

Abstract
CCR5 is a chemokine receptor expressed by T cells and macrophages, which also functions as the principal coreceptor for macrophage (M)-tropic strains of HIV-1. To understand the molecular basis of the binding of chemokines and HIV-1 to CCR5, we developed a number of mAbs that inhibit the various interactions of CCR5, and mapped the binding sites of these mAbs using a panel of CCR5/CCR2b chimeras. One mAb termed 2D7 completely blocked the binding and chemotaxis of the three natural chemokine ligands of CCR5, RANTES (regulated on activation normal T cell expressed and secreted), macrophage inflammatory protein (MIP)-1α, and MIP-1β, to CCR5 transfectants. This mAb was a genuine antagonist of CCR5, since it failed to stimulate an increase in intracellular calcium concentration in the CCR5 transfectants, but blocked calcium responses elicited by RANTES, MIP-1α, or MIP-1β. This mAb inhibited most of the RANTES and MIP-1α chemotactic responses of activated T cells, but not of monocytes, suggesting differential usage of chemokine receptors by these two cell types. The 2D7 binding site mapped to the second extracellular loop of CCR5, whereas a group of mAbs that failed to block chemokine binding all mapped to the NH2-terminal region of CCR5. Efficient inhibition of an M-tropic HIV-1–derived envelope glycoprotein gp120 binding to CCR5 could be achieved with mAbs recognizing either the second extracellular loop or the NH2-terminal region, although the former showed superior inhibition. Additionally, 2D7 efficiently blocked the infectivity of several M-tropic and dual-tropic HIV-1 strains in vitro. These results suggest a complicated pattern of HIV-1 gp120 binding to different regions of CCR5, but a relatively simple pattern for chemokine binding. We conclude that the second extracellular loop of CCR5 is an ideal target site for the development of inhibitors of either chemokine or HIV-1 binding to CCR5.
Chemokines mediate a range of proinflammatory effects on leukocytes, such as chemotaxis, degranulation, and integrin activation (1–3). The chemokines have been divided into four families, based on the configuration of cysteine residues near the NH2 terminus. The CC family, which includes macrophage inflammatory protein (MIP)- 1α,1 MIP-1β, RANTES (regulated on activation normal T cell expressed and activated), monocyte chemotactic protein (MCP)-1, -2, -3, and -4, are generally chemotactic for T cells, monocytes, basophils, and eosinophils (1–5) but not neutrophils. These chemokines attract leukocytes by binding to the seven transmembrane–spanning G-protein coupled receptors CCR1 through CCR8 (1, 6–9). The expression of chemokine receptors on leukocytes directs leukocyte chemotactic responses to particular sets of chemokines, both in vitro and in vivo (5, 10–14). The chemokine receptor CCR5 appears to be one of the important receptors for directing the migration of activated and effector T cells, since these T cells respond robustly to the CCR5 ligands RANTES, MIP-1α, and MIP-1β in chemotaxis assays (15–18), and CCR5 is expressed at high levels on these cells (19). The precise role of other chemokine receptors on T cells has been difficult to assess, since specific reagents or receptor antagonists have not been available.
Chemokine receptors also serve as coreceptors for HIV-1 entry into cells. CCR5 is the principal coreceptor for primary macrophage (M)-tropic HIV-1 strains (20–24) , while CXCR4 supports infection of CD4+ cells by T-tropic HIV-1 strains (25). The envelope glycoprotein gp120 of HIV-1, upon binding to CD4, interacts specifically with the coreceptors (26–28). The importance of CCR5 for HIV-1 transmission is underscored by the findings that individuals who have a defect in CCR5 expression are generally resistant to infection with HIV-1 (29–32). In addition, CD4+ T cells from these individuals are also highly resistant in vitro to the entry of primary M-tropic HIV-1 (29, 33). This resistance results from a defective CCR5 allele that contains an internal 32-bp deletion (CCR5 Δ32). To date, no immunological defects have been noted in either CCR5 Δ32 homozygous or heterozygous individuals. The resistance of CCR5 Δ32 homozygous individuals to infection with HIV-1 has prompted a widespread effort to develop antagonists of CCR5 that may be used therapeutically to inhibit HIV-1 transmission or to delay progression to AIDS (34).
Recently, much attention has been focused on the molecular interactions of CCR5 with HIV-1, as well as the interactions of CCR5 with its natural CC chemokine ligands (35–40). Understanding the nature of these interactions should help in the development of antagonists of CCR5, to inhibit either HIV-1 or chemokine binding. One approach to probe the interactions of CCR5, and to block these interactions, is to use mAbs. A panel of mAbs to CCR5 has recently been produced (19), and these mAbs inhibit M-tropic HIV-1 infection of T cells. Here we used a panel of anti-CCR5 mAbs to inhibit either chemokine binding and function or HIV-1 gp120 binding and HIV-1 infection. We used CCR5/CCR2b chimeras to map the domains on CCR5 recognized by these mAbs, and correlated inhibitory activity with the domain specificity of the mAbs.
Materials and Methods
CCR5/CCR2 Chimeras.
Several CCR5/CCR2 chimeras (C25-01 to C25-14) were constructed by transferring restriction fragments flanked by the common BamHI, AfIII, ClaI, EcoRI, and XbaI sites between CCR5 and CCR2b. The construction and characterization of these chimeras have been previously described (35). The constructs were transferred into a bicistronic vector (41), under dependence of the elongation factor 1a promoter, and transfected in Chinese hamster ovary-K1 cells as previously described (42). G418-resistant cell populations were used in FACS® analyses.
Cells and Cell Lines.
PBMCs were isolated as described (5). The CD3+ blasts were generated using anti-CD3 antibody TR77 and maintained in medium supplemented with recombinant human IL-2 as previously described (19). Other cell lines used included THP-1 and transfectants of the L1.2 murine pre–B cell lymphoma, expressing high levels of CCR5 (19, 26) or CXCR4. The different transfectants were monitored for expression of the relevant receptors, using specific mAbs (15, 19).
Chemokine and HIV-1 gp120 Binding.
