

Abstract
Type I interferons α and β, naturally produced regulators of cell growth and differentiation, have been shown to inhibit IL-7–induced growth and survival of B cell precursors in vitro. After confirming an inhibitory effect on B lymphopoiesis in an ex vivo assay, we treated newborn mice with an active IFN-α2/α1 hybrid molecule to assess its potential for regulating B and T cell development in vivo. Bone marrow and splenic cellularity was greatly reduced in the IFN-α2/α1–treated mice, and B lineage cells were reduced by >80%. The bone marrow progenitor population of CD43+B220+HSA− cells was unaffected, but development of the CD19+ pro–B cells and their B lineage progeny was severely impaired. Correspondingly, IL-7–responsive cells in the bone marrow were virtually eliminated by the interferon treatment. Thymus cellularity was also reduced by >80% in the treated mice. Phenotypic analysis of the residual thymocytes indicated that the inhibitory effect was exerted during the pro–T cell stage in differentiation. In IFN-α/β receptor−/− mice, T and B cell development were unaffected by the IFN-α2/α1 treatment. The data suggest that type I interferons can reversibly inhibit early T and B cell development by opposing the essential IL-7 response.
Band T cell development proceeds in an ordered fashion that can be monitored by the sequential acquisition of gene rearrangements and cell surface markers. B cell development progresses through a series of differentiation and maturation stages in the bone marrow during which sequential rearrangements of immunoglobulin heavy and light chain genes occur (1–3). Via rearrangement of VH, DH, and JH gene elements, B cell progenitors differentiate into pre–B cells that express μ heavy chains in association with the surrogate light chains, encoded by λ5 and VpreB genes, and Igα/β heterodimers (4–6). This progression can be monitored by the differential expression of cell surface markers (7–10). The progenitor and precursor populations can be divided into four fractions according to the expression of CD45R/B220, leukosialin (CD43), heat-stable antigen (HSA),1 and BP-1: fraction A (B220+CD43+HSA−BP-1−); fraction B (B220+CD43+HSA+BP-1−); fraction C (B220+CD43+HSA+BP-1+); and fraction D (B220+CD43− HSA+BP-1+) (9, 11). Early B cell development thus appears to proceed in an ordered sequence from fraction A to fraction D (9). Fraction D cells further develop to IgM+ immature B cells, and finally into IgM+ IgD+ mature B cells.
T lymphocyte development in the thymus correspondingly proceeds from immature CD4−CD8− thymocytes to mature MHC-restricted single-positive CD4+ and CD8+ T cells through positive and negative selection after TCR gene rearrangement (12–14). The CD4−CD8− double negative (DN) population contains multiple progenitor subsets that are distinguishable by their differential expression of CD44 and CD25 (15). The following maturation sequence has been proposed for DN cells: CD4−CD8− CD44+CD25− → CD4−CD8−CD44+CD25+ → CD4− CD8−CD44−CD25+ → CD4−CD8−CD44−CD25− (15, 16). TCR-β gene rearrangements that precede rearrangements in the TCR-α locus begin in the CD44−CD25+ or CD44+CD25+ stages (17–19) with a functional β chain being required for the transition of CD44−CD25+ to the CD44−CD25− DN cell stage (19).
Both B and T cells are developmentally dependent on IL-7 in the mouse (20–26). In contrast, the type I interferons, IFN-α and -β, produced by resident bone marrow macrophages, can inhibit the IL-7–induced growth and survival of early B lineage cells in vitro (27). In vitro analysis also suggests that IFN-α/β can interfere with thymocyte development (28). Although IFN-α/β is well known as a naturally produced regulator of cell growth and differentiation (29–34), few studies have analyzed the potential for an in vivo role of IFN-α/β on development of the immune system. For this purpose, we examined the effects of recombinant human IFN-α2/α1, mutated so as to be active in mice (35). The results indicate that IFN treatment severely inhibits both B and T cell development at comparably early stages, but has no demonstrable effect in mice deficient for the IFN-α/β receptor.
Materials and Methods
Mice and IFN-α2/α1 Treatment.
