

Abstract
It has been believed that the Fas expressed on human peripheral blood T cells (PBT) is nonfunctional, because these cells are insensitive to agonistic anti-Fas/Apo-1 mAbs that efficiently kill in vitro–activated T cells and many Fas-expressing cell lines. Here, we demonstrate that membrane-bound Fas ligand (FasL) kills both fresh and in vitro–activated PBT, indicating that the Fas expressed on fresh PBT is functional. In contrast, soluble FasL kills only the latter. Naive T cells in umbilical cord blood do not express Fas, but can be induced to express Fas by IFN-γ or by a combination of IL-2 and anti-CD28 mAb, after which they acquire sensitivity to membrane but not to soluble FasL. Soluble FasL inhibited the killing of fresh PBT by membrane FasL. These results indicate that the shedding of FasL from the membrane is a mechanism for downregulating at least part of its killing activity.
Fas (also called Apo-1 or CD95) and Fas ligand (FasL) play crucial roles in homeostasis and self-tolerance of lymphocytes in both humans and mice (1). FasL also works as a cytotoxic effector molecule of CTL and NK cells. FasL is constitutively expressed in the testis and eyes, and it has been implicated in the property of immune privilege in these organs (2, 3). At the same time, the Fas-FasL system is a double-edged sword (4) in that both the diminution and enhancement of this system have pathogenic effects. Loss-of-function mutations of Fas and FasL result in lymphoproliferative and autoimmune diseases. FasL expressed on activated T cells seems partly responsible for the tissue damage in fulminant hepatitis (5) and graft-versus-host disease (6, 7). It has been reported that the thyrocytes in patients with Hashimoto disease express both Fas and FasL, and commit suicide similar to the activation-induced cell death of T cells (8). It has been suggested that the Fas-FasL system plays a role in the loss of CD4+ T cells in AIDS patients (9, 10). Therefore, strategies for up- or downregulating the Fas-FasL system might have therapeutic value for these diseases. However, since Fas is expressed rather ubiquitously in a wide range of normal tissues, the side effects of such treatments on normal cells should be carefully evaluated.
FasL is a 40-kD type II transmembrane protein that is homologous to TNF. FasL induces apoptosis by binding to its membrane receptor, Fas (11). TNF-α, which is also synthesized as a type II transmembrane protein, is cleaved into a 17-kD soluble form by proteolysis, and its soluble form causes cachexia. Recently, the novel disintegrin metalloprotease (TNF alpha-converting enzyme, TACE) that mediates this process was identified (12, 13). Similarly, the TNF-homologous portion of membrane FasL is cleaved into a 26-kD soluble form by a metalloprotease (14, 15), although it is unknown whether the FasL cleavage is also performed by TACE. This soluble human FasL efficiently induces apoptosis in some Fas-expressing cells. In contrast, mouse soluble FasL loses its killing activity (16).
The function of Fas expressed on various cells has been investigated using agonistic anti-Fas/Apo-1 mAbs, and the results have indicated that the expression of Fas does not necessarily mean that cells are susceptible to these mAbs (17, 18). A large fraction of the peripheral blood T cells (PBT) freshly isolated from adult humans express Fas. These Fas+ PBT correspond to CD45RO+ PBT cells, which have been postulated to be memory T cells. In spite of having readily detectable levels of Fas expression, these cells are totally resistant to agonistic anti-Fas mAbs (19–21). Virtually all murine T cells from peripheral lymphoid organs constitutively express Fas, but they are also resistant to anti-Fas mAb treatments (22). Therefore, it has been concluded that the Fas expressed on peripheral mature T cells is nonfunctional. However, we recently discovered that a recombinant soluble mouse FasL that carries nearly the complete extracellular region of FasL (termed WX1) efficiently kills mature T cells in freshly isolated mouse splenocytes or lymph node cells (16). It should be noticed that, unlike naturally processed mouse soluble FasL, WX1 is biologically active. In this study, we examined the susceptibility of human PBT to the membrane and soluble forms of human FasL and found that they exhibited differential cytotoxic activity.
Materials and Methods
Preparation of Adult Peripheral and Umbilical Cord Blood Lymphocytes.