125I-labeled human RANTES, MIP-1α, and MIP-1β were purchased from DuPont-NEN (Boston, MA), and unlabeled chemokines were from PeproTech, Inc. (Rocky Hill, NJ). Chemokine binding to target cells was carried out using a modification of a method previously reported (26, 43). CCR5 L1.2 cells or CD3+ blasts were washed and resuspended in binding buffer (50 mM Hepes, pH 7.5, 1 mM CaCl2, 5 mM MgCl2, and 0.5% BSA) at 5 × 106/ml. For each binding reaction (in a final volume of 100 μl), 25 μl of cell suspension (1.25 × 105 cells) was mixed with 0.1 nM radio-labeled chemokine with or without an appropriate amount of anti-CCR5 mAb, or an isotype-matched control mAb. Total binding was in the presence of radiolabeled chemokines only, and nonspecific binding (background) was determined in the presence of 100 nM unlabeled chemokines. The reactions were incubated at room temperature for 45–60 min, and stopped by transferring the mixture to GFB filter plates which were then washed 2–3 times with binding buffer containing 0.5 M NaCl. The plates were dried and MicroScint scintillation fluid (Packard Instrument Company Inc., Meriden, CT) was added before counting. Each sample was assayed in duplicate. The envelope gp120 protein derived from HIV-1 JR-FL (27; gift from Paul Maddon, Progenics Pharmaceuticals, Inc. Tarrytown, NY) was iodinated using solid phase lactoperoxidase to a specific activity of 20 μCi/μg. Binding to target cells was performed similarly as for radiolabeled chemokine binding, except that soluble CD4 was included in the assays, as previously reported (26).
mAbs, Immunofluorescent Staining, and FACS® Analysis.
mAb 2D7 reactive with CCR5 was generated by immunizing mice with L1.2 cells expressing high levels of transfected CCR5, as previously described (19). C57BL6 mice were immunized with 107 cells, intraperitoneally, six times at 2-wk intervals, and 4 d after an intravenous injection, the spleen was removed and cells were fused with the SP2/0 cell line. The mAb generated, 2D7, was determined to be IgG1. Other mAbs used in this study included 5A11, an anti-CCR2b mAb (15), PE-conjugated 12G5, an anti-CXCR4 mAb (44; PharMingen, San Diego, CA), and 3A9, an anti-CCR5 mAb (19). The reactivity of mAbs against transfected cells or leukocytes was assessed using indirect immunofluorescence and flow cytometry techniques as previously described (19).
Chemotaxis Assays.
Chemotaxis experiments with human PBMC, or CD3-activated, IL-2–stimulated T cells, employed the cell line ECV304 to coat Biocoat® Transwell tissue culture inserts (Collaborative Biomedical Products, Bedford, MA), exactly as previously described (5). Chemotaxis with L1.2 receptor transfectant cell lines was carried out similarly except that endothelial cells were not used to coat the Biocoat® Transwell tissue culture inserts and the incubation was for 4–6 h (45). Cells migrating to the bottom chamber of the Transwell were enumerated using the FACScan®, by counting cells for 30 s. Tight forward angle and side scatter gates were set to exclude debris or irrelevant cells.
Measurement of [Ca2+]i.
Cells were labeled with the fluorochrome Fura-2 AM (Molecular Probes, Eugene, OR), as previously described (14). In brief, Fura-2 AM was added to the cell suspension to produce a final concentration of 0.2 mol/106 cells. After incubation at 37°C for 30 min, excess dye was removed by centrifugation and cells were resuspended at a concentration of 106/ml in 125 mM NaCl, 5 mM KCl, 1 mM MgCl2, 1 mM CaCl2, 0.5 mM glucose, 0.025% BSA, and 20 mM Hepes, pH 7.4. [Ca2+]i was measured using excitation at 340 and 380 nm on a Hitachi F-2000 fluorescence spectrometer. Calibration was performed using 1% NP-40 for total fluorophore release and 25 μM EGTA to chelate free Ca2+.
Inhibition of HIV-1 Infection by Anti-CCR5 mAb 2D7.
Inhibition of HIV-1 infection in U87-CD4-CCR5 cells was determined using a virus entry assay based on single-cycle infection. Cells were infected with the env-deficient virus NL4/3 luc (46) complemented in trans with envelope glycoproteins from several clones. Infection of the cells was measured by quantification of luciferase activity. In brief, U87-CD4-CCR5 cells (gift from D. Littman, New York University Medical Center, New York; see reference 47) were split to a concentration of 5 × 104 cells/ml, and 100 μl was added to each well of a 96-well tissue culture plate. The following day, the cells were washed with PBS and preincubated with dilutions of mAb 2D7, an isotype-control (IgG1) mAb, or medium only in a total volume of 40 μl for 1 h at 4°C. 50 μl of HIV-1 (env genes of ADA, JR-FL, DH123, or HxB2, stocks of 100 ng/ml, as measured by p24) was added, and the cells were incubated with the mAb and the virus for 2 h at 37°C. The cells were then washed, fresh medium was added immediately, and added again after 48 h. 72 h after infection, the cells were washed with PBS and lysed in 50 μl of 1× reporter lysis buffer (Promega Corp., Madison, WI). To measure luciferase activity, 100 μl of luciferase substrate (Promega Corp.) was added to 30 μl of the cell lysate.
Results
Generation of Anti-CCR5 mAbs that Recognize Different Domains of CCR5.
We sought to generate mAbs to CCR5 to inhibit the various functions of this molecule, and to understand how different CCR5 domains bind chemokines and HIV-1. We have previously described anti-CCR5 mAbs that inhibit HIV-1 binding, but not chemokine binding (19). New mAbs to CCR5 were generated by immunizing C57BL6 mice with the murine pre–B cell lymphoma line, L1.2, which expresses high levels of transfected human CCR5. An mAb, termed 2D7, reacted with CCR5-transfected L1.2 cells, as well as Chinese hamster ovary cells expressing certain portions of CCR5 (see Fig. 2), but not with L1.2 cells expressing CXCR4 (Fig. 1) or various other receptors, including CCR2b (not shown). Moreover, 2D7 showed an identical pattern of reactivity against human leukocytes, as previously noted for our other anti-CCR5 mAbs (19, 48). In particular, it stained mostly the CXCR4− subset of human PBL (Fig. 1 B), as well as a subset of tissue macrophages (not shown).
Figure 2.

Reactivity of CCR5-specific mAbs with CCR5/CCR2b receptor chimeras. The structures of CCR5/CCR2b chimeras used in this study are shown schematically on the left-hand side. Regions derived from CCR5 are shown in light gray, and regions derived from CCR2b are shown in black. Stable Chinese hamster ovary cell transfectants expressing various CCR5/CCR2b receptor chimeras were stained with anti-CCR5 mAb 3A9, 5C7, 2D7, anti-CCR2b mAb 5A11, or anti-CXCR1 mAb 7D9. The level of staining of the transfectants by the various mAbs was graded + to +++, or negative (neg).
Figure 1.

mAb 2D7 recognizes CCR5 specifically. (A) Reactivity of mAb 2D7 with CCR5 L1.2 cells, but not with CXCR4 L1.2 cells. (B) Two color staining of human PBL with 2D7 (green fluorescence, x-axis) and 12G5 (anti-CXCR4, red fluorescence, y-axis). Quadrants were set on the basis of control and single color stainings.
