

Abstract
T cell maturation requires the rearrangement of clonotypic T cell receptors (TCR) capable of interacting with major histocompatibility complex (MHC) ligands to initiate positive and negative selection. Only 3–5% of thymocytes mature to join the peripheral T cell pool. To investigate the basis for this low success rate, we have measured the frequency of preselection thymocytes capable of responding to MHC. As many as one in five MHC-naive thymocytes show upregulation of activation markers on exposure to MHC-expressing thymic stroma in short-term reaggregate culture. The majority of these cells display physiological changes consistent with entry into the selection process within 24 h. By exposing TCR transgenic thymocytes to a range of MHC–peptide complexes, we show that CD69 induction is indicative of thymocyte selection, positive or negative. Our data provide evidence that the fraction of thymocytes that qualify to enter the thymic selection process far exceeds the fraction that successfully complete it, and suggest that most MHC-reactive thymocytes are actively eliminated in the course of selection.
The realization more than two decades ago that T cells recognize antigen in the context of MHC molecules (1, 2) raised the issue of how efficient cooperation between TCR and MHC molecules is achieved during T cell development. This task is considerable since MHC alleles are highly polymorphic, and not genetically linked to TCR loci. The latter undergo somatic recombination involving essentially random use of multiple germline-encoded gene segments (combinatorial diversity) and deliberate DNA template-independent variability (3–5). These mechanisms, which ensure sufficient TCR diversity, might be predicted to generate only very few cells with receptors competent to productively engage MHC–peptide complexes. Indeed, the vast majority (95–97%) of CD4 CD8 double-positive (DP)1 thymocytes are thought to die in situ (6), and only a small minority of 3–5% thymocytes reach the single-positive (SP) stage (7, 8). However, these figures do not reveal how many thymocytes are initially recruited into the selection process by productive interactions with MHC–peptide complexes. Further, it is not known how many of the initially recruited thymocytes fail to complete their development, be it as a result of negative selection or due to other factors, e.g., the availability of stromal cell environments able to support maturation (8–10), changing activation requirements during the DP to SP transition, (11, 12), or wastage during attempts to match TCR specificity and coreceptor expression (13).
The impact of thymocyte deletion on mature T cell production has been studied in mice with an impaired ability to negatively select. Surh and Sprent (6) analyzed mice that lack MHC molecules (and consequently cannot negatively select) by histological analysis of thymocyte death in situ. They found as many dying thymocytes as in wild-type controls and concluded that negative selection has no measurable impact on thymocyte death, and that the vast majority of thymocytes that die do so because they are not selected at all (6). Recently, Laufer et al. (14) produced mice in which MHC class II molecules were expressed selectively by cortical epithelium, resulting in CD4 T cell maturation in the absence of class II–dependent selection by bone marrow–derived cells. 5% of CD4 T cells generated in these mice responded to syngeneic class II–expressing cells, suggesting that in normal mice, negative selection removes a minimum of 5% of the selected repertoire (∼1/ 500 DP thymocytes, assuming a normal DP to SP conversion rate of 3–5%). Van Meerwijk et al. (15) suggested that the impact of negative selection was in fact 10-fold greater than the estimate by Laufer et al. (14), based on the production of CD4 SP thymocytes in radiation bone marrow chimeras. CD4 SP thymocytes were twice as frequent in chimeras expressing MHC class II on host tissues (including thymic epithelial cells) but lacking class II on bone marrow–derived cells compared to control chimeras expressing class II on both host and graft tissues. The authors attributed this difference to negative selection by bone marrow– derived cells and suggested that in normal mice at least 50% of the initially selected repertoire is subject to deletion (15).
Early suggestions that TCRs might have an intrinsic affinity for MHC (16) were fuelled by the high frequencies with which mature, postselection T cells respond to allogeneic MHC molecules (i.e., MHC molecules they were not selected on; see review in reference 17). The idea was supported by transfection experiments in which combinations of unrelated TCR α and β chains were able to confer allo-recognition (18). Subsequent studies analyzed the repertoire of CD4 T cells selected on a very limited set of MHC–peptide ligands (19–23), or generated in the absence of MHC molecules (24). Invariably, the resulting T cells responded strongly to allogeneic MHC class II, and also to normal, syngeneic class II–peptide complexes. In one study, as many as two thirds of T cell hybridomas generated from CD4 cells selected on a single MHC–peptide complex responded to the same MHC molecule displaying the normal spectrum of associated peptides, providing an estimate of how many thymocytes might have been deleted had the selection process occurred on the normal spectrum of MHC–peptide complexes (22). Based on the assumption that TCR–MHC-peptide interactions which mediate mature T cell activation would delete DP thymocytes, such extrapolations have important limitations. The frequency of cells subject to negative selection may be underestimated because MHC–peptide ligands that delete DP cells do not always activate mature T cells (e.g., reference 25). Vice versa, MHC–peptide ligands that activate mature T cells may not necessarily delete DP thymocytes (e.g., reference 26). Perhaps the most serious problem with using mature T cell reactivity as the basis to estimate TCR–MHC-peptide interactions relevant to thymocytes is that the requirements for positive selection and mature T cell activation are almost invariably different (11, 26–28) and mature T cells do not respond by full activation to the ligands by which they were selected (see review in reference 11). TCR–MHC-peptide interactions that drive positive selection can therefore not be reliably inferred from mature T cell reactivity.
In this report, we present a more direct approach to address this problem. In particular, we determine how many preselection thymocytes are capable of productive interaction with MHC ligands. The frequencies we observe are high and suggest that the low success rate of thymocyte maturation cannot simply be due to a low frequency of cells capable of participating in the selection process, but rather reflects other factors actively limiting thymic T cell production.
MATERIALS AND METHODS
Thymus Reaggregate and Activation Cultures.
Thymic stroma was prepared by trypsinization of deoxyguanosine-treated fetal thymi (29), and centrifugation over 55% Percoll (Pharmacia, Piscataway, NJ). In some experiments, residual CD45-expressing cells were removed using immunomagnetic beads (Dynal, Oslo, Norway). Reaggregates were established from 2 × 105 stromal cells and 4 × 105 thymocytes, and cultured as described (10). In contrast to earlier studies (e.g., references 29 and 30), antibodies to MHC products or to CD3 were not used in the preparation of stromal cells and thymocytes to avoid interference with TCR recognition and alterations in thymocyte behavior (31, 32).
To assess CD69 induction by TCR–MHC-peptide interactions of known affinity, OT-I TCR transgenic/recombination-activating gene RAG-1°/β2 microglobulin (β2m)° thymi were cut into small fragments and cultured in the presence of 5 μg/ml human β2m and the indicated peptides (20 μM) as described (28).
Flow Cytometry.
Cells were counted by phase-contrast microscopy and stained as described (10) using CD4-PE, -FITC (both Caltag Labs., South San Francisco, CA) or –red 613 (GIBCO BRL, Gaithersburg, MD), CD8a-red 613 (GIBCO BRL) or -FITC (Caltag Labs.), CD69-biotin streptavidin-FITC, CD5-FITC, CD24 (HSA)-FITC (all from PharMingen, San Diego, CA), or anti-HY TCR (T3.70) and goat anti–mouse IgG-PE (Caltag Labs.).
To quantitate DP thymocytes activated by MHC encounter in vivo, thymocytes from young adult MHC° and class I+ mice were stained with CD4–red 613, CD5-FITC, and CD69-biotin followed by streptavidin (SA)-PE. Thymocytes from class II+ mice were stained with CD8-red 613, CD5-FITC, and CD69-biotin–SA-PE.
DiOC6 or hydroethidine (Molecular Probes Inc., Eugene, OR) were used in combination with either CD4 or CD8 and CD5 or CD69 as markers for cells entering apoptosis (33). For population turnover studies, thymocytes labeled with PKH26 (Sigma Chemical Co., St. Louis, MO) according to the distributor's instructions were cultured and stained for CD4 and CD8 or activation markers. PKH26 was detected in the FL-2 channel. Samples were analyzed by FACScan® (Becton Dickinson, Mountain View, CA), excluding dead cells by light scatter and uptake of propidium iodide (3 μg/ml).