Heparinized adult peripheral and cord blood were obtained from healthy adult volunteers and newborn babies delivered by Cesarean section, respectively, after obtaining informed consent. Samples were diluted with an equal volume of PBS and the lymphoid fraction was separated by gradient centrifugation using Ficoll-Paque Plus (Pharmacia, Uppsala, Sweden). ConA blasts were prepared from adult PBL by cultivation with 5 μg/ml of ConA (Sigma Chemical Co., St. Louis, MO) and 10 ng/ml of recombinant human IL-2 (Ajinomoto Pharmaceutical Co., Tokyo, Japan) for 48 h, followed by incubation in the presence of IL-2 alone for 5 d. Dead cells were removed by gradient centrifugation using Histopaque 1083 (Sigma). Cells were washed twice and suspended in RPMI 1640 medium (Nikken BioMedical Laboratory, Kyoto, Japan) supplemented with 10% FCS, 100 U/ml benzylpenicillin potassium (Meiji Seika Co., Tokyo, Japan), and 100 μg/ml streptomycin sulfate (Meiji Seika). To induce Fas expression in cord blood lymphocytes, cells were cultured for 20 h with recombinant human IFN-γ (10 ng/ml; PeproTech Inc., Rocky Hill, NJ), IL-2 (10 ng/ml), or in a plate coated with anti-CD28 mAb (PharMingen, San Diego, CA) at 10 μg/ml in 10 mM Tris-HCl, pH 8.0, at 37°C for 1 h. Further analyses of the expression and function of Fas in these cells were performed as described below.
Induction of Apoptosis by Transformants Expressing FasL, Soluble FasL, and Anti-Fas Monoclonal Antibody.
Transformant 1A12, expressing human FasL lacking a cytoplasmic region (amino acids 8-69), was established as previously described (23). Soluble human FasL was purified from the culture medium of the 1A12 cells using an affinity column coupled with anti-human FasL mAb (4H9) (24), and the purified protein was designated as AL-1. Recombinant soluble mouse FasL (WX1) consisting of the signal sequence derived from G-CSF and almost the entire extracellular region of mouse FasL (amino acid 101-279) was prepared as described previously (16), and purified using an anti-mouse FasL mAb affinity column. Anti-human Fas IgM mAb (CH11) was purchased from MBL (Nagoya, Japan). To investigate the susceptibility of the cells to soluble FasL or anti-Fas mAb, 2 × 105 fresh PBL or 6 × 105 ConA blasts were cultured with various concentrations of WX1, AL-1, and CH11 in 200 μl of culture medium in a flat-bottomed 96-well culture plate for 14 h at 37°C in a 5% CO2, humidified atmosphere. The same number of fresh PBL were cocultured with 1A12 cells at various effector/target ratios for 12 h. In some experiments, 1A12 cells were preincubated with 10 μM BB2116 matrix metalloprotease inhibitor (kindly provided by Dr. A.H. Drummond, British Biotech Pharmaceuticals, Oxford, UK) for 4 h, and it was continuously included in the following assay to obstruct the cleavage of membrane FasL.
Evaluation of Fas Expression and Cell Death by Flow Cytometry.
After incubations to induce apoptosis, aliquots of the cells were stained with optimal doses of FITC-conjugated anti-Fas mAb (UB-2, MBL), PE-conjugated mAbs against either CD3, CD4, or CD8 (Becton Dickinson, San Jose, CA), and 2 μg/ml of propidium iodide (PI) in 50 μl of staining solution for 30 min on ice, and then washed 3 times with staining solution. These cells were immediately analyzed in a FACScan® (Becton Dickinson) equipped with a 488-nm argon laser, and the data were analyzed using CELLQuest software. Percent specific cell killing was calculated as follows: % specific cell killing = (% PI+ cells − % spontaneous PI+ cells in medium only) / (100 − % spontaneous PI+ cells in medium only).
Results
WX1, but Not Soluble Human FasL or Anti-Fas mAb Kills Freshly Isolated PBL.