To determine how chemokines or HIV-1 interacts with CCR5, a series of chimeric receptors was generated by replacing extracellular domains of CCR5 with the corresponding domain of CCR2b, or vice versa, using common restriction sites in regions conserved between the two molecules (35). The chimeras of CCR5 and CCR2b were ideal for this purpose, since these two receptors are closely related but have completely different ligand binding properties. The interaction of these chimeras with different strains of HIV-1 has already been reported (35). Fig. 2 shows the panel of chimeras that was used in the present experiments, and the reactivity of these chimeras to several mAbs. The 2D7 mAb reacted with all chimeras that contained the second extracellular loop of CCR5. In particular, C25-14, a chimera comprising CCR2b with the second extracellular loop of CCR5, was stained intensely by mAb 2D7. In contrast, the anti-CCR5 mAb 3A9 (as well as seven other previously reported anti-CCR5 mAbs) reacted only with chimeras that contained the NH2-terminal region of CCR5 (Fig. 2). In addition, mutants of CCR5 lacking the NH2-terminal eight amino acids were not reactive with mAb 3A9, suggesting that the epitope for this mAb was dependent on the very NH2 terminus of the molecule. An mAb to CCR2b, 5A11, stained all chimeras containing the NH2 terminus of CCR2b, consistent with the fact that this mAb was raised against a synthetic peptide comprising the 32 NH2-terminal amino acids of CCR2b (15).
The Second Extracellular Loop mAb, but Not the NH2 Terminus–specific mAbs, Blocks MIP-1α, MIP-1β, and RANTES Binding to CCR5 Transfectants, as well as to Activated T Cells.
A preliminary analysis of a panel of anti-CCR5 mAbs revealed that none of the eight previously identified anti-CCR5 mAbs was able to block the binding of the CCR5 ligands RANTES, MIP-1α, or MIP-1β to CCR5 L1.2 transfectants. The ability of mAb 2D7 to inhibit the binding of these chemokines was assessed. Fig. 3 A shows that 10 μg/ml of mAb 2D7 was able to inhibit completely the binding of 125I-labeled human RANTES, MIP-1α, and MIP-1β to CCR5 L1.2 transfectants. An analysis with decreasing amounts of mAb 2D7 established an inhibitory constant (IC)50 of 23 ng/ml for MIP-1α binding, 41 ng/ml for MIP-1β binding, and 58 ng/ml for RANTES binding (not shown). mAb 3A9, directed to the NH2 terminus of CCR5, caused minimal inhibition of binding of the three chemokines at 10 μg/ml (Fig. 3 A), and only slight inhibition at concentrations up to 100 μg/ml (not shown). THP-1 cells, which do not express CCR5 (19), were also examined as a specific control. These cells bound MIP-1α (Fig. 3 B) and RANTES (not shown), and mAb 2D7 had no effect on the level of binding.
Figure 3.

mAb 2D7 inhibits the binding of 125I–MIP-1α, –MIP-1β, and -RANTES to CCR5. CCR5 L1.2 cells (A), THP1 cells (B), or day 15–activated IL-2–stimulated CD3 blast T cells (C) were incubated with 0.1 nM 125I-labeled–MIP-1α, –MIP-1β, or -RANTES, in the absence (total binding) or presence of either 10 μg/ml of mAb 2D7 (an IgG1 isotype), mAb 3A9, control IgG1 mAb, or 100 nM unlabeled chemokine. After 45–60 min at room temperature, cells were washed and counted as described in Materials and Methods. Data are shown as the percentage of total binding, i.e., in the absence of mAb or unlabeled chemokines.
125I-labeled RANTES, –MIP-1α, and –MIP-1β binding to activated T cells is shown in Fig. 3 C. These three chemokines bound to IL-2–maintained T cells, and this binding could be competed with 100 nM unlabeled chemokine. T cells from day 21 after activation showed the highest level of binding and chemotactic responses to the three chemokines (see below). mAb 2D7 was assessed for its ability to compete for binding of these CC chemokines. At 10 μg/ml, 2D7 completely blocked the MIP-1β binding to these activated T cells. Under the same conditions, 125I-RANTES and 125I–MIP-1α binding were inhibited by 95 and 85%, respectively. This result indicated that CCR5 was responsible for most of the RANTES, MIP-1α, or MIP-1β binding to these T cells. Nevertheless, some variations were noted in the 2D7 inhibition level when using T cells from different time points (10–26 d). At earlier time points, fewer RANTES and MIP-1α binding sites were blocked by mAb 2D7.
mAb 2D7 Inhibits CCR5 Functional Responses to MIP-1α, MIP-1β, and RANTES.
The agonist/antagonist activity of mAb 2D7 was tested on CCR5 L1.2 transfectants, by measuring whether there was a change in intracellular calcium concentration [Ca2+]i of Fura-2–loaded cells in response to various concentrations of mAb 2D7. mAb 2D7 itself failed to stimulate a change in [Ca2+]i in CCR5 L1.2 cells, but was able to inhibit subsequent stimulation by MIP-1α (Fig. 4) as well as by RANTES and MIP-1β (not shown). mAb 2D7 did not inhibit a change in [Ca2+]i after stimulation with stromal cell–derived factor–1, which operates through an endogenous mouse CXCR4 receptor. Incubation of CCR5 L1.2 cells with a control mAb (MOPC-21) had no inhibitory effect. In addition, neither mAb 3A9 nor any of the other CCR5 NH2-terminal–specific mAbs had any inhibitory effect on the RANTES-, MIP-1α– and MIP-1β–mediated Ca2+ flux responses.
Figure 4.
