyte Precursor Migration into the Embryonic Thymus.
The initial wave of progenitor colonization and thymocyte differentiation can be examined in situ in unmanipulated animals. However, analysis of members of the succeeding waves requires a discriminating strategy. Embryonic hemopoietic precursors express the ov antigen in H.B19ov+ birds, and the antigen is maintained on T lineage cells and a subset of B cells. This expression pattern allowed us to examine successive waves of the ov+ progenitor populations in the embryonic thymus and the fate of their T cell progeny in ov− congenic recipients. To examine the second wave of thymus colonization by progenitor cells, E13 bone marrow cells of the ov+ strain were injected into E13 chicken embryos of the congenic H.B19ov− strain, and the thymocyte progenitor influx, migration, and differentiation patterns were examined in thymus sections by immunohistochemistry. By E16, donor cells of bone marrow origin accumulated in the thymic blood vessels, both in interlobular venules and in capillaries located at the corticomedullary junction (Fig. 1); relatively few donor cells were found in the parenchyma of the thymus at this time. E19 ov+ cell invasion and accumulation within the thymic cortex was evident, but by E20 the donor cells had relocated to occupy the outermost cortex of the thymic lobules. This ontogenetic pattern suggests that thymocyte progenitors entering the embryonic thymus either via the corticomedullary junction or the capsular subsequently make their way to the outer cortex of the thymus (Fig.1 C). The donor cells were later found throughout the cortex and by day 23, mature ov+ donor T cells had begun to accumulate in the medulla. This complex intrathymic pattern of migration appears specific to bone marrow– derived thymocyte progenitors, since mature thymocytes and T cells failed to home to the thymus in other adoptive transfer experiments (not shown).
Figure 1.
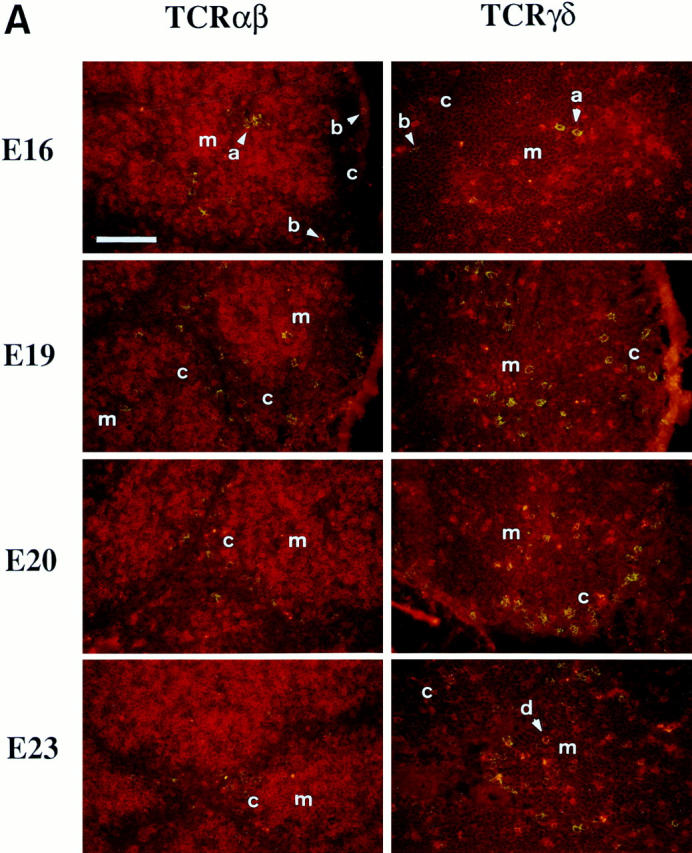


Migration pathways of thymocyte progenitors. Thymus sections from E13 H.B19ov− embryos injected with donor E13 H.B19ov+ bone marrow cells were examined by differential immunofluorescence staining at E16, E19, E20, and E23. Donor ov+ progenitors are labeled with fluorescein; TCR-γ/δ and TCR-α/β–positive cells are labeled with Texas red. Original magnifications: (A) 270, (B) 170. Scale bars correspond to 100 μm. (C) Diagrammatic representation of chicken T cell progenitor migration pathways in the thymus. At E16, T cell progenitors were located either in capillaries at the cortico-medullary junction (a) or close to the thymic capsule (b). Donor cells originally located at the corticomedullary junction or at the capsule were found later (E19) in the cortex, and by E20 had reached the outer cortex. By E23 (2 d after hatching), some donor cells were found in the medulla where they expressed TCR-α/β (Vβ1) or TCR-γ/δ (d). The first TCR-γ+ donor cells were found on E19-20. c, cortex; m, medulla.
Intrathymic Differentiation Kinetics Are Consistently Accelerated for the γ/δ T Cell Subpopulation.
The appearance of γ/δ T cells precedes that of Vβ1 α/β cells by a period of ∼3 d during the initial wave of thymocyte development (18), but studies in chick–quail chimeras suggest this may be a one time occurrence (8). In our studies of the second wave of thymocyte differentiation, the injection of E13 H.B19ov+ bone marrow cells into E13 H.B19ov− embryos led to the appearance of donor γ/δ+ thymocytes 5 d later and donor α/β thymocytes ∼8 d later (Figs. 2 and 3 A). The proportion of ov+ donor thymocytes expressing TCR-γ/δ peaked at 40% on day 21. The first donor α/β (Vβ1+) thymocytes were detected on E20 and these reached a peak level of 57% on E26 (Fig. 3 A). When the third wave of thymocyte differentiation was examined by injection of E18 ov+ bone marrow cells into E18 ov− recipients, the same rule held true; γ/δ T cells appeared 4 d before the α/β T cells (Fig. 3 B). Interestingly, for the cell transfer experiments performed during the second wave of precursor colonization, the level of chimerism was relatively greater for γ/δ than for α/β T cells (Fig. 3 C). However, taking into account the proliferation kinetics for the progeny of each precursor wave, the TCR-γ/δ thymocyte progeny appeared to be 12 to 16 times less numerous than the TCR-α/β (Vβ1) thymocyte progeny (Fig. 3 D, and data not shown). The levels of donor γ/δ and α/β thymocytes peaked at days 23 and 26, respectively, corresponding to the main period of second wave emigration to the periphery. The differential chimerism of γ/δ and α/β T cells thus may reflect the differential emigration kinetics of γ/δ and α/β T cells.
Figure 2.