BALB/c mice purchased from The Jackson Laboratory (Bar Harbor, ME) and wild-type and IFN-α/β receptor−/− 129Sv/Ev mice from B & K Universal Group Ltd. (North Humberside, UK) were bred in rooms equipped with an air filtering system. Cages, bedding, water, and food were sterilized, and the mice were handled with aseptic gloves in sterile hoods. Recombinant human IFN-α2/α1 (35), provided by Dr. Charles Weissman (Institut für Molekularbiologie I, Universität Zürich, Zurich, Switzerland), was intraperitoneally injected. Injections (4 × 105 U) were begun on day 3 and were continued for six days. Mice were killed one day after the final injection. In other experiments designed to examine the recovery from interferon treatment, mice were killed 6, 15, 21, and 28 d after the final injection. Control mice received PBS containing 10 μg/ml mouse serum albumin. The IFN-α2/α1 preparation was tested for the presence of endotoxin (LAL endotoxin testing; BioWhittaker, Inc., Walkersville, MD) and found to contain <10 pg/5 × 106 U.
Antibodies.
Cy-Chrome–labeled anti-B220, FITC-labeled anti-S7/CD43, FITC-labeled anti-CD3, FITC-labeled anti-CD4, FITC-labeled anti-CD8, FITC-labeled anti-Ly-6C, PE-labeled anti-μ, PE-labeled anti-CD44, PE-labeled anti-CD8, PE-labeled DX5, biotin-labeled anti-CD4, biotin-labeled anti-CD25 antibodies, biotin-labeled CD19, and APC-labeled B220 were purchased from PharMingen (San Diego, CA). PE-labeled BP-1, FITC-labeled anti-μ, streptavidin (SA)-Cy-Chrome, and SA-PE were obtained from Southern Biotechnology Associates (Birmingham, AL). SA-PerCP was purchased from Becton-Dickinson (Mountain View, CA).
Flow Cytometry.
Single cell suspensions were prepared from bone marrow and lymphoid tissues, and the nucleated cells were enumerated. Cells stained first with FITC-, PE-, Cy-Chrome–, or biotin-conjugated monoclonal antibodies on ice for 15 min were washed with 1% bovine serum albumin/PBS, and counterstained with SA-Cy-Chrome or SA-PE to reveal biotin conjugates before analysis with Becton-Dickinson FACScan® and FACSCalibur® flow cytometers. The data were analyzed with the Winlist 2.01 (Verity Software House, La Jolla, CA) and WinMDI 2.3 (Trotter@scripps.edu) software programs.
Proliferation Assay.
IL-7, produced by transfecting COS cells with a murine IL-7 expression vector (provided by Dr. Linda Park, Immunex, Seattle, WA), was assayed using an IL-7–dependent cell line, SCID 7, and recombinant IL-7 (Genzyme, Boston, MA) as a standard. Bone marrow cells were cultured in 96-well flat-bottomed plates (105 cells/ml) for 72 h in the absence or presence of IL-7 (0.1–100 ng/ml). Cells were also cultured with or without LPS (20 μg/ml; Sigma Chemical Co., St. Louis, MO). Cells were pulsed with 1 μCi (37 kBq) of [3H]thymidine for the last 8 h, and the incorporated [3H]thymidine was measured for quadruplicate cultures with a liquid scintillation counter.
In vitro B Lymphopoiesis Assay.
The B lymphopoiesis assay, provided by Dr. John F. Kearney (University of Alabama at Birmingham, Birmingham, AL), employed mononuclear cells from 15-d-old embryos. The cells (106/well) were suspended in IL-7– conditioned medium, consisting of 20% supernatant from IL-7– transfected T220 fibroblasts (36) and 80% fresh RPMI 1640 medium with 5% FCS. Cells were then cultured with the T220 transfectants seeded in 6-well plates. IFN-α2/α1 (103 U/ml) was added to the original culture medium and half of the medium was exchanged for fresh medium containing 103 U/ml IFN-α2/α1 every 7 d. Cells were analyzed by flow cytometry on day 0, day 5, day 10, and day 15.
Results
IFN-α2/α1 Inhibition of B Cell Development by Fetal Liver Progenitors.
The effect of IFN-α/β treatment on B cell development was initially examined by incubating fetal liver–derived cells with IFN-α2/α1 (35). The activity of the human IFN-α2/α1 preparation was evaluated in a comparative analysis with mouse IFN-α/β to confirm similar inhibitory effects on IL-7–dependent cells and the absence of significant effect on other types of cytokine-dependent cells (reference 27 and data not shown). B lymphopoiesis by fetal liver progenitors was then monitored by the expression of B220 and IgM. At the time of culture initiation, 18% of the fetal liver cells were B220+ (Fig. 1). After 5 d of culture with the IL-7 fibroblast transfectants, nearly 90% of the cells in control cultures were B220+, whereas ∼50% of the IFN-α2/α1–treated cells were B220+. On days 10 and 15, 25% and 45% of the cells were IgM+B220+ B cells in the control cultures, respectively. By contrast, <10% of the IFN-α2/α1–treated cells were IgM+ by day 15. These results indicate that IFN-α2/α1 is an effective inhibitor of B lymphopoiesis ex vivo.