Reverse Transcriptase PCR.
Thymocytes recovered from reaggregate cultures were stained and sorted according to light scatter and activation markers on a FACSort® modified to allow sample cooling. RNA from 5 × 104 to 2 × 105 cells was reverse transcribed (SuperscriptII; GIBCO BRL) and amplified by PCR (SuperTaq; HT Biotechnology, Cambridge, UK) for 33 cycles, 60°C annealing temperature, with the primers RAG-1/32 CCAAGCTGCAGACATTCTAGCACTC; RAG-1/593 CAACATCTGCCTTCACGTCGATCC; RAG-2/91 CACATCCACAAGCAGGAAGTACAC; RAG-2/562 GGTTCAGGGACATCTCCTACTAAG; hypoxanthine guanine phosphoribosyl transferase (HPRT)/41 GGCTTCCTCCTCAGACCGCTTT; HPR T/742 AGGCTTTGTATTTGGCTTTTCC; CD4-5′ AACTGGTTCGGCATGACACTCTC; CD4-3′ CAGGGGCCACCACTTGAACTAC; CD8-5′ CCCCCACCCAGAGACCCAGAAG; CD8-3′ TCCAAGGCCCAGTCCAAGAAGAGT.
Functional Studies.
Thymi from newborn MHC° or MHC class II+ mice were cut in four and cultured where indicated with 1–2 μg/ml of a bispecific anti-CD3/CD4 reagent incorporating F(ab′) engineered to heterodimerise via fos/jun leucine zippers (34). As described in detail elsewhere (35), this treatment generates phenotypically mature CD4 SP thymocytes in the absence of MHC molecules. Cell suspensions were prepared 96 h later and cultured overnight at 2 × 106 cells/ml to deplete adherent stroma. Limiting dilution assays were done as described (36) with mitomycin C–treated CBA/Ca and MHC° splenocytes as stimulators and controls, respectively. T cell blasts prepared by stimulation of thymocytes with plate-bound TCR antibody (H57, PharMingen), 3 ng/ml PMA, and 20 U/ml IL-2 were fused to TCR-deficient BW5147 thymoma cells as described (37), and those expressing TCR and CD4 were tested for reactivity against MHC class II–transfected L cells in a standard CTLL2 assay (37). A stimulation index >10 over control L cell transfectants in at least two experiments was taken to indicate MHC reactivity.
RESULTS
Visualization of Preselection Thymocyte Responses to MHC.
We examined the MHC reactivity of preselection thymocytes using MHC-naive thymocytes from MHC-deficient (MHC°) mice (13). In these experiments, MHC° CD4+CD8+ thymocytes were exposed to MHC+ thymic stroma in reaggregate culture (10, 29) under conditions that rescue the development of mature, SP thymocytes (Fig. 1, day 5). The frequency of MHC-responsive cells was monitored using the activation marker CD69 (4, 37a, 38). Contrary to our initial expectations, a substantial fraction (15%) of MHC° thymocytes expressed CD69 24 h after exposure to MHC+ (H-2b) thymic stroma, at a time when the number of cells recovered was close to input (see below). This value, when compared to 0.8% and 0.2% CD69-positive cells seen in reaggregates with MHC° stroma or MHC° thymocytes ex vivo (Fig. 1, top) suggests that a considerable proportion of MHC-naive thymocytes are potentially MHC responsive.
Figure 1.
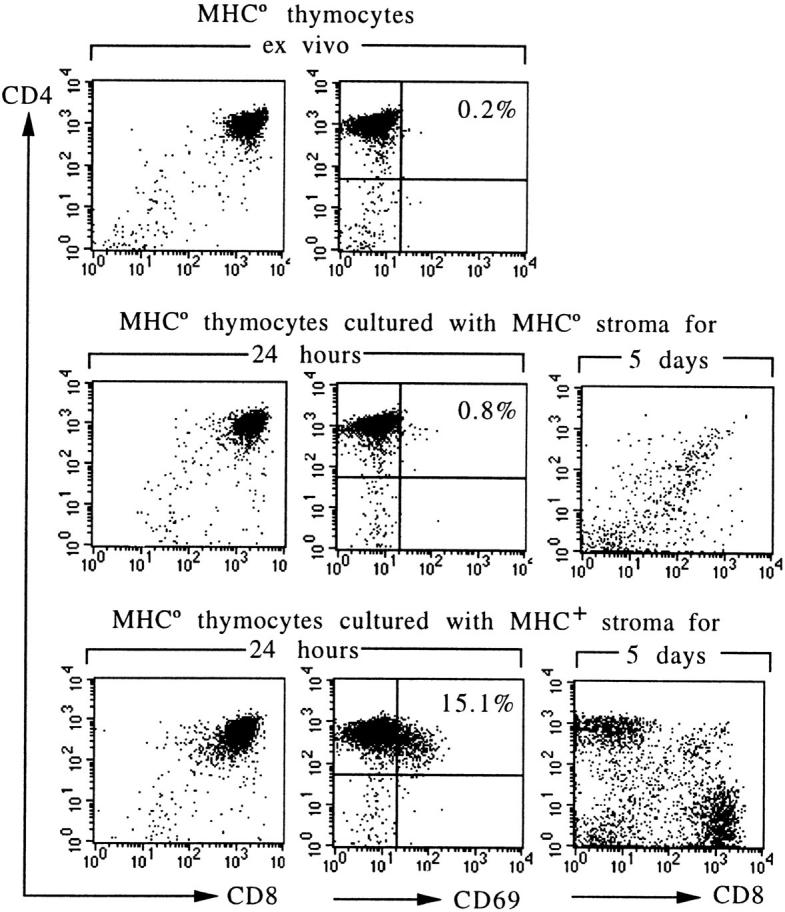
MHC-dependent activation and developmental rescue of MHC-naive thymocytes. Thymocytes from MHC° mice are arrested at the CD4+CD8+ stage and lack selection-associated activation markers. Coculture with MHC+ (H-2b) but not with MHC° thymic stroma results in CD69 expression, and rescues the development of CD4 and CD8 single-positive HSAlow cells (5-d displays are gated on HSAlow cells).
Defining the Requirements for CD69 Induction.
To determine whether the CD69 upregulation measured in these experiments is likely to reflect biologically relevant interactions, we took advantage of a well-characterized TCR transgenic system. The OT-I TCR (28), specific for an ovalbumin-derived peptide in the context of Kb, was crossed onto a recombination-deficient (Rag-1°; reference 39) and nonselecting background (β2m°; reference 40). Relevant MHC–peptide complexes can be restored by the addition in thymic organ culture of exogenous Kb-binding peptides and β2m (28). Since both the biological effects of the resulting Kb–peptide complexes on thymocyte maturation and mature T cell activation (4, 41–43) and their ability to bind the OT-I TCR (44) are extensively documented, we sought to compare CD69 induction with these parameters. The agonist, negatively selecting peptide SIINFEKL caused strong CD69 expression by many of the remaining cells (Fig. 2). Similarly, CD69 was strongly induced by the E1 analogue, a peptide that mediates positive (and in our tests, also some negative) selection of OT-I transgenic thymocytes, and is an antagonist/partial agonist for mature OT-I transgenic T cells. The Kd (off/on) values of OT-I/Kb–SIINFEKL and OT-I/Kb–E1 interactions are 6.5 and 22.6 μM, respectively (44). Other positively selecting antagonist peptides, including V-OVA (Kd 29.8 μM) and R4 (not shown; Kd 57.1 μM) also stimulated substantial CD69 expression (but no apparent deletion). Moderate expression of CD69 was induced by a CPα1-derived peptide, a candidate endogenous ligand for positive selection of OT-I thymocytes (41). In contrast, K4, an analogue peptide that despite a measurable affinity for the OT-I TCR (Kd OT-I/Kb–K4 ∼360 μM; reference 44) fails to drive OT-I transgenic thymocyte maturation did not induce CD69 upregulation in our assay, and neither did a Kb-binding control peptide, vesicular stomatitis virus (VSV). Thus, in this experimental system we find a very strong correlation between biologically relevant TCR–MHC-peptide interactions and CD69 upregulation. Interactions that drive thymocyte positive or negative selection resulted in CD69 upregulation, whereas interactions without effect on thymocyte maturation did not.