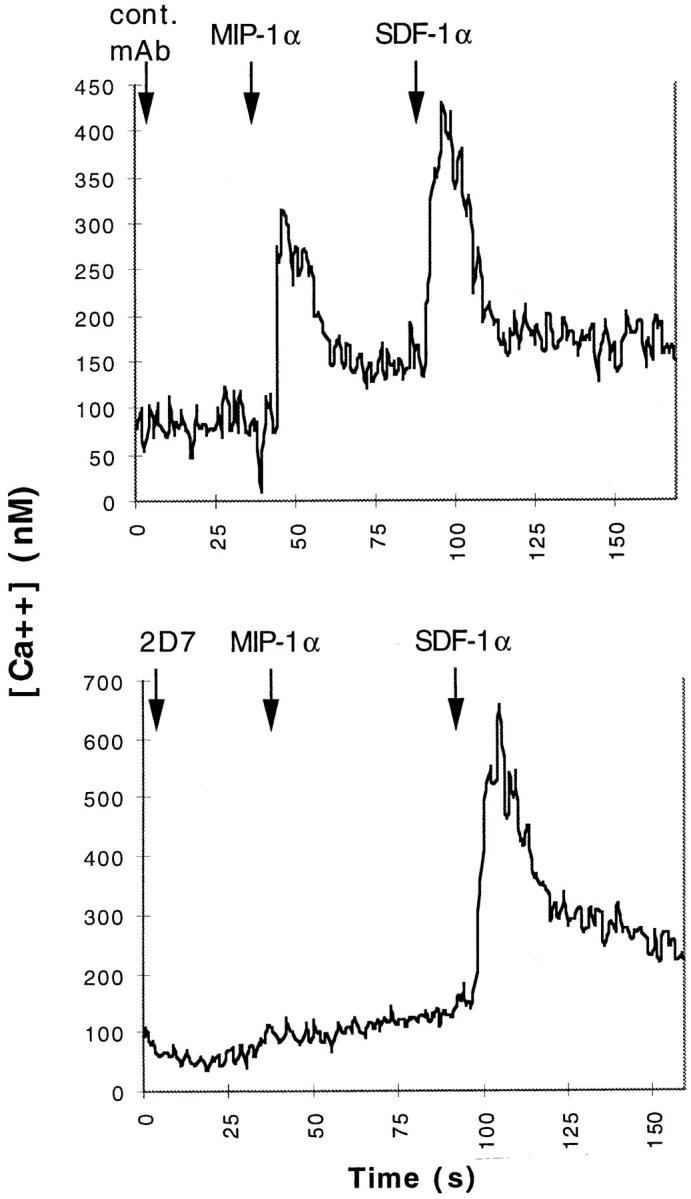
mAb 2D7 inhibits [Ca2+]i flux in CCR5 L1.2 cells in response to MIP-1α. CCR5 L1.2 cells were labeled with Fura-2 as described in Materials and Methods, and stimulated sequentially with mAb, followed 40 s later with MIP-1α, and 100 s with SDF-1. [Ca2+]i fluorescence changes were recorded using a spectrofluorometer. The tracings were representative of three separate experiments. In the top panel, an irrelevant mAb (MOPC-21) was used, and in the bottom panel, mAb 2D7. Antibodies were used at a final concentration of 20 μg/ml. MIP-1α was used at 100 nM and SDF-1 was used at 200 nM.
mAb 2D7 inhibited the chemotaxis of CCR5 L1.2 cells to RANTES, MIP-1α, and MIP-1β, in a dose-dependent manner (Fig. 5 A). Incubation of cells with 20 μg/ml of mAb in the top chamber was sufficient to achieve complete inhibition of migration to all three chemokines. The ability to achieve complete inhibition with 2D7 allowed us to examine the significance of CCR5 for lymphocyte, monocyte, and activated T cell responses to RANTES, MIP-1α, and MIP-1β. As shown in Fig. 5 B, chemotactic responses by blood lymphocytes to MIP-1β were almost completely inhibited by 2D7, consistent with the notion that MIP-1β binds only CCR5 and not other receptors. RANTES responses were also inhibited in most individuals, but MIP-1α responses were not. mAb 2D7 showed no dose-dependent inhibition of monocytes to RANTES or MIP-1α (Fig. 5 C). This result is consistent with previous studies showing minimal expression of CCR5 on most monocytes (19). T cells stimulated in vitro with anti-CD3 and maintained with IL-2 for 3 wk showed a very robust chemotactic response to RANTES, MIP-1α, and MIP-1β (Fig. 5 D). T cells maintained in culture for shorter periods of time also responded, although not quite as robustly (not shown). Importantly, mAb 2D7 was able to inhibit most of the functional chemotactic responses of these T cells to RANTES and MIP-1β, and ∼60–80% of the chemotactic response to MIP-1α. However we did note individual-to-individual variation. These results were supported by studies with T cell lines from Δ32 homozygous individuals, in that the MIP-1α and RANTES chemotactic responses were markedly impaired in these cells (data not shown).
Figure 5.

Inhibition of chemotactic responses of various cell types to MIP-1α, MIP-1β, and RANTES, using mAb 2D7. A, CCR5 L1.2 cells; B, blood lymphocytes; C, blood monocytes; and D, day 21 activated, IL-2–stimulated T cells. For these experiments, 106 cells were placed in the top chamber of the Transwell and an optimal concentration of chemokine (usually 12.5 nM) was placed in the bottom chamber. Various concentrations of 2D7 mAb were placed in the top well. After various periods of time (1–4 h) the cells migrating to the bottom chamber were counted using flow cytometry. The results are representative of at least four separate experiments. Untransfected L1.2 cells showed no migration to MIP-1α, MIP-1β, or RANTES (data not shown). Chemotactic index was calculated by dividing the number of migrated cells in response to a specific chemokine by that in the absence of chemokine (background). The background values for these cells are: 28 (CCR5 L1.2), 136 (lymphocytes), 507 (monocytes), and 615 (CD3 blasts).
Inhibition of gp120 Binding to CCR5 and HIV-1 Infection Is Manifest by mAbs Recognizing Either the NH2 Terminus or the Second Extracellular Loop.
The exterior envelope glycoprotein gp120 of M-tropic primary HIV-1, upon binding to CD4, can interact with CCR5 specifically and with high affinity (26, 27). To assess the ability of the various anti-CCR5 mAbs to inhibit this interaction, we performed binding assays using 125I-labeled gp120 derived from HIV-1 JR-FL (an M-tropic strain; reference 27) in the presence or absence of 3A9 or 2D7 (Fig. 6 A). mAb 2D7 inhibited efficiently the binding of 125I-gp120 to CCR5 L1.2 cells, in the presence of soluble CD4, with an inhibitory constant (IC)50 of ∼20 ng/ml. At a concentration of 100 ng/ml, 2D7 inhibited the 125I-gp120 binding to the same level as that obtained with excess unlabeled gp120. In contrast, mAb 3A9, at the low concentration range, had a moderate inhibitory effect on the 125I-gp120 binding, but inhibited completely at a higher concentration (⩾100 μg/ml). An isotype-control mAb had no significant inhibitory effect at a concentration up to 100 μg/ml.
Figure 6.

Inhibition of 125I-gp120 binding and HIV-1 infection by anti-CCR5 mAbs. (A) Inhibition of 125I-labeled M-tropic HIV-1 JR-FL gp120 binding to CCR5 L1.2 transfectants by mAb 2D7 and 3A9. CCR5 L1.2 cells were incubated with 0.2 nM 125I-labeled gp120 and 20 nM sCD4 in the absence or presence of increasing concentrations of mAb 2D7 or 3A9. After 60 min at room temperature, cells were washed and counted as described in Materials and Methods. An IgG1 control mAb was used as a control. 100% of inhibition was defined as that caused by 100 nM of unlabeled gp120. (B) Inhibition of HIV-1 infection in U87-CD4-CCR5 cells by mAb 2D7. The infectability of U87-CD4-CCR5 cells by M-tropic (ADA and JR-FL), dual-tropic (DH123), and T-tropic (HxB2) HIV-1 strains, in the absence or presence of increasing concentrations of 2D7 or 50 μg/ml of an IgG1 control mAb, was determined using a virus entry assay based on single-cycle infection as described in Materials and Methods. Infection of the cells was measured by quantification of luciferase activity.