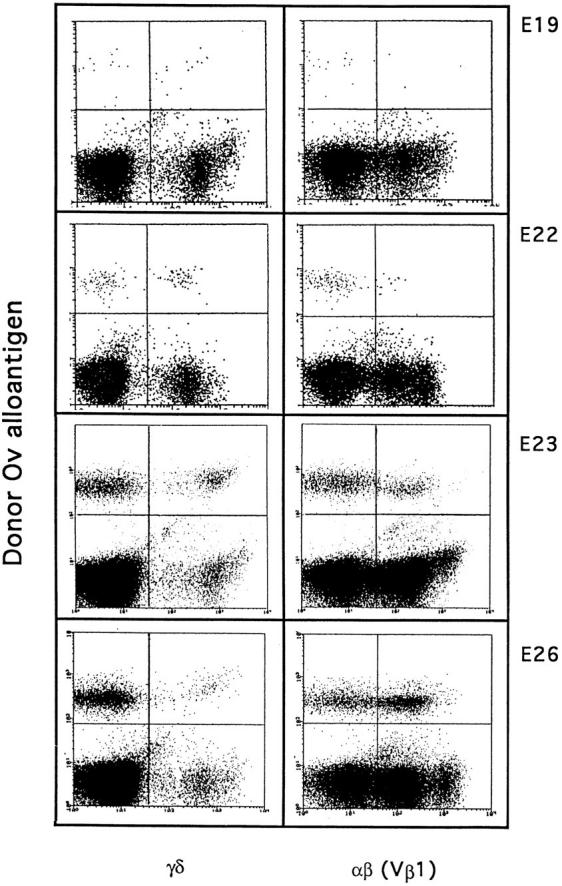
Differentiation kinetics of the second wave of thymocyte progenitors. After adoptive transfer of E13 H.B19ov+ bone marrow (2.0 × 106 cells) into E13 H.B19ov− embryos, the donor cells were examined for T cell expression as a function of developmental age. Thymocytes of recipients were analyzed by immunofluorescence flow cytometry: ov, TCR-γ/δ and ov, and TCR Vβ1. Each dot plot presents 50,000 events for the gated thymocyte population. Hatching occurs at E21.
Figure 3.

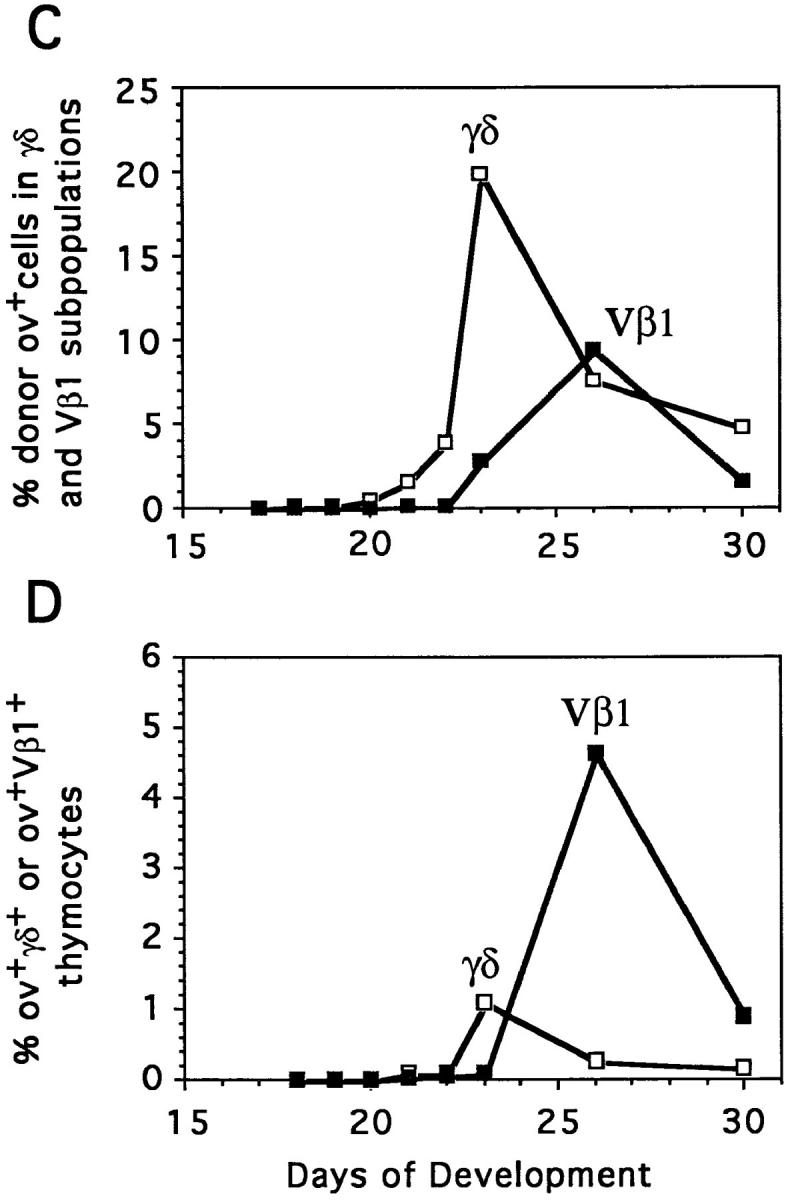
Comparative differentiation kinetics of the second and third waves of thymocyte progenitors: appearance of TCR-γ/δ+ and TCR-α/β (Vβ1)+ T cell progeny. (A and B) Proportion of thymocytes expressing TCR-γ/δ or TCR-α/β (Vβ1) among donor ov+ thymocytes was determined by immunofluorescence flow cytometry. Each point corresponds to the mean value for three to five animals in four independent experiments. The differentiation of second wave T cell progenitors was analyzed by adoptive transfer of E13 H.B19ov+ bone marrow into E13 H.B19ov− embryos, and for analysis of the third wave, E18 H.B19ov+ bone marrow cells were injected into E18 H.B19ov− embryos (injections of 2.0 × 107 cells). (C) Proportions of donor ov+ thymocytes that express TCR-γ/δ or TCR-α/β (Vβ1) during the differentiation of the second wave of progenitors. (D) Proportions of donor ov+ TCR-γ/δ+ and ov+ TCR-α/β (Vβ1)+ thymocytes derived from second wave thymocyte progenitors among the total thymocyte population in the recipient. The realistic efficiency of the γ/δ and Vβ1 T cell differentiation pathways followed by the progeny of the second wave of thymocyte progenitors can be estimated, since the areas under these curves are roughly proportional to the numbers of γ/δ and Vβ1 thymocytes produced by the donor second wave progenitors. This assessment suggests that the donor progenitors produced >15 times more α/β (Vβ1) than γ/δ thymocytes.