Figure 2.
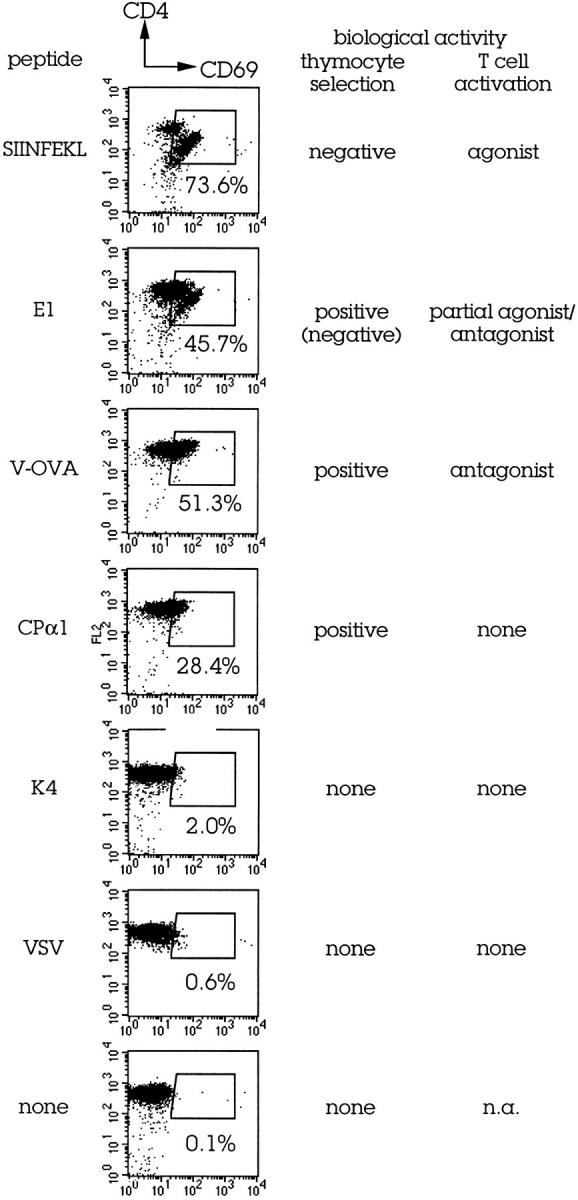
CD69 expression by CD4 CD8 DP thymocytes is triggered by biologically relevant TCR-MHC–peptide contact. Young adult OT-I TCR transgenic/RAG-1°/β2m° thymi were cut into fragments and cultured with or without exogenous peptides (20 μM) and exogenous β2m to allow the reconstitution of Kb–peptide complexes as described (28). CD69 expression was determined 24 h later. Data on thymocyte selection and mature T cell activation are from references 28, 34–36. Kd (off/on) values are listed in the text.
Additive Responses to MHC Class I and Class II.
CD69 induction was used to further quantitate and dissect MHC responses by preselection thymocytes. To estimate how many thymocytes have receptors capable of interacting with either MHC class I or II, MHC° thymocytes were reaggregated with stroma selectively expressing either class I or II molecules. As shown in Fig. 3 a (and summarized in Table 1), 4.7, 7.0, and 14.9% of thymocytes were responsive to stroma bearing class I or class II (Ab) alone or the entire H-2b haplotype, as judged by CD69 expression. Upregulation of the thymocyte activation/maturation marker CD5 (4, 31, 45) was seen with similar frequencies (Fig. 3 a, Table 1). As shown in Fig. 3 a and summarized in Table 1, the frequency of thymocytes responsive to MHC class I and II individually or in combination was approximately additive, indicating that most MHC responses in this assay are selective for either class I or II molecules. In experiments summarized in Table 1, we asked whether the provision of stroma bearing two MHC haplotypes could elevate the proportion of activated thymocytes still further. As shown, reaggregation with stroma expressing both H-2b and H-2k induced upregulation of CD69 and CD5 in 17.7 and 22.9% of MHC° thymocytes, as compared to 10.1 and 18.5% in response to H-2b alone (Table 1). Therefore, at least one in five MHC-naive thymocytes appear capable of MHC recognition.
Figure 3.


Preselection thymocyte responses to MHC class I and II are largely additive and, in addition to CD69, involve CD5 upregulation, downregulation of RAG-1 and -2 mRNA, and perturbed CD4 and CD8 expression. (a) Elevated CD5 and CD69 expression by MHC-naive thymocytes 24 h after contact with MHC. Note that responses to class I and II (Ab) are approximately additive. These responses are independent of superantigens, as verified using MHC-expressing cells from mice without mouse mammary tumor virus integrants (provided by Dr. E. Simpson, data not shown). (b) Downregulation of RAG-1 and -2 mRNA 24 h after MHC contact. RT-PCR analysis of thymocytes isolated by fluorescence activated cell sorting from reaggregates with MHC-deficient (lane 1) or MHC+ stroma (lanes 2 and 3). The latter were separated into CD69-negative (lane 2) and -positive cells (lane 3). RAG-1 and -2 signals are barely detectable in CD69-positive thymocytes; CD4 and, to a lesser extent, CD8a mRNA appear downregulated. RT-PCR for HPRT serves as a control; lane 4 contains no cDNA.
Table 1.
Incremental Recruitment of MHC-naive Thymocytes by MHC Class I, Class II, One Haplotype (H-2b), and Two Haplotypes (Hbxk)
| Stroma | % CD69 induction | % CD5 induction | ||||||
|---|---|---|---|---|---|---|---|---|
| mean ± 1 SD | n | mean ± 1 SD | n | |||||
| MHC° | 1.1 ± 0.6 | (23) | 1.8 ± 0.7 | (13) | ||||
| Class I+ (H-2b) | 5.7 ± 2.0 | (22) | 10.3 ± 6.2 | (12) | ||||
| Class II+ (Ab) | 8.2 ± 3.3 | (19) | 11.2 ± 3.7 | (11) | ||||
| MHC+ (H-2b) | 10.1 ± 2.5 | (15) | 18.5 ± 2.9 | (6) | ||||
| MHC+ (H-2bxk) | 17.7 ± 7.0 | (18) | 22.9 ± 7.9 | (13) | ||||
One of the least predictable aspects of the TCR rearrangement process is template-independent variability due to the activity of TdT (see review in reference 5). Since TdT activity is developmentally regulated with a postnatal onset, we compared MHC responses by MHC-naive thymocytes isolated from newborn mice and from 2-wk-old mice in parallel. As summarized in Table 2, the frequency of MHC responses was not substantially affected by TdT activity.
Table 2.
MHC-naive Thymocytes Isolated from Mice Aged <3 d or >2 wk (Before and After the Onset of TdT Expression) Respond to MHC with Similar Frequencies
| % CD69 induction | ||||||||
|---|---|---|---|---|---|---|---|---|
| Pre-TdT (<3 d) | Post-TdT (>2 wk) | |||||||
| mean ± 1 SD | n | mean ± 1 SD | n | |||||
| MHC° | 1.4 ± 0.5 | (6) | 1.0 ± 0.7 | (7) | ||||
| Class I+ (H-2b) | 6.6 ± 1.3 | (9) | 6.1 ± 2.7 | (5) | ||||
| Class II+ (Ab) | 10.5 ± 2.6 | (5) | 8.3 ± 3.4 | (7) | ||||
| MHC+ (H-2b) | 11.1 ± 1.3 | (5) | 10.4 ± 3.5 | (4) | ||||
| MHC+ (H-2bxk) | 15.8 ± 2.7 | (5) | 13.0, 21.2 | (2) | ||||
MHC-responsive Thymocytes Have Features Consistent with Entry into the Selection Process.