Figure 1.
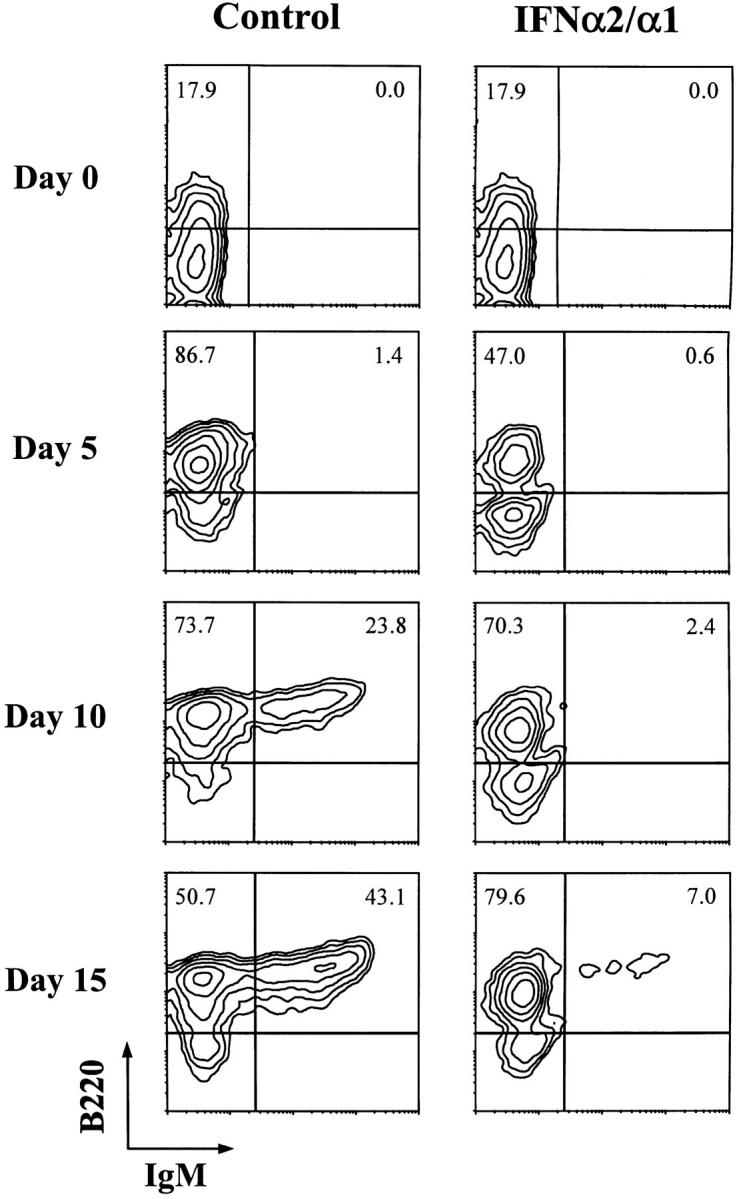
In vitro analysis of B lymphopoiesis by fetal liver progenitor cells in the presence or absence of IFN-α2/α1. IFN-α2/α1 (103 U/ml) was added at the time of culture initiation and half of the medium was exchanged for fresh medium containing 103 U/ml IFN-α2/α1 every 7 d. B220 and IgM expression was examined for the fetal liver cells derived from day 15 embryos. The cells were cultured over a confluent layer of IL-7–transfected T220 cells. Percentages of B220+ and IgM+ cells are indicated in the corresponding quadrants.
General Effects of IFN-α2/α1 Treatment in Newborn Mice.
The systemic effects of IFN-α/β were examined after a 6 d course of treatment initiated on the third day of life. When cells from the bone marrow, spleen, liver, and thymus were examined for Ly-6C expression, enhanced expression of Ly-6C was observed for all of the different cell types (Fig. 2), indicating a strong physiological response to the exogenous IFN-α2/α1 (37).
Figure 2.