Mature γ/δ and α/β Thymocytes Migrate to the Periphery in Alternating Waves.
The colonization of the thymus in discrete waves (7, 8), and the differences in differentiation and emigration kinetics of γ/δ and α/β thymocytes suggest interspersed emigration of the mature γ/δ and α/β thymocyte subsets (11–13). To test this hypothesis, we examined the phenotype of recent thymocyte migrants at different developmental ages. In these experiments, thymocytes of chicks at 21 (hatching)–30 d were labeled in situ by intrathymic injection of FITC. Blood samples were obtained 12 h later and labeled lymphocytes in the circulation were analyzed for expression of TCR-γ/δ or TCR-α/β (Vβ1) (Fig. 4). The FITC-labeled cells represented 3–10% of the peripheral blood lymphocytes. Approximately 75% of these were γ/δ or α/β (Vβ1+) T cells; of the remaining 25% approximately half were α/β (Vβ2+) T cells and the rest were TCR−. Peaks of recent γ/δ thymocyte emigrants were detected on days 21–23 and 27–28, and a peak of recent α/β thymocyte emigrants was observed on days 24–26. The frequency of FITC-labeled γ/δ thymocyte emigrants reached a maximum of 20%, whereas the peak of labeled Vβ1 emigrants reached a maximum of 70%. These figures reflected the fact that each precursor wave gives rise to ∼5% γ/δ and 75% α/β (Vβ1+) thymocytes, respectively. Thymocyte progenitors in each colonization period thus gave rise to γ/δ T cell progeny within 9 d and α/β (Vβ1+) T cell progeny within 12 d, and these migrated in the same sequence to the periphery. Since each colonization period is followed by a refractory interval of ∼4 d, the end result is alternating emigrant waves of γ/δ and α/β (Vβ1) T cells, with minimal overlap of migrant cells representing each of the three embryonic waves of thymocyte progenitors.
Figure 4.

Phenotypic analysis of recent thymocyte emigrants in the periphery. Thymocytes were randomly labeled by intrathymic injection of fluorescein. Blood samples obtained 12 h later were analyzed by immunofluorescence flow cytometry. Around 3–10% of the peripheral blood lymphocytes were FITC labeled. TCR-γ/δ or TCR-α/β (Vβ1) were detected by phycoerythrin-coupled antibodies. Results are expressed in percent of fluorescein-labeled cells expressing either TCR-γ/δ and TCR-α/β Vβ1 cells, and each point corresponds to the mean value for four recipients. Hatched areas indicate the peak periods of splenic and intestinal colonization by γ/δ and α/β(Vβ1) T cells derived from the thymus (13, 21).
Each Thymocyte Progenitor Wave Contributes a Characteristic TCR-γ Repertoire.
The TCR-γ repertoire expressed by the thymocyte progeny of each progenitor wave was addressed in these experiments. Simple sorting of E13 thymocytes reactive with the anti–TCR-γ/δ antibody was sufficient to isolate first wave progeny for repertoire analysis. Since the progeny of successive waves may accumulate in the thymus during development, isolation of the second and the third wave progeny required the more complex strategy of adoptive transfer between the H.B19ov+/− congenic chicken strains. The progeny of second wave progenitors was isolated after the injection of E13 bone marrow cells from H.B19ov+ donors into age-matched H.B19ov− recipients. The ov+ TCR-γ/δ+ thymocytes (14,000 cells) were then sampled on day 22. To purify the progeny of the third progenitor wave, a similar adoptive transfer was made into E18 embryos, and ov+ TCR-γ/δ+ thymocytes in the recipients were sorted (1,400 cells) on day 28.
To examine the TCR-γ repertoire expressed in each developmental wave of thymocytes, we performed a PCR using a 3′ primer specific for Cγ and 5′ primers specific for Vγ1, Vγ2, or Vγ3 segments. First, we determined that Vγ1-Jγ-Cγ, Vγ2-Jγ-Cγ, and Vγ3-Jγ-Cγ transcripts were present as early as E10-11 in ov+ embryos, confirming in this congenic strain that the three different Vγ subfamilies undergo rearrangment simultaneously (Fig. 5 A). A similar analysis of the second and third wave progeny indicated that all three Vγ gene subfamilies also undergo rearrangement in these waves (Fig. 5 B). However, the Vγ subfamily representation differed among three waves. The TCR-γ repertoire of the second wave was composed mainly of Vγ2 transcripts, whereas transcripts containing the Vγ1 and Vγ3 gene segments were weakly represented. In contrast, Vγ1, Vγ2, and Vγ3 transcripts appeared to be equally well represented in the first and the third waves of thymus colonization (Fig. 5 B).
Figure 5.