The early stages of thymocyte selection are associated with characteristic changes in gene expression (4). In particular, the described loss of Rag-1 and -2, and the transient downregulation of CD4 and CD8 messenger RNA (mRNA; 38, 46, 47) provided an opportunity to assess at the population level the consequences of MHC recognition in our assay (with the rationale that loss of expression would be visible only if it affected a majority of the population analyzed). Thymocytes recovered from short-term reaggregate cultures were separated into CD69-positive and -negative populations and analyzed by reverse transcriptase (RT)-PCR. The CD69-positive population had strongly reduced levels of mRNA for the recombination-activating genes RAG-1 and -2 (Fig. 3 b, lane 3), whereas RAG transcripts remained abundant in activation marker–negative cells from the same cultures (Fig. 3 b, lane 2) and in MHC° thymocytes exposed to MHC-deficient stroma (Fig. 3 b, lane 1). Similarly, expression of CD4 and CD8 was perturbed in activation marker–positive thymocytes (Fig. 3 b, lane 3). HPRT served as a control and remained unaffected throughout. It appears that the 15– 18% of thymocytes that respond to MHC encounter in reaggregate culture do so in a manner indicative of TCR engagement and entry into the early stages of thymic selection in vivo (4, 31, 37a, 38, 45, 46).
Recruitment of Preselection Thymocytes by MHC Is Cell Autonomous and Reflects Cognate TCR Interactions.
To clarify whether the activation marker upregulation analyzed in previous experiments requires (and reflects) cognate TCR– MHC interactions by individual thymocytes, we exploited thymocytes manipulated to express a single, HY-specific TCR (4, 39, 40). Here, as shown in Fig. 4 a, CD5 was substantially upregulated by contact with stroma bearing selecting (H-2b) MHC molecules (71.6%), but not by exposure to nonselecting (H-2d) stroma (3.0%), indicating a requirement for cognate TCR–MHC interactions. To address the possibility that the high frequency of MHC-naive thymocytes activated on exposure to MHC-expressing stroma (Figs. 1 and 3, Table 1) might result from indirect “bystander” effects (where a cell activates its neighbors and thereby amplifies the response), we compared MHC responses in reaggregate culture by HY TCR–transgenic thymocytes and non-TCR–transgenic, MHC-naive thymocytes cultured separately (Fig. 4 a, top and bottom, respectively), or mixed together (Fig. 4 a, middle). The presence of HY TCR–transgenic thymocytes did not appreciably alter the frequency of MHC responses among non-TCR– transgenic, MHC-naive cells (HY TCR−), or vice versa.
Figure 4.
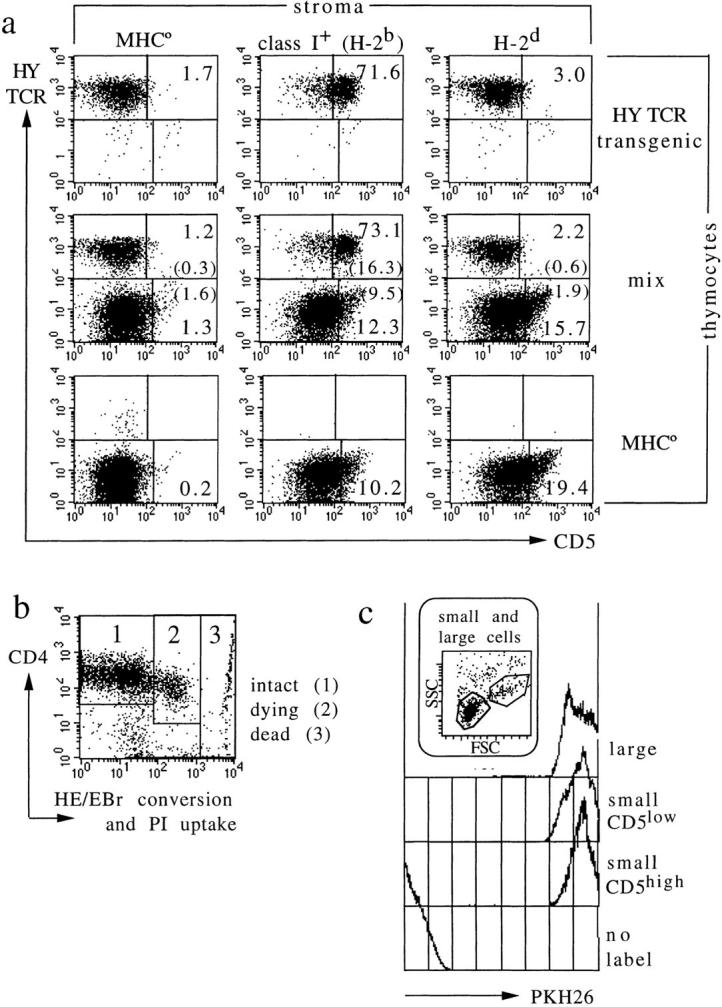
Bystander activation, unscheduled display of activation markers during apoptosis, or preferential expansion do not account for the frequency of MHC responsive thymocytes. (a) CD5 induction by cognate TCR–MHC interactions. HY TCR transgenic thymocytes from RAG-1–deficient mice lacking MHC class I expression respond strongly to selecting (H-2b), but not to nonselecting (H-2d) MHC molecules (top; Note that in contrast to an earlier study [45], our experiment excludes endogenous TCR rearrangements by using RAG-1–deficient thymocytes). Reaggregates containing mixtures of HY TCR transgenic and MHC-naive, TCR nontransgenic thymocytes (middle) show that both populations respond independently, and without significant bystander activation (compare top and bottom to middle). The percentage of cells in each region is indicated. In the middle panel, the percentage of activated cells among TCR transgenic and nontransgenic thymocytes is calculated separately (percentage of total cells is given in brackets). (b) Most cells recovered from short-term aggregate cultures are viable (i.e., exclude propidium iodide; PI) and metabolically intact, as indicated by low oxidative conversion of hydroethidine (HE) to ethidium bromide (EBr ; reference 33). Displays in Fig. 3 a and data in Table 1 are gated on the region marked 1. (c) MHC-responsive thymocytes do not selectively expand in short-term culture. Thymocytes were labeled with the fluorescent membrane dye PKH26 (Sigma Chemical Co.) before coculture with stroma for 24 h. Control traces are of unlabeled and large (cycling) thymocytes; the latter illustrating dye dilution on cell division. PKH26 fluorescence is the same for MHC-responsive and -nonresponsive cells (defined by CD5 expression level).
To exclude that our estimates are prejudiced by the preferential survival or expansion of MHC-responsive cells in reaggregate cultures, a detailed analysis of cell recovery, viability, and turnover was performed. After 24 h, the recovery of viable cells from reaggregates was close to input, and independent of whether the stroma expressed relevant MHC molecules (recovery was 86% of MHC-naive thymocytes cultured with MHC° stroma, 81% of MHC-naive thymocytes cultured with MHC+ stroma, 78% of TCR transgenic thymocytes cultured with nonselecting stroma, and 75% of TCR transgenic thymocytes cultured with selecting stroma). Between 77 and 83% of the cells recovered from cultures with MHC° or MHC+ stroma were lymphoid, and <90% of these were metabolically intact as indicated by normal mitochondrial membrane potential (not shown) and low hydroethidine/ethidium bromide conversion (Fig. 4 b, region 1; reduced mitochondrial membrane potential and increased hydroethidine/ethidiumand conversion are features of early apoptotic cells before DNA fragmentation; reference 33). Furthermore, the MHC-dependent upregulation of CD5 and CD69 reported here was blocked by cycloheximide (not shown), and is therefore distinct from the aberrant expression of activation markers reported to occur in the absence of de novo protein synthesis during thymocyte apoptosis (48). Finally, when thymocytes were labeled before culture with the membrane dye PKH26 (which is distributed between daughter cells on cell division), MHC-responsive and -unresponsive cells retained equal amounts of dye, excluding the possibility that MHC-responsive cells are preferentially expanded in reaggregate cultures (Fig. 4 c).
Similar Frequencies of MHC-responsive Thymocytes In Vitro and In Vivo.