Enhanced Ly-6C antigen expression after IFN-α2/α1 treatment. 3-d-old mice received intraperitoneal injections of IFN-α2/α1 (4 × 105 U) or PBS for 6 d, and viable cells from the indicated tissues were stained with anti–Ly-6C antibody 1 d after the last injection. Thin lines represent the staining patterns of cells from control mice; thick lines represent cells from IFN-α2/α1–treated mice.
The IFN-α2/α1 treatment resulted in a pronounced decrease in cellularity of the bone marrow, spleen, and thymus in IFN-α2/α1–treated mice when compared with these tissues in control mice treated with PBS injections (Table 1). The weights of the spleen and thymus were also significantly decreased in mice treated with IFN-α2/α1 (mean ± 1 SD = 39.3 ± 7.5 for experimental spleens versus 52.9 ± 4.3 mg for control spleens, P <0.05; and 10.4 ± 3.2 versus 32.1 ± 2.9 mg for control thymuses, P <0.01). Relatively small differences were seen in the total body weights of control and IFN-α2/α1–treated mice (5.8 ± 0.2 versus 5.2 ± 0.4 g, P <0.05), and both groups of mice were healthy in appearance.
Table 1.
Effect of IFNα2/α1 Treatment on the Cellularity of Hematopoietic and Lymphoid Organs
| Organ | Number of nucleated cells (× 106) | |||
|---|---|---|---|---|
| PBS treatment* | IFN treatment* | |||
| Bone marrow | 27.3 ± 5.5‡ | 7.5 ± 1.3§ | ||
| Spleen | 73.4 ± 14.7 | 20.4 ± 5.2§ | ||
| Thymus | 60.9 ± 15.3 | 8.5 ± 5.2§ | ||
Peritoneal injections of PBS or IFN-α2/α1 (4 × 105 U) were given daily from 3–8 d of age and the mice were killed at 9 d of age.
Mean ± 1 standard error for six mice.
P <0.01, unpaired Student's t test.
Impaired B Cell Development in Mice Treated with IFN-α2/ α1.
When the percentages of B220+ B lineage cells were determined in the bone marrow and spleen of IFN-α2/α1 treated mice and control mice, a 45–60% decrease was observed in the percentages of B220+ B lineage cells in mice treated with IFN-α2/α1 (Table 2). The absolute numbers of B220+ cells, calculated on the basis of bone marrow and spleen cellularity (Table 1), were decreased to an even greater extent (>80%) in IFN-α2/α1–treated mice. When the Mac-1 antigen was employed as a marker of myeloid cells, the percentages of Mac-1+ cells were increased in bone marrow and spleens of IFN-α2/α1–treated mice, whereas a significant decrease was observed in the percentage of Ter119+ erythroid cells (Table 2).
Table 2.
Effect of IFN-α2/α1 Treatment on Lymphoid, Myeloid, and Erythroid Development in Bone Marrow and Spleen
| Treatment* | Bone marrow | Spleen | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B220+ | Mac-1+ | Ter119+ | B220+ | Mac-1+ | Ter119+ | |||||||
| PBS | 21.8 ± 1.8‡ | 23.3 ± 3.0 | 40.3 ± 1.6 | 11.0 ± 2.7 | 3.6 ± 0.4 | 69.7 ± 8.4 | ||||||
| IFNα2/α1 | 8.7 ± 0.9§ | 26.9 ± 2.6 | 26.4 ± 4.5§ | 6.0 ± 0.5§ | 7.4 ± 0.4§ | 45.8 ± 3.4§ | ||||||
Peritoneal injections of PBS or IFN-α2/α1 (4 × 105 U) were administered daily from 3–8 d of age and the mice were killed at 9 d of age.
Mean percentage of marker-positive cells ±1 SE for six mice.
P ⩽0.01, unpaired Student's t test.
An ∼50% decrease in the percentage of B220+IgM+ B cells was observed for the bone marrow and splenic lymphocyte populations in the IFN-α2/α1–treated neonates (Fig. 3). Reductions in the absolute number of B cells were estimated to be 89 and 84% in the bone marrow and spleen, respectively. Although the percentage of residual B220+IgM− B lineage cells was only modestly decreased in the bone marrow of IFN-α2/α1–treated mice (Fig. 3), differences in the staining profiles of B220lo and B220hi populations suggested treatment-induced alteration of the constituent B lineage progenitor subpopulations. Further analysis of bone marrow cells with lymphocyte light-scatter characteristics indicated a twofold reduction in the percentage of CD43−B220+ cells in IFN-α2/α1–treated mice, whereas a twofold increase was observed in the percentage of CD43+B220+ (fractions A–C in the Hardy scheme, reference 9) cells (Fig. 4 A). When the CD43+B220+ population was analyzed for HSA expression, the percentage of CD43+B220+HSA+ cells (fractions B and C) was greatly decreased while the proportion of fraction A cells was increased (Fig. 4 B).