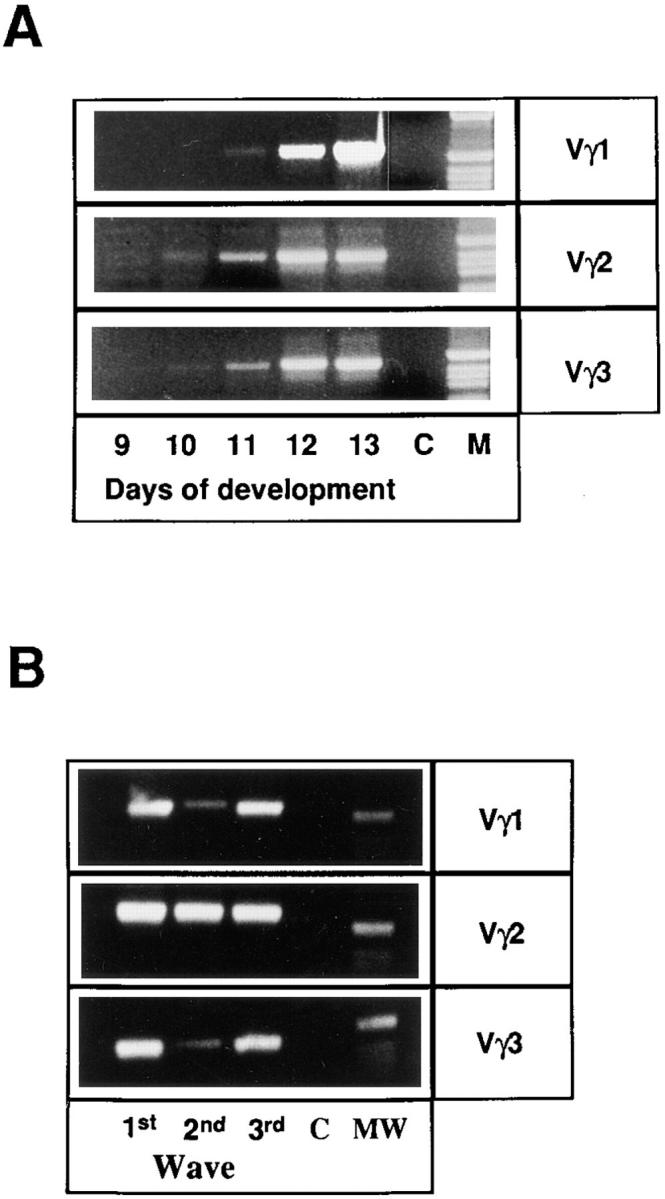
Ontogeny of TCR-γ transcripts. Identification of Vγ1-Jγ-Cγ, Vγ2-Jγ-Cγ, and Vγ3-Jγ-Cγ transcripts by PCR amplification. Actin expression was used to standardize cDNA amounts and to allow semiquantitative analysis. Templates were cDNA from thymus at various stages of development. After 32 cycles of amplification, PCR products were electrophoresed, stained with ethidiumbromide, and photographed. (A) Ontogeny of the different Vγ transcripts in chicken embryos. (B) Comparative analysis of the different Vγ transcripts in the three developmental waves of thymocytes. Thymocytes derived from the first, second, and third wave of precursors were from E13 embryos, 1-d-old chicks (day 22), and 1-wk-old chicks (day 28), respectively. First, second, and third wave thymocytes were obtained from multiple embryos.
A more detailed analysis of the representative TCR-γ repertoires was performed by cloning of the PCR products and sequencing ∼30 clones for each Vγ gene subfamily/ thymocyte wave (Figs. 6 and 7). In the H.B19ov+ chicken strain, six Vγ1, 12 Vγ2, and 10 Vγ3 members were detected. The Vγ2 subfamily was divided into eight Vγ2a members, three Vγ2b members, and a new member, Vγ2c, that differs significantly from the Vγ2a and Vγ2b subfamilies. In contrast to the differences in Vγ subfamily usage (Fig. 5 B), significant differences were not seen in the representation of members within a given Vγ subfamily for the three thymocyte waves (Fig. 7). Different members of the three Vγ families were found to rearrange with all three different Jγ segments. However, preferential pairings of Vγ and Jγ segments were observed. In each of developmental waves, Vγ2 segments were rearranged with Jγ2 segments (Table 1). In addition, variable Jγ usage patterns were seen in Vγ1- and Vγ3- containing transcripts for the three thymocyte waves. A high frequency of Vγ1/Jγ2 rearrangements in the first thymocyte wave was rearranged with the Jγ1 segment in the first wave and with the Jγ3 segment in the following waves.
Figure 6.

TCR Vγ segments observed in the H.B19ov+ chicken strain. Comparison of COOH-terminal amino acid sequences of the Vγ1, Vγ2, and Vγ3 subfamily members identified for the H.B19ov+ chicken strain by sequence analysis of PCR-derived cDNAs. Residues identical to the prototypic sequence are represented by dashes. Vγ segments are numbered according to Six et al. (16). §, Vγ1.1 and Vγ3.1 segment references were not encountered in our study; = new Vγ members. Some members differ by several nucleotides, but code for the same amino acid sequence. The Vγ2b2 member presents a stop codon (*) and might correspond to a pseudogene.
Figure 7.

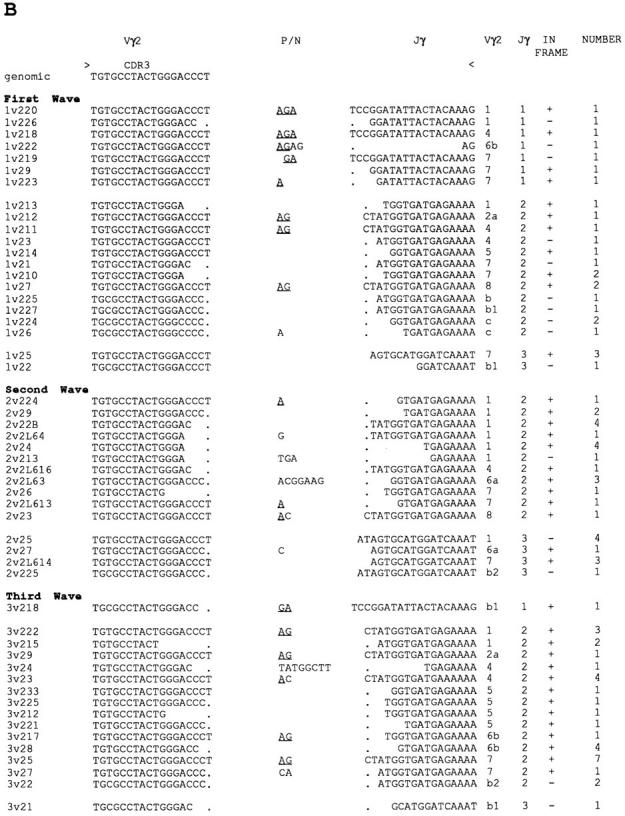
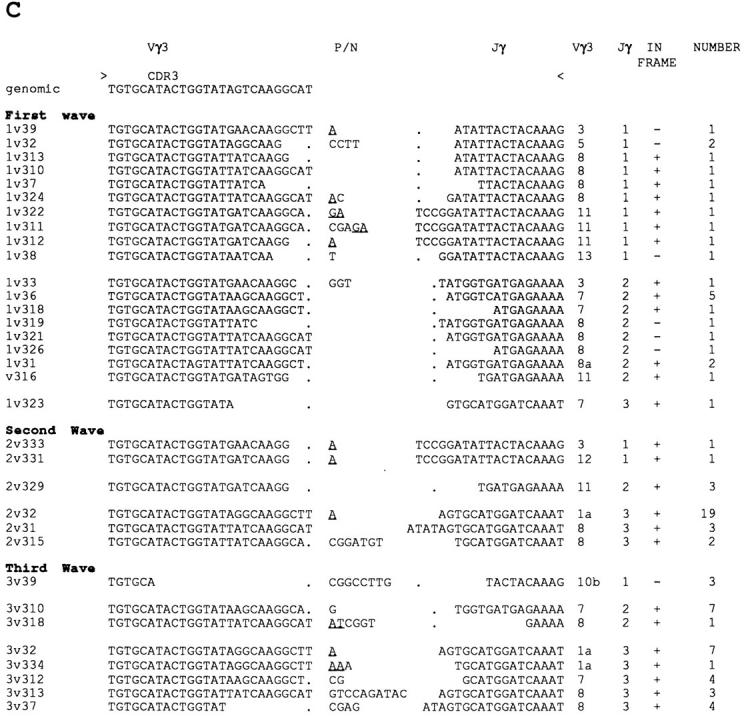
TCR-γ repertoires comparison for the three developmental waves of thymocyte precursors. Nucleotide sequences are shown for the Vγ–Jγ junctions. Vγ members, Jγ members, and the number of sequenced clones are indicated. The in frame clones are indicated by +. Assignment of nucleotides to the Vγ, Jγ1, and Jγ2 segments is based on germline sequences. Jγ3 region assignment is based on a consensus sequence from a TCR-γ cDNA clone containing the Jγ3 segment. Nucleotides that cannot be assigned to either Vγ or Jγ gene segments are indicated as nontemplate/palindromic additions. Putative P (palindromic) nucleotides are underlined. Thymocytes derived from the first, second, and third wave of precursors correspond to thymocytes from embryos (day 13), 1-d-old chicks (day 22), and 1-wk-old chicks (day 28), respectively. (A) TCR Vγ1 repertoires, (B) TCR Vγ2 repertoires, and (C) TCR Vγ3 repertoires.