The data presented above suggest that a large fraction of thymocytes can respond to MHC under in vitro conditions that support thymocyte differentiation. To determine if this potential is actually realized in vivo, we analyzed the expression of activation markers on CD4+CD8+ DP thymocytes freshly isolated ex vivo. Compared to MHC° thymocytes, 27.6 ± 3.5% (n = 9) of C57Bl/10 (H-2b) DP thymocytes expressed elevated CD5 levels (not shown), and elevated CD5 and/or CD69 expression was detectable in >20% of DP thymocytes from mice expressing MHC class I or II (Ab) selectively (Fig. 5). These frequencies are slightly higher than those determined using MHC° thymocytes in vitro, possibly because successful TCR engagement can halt further TCR α chain rearrangements (see review in reference 4), and thereby permit the accumulation of MHC-responsive receptors in vivo.
Figure 5.

In vivo expression of selection markers by CD4 CD8 DP thymocytes. Thymocytes taken from young adult mice lacking MHC (MHC°, left), or expressing MHC class I (middle) or class II (Ab; right), respectively, were stained for CD5 and CD69. Displays are gated on CD4+CD8+ double-positive cells as defined by the expression of CD4 (for thymocytes from MHC° and class I+ mice, left and middle) or CD8 (for thymocytes from class II+ mice, right). The percentages of cells expressing elevated levels of CD5, CD69, or both are indicated in the respective quadrants. Although this analysis is representative of H-2b mice, up to 50% of DP thymocytes are apparently engaged in the selection process in MHC haplotypes expressing class II E products (not shown).
Analysis of the Preselection Repertoire Using Mature T Cells.
Previous estimates suggest that as many as 1–10% of mature, postselection T cells can respond to allogeneic MHC products, and it has been argued that this observation is not readily accounted for by models for thymic selection (17). To explore the nature of this responsiveness to MHC, we attempted a direct comparison of MHC reactivity among MHC-naive and MHC class II–selected CD4 T cells (Fig. 6). Preculture with bispecific (CD3/CD4) antibodies (34) was used to drive thymocyte maturation in the absence of MHC selection (24, 35, 49, 50). In contrast to untreated MHC° thymi, MHC° thymi exposed to CD3/ CD4 bispecific antibodies generated percentages of CD4 SP thymocytes similar to MHC class II+ thymi (Fig. 6 a). Comparing MHC-naive and MHC class II-selected CD4 T cells by limiting dilution analysis, we found similar frequencies of alloreactive cells (1/757 and 1/691 thymocytes, respectively, or 1/621 and 1/587 CD4 cells, Fig. 6, b and c).
Figure 6.
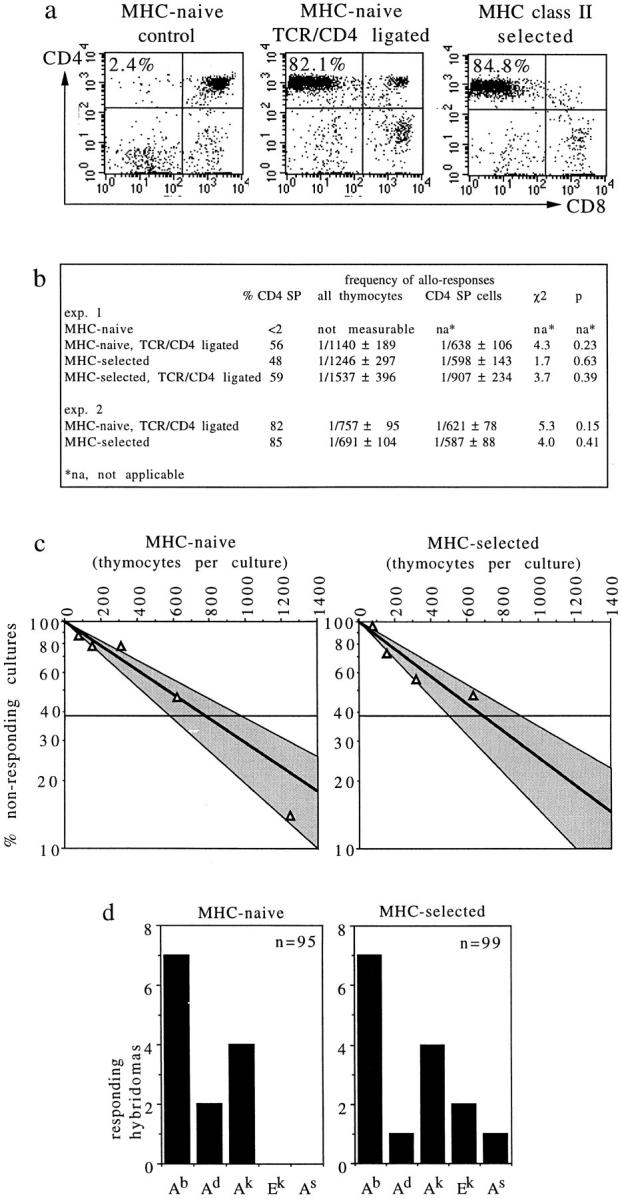
MHC-naive CD4 T cells respond to MHC. (a) Treatment of MHC° thymi with CD3/CD4 bispecific antibodies generates CD4 SP cells phenotypically and functionally similar to CD4 SP cells selected by MHC class II. (b) Limiting dilution analysis indicates that MHC-naive and class II–selected CD4 cells mount proliferative responses to allogeneic CBA/Ca splenocytes with similar frequencies, whereas MHC-naive thymocytes not treated with bispecific antibodies mount no detectable responses. (c) Graphic representation of experiment 2. Shaded areas indicate 95% confidence limits. (d) T cell hybridomas were generated from MHC-naive and class II–selected CD4 cells, and tested for reactivity to a panel of MHC class II transfected L cells. 8 of 95 (8.4%) and 13 of 99 (13.1%), respectively, responded to at least one class II transfectant, five hybridomas generated from MHC-naive cells and two hybridomas generated from class II–selected cells responded to two transfectants.
T cell hybridomas were generated from MHC-naive (CD3/4 hybrid antibody treated) and MHC class II–selected CD4 cells and tested for MHC reactivity. To exclude responses to class II–presented superantigens, a panel of MHC class II transfected L cells was used as stimulator cells (Fig. 6 d). 8 of 95 (8.4%) and 13 of 99 (13.1%) T cell hybridomas generated from MHC-naive and class II–selected CD4 cells, respectively, responded to at least one of five class II transfectants. Several Ab-selected thymocyte hybridomas responded to Ab-transfected L cells, probably reflecting an altered spectrum of MHC-associated peptides displayed by L cells in the absence of H-2M and invariant chain (51), since none of twenty Ab-selected hybridomas, but 2 of 20 antibody-generated thymocyte hybridomas tested responded to H-2b spleen cells (data not shown). Five hybridomas generated from MHC-naive cells and two hybridomas generated from class II–selected cells responded to more than one transfectant, perhaps indicating a trend towards greater cross-reactivity in the preselection as compared to the postselection repertoire. In conclusion, the preselection repertoire clearly contains TCRs able to confer MHC reactivity to mature T cells, and at frequencies similar to the repertoire that results from selection by MHC.
DISCUSSION
The extent to which the preselection TCR repertoire is MHC responsive had previously been addressed by extrapolation from mature T cell reactivity (14, 22, 24; see introduction). This approach is ultimately limited because of potential and reported differences in the responsiveness of preselection thymocytes versus mature T cells; MHC–peptide ligands may delete DP cells without activating mature T cells (25) and vice versa (26), and TCR-MHC-peptide interactions that drive positive selection do not generally meet the activation requirements of mature T cells (11, 26– 28, 41, 43).