Figure 3.

Immunofluorescence profile analysis of B lineage cells in the bone marrow and spleen of PBS- and IFN-α2/α1–treated mice. Cells were stained with FITC-conjugated anti-μ and Cy-chrome–labeled anti-B220 and the immunocytometric analysis was limited to cells with the forward and side light scattering characteristics of lymphocytes. Percentages of IgM−B220+ and IgM+B220+ cells in the boxed areas are indicated.
Figure 4.
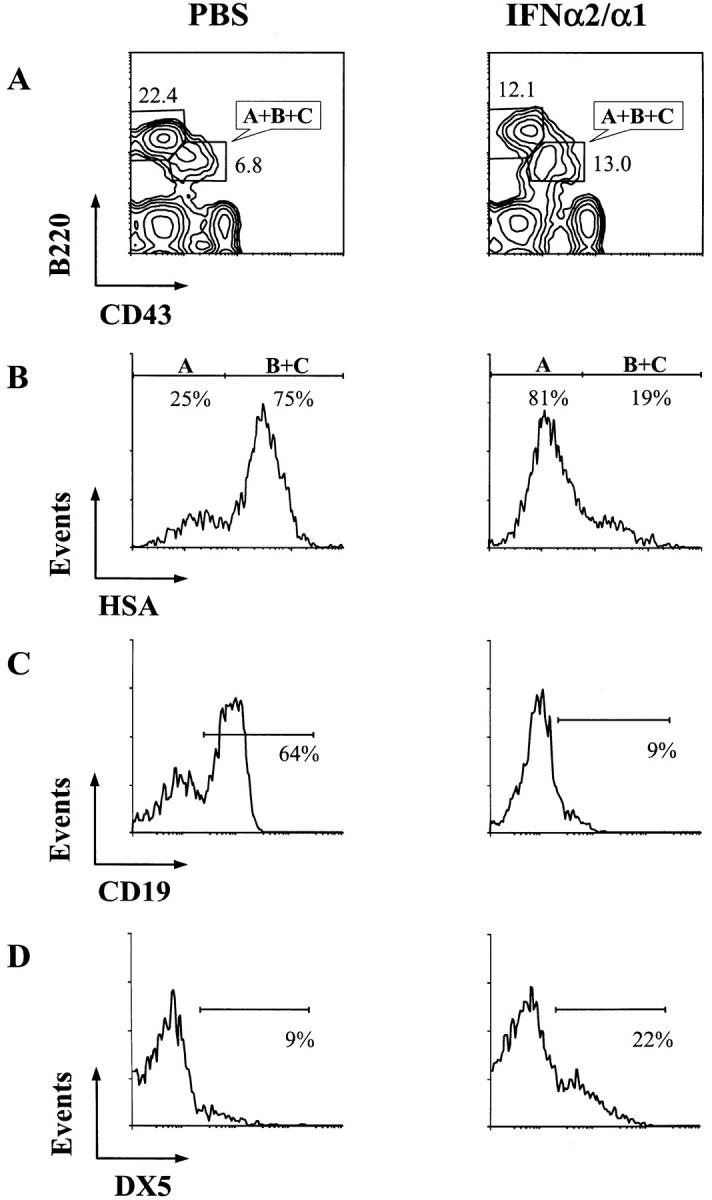
Immunofluorescence profile analysis of B lineage cells in the bone marrow of PBS- and IFN-α2/α1–treated mice. Bone marrow cells were stained with the CD43, HSA, CD19, DX5, and B220 antibodies. (A) Lymphocyte gates were set for forward and side light scattering, and the CD43−B220+ and CD43+B220+ (fractions A, B, and C) subpopulations are indicated as percentages of cells in the two boxes. (B) Fraction A is distinguished from fractions B and C in the CD43+B220+ subpopulation of cells by HSA expression. Fraction A cells were further analyzed for their expression of CD19 as a B lineage marker (C) and of DX5 as an NK lineage marker (D).