About 50% of CD4+ and CD8+ T cells in freshly isolated PBL from a healthy adult volunteer and virtually all ConA blasts generated from the PBL expressed readily detectable levels of Fas (data not shown). Consistent with previous reports, agonistic anti-Fas mAb (CH11) efficiently killed ConA blasts, whereas fresh PBT were resistant to the mAb (Fig. 1 a). We previously established a transfectant that expresses a human FasL lacking most of its cytoplasmic region. This cell line (1A12) secretes soluble FasL by a proteolytic mechanism (23). The soluble FasL was purified using an affinity column with anti-human FasL mAb. This purified soluble human FasL (named AL-1) showed similar target specificity to CH11 (Fig. 1 b). In contrast, WX1, the biologically active, recombinant soluble mouse FasL, killed both ConA blasts and fresh PBT (Fig. 1, c–g). Since WX1 specifically killed Fas+ cells in PBT, which represent ∼50% of the total PBT (Fig. 1, d and e), the dose-response curves of WX1-induced death in ConA blasts and fresh Fas+ PBT are comparable. There was no difference between CD4 and CD8 T cells in terms of susceptibility to various Fas agonists.
Figure 1.
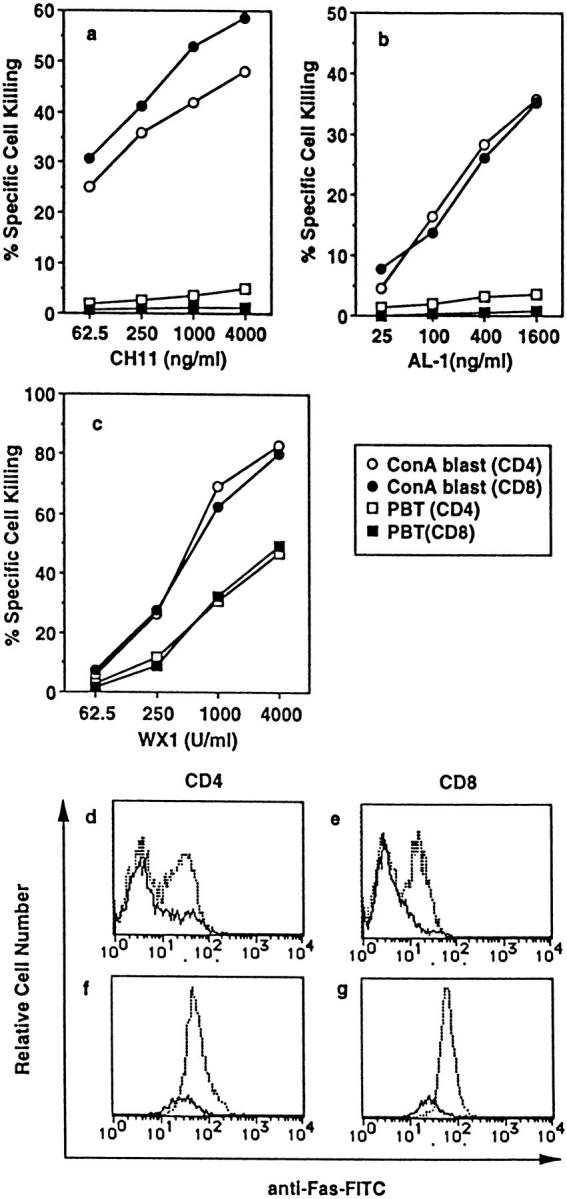
Susceptibility of fresh PBT and ConA blasts to anti-Fas mAb (CH11), soluble human FasL (AL-1), and recombinant soluble mouse FasL (WX1). (a–c), Fresh PBL or ConA blasts were cultured in the presence of the indicated concentrations of CH11 (a), AL-1 (b), and WX1 (c) for 14 h. Cells were then stained with FITC-anti-Fas mAb, PE-conjugated mAb against either CD4 or CD8, and PI. Percent specific cell killing of CD4+ and CD8+ cells was determined as described in Materials and Methods. (d–g) Fresh PBL (d and e) and ConA blasts (f and g) cultured with (solid line) or without (dotted line) 4,000 units/ml of WX1 for 14 h were stained as described above, and 5 × 104 total cells (including both viable and dead cells) were analyzed in a FACScan®. Staining profiles for FITC-anti-Fas mAb of viable (PI−) CD4+ (d and f) or CD8+ cells (e and g) are shown.
Membrane FasL Kills Freshly Isolated PBL, and Soluble FasL Inhibits This Activity.