We have taken a direct approach to evaluate the MHC responsiveness of preselection thymocytes. MHC-naive thymocytes were exposed to MHC-expressing thymic stroma in short-term reaggregate cultures, and the induction of the activation/selection markers CD69 and CD5 was measured by flow cytometry under conditions where most thymocytes seeded into the cultures could be recovered for quantitative analysis. We show that the frequency with which preselection thymocytes respond to MHC is high, exceeding or at least matching the highest estimates made by extrapolations from responses of normal postselection T cells to allogeneic MHC (17), MHC responses by CD4 T cells selected on restricted MHC–peptide complexes (22), or CD4 T cells generated independently of MHC (24). In our experiments, on average one in five preselection thymocytes responded to stroma expressing two parental MHC haplotypes. Control experiments ruled out several trivial explanations for this high frequency, including noncognate (e.g., coreceptor–MHC) interactions, the selective expansion or survival of MHC-responsive cells, the activation of bystanders that do not themselves see MHC, and the aberrant expression of activation markers by thymocytes in the early stages of apoptosis. Consistent with our observations in vitro, DP thymocytes analyzed ex vivo showed similar frequencies of activation marker–positive cells.
It has been estimated that four of nine thymocytes lack a functional TCR because they fail to produce in-frame TCR α chain rearrangements (52). In this scenario, as many as one in three successfully rearranged TCR would have to confer MHC reactivity to preselection thymocytes to account for the frequencies of MHC-responsive thymocytes reported here (if rearrangement is attempted for both alleles, and the net effect of sequential rearrangements is nil in the absence of MHC molecules to provide feedback on the rearrangement process; reference 4). Since the MHC responses observed were largely additive (Fig. 2 a, Table 1), additional MHC products would be expected to reveal an even greater proportion of responsive TCRs.
It appears that the evolutionary partnership between TCR and MHC has streamlined TCR rearrangement towards maximal variability in domains interacting predominantly with MHC-bound peptides, while minimally interfering with MHC interactions. These conditions would be met by the preferential use of germline-encoded TCR segments for MHC contacts and TCR–MHC orientation, as suggested by theoretical considerations (16, 53) and functional (3, 54) as well as structural data (55, 56). This point is illustrated by our observation that the frequency of MHC responses was not substantially affected by TdT activity, even though template-independent variability due to TdT activity represents one of the least predictable aspects of the TCR rearrangement process (see review in reference 5).
To ask to what extent preselection TCRs would confer MHC reactivity to mature T cells, we generated CD4 SP thymocytes from preselection thymocytes by limited CD3/ CD4 cross-linking (35) and compared their MHC reactivity to CD4 SP thymocytes that had been selected on MHC class II. The frequencies of MHC-responsive cells estimated by limiting dilution analysis and T cell hybridoma assays were similar for both populations, confirming that MHC selection is not required for MHC reactivity (24). Collectively, these data suggest a minimalist interpretation of allo-reactivity, in which mature T cells respond to allogeneic MHC products not because allo-reactive TCR are somehow selected for, but because allo-reactivity is not selected against.
CD4 SP thymocytes expressing pre- or postselection TCRs were equally MHC reactive, but at frequencies markedly lower than pre-selection thymocytes. Only one in several hundred CD4 SP thymocytes responded to allogeneic stimulator cells in proliferation assays under limiting dilution conditions, and individual class II transfectants were recognized on average by 2.6 of 95 (2.7%) and 3 of 99 (3.0%) pre- and postselection hybridomas, respectively. 9.7 ± 1.8% MHC-naive thymocytes responded to each L cell transfectant by expressing CD69 (compared to 2.1% CD69 induction by control L cells expressing endogenous H-2k MHC class I; data not shown). Perhaps predictably (if mature and immature T cells have different activation requirements), it appears that while the preselection repertoire confers moderate MHC reactivity to mature T cells, it is used to greater effect by DP thymocytes to attain MHC responsiveness, and a chance to enter the selection process.
Several lines of evidence link the observed MHC responsiveness of preselection thymocytes to thymocyte selection. First, when the requirements for CD69 induction were checked against a set of standards in the form of MHC–peptide complexes with well-characterized effects on thymocyte maturation, we found concordance between interactions that trigger CD69 expression and thymocyte selection, both positive and negative. In contrast, peptides without detectable effects on thymocyte selection failed to induce CD69, including one for which weak, but measurable TCR/MHC-peptide interactions can be demonstrated (44). Second, the majority of activation marker–positive thymocytes undergo physiological changes indicative of thymocyte selection in vivo, including downregulation of RAG-1 and -2 (both involved in TCR rearrangement and selection), as well as CD4 and CD8 mRNA (a feature of thymocytes in the DP to SP transition; references 46, 47). Third, the expression of activation markers in reaggregate culture was correlated with successful T cell maturation (as demonstrated in reaggregate cultures maintained for 5 d), and both were MHC dependent. CD69 and CD5 are expressed by selection intermediates in vivo (4, 37a, 38, 46) as well as in reaggregate cultures (30).
If more thymocytes enter the selection process than successfully complete it (7, 8), it is pertinent to consider evidence for mechanisms that may operate to limit mature T cell production. Even in TCR transgenic mice where all thymocytes are potentially selectable, only 20% of DP cells enter the SP pool (8), and the absolute number of mature T cells produced is similar to that in normal mice (9). It appears that access to relevant MHC–peptide complexes is not a limiting factor, because RAG expression is strongly downregulated in the majority of cortical thymocytes (38). Perhaps stromal cell microenvironments that can sustain the DP to SP transition are limiting, at least in a TCR transgenic scenario (8, 10). Since thymic selection is aborted when thymocytes are removed from their selecting environment, or thymocyte access to selecting ligands is blocked experimentally (38), it is possible that thymocytes are vulnerable because of a requirement for prolonged signaling, which in turn may facilitate the matching between TCR specificity and coreceptor expression (4, 13) as well as the fine tuning of T cell responsiveness during the DP to SP transition (11, 12). One can speculate that a particular TCR may allow entry into the selection process, but fail to sustain maturation at a later stage. Finally, negative selection must be an important candidate for limiting mature T cell production, but, as already discussed, estimates of its impact vary from negligible (6) to moderate (14) to overwhelming (15, 22).
The thymus reaggregate system used in this study does not distinguish between these possibilities because the method to prepare thymic stroma (preculture in deoxyguanosine) depletes dendritic cells, important mediators of negative selection. Nevertheless, it is provocative that the frequency of thymocytes recruited into the thymic selection process (this study) could theoretically accommodate even the highest estimates (15, 22). We are tempted to speculate that the true impact of negative selection may be closer to the estimates of Ignatowicz et al. (22) and van Meerwijk et al. (15) than to those of Surh and Sprent (6) or Laufer et al. (14). Surh and Sprent's conclusion is based on the observation that apoptotic thymocytes occur with similar frequencies in normal mice and in mice that can not negatively select due to lack of MHC expression. Another view of this result would be that, regardless of how many thymocytes normally die by negative selection, thymocyte death in MHC-deficient mice must be at least as common as in normal mice because DP thymocytes can not mature in the absence of MHC expression. The estimate of Laufer et al. (14) is based on limiting dilution analysis, an approach likely to yield lower figures than assays on preactivated T cell clones or hybridomas (e.g., reference 24 and this study), which according to our data, more accurately reflect thymocyte response to MHC.
Acknowledgments
This work was supported by the Medical Research Council (UK).
Footnotes
Drs. D. Mathis, C. Benoist, B. Koller, H. von Boehmer, D. Kioussis, E. Spanopoulou, M. Owen, E. Simpson, W. Heath, and F. Carbone are thanked for mice; Drs. R. Hilton, R. Lechler, and D. Gray for cells; Dr. R. Edwards for help with statistics; and Drs. P. Travers and S. Jameson for communicating results before publication.
Abbreviations used in this paper: β2m, β2 microglobulin; °, deficient; DP, double positive; HPRT, hypoxanthine guanine phosphoribosyl transferase; mRNA, messenger RNA; rag, recombination-activating gene; RT, reverse transcriptase; SP, single positive; TdT, terminal deoxynucleotidyl transferase.