Table 1.
Vγ Jγ Usage as a Function of Thymocyte Developmental Waves
| Jγ1 | Jγ2 | Jγ3 | ||||
|---|---|---|---|---|---|---|
| Vγ1 | ||||||
| First wave | 2 | 12 | 1 | |||
| Second wave | 1* | 0 | 0 | |||
| Third wave | 2* | 2 | 2 | |||
| Vγ2 | ||||||
| First wave | 4 | 6 | 1 | |||
| Second wave | 0 | 10 | 2 | |||
| Third wave | 1 | 13 | 0 | |||
| Vγ3 | ||||||
| First wave | 7 | 5 | 1 | |||
| Second wave | 2 | 1 | 3* | |||
| Third wave | 0 | 2 | 5* |
Results indicate the number of independent productively rearranged cDNA that corresponded to each type of Vγ Jγ rearrangement and presented for each Vγ subfamily.
The presence of recurrent clones.
A striking feature of the TCR-γ transcript analysis was the occurrence of recurrent clones exhibiting the same Vγ– Jγ junction in the second and third thymocyte waves (Fig. 7). (a) 30 identical Vγ1-Jγ1-Cγ clones (2v12) were found in the second thymocyte wave and 9 such clones (3v14) were found in the third thymocyte wave. (b) 19 identical Vγ3-Jγ3-Cγ clones were found in the second wave (2v32), and 7 clones of this type (3v32) were encountered in the third thymocyte wave. Thus, the low frequency of Vγ1 and Vγ3 subfamily usage in the second thymocyte wave was associated with an increased representation of repetitive clones.
Differences in CDR3 length were not observed in the repertoires generated by the three embryonic waves. However, an apparent increase in N/P nucleotide insertions at the Vγ-Jγ junction suggested an increase in CDR3 diversity in the third thymocyte wave (Table 2).
Table 2.
Analysis of CDR3
| Thymocyte wave | Vγ 3′ nucleotide deletions | Jγ 5′ nucleotide deletions | Junctional N/P nucleotides | |||
|---|---|---|---|---|---|---|
| First wave E13 | 1.87 | 2.63 | 1.13 | |||
| Second wave E22 | 1.90 | 3.28 | 1.19 | |||
| Third wave E28 | 2.07 | 3.80 | 2.26 |
Results correspond to means obtained in independent productive rearrangements. Only clones using Jγ1 and Jγ2, for which genomic borders were known (16), were taken into account for Jγ5′ nucleotide deletions and junctional N/P nucleotides.
DISCUSSION
Three discrete waves of thymocyte progenitors enter the embryonic chick thymus to generate three successive waves of T cell progeny members which leave the thymus to colonize peripheral organs such as the spleen and the intestine. The present studies indicate that each wave of progenitors gives rise first to γ/δ thymocytes and then α/β (Vβ1) thymocytes 3 or 4 d later so that the γ/δ and α/β (Vβ1) T cells migrate asynchronously from the thymus. Progenitor colonization of the thymus in waves and an accelerated rate of γ/δ T cell differentiation thus contribute to the alternating emigration of first γ/δ and then α/β T cells to the periphery (Fig. 8). Analysis of the TCR-γ repertoire for the different thymocyte waves suggests that they differ in Vγ and Jγ usage as well as in CDR3 diversity.
Figure 8.

Model for embryonic traffic of T lineage cells. Colonization of the embryonic thymus occurs in three waves (8). The present study indicates for all three waves the sequential generation of mature γ/δ T cells in ∼9 d, and mature α/β T cells in 12 d. Thymocyte emigration analysis, in agreement with the peaks of emigration period of thymocytes to the spleen and intestine reported previously (11–13), reveals that γ/δ and α/β T cells emigrate from thymus to periphery in alternating waves.
Thymus colonization by waves of hemopoietic progenitors also appears to occur in mammals (35, 36), but can be examined in greater detail in the chicken because of embryo accessibility and the opportunity to purify the progeny of individual waves of progenitors. Adoptive transfer of alloantigen-marked progenitors allowed us to elucidate the homing routes whereby these enter the embryonic thymus. This analysis indicates that progenitors of bone marrow origin enter the thymus either via interlobular venules or capillaries located at the corticomedullary junction. Both routes have been described in the mouse, but were not shown to be used simultaneously, the transcapsule route thought to be restricted to thymus colonization before its vascularization (37, 38). In the congenic chick chimeras, progenitors entering at the corticomedullary junction subsequently migrated to the outer cortex of the thymus, where precursors entering through the capsule were also found. This outer cortical homing pattern of thymocyte precursors has also been noted after direct needle injection in mice. As has been described in mammals (37), thymocytes then migrate from the outer cortex as they undergo T cell differentiation en route to the thymic medulla.