The cytotoxicity of membrane FasL to fresh PBT was investigated using a transfectant (1A12) expressing human FasL and its parental cell line (WR19L). The metalloprotease inhibitor BB2116 prevents proteolytic secretion of FasL from 1A12 cells (23). As shown in Fig. 2 a, 1A12 killed fresh PBT in a dose-dependent manner, and BB2116 dramatically enhanced the cytotoxicity of 1A12. WR19L showed no cytotoxicity even in the presence of BB2116. BB2116 did not enhance the cytotoxicity of WX1 (data not shown), excluding the possibility that the weak toxicity of BB2116 rather than its protease inhibitor activity caused the enhancement of 1A12-mediated killing. Fas+ PBT were specifically killed by 1A12 (Fig. 2, b–d). We then tested whether soluble FasL inhibits the cytotoxicity of 1A12 against PBT. As shown in Fig. 2 e, exogenously added soluble FasL inhibited the cytotoxicity of 1A12 in the presence of BB2116 in a dose-dependent manner. Similarly, anti-Fas mAb inhibited the killing of PBT by 1A12 (Fig. 2 f). These results indicate that membrane FasL is capable of killing fresh Fas+ PBT, whereas soluble FasL and anti-Fas mAb behave as Fas-antagonists for these cells under the conditions used.
Figure 2.

Membrane FasL kills fresh PBT and soluble FasL inhibits the killing. (a) Fresh PBL were cultured with the transfectant expressing human FasL (1A12) or its parental cell line (WR19L) at the indicated effector/target (E/T) ratios for 12 h in the presence or absence of 10 μM BB2116. Cells were then stained with FITC anti-Fas, PE-anti-CD3 mAb and PI, and analyzed in a FACScan®. (b–d) FITC anti-Fas mAb staining profiles for PI−CD3+ cells after culturing with WR19L in the absence of BB2116 (b), or with 1A12 in the absence (c) or presence (d) of BB2116 at an E/T ratio of 1.0 are shown. Dotted profiles indicate negative controls stained with FITC-labeled mouse IgG1. (e and f). Fresh PBL were cultured with 1A12 plus 10 mM BB2116 or with WR19L cells at the E/T ratio of 1.0 in the presence of the indicated concentrations of purified AL-1 (e) or CH11 (f) for 12 h.
IFN-γ or a Combination of IL-2 and Immobilized Anti-CD28 mAb Induces Fas Expression and Susceptibility to 1A12 Cells and WX1, but Not to AL-1 or CH11 in PBL.
Essentially all cord blood T cells (CBT) are CD45RO− naive T cells and do not express Fas at detectable levels. Accordingly, they are not killed by any form of FasL or anti-Fas mAb (data not shown). Previously, it was reported that CBT cultured with phytohemagglutinin for 5 d express Fas (20). Here, we found that IFN-γ induces Fas expression on both CD4+ and CD8+ CBT within 20 h (Fig. 3, a and e). IL-2 alone induced FasL weakly, while other stimuli including IL-1, IL-4, IL-6, TNF-α, and immobilized anti-CD28 mAb did not, when added separately (Fig. 3, b, c, f, and g, and data not shown). However, a combination of IL-2 and anti-CD28 mAb showed Fas-inducing activity comparable to IFN-γ (Fig. 3, d and h). The levels of Fas expression induced by IFN-γ plus anti-CD28 mAb were not much different from those induced by IFN-γ alone (data not shown).
Figure 3.

Function of Fas on CBT induced by IFN-γ or a combination of IL-2 and immobilized anti-CD28 mAb. (a–h) Cord blood lymphocytes were cultured for 20 h with medium only (dotted line in each panel), IFN–γ (10 ng/ml, a and e), IL-2 (10 ng/ml, b and f), immobilized anti-CD28 mAb (coated on plate at 10 μg/ml, c and g), or IL-2 plus immobilized anti-CD28 mAb (d and h), respectively. Cells were then stained with FITC–anti-Fas mAb, PE-conjugated mAb against either CD4 or CD8, and PI. FITC–anti-Fas mAb staining profiles of viable (PI−) CD4+ (a–d) or CD8+ cells (e–h) are shown. (i and j) CBT treated with IFN-γ or with IL-2 plus anti-CD28 mAb as described above were cultured with CH11, WX1, AL-1, or 1A12.