To study the inhibitory effect of 2D7 on HIV-1 entry, we employed a virus entry assay based on single-cycle infection using several viral strains. As shown in Fig. 6 B, the U87MG-CD4+ cells expressing transfected CCR5 (47) can be efficiently infected by M-tropic (ADA and JR-FL env) and dual-tropic (DH123 env) chimeric viruses, which can use CCR5 as coreceptor, but not by the T cell–tropic chimera (HxB2), which uses only CXCR4. mAb 2D7 efficiently inhibited the entry of the dual-tropic DH123 chimera (>90% inhibition at 1 μg/ml), whereas a higher concentration of 2D7 (∼10 μg/ml for ∼90% inhibition) was required for inhibiting the M-tropic (ADA and JR-FL) strains. Under the same conditions, the isotype-control mAb had no significant effect. mAb 3A9 showed little effect on the HIV-1 entry of U87-CD4-CCR5 cells, but it could effectively neutralize the infection of PBMC by M-tropic HIV-1 strains as previously shown (19).
Discussion
In this study, we sought to develop and use blocking anti-CCR5 mAbs to determine the important regions on CCR5 for chemokine binding and HIV-1 gp120 interactions. Additionally, we used the blocking mAb to determine the importance of CCR5 for T cell and monocyte responses to RANTES, MIP-1α, and MIP-1β.
Identifying regions of CCR5 that facilitate HIV-1 gp120 binding, or chemokine binding, is important for understanding the molecular basis of CCR5 interactions, and for developing inhibitors of these interactions. Results from several studies imply that gp120 binds to CCR5 via multiple domains (35, 36, 38, 40). However, different conclusions have been reached as to the importance of the various extracellular domains of CCR5 for HIV-1 interactions. Previous studies with the panel of CCR5/CCR2b receptor chimeras used here showed that the NH2 terminus of CCR5, as well as the first extracellular loop (but not the second extracellular loop), were important for M-tropic HIV-1 binding (35). In contrast, studies by Bieniasz et al. (38), using 89.6 and ADA, found that the second extracellular loop was important for binding. Using human/mouse CCR5 chimeras, they showed that ADA could utilize a chimeric receptor containing any single human CCR5 extracellular domain, while BaL required any two human extracellular domains, and 89.6 required three domains (38).
Regardless of the way different strains of HIV-1 bind to CCR5, another issue is how to develop agonists or antagonists that can inhibit HIV-1 binding to various regions of CCR5. To date, the most effective inhibitors of HIV-1 binding to CCR5 have been CC chemokines, or modified forms of them (21, 23, 24, 49–51). Here we show that an anti-CCR5 mAb 2D7, which recognizes the second extracellular loop, was able to completely inhibit the binding of JR-FL gp120 to CCR5, and to efficiently block the entry of M-tropic (ADA and JR-FL) and dual-tropic (DH123) HIV-1 strains. Anti-CCR5 mAbs that mapped to the NH2 terminus were also able to inhibit M-tropic HIV-1 gp120 binding or viral entry into T cells, albeit somewhat less potently. This is consistent with studies using CCR5/CCR2b chimeras which show that M-tropic HIV-1 binding relies on the NH2 terminus and the first extracellular loop (35).
It has been shown recently that the binding of HIV-1 and chemokines to the coreceptor are nonoverlapping functions of CCR5 (36, 37, 39). All evidence implicates the second extracellular loop of CCR5 as the principal binding element for RANTES, MIP-1α, or MIP-1β (51a; Charo, I., personal communication). Consistent with this, mAb 2D7 to the second extracellular loop was able to inhibit totally the binding of RANTES, MIP-1α, and MIP-1β, whereas mAbs recognizing the NH2 terminus of CCR5 were ineffective. The different effect between the two panels of mAbs is unlikely due to their different binding affinities to the receptor, since all eight mAbs directed to the NH2 terminus showed a similar lack of inhibitory activity. Furthermore, the inhibitory effect of 2D7 on chemokine and HIV-1 binding did not result from receptor downmodulation, since the same level of CCR5 cell surface staining was detected on CCR5 L1.2 cells that were preincubated with 2D7 for 1 h at room temperature and on those that were not preincubated (Kassam, N., and L. Wu, unpublished results). Therefore, whereas the specificity of CC chemokine binding to CCR5 is determined by a single domain, the gp120 binding site is more complex and probably involves at least two domains, although changes in the NH2 terminus can be sufficient and can significantly affect gp120 binding and HIV-1 entry. It is conceivable that the ability of an mAb to inhibit HIV-1 binding may have more to do with steric hindrance rather than with direct interruption of the binding sites.
We predict that mAb 2D7 will block the binding to CCR5 of most HIV-1 strains that can use this coreceptor, as exemplified here with ADA, JR-FL, and DH123, and by other studies which indicate that 2D7 is able to block the replication of a wide range of primary isolates in PBMC (Wilkinson, P., and R. Koup, personal communication). The potential to disrupt HIV-1 gp120 binding with agents that interfere with either the NH2 terminus or the second extracellular loop suggests that small molecule antagonists that can bind to different parts of CCR5 might also be effective at blocking CCR5-gp120 interactions. Since mAb 2D7 can efficiently block all known functions of CCR5, the second extracellular loop might be a particularly good target for the development of small molecule antagonists.
Much confusion has surrounded the chemotactic responses of T cells to chemokines (52), as well as the qualitative and quantitative aspects of chemokine receptor expression. mAbs have proven to be a powerful tool for dissecting chemokine receptor expression and function on leukocytes (14). Our results and those of others show that a subset of memory/effector T cells responds to RANTES, MIP-1α, and MIP-1β in chemotaxis assays (15, 53). Candidate receptors that could be mediating these responses include CCR1, CCR3, CCR4, and CCR5. The use of anti-CCR5 mAbs demonstrated that CCR5 is expressed intensely on a subset of memory T cells, as well as on activated, IL-2–stimulated T cell lines (19). In addition, CCR5 is the only receptor yet known to bind MIP-1β. We do not now have reagents to examine CCR1 or CCR4 expression on the cell surface, so our approach in this study was to inhibit CCR5 function and assess the remaining ligand binding and functional responses to RANTES, MIP-1α, and MIP-1β by various cell types. The mAb 2D7 inhibited most of the MIP-1β (>95%) and RANTES (>90%), and partially inhibited the MIP-1α (60–80%) chemotactic responses by activated T cells. These results suggest that although CCR5 is probably the only MIP-1β receptor and the predominant RANTES receptor on T cells, other receptors as well as CCR5 play a role for MIP-1α responses. CCR1 responds to MIP-1α and its message is clearly upregulated in IL-2–stimulated T cell lines, and is thus a likely candidate. However, the assessment of actual receptor expression will have to await the development of specific mAbs.