REFERENCES
- 1.Zinkernagel RM, Doherty PC. MHC-restricted cytotoxic T cells: studies on the biological role of polymorphic major transplantation antigens determining T-cell restriction-specificity, function, and responsiveness. Adv Immunol. 1979;27:51–177. doi: 10.1016/s0065-2776(08)60262-x. [DOI] [PubMed] [Google Scholar]
- 2.Bevan MJ. In a radiation chimaera, host H-2 antigens determine immune responsiveness of donor cytotoxic cells. Nature (Lond) 1977;269:417–418. doi: 10.1038/269417a0. [DOI] [PubMed] [Google Scholar]
- 3.Jorgensen JL, Reay PA, Ehrich EW, Davis MM. Molecular components of T-cell recognition. Annu Rev Immunol. 1992;10:835–873. doi: 10.1146/annurev.iy.10.040192.004155. [DOI] [PubMed] [Google Scholar]
- 4.Kisielow P, von Boehmer H. Development and selection of T cells: facts and puzzles. Adv Immunol. 1995;58:87–209. doi: 10.1016/s0065-2776(08)60620-3. [DOI] [PubMed] [Google Scholar]
- 5.Gilfillan S, Benoist C, Mathis D. Mice lacking terminal deoxynucleotidyl transferase: adult mice with a fetal antigen receptor repertoire. Immunol Rev. 1995;148:201–219. doi: 10.1111/j.1600-065x.1995.tb00099.x. [DOI] [PubMed] [Google Scholar]
- 6.Surh CD, Sprent J. T-cell apoptosis detected in situ during positive and negative selection in the thymus. Nature (Lond) 1994;372:100–103. doi: 10.1038/372100a0. [DOI] [PubMed] [Google Scholar]
- 7.Shortman K, Vremec D, Egerton M. The kinetics of T cell antigen receptor expression by subgroups of CD4+8+thymocytes: delineation of CD4+8+3(2+) thymocytes as post-selection intermediates leading to mature T cells. J Exp Med. 1991;173:323–332. doi: 10.1084/jem.173.2.323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Huesmann M, Scott B, Kisielow P, von Boehmer H. Kinetics and efficacy of positive selection in the thymus of normal and T cell receptor transgenic mice. Cell. 1991;66:533–540. doi: 10.1016/0092-8674(81)90016-7. [DOI] [PubMed] [Google Scholar]
- 9.Kelly KA, Pircher H, von Boehmer H, Davis MM, Scollay R. Regulation of T cell production in T cell receptor transgenic mice. Eur J Immunol. 1993;23:1922–1928. doi: 10.1002/eji.1830230829. [DOI] [PubMed] [Google Scholar]
- 10.Merkenschlager M, Benoist C, Mathis D. Evidence for a single niche model of positive selection. Proc Natl Acad Sci USA. 1994;91:11694–11698. doi: 10.1073/pnas.91.24.11694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jameson SC, Hogquist KA, Bevan MJ. Positive selection of thymocytes. Annu Rev Immunol. 1995;13:93–126. doi: 10.1146/annurev.iy.13.040195.000521. [DOI] [PubMed] [Google Scholar]
- 12.Grossman Z, Singer A. Tuning of activation thresholds explains flexibility in the selection and development of T cells in the thymus. Proc Natl Acad Sci USA. 1996;93:14747–1452. doi: 10.1073/pnas.93.25.14747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chan S, Cosgrove D, Waltzinger C, Benoist C, Mathis D. Another view of the selective model of thymocyte selection. Cell. 1993;73:225–236. doi: 10.1016/0092-8674(93)90225-f. [DOI] [PubMed] [Google Scholar]
- 14.Laufer TM, DeKoning J, Markowitz JS, Lo D, Glimcher LH. Unopposed positive selection and autoreactivity in mice expressing class II MHC only on thymic cortex. Nature (Lond) 1996;383:81–85. doi: 10.1038/383081a0. [DOI] [PubMed] [Google Scholar]
- 15.van Meerwijk JPM, Marguerat S, Lees RK, Germain RN, Fowlkes BJ, MacDonald HR. Quantitative impact of thymic clonal deletion on the T cell repertoire. J Exp Med. 1997;185:377–383. doi: 10.1084/jem.185.3.377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Jerne NK. The somatic generation of immune regulation. Eur J Immunol. 1971;1:1–9. doi: 10.1002/eji.1830010102. [DOI] [PubMed] [Google Scholar]
- 17.Matzinger P. Why positive selection? . Immunol Rev. 1993;135:81–117. doi: 10.1111/j.1600-065x.1993.tb00645.x. [DOI] [PubMed] [Google Scholar]
- 18.Blackman M, Yague J, Kubo R, Gay D, Coleclough C, Palmer E, Kappler J, Marrack P. The T cell repertoire may be biased in favor of MHC recognition. Cell. 1986;47:349–357. doi: 10.1016/0092-8674(86)90591-x. [DOI] [PubMed] [Google Scholar]
- 19.Miyazaki T, Wolf P, Tourne S, Waltzinger C, Dierich A, Barois N, Ploegh H, Benoist C, Mathis D. Mice lacking H2-M complexes, enigmatic elements of the MHC class II peptide-loading pathway. Cell. 1996;84:531–541. doi: 10.1016/s0092-8674(00)81029-6. [DOI] [PubMed] [Google Scholar]
- 20.Martin WD, Hicks GG, Mendiratta SK, Leva HI, Ruley HE, van Kaer L. H2-M mutant mice are defective in the peptide loading of class II molecules, antigen presentation, and T cell repertoire selection. Cell. 1996;84:543–550. doi: 10.1016/s0092-8674(00)81030-2. [DOI] [PubMed] [Google Scholar]
- 21.Fung-Leung WP, Surh CD, Liljedahl M, Pang J, Leturcq D, Peterson PA, Webb SR, Karlsson L. Antigen presentation and T cell development in H2-M–deficient mice. Science (Wash DC) 1996;271:1278–1281. doi: 10.1126/science.271.5253.1278. [DOI] [PubMed] [Google Scholar]
- 22.Ignatowicz L, Kappler J, Marrack P. The repertoire of T cells shaped by a single MHC/peptide ligand. Cell. 1996;84:521–529. doi: 10.1016/s0092-8674(00)81028-4. [DOI] [PubMed] [Google Scholar]
- 23.Vidovic D, Boulanger N, Kuye O, Toral J, Ito K, Guenot J, Bluethmann H, Nagy ZA. The helper T cell repertoire of mice expressing class II major histocompatibility complex beta chains in the absence of alpha chains. Immunogenetics. 1997;45:325–335. doi: 10.1007/s002510050212. [DOI] [PubMed] [Google Scholar]
- 24.Zerrahn J, Held W, Raulet DH. The MHC reactivity of the T cell receptor repertoire prior to positive and negative selection. Cell. 1997;88:627–636. doi: 10.1016/s0092-8674(00)81905-4. [DOI] [PubMed] [Google Scholar]
- 25.Pircher H, Rohrer UH, Moskophidis D, Zinkernagel RM, Hengartner H. Lower receptor avidity required for thymic clonal deletion than for effector T-cell function. Nature (Lond) 1991;351:482–485. doi: 10.1038/351482a0. [DOI] [PubMed] [Google Scholar]
- 26.Sebzda E, Kündig TM, Thomson CT, Aoki K, Mak SY, Mayer JP, Zamborelli T, Nathenson SG, Ohashi PS. Mature T cell reactivity altered by peptide agonist that induces positive selection. J Exp Med. 1996;183:1093–1104. doi: 10.1084/jem.183.3.1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ashton-Rickardt P, Bandeira A, Delaney JR, van Kaer L, Pircher H-P, Zinkernagel RM, Tonegawa S. Evidence for a differential avidity model of T cell selection in the thymus. Cell. 1994;76:651–663. doi: 10.1016/0092-8674(94)90505-3. [DOI] [PubMed] [Google Scholar]
- 28.Hogquist KA, Jameson SC, Heath WR, Howard JL, Bevan MJ, Carbone FR. T cell antagonist peptides induce positive selection. Cell. 1994;76:17–27. doi: 10.1016/0092-8674(94)90169-4. [DOI] [PubMed] [Google Scholar]