Finally, we tested the susceptibility of Fas-induced CBT to various forms of FasL and anti-FasL mAb. As shown in Fig. 3, i and j, CBT treated with IFN-γ or a combination of IL-2 and anti-CD28 mAb were killed by 1A12 cells in the presence of BB2116 and WX1, whereas the same cells were resistant to AL-1 and CH11.
Discussion
We have shown that the membrane form of FasL is capable of killing Fas+ PBT. The Fas+ fraction of PBT corresponds to CD45RO+ memory T cells (20). In contrast, naive T cells in adult or cord blood do not express Fas, hence they are not killed by any types of FasL or anti-Fas mAb. However, IFN-γ and a combination of IL-2 and anti-CD28 mAb rapidly induce Fas expression in naive T cells from cord blood, and as a result, these cells become sensitive to membrane FasL. We have previously reported that freshly isolated mouse T cells from peripheral lymphoid organs, which are resistant to anti-mouse Fas mAb, are susceptible to WX1 (16). Collectively, these results demonstrate that the Fas expressed on the peripheral T cells of humans and mice is functional. The cytotoxicity of membrane FasL against peripheral T cells is not specific to transfectants expressing high levels of FasL, because activated T cells from B10-Thy1a (Thy1.1) mice induced cell death in naive T cells from B10 (Thy1.2) mice in a FasL-dependent manner (data not shown). Why should fresh PBT, which are potentially useful cells, be killed by membrane FasL? T cells are activated through TCR in an Ag-specific manner. However, once T cells are activated, they produce a variety of cytokines and cell surface costimulative molecules that can activate bystander T cells in an Ag-nonspecific manner. Therefore, when some naive or memory T cells with inappropriate Ag-specificity accidentally encounter activated T cells at an inflammatory site, they should be killed, probably through the FasL expressed by activated T cells; otherwise, they would be activated Ag-nonspecifically. We previously demonstrated that TCR engagement induces FasL-resistance in mouse splenic T cells. Therefore, naive or memory T cells with appropriate TCR, which are thereby acquiring antigenic signals, would not be killed by activated T cells (16).
Soluble human FasL is biologically active in that it has potent cytotoxicity against some Fas-expressing cell lines (14). In contrast, its mouse counterpart is inactive. Since Fas is constitutively expressed in various tissues, if soluble FasL is fully functional, it may be detrimental to humans. Here, we found that soluble human FasL possesses a selective killing activity. Potentially useful PBT are not killed by AL-1. In contrast, ConA blasts are killed by AL-1. The latter cells may represent useless or even dangerous senescent activated T cells. Human recombinant soluble FasL produced in yeast, which has a target selectivity similar to naturally processed soluble FasL (data not shown), induces lethal hepatic failure, but only when a high dose (500 μg/mouse) of the FasL is intravenously injected in mice (24). In contrast, the injection of a small number of CTL can induce lethal hepatitis in mice (25), and the lethal activity is mediated by FasL (5). These results suggest that the cytotoxicity of soluble FasL against hepatocytes is also limited, compared with that of membrane FasL. This notion has been confirmed by an in vitro cytotoxicity assay against primary culture hepatocytes (Tanaka, M., and S. Nagata, manuscript in preparation). It has been shown that membrane TNF-α is far more efficient than soluble TNF-α in activating type II TNF receptors (26). These results indicate that the membrane forms of FasL and TNFα are the primary activators of their receptors. We discovered that soluble human FasL competitively inhibits the killing of PBT by membrane FasL. Collectively, these results indicate that the cleaving of membrane ligands is a mechanism that partly downregulates their activities.