Fraction A cells, although B220+, include nonlineage cells as well as B lineage progenitors. This population contains cells that do not express the B lineage marker CD19; some of these CD19− cells belong to the natural killer cell lineage (38). Therefore, CD19 and DX5, a pan–NK cell marker, were employed to analyze the phenotype of fraction A cells. Only 10% of fraction A cells from IFN-α2/ α1–treated mice expressed CD19, whereas in control mice 64% of the cells in this fraction expressed this B lineage marker (Fig. 4 C). In IFN-treated mice there was a 2.5-fold increase in the percentage of DX5+ cells (Fig. 4 D). The reduction of CD19+ B progenitors combined with the reduction in cells representative of later B lineage stages indicates that the IFN-α2/α1 treatment interfered with B lineage development beginning at the pro-B cell stage.
Decreased IL-7 Responsiveness of Bone Marrow Cells from IFN-α2/α1 Treated Mice.
To examine the effects of IFN-α2/α1 treatment on pre-B and B cell response, bone marrow B cells were stimulated with LPS and IL-7 at different concentrations. The proliferative response of early B lineage cells to IL-7 was dramatically inhibited in mice treated with IFN-α2/α1 (Fig. 5), thus providing functional evidence for the early abortion of B lineage differentiation. Diminished LPS responsiveness was also observed for the bone marrow cells in treated mice (data not shown).
Figure 5.

IL-7 responses of bone marrow cells from PBS- and IFN-α2/α1–treated mice. Bone marrow cells were cultured with different concentrations of IL-7 for 72 h, and pulsed with [3H]thymidine for the final 8 h. Results are shown as mean cpm (±SD) from triplicate or quadruplicate cultures.
Impaired T Cell Development in IFN-α2/α1–Treated Mice.
Thymic cellularity was reduced by >85% in IFN-α2/α1– treated mice (Table 1). When IFN-α2/α1 effects on the different thymocyte subpopulations were evaluated by determining the CD4 and CD8 expression, a 50% decrease in the percentage of CD4+CD8+ thymocytes was observed, whereas the relative proportions of CD4+CD8− and CD4−CD8+ thymocytes were increased (Fig. 6 A). To determine more precisely where IFN-α2/α1 inhibited the progression of thymocyte differentiation, we examined the differential expression of CD44 and CD25, cell surface markers that serve to discriminate the CD4−CD8− subpopulations of thymocyte progenitors (15, 16). In this analysis, an increased percentage of the relatively immature CD44+CD25− cells was observed along with a decreased percentage of the more mature CD44−CD25− cells within the CD4−CD8− thymocyte population in IFN-α2/α1–treated mice (Fig. 6 B). The early CD44+CD25− population of thymocytes contains precursors for T cells, NK cells, and dendritic cells (39–41), and the increased frequency of CD44+CD25− cells in the thymus of IL-7−/− mice could in part reflect an increased frequency of NK cells (42). When the CD44+CD25− cells from IFN-treated mice were examined, a twofold increase was indeed observed in the percentage of DX5+ cells (Fig. 6 C). Thus, the composite data indicate that the IFN-α2/ α1 inhibition of T cell development begins at an early progenitor stage.
Figure 6.

Immunofluorescence profile analysis of thymocyte subpopulations from PBS- and IFN-α2/α1–treated mice. Cells were stained with anti-CD4, anti-CD8, anti-CD44, and anti-CD25 antibodies. Percentages of cells within each quadrant are indicated. (A) Analysis of the total thymocyte population for CD4 and CD8 expression; (B) CD25 and CD44 expression by the CD4−CD8− thymocyte subpopulation, and (C) DX5 expression by the CD44+CD25− subpopulation of the CD4−CD8− thymocytes.
Recovery of T and B Cell Development after Cessation of IFN-α2/α1 Treatment.
When T and B cell development was examined as a function of time after discontinuation of the IFN-α2/α1 treatment, no recovery was observed for the T lineage cells in the thymus and spleen or of the B lineage cells in the bone marrow and spleen in the first week after the 6 d course of interferon therapy (Fig. 7). However, by 2 wk after the final injection, mice treated with IFN-α2/ α1 showed significant signs of recovery as reflected by an increase in T and B cell numbers in the thymus, bone marrow, and spleen. By 1 mo after the neonatal treatment, the mice appeared to recover from the effects of the IFN-α2/ α1 in terms of T and B cell numbers.