- 29.Jenkinson E, Anderson G, Owen JJT. Studies on T cell maturation on defined thymic stromal cell populations in vitro. J Exp Med. 1992;176:845–853. doi: 10.1084/jem.176.3.845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Anderson G, Owen JJ, Moore NC, Jenkinson EJ. Characteristics of an in vitro system of thymocyte positive selection. J Immunol. 1994;153:1915–1920. [PubMed] [Google Scholar]
- 31.Punt JA, Osborne BA, Takahama Y, Sharrow SO, Singer A. Negative selection of CD4+CD8+thymocytes by T cell receptor–induced apoptosis requires a costimulatory signal that can be provided by CD28. J Exp Med. 1994;179:709–713. doi: 10.1084/jem.179.2.709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Levelt CN, Eichmann K. Receptors and signals in early thymic selection. Immunity. 1995;3:667–672. doi: 10.1016/1074-7613(95)90056-x. [DOI] [PubMed] [Google Scholar]
- 33.Zamzami N, Marchetti P, Castedo M, Decaudin D, Macho A, Hirsch T, Susin SA, Petit PX, Mignotte B, Kroemer G. Sequential reduction of mitochondrial transmembrane potential and generation of reactive oxygen species in early programmed cell death. J Exp Med. 1995;182:367–377. doi: 10.1084/jem.182.2.367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kostelny SA, Cole MS, Tso JY. Formation of a bispecific antibody by the use of leucine zippers. J Immunol. 1992;148:1547–1553. [PubMed] [Google Scholar]
- 35.Bommhardt U, Cole MS, Tso JY, Zamoyska R. Signals through CD4 or CD8 can induce commitment to the CD4 lineage in the thymus. Eur J Immunol. 1997;27:1152–1163. doi: 10.1002/eji.1830270516. [DOI] [PubMed] [Google Scholar]
- 36.Merkenschlager M, Terry L, Edwards R, Beverley PCL. Limiting dilution analysis of proliferative responses in human lymphocyte populations defined by the monoclonal antibody UCHL1: implications for differential CD45 expression in T cell memory formation. Eur J Immunol. 1988;18:1653–1661. doi: 10.1002/eji.1830181102. [DOI] [PubMed] [Google Scholar]
- 37.White J, Blackman M, Bill J, Kappler J, Marrack P, Gold DP, Born W. Two better cell lines for making hybridomas expressing specific T cell receptors. J Immunol. 1989;143:1822–1825. [PubMed] [Google Scholar]
- 37a.Bendelac A, Matzinger P, Seder RA, Paul WE, Schwartz RH. Activation events during thymic selection. J Exp Med. 1992;175:731–742. doi: 10.1084/jem.175.3.731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Brändle D, Müller S, Müller C, Hengartner H, Pircher H. Regulation of RAG-1 and CD69 expression in the thymus during positive and negative selection. Eur J Immunol. 1994;24:145–151. doi: 10.1002/eji.1830240122. [DOI] [PubMed] [Google Scholar]
- 39.Spanopoulou E, Cortes P, Shih C, Huang CM, Silver DP, Svec P, Baltimore D. Localization, interaction, and RNA binding properties of the V(D)J recombination-activating proteins RAG1 and RAG2. Immunity. 1995;3:715–726. doi: 10.1016/1074-7613(95)90061-6. [DOI] [PubMed] [Google Scholar]
- 40.Koller BH, Marrack P, Kappler JW, Smithies O. Normal development of mice deficient in beta 2m, MHC class I proteins, and CD8+T cells. Science (Wash DC) 1990;248:1227–1230. doi: 10.1126/science.2112266. [DOI] [PubMed] [Google Scholar]
- 41.Hogquist KA, Tomlinson AJ, Kieper WC, McGargill MA, Naylor S, Jameson SC. Identification of a naturally occurring ligand for positive selection. Immunity. 1997;6:389–399. doi: 10.1016/s1074-7613(00)80282-4. [DOI] [PubMed] [Google Scholar]
- 42.Jameson SC, Carbone FR, Bevan MJ. Clone-specific T cell receptor antagonists of major histocompatibility complex class I–restricted cytotoxic T cells. J Exp Med. 1993;1993:1541–1550. doi: 10.1084/jem.177.6.1541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Jameson SC, Hogquist KA, Bevan MJ. Specificity and flexibility in thymic selection. Nature (Lond) 1994;369:750–752. doi: 10.1038/369750a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Alam SM, Travers PJ, Wung JL, Nasholds W, Redpath S, Jameson SC, Gascoigne NR. T-cell–receptor affinity and thymocyte positive selection. Nature (Lond) 1996;381:616–620. doi: 10.1038/381616a0. [DOI] [PubMed] [Google Scholar]
- 45.Dutz JP, Ong CJ, Marth J, Teh H-S. Distinct differentiative stages of CD4+CD8+thymocyte development defined by the lack of coreceptor binding in positive selection. J Immunol. 1995;154:2588–2599. [PubMed] [Google Scholar]
- 46.Lucas B, Germain RN. Unexpectedly complex regulation of CD4/CD8 coreceptor expression supports a revised model for CD4+CD8+thymocyte differentiation. Immunity. 1996;5:461–477. doi: 10.1016/s1074-7613(00)80502-6. [DOI] [PubMed] [Google Scholar]
- 47.Cibotti R, Punt JA, Dash KS, Sharrow SO, Singer A. Surface molecules that drive T cell development in vitro in the absence of thymic epithelium and in the absence of lineage specific signals. Immunity. 1997;6:245–255. doi: 10.1016/s1074-7613(00)80327-1. [DOI] [PubMed] [Google Scholar]
- 48.Kishimoto H, Surh CD, Sprent J. Upregulation of surface markers on dying thymocytes. J Exp Med. 1995;181:649–655. doi: 10.1084/jem.181.2.649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Müller KP, Kyewski BA. T cell receptor targeting to thymic cortical epithelial cells in vivo induces survival, activation and differentiation of immature thymocytes. Eur J Immunol. 1993;23:1661–1670. doi: 10.1002/eji.1830230740. [DOI] [PubMed] [Google Scholar]
- 50.Takahama Y, Suzuki H, Katz KS, Grusby MJ, Singer A. Positive selection of CD4+ T cells by TCR ligation without aggregation even in the absence of MHC. Nature (Lond) 1994;371:67–70. doi: 10.1038/371067a0. [DOI] [PubMed] [Google Scholar]
- 51.Katz JF, Stebbins C, Appella E, Sant AJ. Invariant chain and DM edit self-peptide presentation by major histocompatibility complex (MHC) class II molecules. J Exp Med. 1996;184:1747–1753. doi: 10.1084/jem.184.5.1747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Malissen, B., and M. Malissen. 1995. Allelic exclusion of T cell receptor genes. In T Cell Receptors. J.I. Bell, M.J. Owen, and E. Simpson, editors. Oxford University Press, Oxford. 352–365.
- 53.Davis MM, Bjorkman P. T-cell antigen receptor genes and T-cell recognition. Nature (Lond) 1988;334:395–402. doi: 10.1038/334395a0. [DOI] [PubMed] [Google Scholar]
- 54.Sant'Angelo DB, Waterbury G, Preston-Hurlburt P, Yoon ST, Medzhitov R, Hong SC, Janeway CA., Jr The specificity and orientation of a TCR to its peptide–MHC class II ligands. Immunity. 1996;4:367–376. doi: 10.1016/s1074-7613(00)80250-2. [DOI] [PubMed] [Google Scholar]
- 55.Garboczi DN, Ghosh P, Utz U, Fan QR, Biddison WE, Wiley DC. Structure of the complex between human T cell receptor, viral peptide and HLA A2. Nature (Lond) 1996;384:134–141. doi: 10.1038/384134a0. [DOI] [PubMed] [Google Scholar]
- 56.Garcia KC, Degano M, Stanfield RL, Brunmark A, Jackson MR, Peterson PA, Teyton L, Wilson IA. An alpha beta T cell receptor structure at 2.5 Å and its orientation in the TCR–MHC complex. Science (Wash DC) 1996;274:209–219. doi: 10.1126/science.274.5285.209. [DOI] [PubMed] [Google Scholar]