In contrast to T cells, monocytes responded to RANTES and MIP-1α through a receptor other than CCR5. This result is consistent with the low level of CCR5 expression that is found on most circulating monocytes (19). However, CCR5 is upregulated on monocytes after their activation in vitro (our unpublished results), and is also expressed by a population of tissue macrophages (19). The receptor on circulating monocytes which mediates the RANTES and MIP-1α function could be CCR1. Thus, there appears to be a relatively good dichotomy in receptor expression; CCR5 on T cells, and CCR1 on monocytes, although these receptors are not restricted to these two leukocyte lineages. The role of CCR1 on monocytes and T cells will be understood better after the development of a reliable antagonistic mAb.
In conclusion, we identified the second extracellular loop of CCR5 as an important target site for developing inhibitors of CC chemokine binding as well as HIV-1 gp120 binding to CCR5. Hopefully, future studies will determine the suitability of CCR5 antagonists as therapeutics for inflammation or HIV-1 infection.
Acknowledgments
We thank Dr. Dan Littman for providing U87MG-CCR5 transfectants, Dr. Paul Maddon for JR-FL gp120, Dr. Shixin Qin for CXCR4 L1.2 transfectants, and Dr. Antonio Lanzavecchia for recombinant human IL-2. We also wish to thank Dr. Walter Newman, Dr. Tim Springer, Dr. Tatjana Dragic, and Dr. Rick Koup for helpful discussions and critical review.
Footnotes
M. Samson and M. Parmentier at the I.R.I.B.H.N. were supported by the Belgian programme on Interdisciplinary Poles of Attraction and the French Agence Nationale de Recherche contre le SIDA. J.P. Moore was supported by National Institutes of Health grant AI-41420 and by the Pediatric AIDS foundation, of which he is an Elizabeth Glaser Scientist.
Abbreviations used in this paper: M, macrophage; MCP, monocyte chemotactic protein; MIP, macrophage inflammatory protein; RANTES, regulated on activation normal T cell expressed and secreted.
References
- 1.Baggiolini M, Dewald B, Moser B. IL-8 and related chemotactic cytokines-CXC and CC chemokines. Adv Immunol. 1994;55:97–179. [PubMed] [Google Scholar]
- 2.Oppenheim JJ, Zachariae CO, Mukaida N, Matsushima K. Properties of the novel proinflammatory supergene “intercrine” cytokine family. Annu Rev Immunol. 1991;9:617–648. doi: 10.1146/annurev.iy.09.040191.003153. [DOI] [PubMed] [Google Scholar]
- 3.Miller MD, Krangel MS. Biology and biochemistry of the chemokines: a family of chemotactic and inflammatory cytokines. Crit Rev Immunol. 1992;12:17–46. [PubMed] [Google Scholar]
- 4.Jose PJ, Griffiths-Johnson DA, Collins PD, Walsh DT, Moqbel R, Totty NF, Truong O, Hsuan JJ, Williams TJ. Eotaxin: a potent eosinophil chemoattractant cytokine detected in a guinea pig model of allergic airways inflammation. J Exp Med. 1994;179:881–887. doi: 10.1084/jem.179.3.881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ponath PD, Qin S, Ringler DJ, Clark-Lewis I, Wang J, Kassam N, Smith H, Shi X, Gonzalo J-A, Newman W, et al. Cloning of the human eosinophil chemoattractant, eotaxin. Expression, receptor binding, and functional properties provide a mechanism for the selective recruitment of eosinophils. J Clin Invest. 1996;97:604–612. doi: 10.1172/JCI118456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Murphy PM. The molecular biology of leukocyte chemoattractant receptors. Annu Rev Immunol. 1994;12:593–633. doi: 10.1146/annurev.iy.12.040194.003113. [DOI] [PubMed] [Google Scholar]
- 7.Schall TJ, Bacon KB. Chemokines, leukocyte trafficking, and inflammation. Curr Opin Immunol. 1994;6:865–873. doi: 10.1016/0952-7915(94)90006-x. [DOI] [PubMed] [Google Scholar]
- 8.Gerard C, Gerard NP. The pro-inflammatory seven-transmembrane segment receptors of the leukocyte. Curr Opin Immunol. 1994;6:140–145. doi: 10.1016/0952-7915(94)90045-0. [DOI] [PubMed] [Google Scholar]
- 9.Mackay, C.R. 1997. What chemokine is that? Curr. Biol. In press. [DOI] [PubMed]
- 10.Ponath PD, Qin S, Post TW, Wang J, Wu L, Gerard NP, Newman W, Gerard C, Mackay CR. Molecular cloning and characterization of a human eotaxin receptor expressed selectively on eosinophils. J Exp Med. 1996;183:2437–2448. doi: 10.1084/jem.183.6.2437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Broaddus VC, Boylan AM, Hoeffel JM, Kim KJ, Sadick M, Chuntharapai A, Hebert CA. Neutralization of IL-8 inhibits neutrophil influx in a rabbit model of endotoxin-induced pleurisy. J Immunol. 1994;152:2960–2967. [PubMed] [Google Scholar]
- 12.Mulligan MS, Jones ML, Bolanowski MA, Baganoff MP, Deppeler CL, Meyers DM, Ryan US, Ward PA. Inhibition of lung inflammatory reactions in rats by an anti-human IL-8 antibody. J Immunol. 1993;150:5585–5595. [PubMed] [Google Scholar]
- 13.Sekido N, Mukaida N, Harada A, Nakanishi I, Watanabe Y, Matsushima K. Prevention of lung reperfusion injury in rabbits by a monoclonal antibody against interleukin-8. Nature (Lond) 1993;365:654–657. doi: 10.1038/365654a0. [DOI] [PubMed] [Google Scholar]
- 14.Heath H, Qin S, Rao P, Wu L, LaRosa G, Kassam N, Ponath PD, Mackay CR. Chemokine receptor usage by human eosinophils. The importance of CCR3 demonstrated using an antagonistic monoclonal antibody. J Clin Invest. 1997;99:178–184. doi: 10.1172/JCI119145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Qin S, LaRosa G, Campbell JJ, Smith-Heath H, Kassam N, Shi X, Zeng L, Butcher EC, Mackay CR. Expression of monocyte chemoattractant protein–1 and interleukin-8 receptors on subsets of T cells: correlation with transendothelial chemotactic potential. Eur J Immunol. 1996;26:640–647. doi: 10.1002/eji.1830260320. [DOI] [PubMed] [Google Scholar]