An interesting question concerns whether each embryonic wave of precursors generates the same or different T cell repertoires. Studies of the TCR-γ repertoire generated during mouse development indicate sequential usage of Vγ genes. The first γ/δ T cells generated during embryonic development express Vγ5-Cγ1 transcripts, the second population of γ/δ T cells express Vγ6-Cγ1 transcripts, and γ/δ T cells become more heterogeneous for Vγ usage after birth (22, 25). In similar fashion, the Vβ1 gene segments are rearranged before the Vβ2 gene segments during avian ontogeny (39). However, the Vγ gene families do not undergo sequential rearrangement during ontogeny. The chicken TCR-γ locus consists of 3 Vγ families with ∼10 members each, 3 Jγ segments, and 1 Cγ segments (15, 16). The first wave of thymocyte progeny rearrange all three Vγ families and Jγ genes as early as E10-E11. This type of unrestricted TCR-γ rearrangement pattern has also been suggested in sheep and humans (40, 41). On the other hand, preferential Vγ Jγ pairings were observed for the three developmental waves of thymocytes, whereas preferred TCR Vβ1/D/Jβ rearrangements were not apparent during ontogeny (13). The Vγ Jγ junctional variations (CDR3) for all three embryonic thymocyte waves were more limited than in the adult (16). Such differences between embryonic and adult repertoires have also been found in mammals (42). Finally, the nonproductively rearranged TCR-γ transcripts observed in TCR-γ/δ+ov+ thymocytes indicate that Vγ Jγ rearrangements occur on both alleles in avian γ/δ T cells.
A high frequency of repetitive TCR-γ transcripts was found in the second and third waves of thymocyte progeny, particularly in the second wave of thymocytes where two clones, 2v12 and 2v32, represented 97 and 66% of the Vγ1 and Vγ3 repertoires, respectively. This result could represent a PCR artefact or a sampling error due to the limited numbers of cells being analyzed. However, several observations suggest that the TCR-γ repertoire may differ for the different waves. (a) The Vγ1 and Vγ3 repertoire diversities were higher in the third wave than in second, even though the number of γ/δ+ donor-derived thymocytes examined in the second wave was ten times higher than for the third wave (14 × 103 versus 1.4 × 103 γ/δ cells). (b) Repetitious clones were also encountered in the third wave repertoire, albeit at lower frequencies (31% for 2v12-3v14, 23% for 2v32-3v32). (c) The cDNA synthesis, PCR amplification, and product cloning procedures were repeated three times to confirm the findings. (d) Repetitive transcripts were not encountered in the Vγ2 repertoire preferentially used by the second thymocyte wave. Thus, the high frequency of repetitive invariant clones in Vγ1 and Vγ3 repertoires created by cells of the second wave is in agreement with a lower usage of Vγ1 and Vγ3 families in this wave.
Differences in the TCR-γ repertoire generated in each progenitor wave could reflect differences in the colonizing progenitors. The first wave of thymocyte progenitor are derived from multipotent hematopoietic stem cells that arose in the paraaortic region of the embryo (3), whereas the second and third wave of thymocyte progenitors were derived from the bone marrow. The generation of different TCR-γ repertoires could therefore reflect differences in the progenitor cells themselves. The differentiation kinetics of γ/δ and α/β (Vβ1+) thymocytes were conserved for the three developmental waves of thymocyte progenitors. The times required for differentiation of γ/δ and α/β (Vβ1) thymocytes were ∼9 and 12 d, respectively, in the chicken. The accelerated differentiation of avian γ/δ versus α/β thymocytes has also been noted for mouse thymocyte precursors (43, 44). Different selection processes for the α/β and the γ/δ thymocytes may contribute to the differences in differentiation kinetics (45, 46). Different growth requirements during γ/δ and α/β T cell differentiation, such as IL-7 requirement, may also promote different differentiation kinetics (47).
The thymocyte emigration model whereby peripheral tissues are colonized by γ/δ T cells before α/β (Vβ1+) T cells may favor harmonious development of a strategic immune defense system mediated by interacting lymphocyte subpopulations. The interaction of γ/δ T cells with cells in the peripheral organs could provoke microenvironmental modification that favors the homing of α/β T cells. The location of γ/δ and α/β T cells in peripheral tissues may play an important role in the establishment of an immune response, in that recent evidence suggests γ/δ T cells may function as regulatory cells for α/β T cells (48–50). During organogenesis, new niches for lymphocyte homing may appear throughout development. The interspersed emigration of γ/δ and Vβ1 thymocytes might provide a mechanism to fill these niches by γ/δ and α/β cells in developing organs. The observed interspersed emigration of γ/δ and α/β (Vβ1+) thymocytes could also affect the homing patterns of specific thymocyte subpopulations. For example, two thymocyte subpopulations are emigrating from the thymus at day 21–23, the γ/δ thymocytes generated by the second wave of precursors and the minor second subpopulation of α/β (Vb2+) T cells generated by the first wave of progenitors (8). Interestingly, γ/δ thymocytes colonize the intestine in massive numbers, whereas Vβ2 thymocytes are rarely seen in that organ (20). These two populations may compete for homing sites, thus diverting the Vβ2 population to other organs. Since an optimal immune response may require collaboration between γ/δ and α/β T cells, sequential migration of γ/δ and α/β thymocyte subpopulations may provide an efficient means to maintain a physiological balance between the two cell populations during development.
Acknowledgments
The authors thank Mark Dessing, Victor Hasler, Barbara Ecabert, and Birgit Kugelberg for excellent technical assitance; Jean Desrosiers, Hans Spalinger, and Beatrice Pfeiffer for photography; and Drs. Klaus Karjalainen, Hans Reimer Rodewald, and Charley Steinberg for critical reading and improvement of the manuscript.
This work was supported by the Association pour la Recherche contre le Cancer, Association pour la Recherche contre le Cancer (ARC), the Human Frontier Science Programme Organization, Human Fronten Science Programme Organisation (HFSP), and the Academy of Finland. The Basel Institute for Immunology was founded and is supported by F. Hoffmann-La Roche (Basel, Switzerland).