Why do membrane and soluble FasL have different sets of target cells? In relation to this phenomenon, various Fas agonists including soluble and membrane FasL, and anti-Fas mAbs, can be separated into two groups based on their killing activity against fresh PBT and ConA blasts. Membrane FasL and WX1 kill both fresh PBT and ConA blasts, whereas “agonistic” anti-Fas mAbs and AL-1 kill ConA-activated T cells but not fresh PBT. Furthermore, the latter group inhibits the killing activity of the first group, indicating that they can bind to Fas on fresh PBT. Since both AL-1 and WX1 are soluble proteins, solubility is not the reason for the different activity of the two groups. The fact that purified WX1 can kill fresh PBT makes it unlikely that an unidentified molecule coexpressed with membrane FasL on 1A12 cells is required as a costimulative molecule to facilitate the FasL-mediated death of fresh PBT. Analytical gel filtration indicates that AL-1 exists as a trimer (Tanaka, M., and S. Nagata, unpublished observation) whereas WX1 mostly exists as an oligomer larger than a decamer (16). Therefore, it is possible that the trimerization of Fas is sufficient for it to kill ConA blasts, whereas the formation of larger complexes of Fas is required for it to kill fresh PBT. However, this possibility seems unlikely because the anti-human Fas mAb, CH11, which belongs to the second group, is a decavalent IgM Ab. Furthermore, the super-cross-linking of Fas on PBT by sequential treatments with CH11 and anti-mouse IgM antibodies induces only minimal death in CD4+ cells, and it has no effect on CD8+ cells (data not shown). The affinities or avidities of various agonists for Fas may determine their spectrum of target cells. Recently, Fadeel et al. (27) reported that various anti-Fas mAbs that recognize the same epitope exhibit different effects on Fas-mediated apoptosis, and that those mAbs with a moderate affinity killed Fas-expressing cells, while high-affinity Abs were antagonistic and low-affinity Abs showed no biological effect. However, the reason why AL-1 inhibits the cytotoxicity of membrane FasL is probably not because AL-1 has a very high affinity, since rather large doses of AL-1 were required to inhibit cytotoxicity of membrane FasL. Alternatively, membrane FasL and WX1 but not anti-Fas mAbs or AL-1 may interact with, in addition to Fas, another molecule expressed in fresh PBT, and this interaction may be necessary to kill fresh PBT. Since WX1, which is a soluble protein, somehow mimics membrane FasL rather than soluble FasL, a careful comparison of WX1 and AL-1 may elucidate this possibility. WX1 is also a useful tool for investigating the Fas sensitivity of a group of cells that are only killed by membrane FasL or WX1.
Since agonistic anti-Fas/Apo-1 mAbs show strong cytotoxicity against a variety of tumors (28, 29) or abnormally proliferating synovial cells in rheumatoid arthritis (30), a possible therapeutic value of these mAbs for these diseases has been discussed. The main concern has been the toxicity of these molecules against normal cells. Our finding that some soluble FasL and an anti-Fas mAb show selective killing activity opens up the possibility that some tumor cells or abnormally activated cells are more sensitive than normal cells to these molecules. If this is the case, these molecules at a sublethal dose may be therapeutic for tumor or rheumatoid arthritis patients.
Acknowledgments
We thank Dr. N. Matuzaki (Ikeda City Hospital, Department of Obstetrics and Gynecology) for kindly providing natal umbilical cord blood, and Ms. H. Fujiwara for secretarial assistance.
This work was supported in part by grants-in-aid from the Ministry of Education, Science and Culture of Japan, by Special Coordination Funds of the Science and Technology Agency of the Japanese Government to T. Suda and S. Nagata, and by the Program for Promotion of Fundamental Studies in Health Sciences of the Organization for Drug ADR Relief, R&D Promotion and Product Review of Japan to T. Ochi.
Footnotes
The first two authors contributed equally to this manuscript.