- 16.Taub DD, Lloyd AR, Conlon K, Wang JM, Ortaldo JR, Harada A, Matsushima K, Kelvin DJ, Oppenheim JJ. Recombinant human interferon–inducible protein 10 is a chemoattractant for human monocytes and T lymphocytes and promotes T cell adhesion to endothelial cells. J Exp Med. 1993;177:1809–1814. doi: 10.1084/jem.177.6.1809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Taub DD, Proost P, Murphy WJ, Anver M, Longo DL, van Damme J, Oppenheim JJ. Monocyte chemotactic protein-1 (MCP-1), -2, and -3 are chemotactic for human T lymphocytes. J Clin Invest. 1995;95:1370–1376. doi: 10.1172/JCI117788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Schall TJ, Bacon K, Toy KJ, Goeddel DV. Selective attraction of monocytes and T lymphocytes of the memory phenotype by cytokine RANTES. Nature (Lond) 1990;347:669–671. doi: 10.1038/347669a0. [DOI] [PubMed] [Google Scholar]
- 19.Wu L, Paxton WA, Kassam N, Ruffing N, Rottman JB, Sullivan N, Choe H, Sodroski J, Newman W, Koup RA, Mackay CR. CCR5 levels and expression pattern correlate with infectability by macrophage-tropic HIV-1, in vitro. J Exp Med. 1997;185:1681–1691. doi: 10.1084/jem.185.9.1681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Choe H, Farzan M, Sun Y, Sullivan N, Rollins B, Ponath PD, Wu L, Mackay CR, LaRosa G, Newman W, et al. The β-chemokine receptors CCR3 and CCR5 facilitate infection by primary HIV-1 isolates. Cell. 1996;85:1135–1148. doi: 10.1016/s0092-8674(00)81313-6. [DOI] [PubMed] [Google Scholar]
- 21.Alkhatib G, Combadière C, Broder CC, Feng Y, Kennedy PE, Murphy PM, Berger EA. CC CKR5: a RANTES, MIP-1α, MIP-1β receptor as a fusion cofactor for macrophage-tropic HIV-1. Science (Wash DC) 1996;272:1955–1958. doi: 10.1126/science.272.5270.1955. [DOI] [PubMed] [Google Scholar]
- 22.Doranz BJ, Rucker J, Yi Y, Smyth RJ, Samson M, Peiper SC, Parmentier M, Collman RG, Doms RW. A dual-tropic primary HIV-1 isolate that uses fusin and the β-chemokine receptors CKR-5, CKR-3, and CKR-2b as fusion cofactors. Cell. 1996;85:1149–1158. doi: 10.1016/s0092-8674(00)81314-8. [DOI] [PubMed] [Google Scholar]
- 23.Deng H, Liu R, Ellmeier W, Choe S, Unutmaz D, Burkhart M, Di Marzio P, Marmon S, Sutton RE, Hill CM, et al. Identification of a major co-receptor for primary isolates of HIV-1. Nature (Lond) 1996;381:661–666. doi: 10.1038/381661a0. [DOI] [PubMed] [Google Scholar]
- 24.Dragic T, Litwin V, Allaway GP, Martin SR, Huang Y, Nagashima KA, Cayanan C, Maddon PJ, Koup RA, Moore JP, Paxton WA. HIV-1 entry into CD4+cells is mediated by the chemokine receptor CC-CKR-5. Nature (Lond) 1996;381:667–673. doi: 10.1038/381667a0. [DOI] [PubMed] [Google Scholar]
- 25.Feng Y, Broder CC, Kennedy PE, Berger EA. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein–coupled receptor. Science (Wash DC) 1996;272:872–877. doi: 10.1126/science.272.5263.872. [DOI] [PubMed] [Google Scholar]
- 26.Wu L, Gerard N, Wyatt R, Choe H, Parolin C, Ruffing N, Borsetti A, Cardoso AA, Desjardin E, Newman W, et al. CD4-induced interaction of primary HIV-1 gp120 glycoproteins with the chemokine receptor CCR-5. Nature (Lond) 1996;384:179–183. doi: 10.1038/384179a0. [DOI] [PubMed] [Google Scholar]
- 27.Trkola A, Dragic T, Arthos J, Binley JM, Olson WC, Allaway GP, Cheng-Mayer C, Robinson J, Maddon PJ, Moore JP. CD4-dependent, antibody-sensitive interactions between HIV-1 and its co-receptor CCR5. Nature (Lond) 1996;384:184–186. doi: 10.1038/384184a0. [DOI] [PubMed] [Google Scholar]
- 28.Lapham CK, Ouyang J, Chandrasekhar B, Nguyen NY, Dimitrov DS, Golding H. Evidence for cell-surface association between fusin and the CD4-gp120 complex in human cell lines. Science (Lond) 1996;274:602–605. doi: 10.1126/science.274.5287.602. [DOI] [PubMed] [Google Scholar]
- 29.Liu R, Paxton WA, Choe S, Ceradini D, Martin SR, Horuk R, MacDonald ME, Stuhlmann H, Koup RA, Landau NR. Homozygous defect in HIV-1 coreceptor accounts for resistance of some multiply-exposed individuals to HIV-1 infection. Cell. 1996;86:367–377. doi: 10.1016/s0092-8674(00)80110-5. [DOI] [PubMed] [Google Scholar]
- 30.Samson M, Libert F, Doranz BJ, Rucker J, Liesnard C, Farber CM, Saragosti S, Lapoumeroulie C, Cognaux J, Forceille C, et al. Resistance to HIV-1 infection in caucasian individuals bearing mutant alleles of the CCR-5 chemokine receptor gene. Nature (Lond) 1996;382:722–725. doi: 10.1038/382722a0. [DOI] [PubMed] [Google Scholar]
- 31.Dean M, Carrington M, Winkler C, Huttley GA, Smith MW, Allikmets R, Goedert JJ, Buchbinder SP, Vittinghoff E, Gomperts E, et al. Genetic restriction of HIV-1 infection and progression to AIDS by a deletion allele of the CKR5 structural gene. Science (Wash DC) 1996;273:1856–1862. doi: 10.1126/science.273.5283.1856. [DOI] [PubMed] [Google Scholar]
- 32.Huang Y, Paxton WA, Wolinsky SM, Neumann AU, Zhang L, He T, Kang S, Ceradini D, Jin Z, Yazdanbakhsh K, et al. The role of a mutant CCR5 allele in HIV-1 transmission and disease progression. Nat Med. 1996;2:1240–1243. doi: 10.1038/nm1196-1240. [DOI] [PubMed] [Google Scholar]
- 33.Paxton WA, Martin SR, Tse D, O'Brien TR, Skurnick J, VanDevanter NL, Padian N, Braun JF, Kotler DP, Wolinsky SM, Koup RA. Relative resistance to HIV-1 infection of CD4 lymphocytes from persons who remain uninfected despite multiple high-risk sexual exposure. Nat Med. 1996;2:412–417. doi: 10.1038/nm0496-412. [DOI] [PubMed] [Google Scholar]
- 34.Cohen J. Exploiting the HIV-chemokine nexus. Science (Wash DC) 1997;275:1261–1264. doi: 10.1126/science.275.5304.1261. [DOI] [PubMed] [Google Scholar]