Abbreviation used in this paper
- E
day of embryonic life
REFERENCES
- 1.Cooper M, Peterson D, Good R. Delineation of the thymic and bursal lymphoid system in the chicken. Nature (Lond) 1965;205:143–146. doi: 10.1038/205143a0. [DOI] [PubMed] [Google Scholar]
- 2.Moore MAS, Owen JJT. Experimental studies on the development of the thymus. J Exp Med. 1967;126:715–726. doi: 10.1084/jem.126.4.715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Dieterlen-Lièvre F, Godin I, Pardanaud L. Ontogeny of hematopoiesis in the avian embryo: a general paradigm. Curr Top Microbiol Immunol. 1996;212:119–128. doi: 10.1007/978-3-642-80057-3_11. [DOI] [PubMed] [Google Scholar]
- 4.Jotereau FV, Houssaint E, Le Douarin NM. Lymphoid stem cell homing to the early thymic primordium of the avian embryo. Eur J Immunol. 1980;10:620–627. doi: 10.1002/eji.1830100809. [DOI] [PubMed] [Google Scholar]
- 5.Jotereau FV, Le Douarin NM. Demonstration of a cyclic renewal of the lymphocyte precursor cells in the quail thymus during embryonic and perinatal life. J Immunol. 1982;129:1869–1877. [PubMed] [Google Scholar]
- 6.Le Douarin, N.M., F.V. Jotereau, E. Houssaint, and J.P. Thiery. 1984. Primary lymphoid organ ontogeny in birds. In Chimeras in Developmental Biology. N.M. Le Douarin and A. McLaren, editors. Academic Press, London. 179–216.
- 7.Coltey M, Jotereau FV, Le Douarin NM. Evidence for a cyclic renewal of lymphocyte precursor cells in the embryonic chick thymus. Cell Differ. 1987;22:71–82. doi: 10.1016/0045-6039(87)90414-3. [DOI] [PubMed] [Google Scholar]
- 8.Coltey M, Bucy RP, Chen CH, Cihak J, Lösch U, Char D, Le Douarin NM, Cooper MD. Analysis of the first two waves of thymus homing stem cells and their T cell progeny in chick–quail chimeras. J Exp Med. 1989;170:543–557. doi: 10.1084/jem.170.2.543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dunon D, Kaufman J, Salomonsen J, Skjoedt K, Vainio O, Thiery JP, Imhof BA. T cell precursor migration towards β2-microglobulin is involved in thymus colonization of chicken embryos. EMBO (Eur Mol Biol Organ) J. 1990;9:3315–3322. doi: 10.1002/j.1460-2075.1990.tb07531.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Vainio O, Dunon D, Aïssi F, Dangy JP, McNagny KM, Imhof BA. HEMCAM, an adhesion molecule expressed by c-kit+ hemopoietic progenitors. J Cell Biol. 1996;135:1655–1668. doi: 10.1083/jcb.135.6.1655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dunon D, Cooper M, Imhof B. Thymic origin of embryonic intestinal γ/δ T cells. J Exp Med. 1993;177:257–263. doi: 10.1084/jem.177.2.257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Dunon D, Cooper MD, Imhof BA. Migration patterns of thymus derived γδ T cells during chicken development. Eur J Immunol. 1993;23:2545–2550. doi: 10.1002/eji.1830231025. [DOI] [PubMed] [Google Scholar]
- 13.Dunon D, Schwager J, Dangy JP, Cooper MD, Imhof BA. T cell migration during development: homing is not related to TCR Vβ1 repertoire selection. EMBO (Eur Mol Biol Organ) J. 1994;13:808–815. doi: 10.1002/j.1460-2075.1994.tb06323.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chen CH, Six A, Kubota T, Tsuji S, Kong FK, Göbel TWF, Cooper MD. T cell receptors and T cell development. Curr Top Microbiol Immunol. 1996;212:37–53. doi: 10.1007/978-3-642-80057-3_5. [DOI] [PubMed] [Google Scholar]
- 15.Rast J, Haire R, Litman R, Pross S, Litman G. Identification and characterization of T-cell antigen receptor–related genes in phylogenetically diverse vertebrate species. Immunogenetics. 1995;42:204–212. doi: 10.1007/BF00191226. [DOI] [PubMed] [Google Scholar]
- 16.Six A, Rast J, McCormack W, Dunon D, Courtois D, Li Y, Chen C, Cooper M. Characterization of avian T cell receptor γ genes. Proc Natl Acad Sci USA. 1996;93:15329–15334. doi: 10.1073/pnas.93.26.15329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sowder JT, Chen CH, Ager LL, Chan MM, Cooper MD. A large subpopulation of avian T cells express a homologue of the mammalian T γ/δ receptor. J Exp Med. 1988;167:315–321. doi: 10.1084/jem.167.2.315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bucy RP, Chen CL, Cooper MD. Ontogeny of T cell receptors in the chicken thymus. J Immunol. 1990;144:1161–1168. [PubMed] [Google Scholar]
- 19.Chen CH, Cihak J, Lösch U, Cooper MD. Differential expression of two T cell receptors, TcR1 and TcR2, on chicken lymphocytes. Eur J Immunol. 1988;18:539–543. doi: 10.1002/eji.1830180408. [DOI] [PubMed] [Google Scholar]
- 20.Char D, Sanchez P, Chen C, Bucy R, Cooper M. A third sublineage of avian T cells can be identified with a T cell receptor–3–specific antibody. J Immunol. 1990;145:3547–3555. [PubMed] [Google Scholar]
- 21.Dunon D, Imhof BA. T cell migration during ontogeny and T cell repertoire generation. Curr Top Microbiol Immunol. 1996;212:79–93. doi: 10.1007/978-3-642-80057-3_8. [DOI] [PubMed] [Google Scholar]
- 22.Haas W, Tonegawa S. Development and selection of γδ T cells. Curr Opin Immunol. 1992;4:147–155. doi: 10.1016/0952-7915(92)90004-x. [DOI] [PubMed] [Google Scholar]
- 23.Ikuta K, Kina T, MacNeil I, Uchida N, Peault B, Chien Y, Weissman I. A developmental switch in thymic lymphocyte maturation potential occurs at the level of hematopoietic stem cells. Cell. 1990;62:863–874. doi: 10.1016/0092-8674(90)90262-d. [DOI] [PubMed] [Google Scholar]
- 24.Ikuta K, Weissman I. The junctional modifications of a T cell receptor γ chain are determined at the level of thymic precursors. J Exp Med. 1991;174:1279–1282. doi: 10.1084/jem.174.5.1279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ikuta K, Uchida N, Friedman J, Weissman I. Lymphocyte development from stem cells. Annu Rev Immunol. 1992;10:759–783. doi: 10.1146/annurev.iy.10.040192.003551. [DOI] [PubMed] [Google Scholar]
- 26.Vainio, O., T.V. Veromaa, E. Eerola, and P. Toivanen. 1987. Characterization of two monoclonal antibodies against chicken T lymphocytes surface antigen. In Avian Immunology. W.T. Weber and D.L. Ewert, editors. Alan R. Liss Inc., New York. 99–108.