References
- 1.Nagata S, Suda T. Fas and Fas ligand: lpr and gldmutations. Immunol Today. 1995;16:39–43. doi: 10.1016/0167-5699(95)80069-7. [DOI] [PubMed] [Google Scholar]
- 2.Bellgrau D, Gold D, Selawry H, Moore J, Franzusoff A, Duke RC. A role for CD95 ligand in preventing graft rejection. Nature. 1995;377:630–632. doi: 10.1038/377630a0. [DOI] [PubMed] [Google Scholar]
- 3.Griffith TS, Brunner T, Fletcher SM, Green DR, Ferguson TA. Fas ligand-induced apoptosis as a mechanism of immune privilege. Science. 1995;270:1189–1192. doi: 10.1126/science.270.5239.1189. [DOI] [PubMed] [Google Scholar]
- 4.Nagata S. Apoptosis by death factor. Cell. 1997;88:355–365. doi: 10.1016/s0092-8674(00)81874-7. [DOI] [PubMed] [Google Scholar]
- 5.Kondo T, Suda T, Fukuyama H, Adachi M, Nagata S. Essential roles of the Fas ligand in the development of hepatitis. Nat Med. 1997;3:409–413. doi: 10.1038/nm0497-409. [DOI] [PubMed] [Google Scholar]
- 6.Via CS, Nguyen P, Shustov A, Drappa J, Elkon KB. A major role for the Fas pathway in acute graft-versus-host disease. J Immunol. 1996;157:5387–5393. [PubMed] [Google Scholar]
- 7.Braun MY, Lowin B, French L, Acha OH, Tschopp J. Cytotoxic T cells deficient in both functional fas ligand and perforin show residual cytolytic activity yet lose their capacity to induce lethal acute graft-versus-host disease. J Exp Med. 1996;183:657–661. doi: 10.1084/jem.183.2.657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Giordano C, Stassi G, De MR, Todaro M, Richiusa P, Papoff G, Ruberti G, Bagnasco M, Testi R, Galluzzo A. Potential involvement of Fas and its ligand in the pathogenesis of Hashimoto's thyroiditis. Science. 1997;275:960–963. doi: 10.1126/science.275.5302.960. [DOI] [PubMed] [Google Scholar]
- 9.Sloand EM, Young NS, Kumar P, Weichold FF, Sato T, Maciejewski JP. Role of Fas ligand and receptor in the mechanism of T-cell depletion in acquired immunodeficiency syndrome: effect on CD4+lymphocyte depletion and human immunodeficiency virus replication. Blood. 1997;89:1357–1363. [PubMed] [Google Scholar]
- 10.Silvestris F, Cafforio P, Frassanito MA, Tucci M, Romito A, Nagata S, Dammacco F. Overexpression of Fas antigen on T cells in advanced HIV-1 infection: differential ligation constantly induces apoptosis. AIDS. 1996;10:131–141. doi: 10.1097/00002030-199602000-00002. [DOI] [PubMed] [Google Scholar]
- 11.Suda T, Takahashi T, Golstein P, Nagata S. Molecular cloning and expression of the Fas ligand: a novel member of the tumor necrosis factor family. Cell. 1993;75:1169–1178. doi: 10.1016/0092-8674(93)90326-l. [DOI] [PubMed] [Google Scholar]
- 12.Black RA, Rauch CT, Kozlosky CJ, Peschon JJ, Slack JL, Wolfson MF, Castner BJ, Stocking KL, Reddy P, Srinivasan S, et al. A metalloproteinase disintegrin that releases tumour-necrosis factor- alpha from cells. Nature. 1997;385:729–733. doi: 10.1038/385729a0. [DOI] [PubMed] [Google Scholar]
- 13.Moss ML, Jin SL, Milla ME, Burkhart W, Carter HL, Chen WJ, Clay WC, Didsbury JR, Hassler D, Hoffman CR, et al. Cloning of a disintegrin metalloproteinase that processes precursor tumour-necrosis factor-alpha. Nature. 1997;385:733–736. doi: 10.1038/385733a0. [DOI] [PubMed] [Google Scholar]
- 14.Tanaka M, Suda T, Takahashi T, Nagata S. Expression of the functional soluble form of human Fas ligand in activated lymphocytes. EMBO (Eur Mol Biol Organ) J. 1995;14:1129–1135. doi: 10.1002/j.1460-2075.1995.tb07096.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kayagaki N, Kawasaki A, Ebata T, Ohmoto H, Ikeda S, Inoue S, Yoshino K, Okumura K, Yagita H. Metalloproteinase-mediated release of human Fas ligand. J Exp Med. 1995;182:1777–1783. doi: 10.1084/jem.182.6.1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Suda T, Tanaka M, Miwa K, Nagata S. Apoptosis of mouse naive T cells induced by recombinant soluble Fas ligand and activation-induced resistance to Fas ligand. J Immunol. 1996;157:3918–3924. [PubMed] [Google Scholar]