- 35.Rucker J, Samson M, Doranz BJ, Libert F, Berson JF, Yi Y, Smyth RJ, Collman RG, Broder CC, Vassart G, et al. Regions in β-chemokine receptors CCR5 and CCR2b that determine HIV-1 cofactor specificity. Cell. 1996;87:437–446. doi: 10.1016/s0092-8674(00)81364-1. [DOI] [PubMed] [Google Scholar]
- 36.Atchison RE, Gosling J, Monteclaro FS, Franci C, Digilio L, Charo IF, Goldsmith MA. Multiple extracellular elements of CCR5 and HIV-1 entry: dissociation from response to chemokines. Science (Wash DC) 1996;274:1924–1926. doi: 10.1126/science.274.5294.1924. [DOI] [PubMed] [Google Scholar]
- 37.Farzan M, Choe H, Martin KA, Sun Y, Sidelko M, Mackay CR, Newman W, Gerard NP, Sodroski J, Gerard C. HIV-1 entry and macrophage inflammatory protein-1β–mediated signaling are independent functions of the chemokine receptor CCR5. J Biol Chem. 1997;272:6854–6857. doi: 10.1074/jbc.272.11.6854. [DOI] [PubMed] [Google Scholar]
- 38.Bieniasz PD, Fridell RA, Aramori I, Ferguson SSG, Caron MG, Cullen BR. HIV-1–induced cell fusion is mediated by multiple regions within both the viral envelope and the CCR-5 coreceptor. EMBO (Eur Mol Biol Organ) J. 1997;16:2599–2609. doi: 10.1093/emboj/16.10.2599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gosling J, Monteclaro FS, Atchison RE, Arai H, Tsou C, Goldsmith MA, Charo IF. Molecular uncoupling of C-C chemokine receptor 5–induced chemotaxis and signal transduction from HIV-1 coreceptor activity. Proc Natl Acad Sci USA. 1997;94:5061–5066. doi: 10.1073/pnas.94.10.5061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lu ZH, Berson JF, Chen YH, Turner JD, Zhang TY, Sharron M, Jenks MH, Wang ZX, Kim J, Rucker J, et al. Evolution of HIV-1 coreceptor usage through interactions with distinct CCR5 and CXCR4 domains. Proc Natl Acad Sci USA. 1997;94:6426–6431. doi: 10.1073/pnas.94.12.6426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ghattas IR, Sanes JR, Majors JE. The encephalomyocarditis virus internal ribosome entry site allows efficient coexpression of two genes from a recombinant provirus in cultured cells and in embryos. Mol Cell Biol. 1991;11:5848–5859. doi: 10.1128/mcb.11.12.5848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Perret J, Ludgate M, Libert F, Gerard C, Dumont JE, Vassart G, Parmentier M. Stable expression of the human TSH receptor in CHO cells and characterization of differentially expressing clones. Biochem Biophys Res Commun. 1990;17:1044–1050. doi: 10.1016/0006-291x(90)90789-p. [DOI] [PubMed] [Google Scholar]
- 43.Van Riper G, Siciliano S, Fischer PA, Meurer R, Springer MS, Rosen H. Characterization and species distribution of high affinity GTP-coupled receptors for human rantes and monocyte chemoattractant protein 1. J Exp Med. 1993;177:851–856. doi: 10.1084/jem.177.3.851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Endres MJ, Clapham PR, Marsh M, Ahuja M, Turner JD, McKnight A, Thomas JF, Stoebenau-Haggarty B, Choe S, Vance PJ, et al. CD4-independent infection by HIV-2 is mediated by fusin/CXCR4. Cell. 1996;87:745–756. doi: 10.1016/s0092-8674(00)81393-8. [DOI] [PubMed] [Google Scholar]
- 45.Wu L, Ruffing N, Shi X, Newman W, Soler D, Mackay CR, Qin S. Discrete steps in binding and signaling of interleukin-8 with its receptor. J Biol Chem. 1996;271:31202–31209. doi: 10.1074/jbc.271.49.31202. [DOI] [PubMed] [Google Scholar]
- 46.Connor RI, Chen BK, Choe S, Landau NR. Vpr is required for efficient replication of human immunodeficiency virus type-1 in mononuclear phagocytes. Virology. 1995;206:935–944. doi: 10.1006/viro.1995.1016. [DOI] [PubMed] [Google Scholar]
- 47.Hill CM, Deng H-K, Unutmaz D, Kewalramani V, Bastiani L, Zolla-Pazner S, Littman DR. Envelope glycoproteins from HIV-1, HIV-2, and SIV can use human CCR5 as a cofactor for viral entry and make direct CD4- dependent interactions with this chemokine receptor. J Virol. 1997;71:6296–6304. doi: 10.1128/jvi.71.9.6296-6304.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Bleul CC, Wu L, Hoxie JA, Springer TA, Mackay CR. The chemokine receptors CXCR4 and CCR5 are differentially expressed and regulated on T cells. Proc Natl Acad Sci USA. 1997;94:1925–1930. doi: 10.1073/pnas.94.5.1925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Cocchi F, DeVico AL, Garzino-Demo A, Arya SK, Gallo RC, Lusso P. Identification of RANTES, MIP-1α, and MIP-1β as the major HIV-suppressive factors produced by CD8+T cells. Science (Wash DC) 1995;270:1811–1815. doi: 10.1126/science.270.5243.1811. [DOI] [PubMed] [Google Scholar]
- 50.Simmons G, Clapham PR, Picard L, Offord RE, Rosenkilde MM, Schwartz TW, Buser R, Wells TNC, Proudfoot AEI. Potent inhibition of HIV-1 infectivity in macrophages and lymphocytes by a novel CCR5 antagonist. Science (Wash DC) 1997;276:276–279. doi: 10.1126/science.276.5310.276. [DOI] [PubMed] [Google Scholar]
- 51.Arenzana-Seisdedos F, Virelizier JL, Rousset D, Clark-Lewis I, Loetscher P, Moser B, Baggiolini M. HIV blocked by chemokine antagonist. Science (Wash DC) 1996;383:400. doi: 10.1038/383400a0. [DOI] [PubMed] [Google Scholar]
- 51a.Samson, M., G. LaRosa, F. Libert, M. Detheux, P. Paindavoine, G. Vassart, and M. Parmentier. 1997. The second extracellular loop of CCR5 is the major determinant of ligand specificity. J. Biol. Chem. In press. [DOI] [PubMed]
- 52.Mackay CR. Chemokine receptors and T cell chemotaxis. J Exp Med. 1996;184:799–802. doi: 10.1084/jem.184.3.799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Roth SJ, Carr MW, Springer TA. C-C chemokines, but not the C-X-C chemokines interleukin-8 and interferon-γ–inducible protein-10, stimulate transendothelial chemotaxis of T lymphocytes. Eur J Immunol. 1995;25:3482–3488. doi: 10.1002/eji.1830251241. [DOI] [PubMed] [Google Scholar]