Figure 7.

Recovery of T and B cell development after neonatal treatment with IFN-α2/α1. 3-d-old mice injected with PBS or IFN-α2/α1 for 6 consecutive d were examined serially after the last treatment. T and B cells were identified by CD3 and B220 expression, respectively. Results of the enumeration of thymocytes, T cells in the spleen, and B cells in the spleen and bone marrow are presented as mean percentages of control values (I = ± standard error; n = 3 mice).
Absence of IFN-α2/α1 Effects in Mice Lacking the Type I IFN Receptor.
To examine the possibility that these inhibitory effects on B and T lymphopoiesis could be attributable to nonspecific cytotoxicity of the recombinant IFN-α2/α1 preparation, we treated type I interferon receptor (IFN-α/ βR) knockout mice (129 Sv/Ev strain) with the recombinant IFN-α2/α1 protein under the same experimental conditions. When treated with IFN-α2/α1, the wild-type 129 mice had an even greater impairment in T and B cell development than did the BALB/c mice, thus suggesting strain differences in interferon sensitivity (Fig. 8). In contrast, T and B cell development was perfectly normal in mice lacking the IFN-α/β receptor after IFN-α2/α1 treatment (Fig. 8). This finding indicates that the inhibitory effect of IFN-α2/α1 on T and B cell development in wild-type mice is mediated via the IFN-α/β receptor.
Figure 8.

Immunofluorescence profiles of bone marrow B cells and thymocytes from PBS- and IFN-α2/α1–treated 129 Sv/Ev wild-type (top) and IFN-α/β receptor−/− mice (bottom). Bone marrow cells were stained with anti-μ and B220 antibodies, and thymocytes were stained with anti-CD4 and anti-CD8. The percentages of cells within each quadrant are indicated.
Discussion
These results indicate that IFN-α2/α1 is a potent, reversible inhibitor of B and T lymphopoiesis in inbred strains of mice. The inhibitory effects begin very early in B and T cell development, being manifested by the pro-B (CD43+B220+CD19+HSA+) and pro-T (CD4−CD8− CD44+CD25+) cell stages. Residual B and T cells were still present in neonatal mice treated with IFN-α2/α1, due either to the limited supply of T and B cells produced before the onset of interferon treatment on the third day of life or to incomplete inhibition of B and T lymphopoiesis by this treatment regimen. In either case, the analysis clearly implies a window of exquisite sensitivity to IFN-α2/α1 during the early stages of T and B cell differentiation.
A trivial explanation of the inhibitory effects would be that the recombinant IFN-α2/α1 preparation contained an unknown cytotoxic contaminant. This possibility was excluded by the fact that IFN-α2/α1 treatment had no effect on the cellular content of the bone marrow, spleen, and thymus in IFN-α/β receptor−/− mice. Specifically, no disruption of B and T cell development was evident in these mice. In addition, expression of the Ly-6C antigen, which serves as a downstream indicator for the interaction of IFN-α/β with its receptor (37), was universally upregulated on hemopoietic cells in wild-type mice treated with IFN-α2/α1. Thus, the observed inhibitory effects of IFN-α2/α1 treatment in wild-type mice can be attributed to the cytokine itself and its interaction with the IFN-α/β receptor.
We previously observed that type I interferons counteract the IL-7–dependent growth and survival of bone marrow B lineage cells, fetal thymocytes, and IL-7–dependent cell lines by reinstating programmed cell death (27, 28, 43). The inhibitory effect of IFN-α/β on IL-7–dependent B lineage cells can be alleviated by upregulated expression of the Bcl-2 gene (27) and upregulated Bcl-2 expression has recently also been shown to rescue T lymphopoiesis in IL-7R−/− mice (44, 45). The possibility that recombinant type I interferon, IFN-α2/α1, exerts its inhibitory effects on lymphocyte development in vivo by counteracting IL-7/ IL-7R–mediated signaling is further supported by evidence indicating that IFN-α2/α1 interferes with B cell development at the CD43+B220+CD19+HSA+ pro–B cell stage and with T cell development at the CD4−CD8−CD44+ CD25+ pro–T cell stage, as these are developmental stages during which IL-7 is required in mice (9, 46–50). A remarkably similar impairment in T and B cell development is seen in mice treated with antibodies to IL-7 or to IL-7R, in mice with IL-7 or IL-7R deficiency (21, 46, 51–53), in common cytokine receptor γ chain (γc)−/− mice (54, 55), and in Jak-3−/− mice (56–58), Jak3 being essential for signal transduction via the IL-7 and other cytokine receptors employing γc (59–63). In IL-7R–deficient mice, B and T cells fail to progress beyond the CD43+B220+HSA− pro-B cell stage in the bone marrow and the CD4−CD8−CD25−, pro-T cell stage in the thymus, respectively (53). B and T cell differentiation are also interrupted at very early stages in mice treated with anti–IL-7 or anti–IL-7R antibodies (21, 46, 51). As in the IFN-α2/α1–treated mice, some mature B220+IgM+ cells are found in the bone marrow and CD4+ and CD8+ thymocytes are not completely eliminated in these mice, unless exposure to the IL-7 antibodies is initiated in utero (46). Thus, the window of sensitivity to IFN-α2/α1 corresponds closely to periods in T and B cell development during which IL-7 → IL-7R signaling is essential. The inhibitory effects of IFN-α2/α1 treatment on T and B cell development thus may reflect counteraction of the IL-7/IL-7R signaling.