- 17.Wong GHW, Goeddel DV. Fas antigen and p55 TNF receptor signal apoptosis through distinct pathways. J Immunol. 1994;152:1751–1755. [PubMed] [Google Scholar]
- 18.Shima Y, Nishimoto N, Ogata A, Fujii Y, Yoshizaki K, Kishimoto T. Myeloma cells express Fas antigen/APO-1 (CD95) but only some are sensitive to anti-Fas antibody resulting in apoptosis. Blood. 1995;85:757–764. [PubMed] [Google Scholar]
- 19.Owen-Schaub LB, Yonehara S, Crump WL, III, Grimm EA. DNA fragmentation and cell death is selectively triggered in activated human lymphocytes by Fas antigen engagement. Cell Immunol. 1992;140:197–205. doi: 10.1016/0008-8749(92)90187-t. [DOI] [PubMed] [Google Scholar]
- 20.Miyawaki T, Uehara T, Nibu R, Tsuji T, Yachie A, Yonehara S, Taniguchi N. Differential expression of apoptosis-related Fas antigen on lymphocyte subpopulations in human peripheral blood. J Immunol. 1992;149:3753–3758. [PubMed] [Google Scholar]
- 21.Robertson MJ, Manley TJ, Pichert G, Cameron C, Cochran KJ, Levine H, Ritz J. Functional consequences of APO-1/Fas (CD95) antigen expression by normal and neoplastic hematopoietic cells. Leuk Lymphoma. 1995;17:51–61. doi: 10.3109/10428199509051703. [DOI] [PubMed] [Google Scholar]
- 22.Nishimura Y, Ishii A, Kobayashi Y, Yamasaki Y, Yonehara S. Expression and function of mouse Fas antigen on immature and mature T cells. J Immunol. 1995;154:4395–4403. [PubMed] [Google Scholar]
- 23.Tanaka M, Suda T, Haze K, Nakamura N, Sato K, Kimura F, Motoyoshi K, Mizuki M, Tagawa S, Ohga S, et al. Fas ligand in human serum. Nat Med. 1996;2:317–322. doi: 10.1038/nm0396-317. [DOI] [PubMed] [Google Scholar]
- 24.Tanaka M, Suda T, Yatomi T, Nakamura N, Nagata S. Lethal effect of recombinant human Fas ligand in mice pretreated with Propionibacterium acnes. . J Immunol. 1997;158:2303–2309. [PubMed] [Google Scholar]
- 25.Ando K, Moriyama T, Guidotti LG, Wirth S, Schreiber RD, Schlicht HJ, Huang S, Chisari FV. Mechanisms of class I restricted immunopathology. A transgenic mouse model of fulminant hepatitis. J Exp Med. 1993;178:1541–1554. doi: 10.1084/jem.178.5.1541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Grell M, Douni E, Wajant H, Lohden M, Clauss M, Maxeiner B, Georgopoulos S, Lesslauer W, Kollias G, Pfizenmaier K, Scheurich P. The transmembrane form of tumor necrosis factor is the prime activating ligand of the 80 kDa tumor necrosis factor receptor. Cell. 1995;83:793–802. doi: 10.1016/0092-8674(95)90192-2. [DOI] [PubMed] [Google Scholar]
- 27.Fadeel B, Thorpe CJ, Yonehara S, Chiodi F. Anti-Fas IgG1 antibodies recognizing the same epitope of Fas/APO-1 mediate different biological effects in vitro. Int Immunol. 1997;9:201–209. doi: 10.1093/intimm/9.2.201. [DOI] [PubMed] [Google Scholar]
- 28.Trauth BC, Klas C, Peters AMJ, Matzuku S, Möller P, Falk W, Debatin K-M, Krammer PH. Monoclonal antibody-mediated tumor regression by induction of apoptosis. Science. 1989;245:301–305. doi: 10.1126/science.2787530. [DOI] [PubMed] [Google Scholar]
- 29.Owen-Schaub LB, Radinsky R, Kruzel E, Berry K, Yonehara S. Anti-Fas on nonhematopoietic tumors: Levels of Fas/APO-1 and bcl-2 are not predictive of biological responsiveness. Cancer Res. 1994;54:1580–1586. [PubMed] [Google Scholar]
- 30.Fujisawa K, Asahara H, Okamoto K, Aono H, Hasunuma T, Kobata T, Iwakura Y, Yonehara S, Sumida T, Nishioka K. Therapeutic effect of the anti-Fas antibody on arthritis in HTLV-1 tax transgenic mice. J Clin Invest. 1996;98:271–278. doi: 10.1172/JCI118789. [DOI] [PMC free article] [PubMed] [Google Scholar]