- 27.Houssaint E, Mansikka A, Vainio O. Early separation of B and T lymphocyte precursors in chick embryo. J Exp Med. 1991;174:397–406. doi: 10.1084/jem.174.2.397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cihak J, Ziegler-Heitbrock HWS, Trainer H, Schranner I, Merkenschlager M, Lösch U. Characterization and functional properties of a novel monoclonal antibody which identifies a T cell receptor in chickens. Eur J Immunol. 1988;18:533–537. doi: 10.1002/eji.1830180407. [DOI] [PubMed] [Google Scholar]
- 29.Dunon, D., O. Vainio, and B.A. Imhof. 1997. Lymphocyte migration in vivo: the chicken embryo model. In Immunology Manual Methods. I. Lefkovits, editor. Academic Press Ltd., London. 1345–1353.
- 30.Sambrook, J., E.F. Fritsch, and T. Maniatis. 1989. Molecular cloning. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
- 31.Keller G, Kennedy M, Papayannopoulo T, Wiles M. Hematopoietic commitment during embryonic stem cell differentiation in culture. Mol Cell Biol. 1993;13:473–486. doi: 10.1128/mcb.13.1.473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kost T, Theodorakis N, Hughes S. The nucleotide sequence of the chick cytoplasmic beta-actin gene. Nucleic Acids Res. 1983;11:8287–8301. doi: 10.1093/nar/11.23.8287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chen E, Seeburg P. Supercoil sequencing: a fast and simple method for sequencing plasmid DNA. DNA (NY) 1985;4:165–170. doi: 10.1089/dna.1985.4.165. [DOI] [PubMed] [Google Scholar]
- 34.Sanger F, Nicklen S, Coulson A. DNA sequencing with chain terminating inhibitors. Proc Natl Acad Sci USA. 1977;74:5463–5467. doi: 10.1073/pnas.74.12.5463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Jotereau F, Heuze F, Salomon-Vie V, Gascan H. Cell kinetics in the fetal mouse thymus: precursor cell input, proliferation, and emigration. J Immunol. 1987;138:1026–1030. [PubMed] [Google Scholar]
- 36.Morin C, Jotereau F, Augustin A. Patterns of responsiveness of T cell lines and thymocytes reveal waves of specific activity in the post-natal thymus. Int Immunol. 1992;4:1091–1101. doi: 10.1093/intimm/4.10.1091. [DOI] [PubMed] [Google Scholar]
- 37.Kyewski BA. Seeding of thymic microenvironments defined by distinct thymocyte–stromal interactions is developmentally controlled. J Exp Med. 1987;166:520–538. doi: 10.1084/jem.166.2.520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.van Ewijk W. T-cell differentiation is influenced by thymic microenvironments. Annu Rev Immunol. 1991;9:591–615. doi: 10.1146/annurev.iy.09.040191.003111. [DOI] [PubMed] [Google Scholar]
- 39.Pickel JM, McCormack WT, Chen CH, Cooper MD, Thompson CB. Differential regulation of V(D)J recombination during development of avian B and T cells. Int Immunol. 1993;5:919–927. doi: 10.1093/intimm/5.8.919. [DOI] [PubMed] [Google Scholar]
- 40.Hein W, Dudler L. Divergent evolution of T cell repertoires: extensive diversity and developmentally regulated expression of the sheep γδ T cell receptor. EMBO (Eur Mol Biol Organ) J. 1993;12:715–724. doi: 10.1002/j.1460-2075.1993.tb05705.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.McVay L, Carding S, Bottomly K, Hayday A. Regulated expression and structure of T cell receptor γ/δ transcripts during human ontogeny. EMBO (Eur Mol Biol Organ) J. 1991;10:83–91. doi: 10.1002/j.1460-2075.1991.tb07923.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bogue M, Candéias S, Benoist C, Mathis D. A special neo-repertoire in neonatal mice. EMBO (Eur Mol Biol Organ) J. 1991;10:3647–3654. doi: 10.1002/j.1460-2075.1991.tb04931.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Petrie H, Scollay R, Shortman K. Commitment to the T-cell receptor α/β or γ/δ lineages can occur just prior to the onset of CD4 and CD8 expression among immature thymocytes. Eur J Immunol. 1992;22:2185–2188. doi: 10.1002/eji.1830220836. [DOI] [PubMed] [Google Scholar]
- 44.Wu L, Scollay R, Egerton M, Pearse M, Spangrude G, Shortman K. CD4 expresssed on earliest T-lineage precursor cells in the adult murine thymus. Nature (Lond) 1991;349:71–74. doi: 10.1038/349071a0. [DOI] [PubMed] [Google Scholar]
- 45.Cooper M, Chen C, Bucy R, Thompson C. Avian T cell ontogeny. Adv Immunol. 1991;50:87–117. doi: 10.1016/s0065-2776(08)60823-8. [DOI] [PubMed] [Google Scholar]
- 46.Schweighoffer E, Fowlkes BJ. Positive selection is not required for thymic maturation of transgenic γ/δ T cells. J Exp Med. 1996;183:2033–2041. doi: 10.1084/jem.183.5.2033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Maki K, Sunaga S, Komagata Y, Kodaira Y, Mabuchi A, Karasuyama H, Yokomuro K, Miyazaki J, Ikuta K. Interleukin receptor–deficient mice lack γ/δ T cells. Proc Natl Acad Sci USA. 1996;93:7172–7177. doi: 10.1073/pnas.93.14.7172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kasahara Y, Chen CH, Cooper MD. Growth requirements for avian γ/δ T cells include exogenous cytokines, receptor ligation and in vivo priming. Eur J Immunol. 1993;23:2230–2236. doi: 10.1002/eji.1830230927. [DOI] [PubMed] [Google Scholar]
- 49.Raulet DH. How γ/δ T cells make a living. Curr Biol. 1994;4:246–248. doi: 10.1016/s0960-9822(00)00055-5. [DOI] [PubMed] [Google Scholar]
- 50.Tsuji S, Char D, Bucy RP, Simonsen M, Chen CH, Cooper MD. γ/δ T cells are secondary participants in acute graft-versus-host reactions initiated by CD4+α/β T cells. Eur J Immunol. 1996;26:420–427. doi: 10.1002/eji.1830260223. [DOI] [PubMed] [Google Scholar]