The mechanism for type I interferon interference with the signaling pathways triggered by ligated IL-7Rs is unknown. However, interferon regulatory factor 1 (IRF-1), a transactivation factor whose expression is induced by type I and type II interferons (64, 65), may play an important role in this antagonistic effect. IRF-1 binding sites have been identified in the promoter regions of a number of IFN-inducible genes (66). Moreover, a very specific arrest in B lineage differentiation was observed in transgenic mice carrying the human IRF-1 gene driven by a juxtaposed human immunoglobulin heavy-chain enhancer (67). In these transgenic mice, B220+ cells were dramatically reduced in the bone marrow, as well as in the peripheral lymphoid tissues, and bone marrow cells responsive to IL-7 were virtually eliminated. Therefore, IRF-1 overexpression in B lineage cells aborts differentiation at the stage when these cells become IL-7 responsive. Collectively, these observations suggest that one of the key steps in the IFN-α/β inhibition of T and B cell development is the upregulation of IRF-1 activity.
In addition to its inhibitory effects on T and B cell differentiation, IFN-α/β may negatively regulate growth or differentiation of other hematopoietic cells (30–34), as was observed for erythroid cells in this analysis. Interestingly, IFN-α and IFN-β genes are frequently deleted in certain types of malignancies (68, 69). In spite of the compelling evidence for an important IFN-α/β role in regulation of cellular proliferation and differentiation, the only overt phenotypic abnormality in IFN-α/β receptor−/− mice is their diminished resistance to certain viral infections (70). Normal levels of T and B lymphocytes in the IFN-α/β receptor−/− mice could reflect redundancy or compensating activities of other negative regulators, such as IFN-γ (71, 72) or TGF-β (73, 74), which could also interfere with B or T cell development while playing different roles in the immune response to different pathogens. The absence of dramatic phenotypic changes in IFN-α/β receptor−/− mice could reflect the possibility that the interferon effects on B and T cell development are subtle ones in the normal physiological setting. However, during viral infections, severe depression of hematopoiesis has been reported to be a direct effect of enhanced IFN-α/β production (75), thus suggesting a role for type I interferons in the regulation of hematopoietic cellular proliferation and differentiation under conditions of pathological stress. The reversible inhibitory effects observed in IFN-α2/α1–treated mice may therefore be relevant to the pathogenesis of pathogen-induced disturbances in lympho/hemopoiesis.
Acknowledgments
We thank Drs. B.E. Rogers and M.B. Khazaeli for performing the endotoxin measurements, Drs. Peter D. Burrows and Jiyang Wang for critical comments, and Ann Brookshire for help in preparation of the manuscript.
This work has been supported in part by National Institutes of Health grants AI-39816 and AI-34568. M.D. Cooper is a Howard Hughes Medical Institute Investigator.
Footnotes
Abbreviations used in this paper: DN, double negative; HSA, heat-stable antigen; IRF, interferon regulatory factor.
References
- 1.Rajewsky K. Clonal selection and learning in the antibody system. Nature. 1996;381:751–758. doi: 10.1038/381751a0. [DOI] [PubMed] [Google Scholar]
- 2.Tonegawa S. Somatic generation of antibody diversity. Nature. 1983;302:575–581. doi: 10.1038/302575a0. [DOI] [PubMed] [Google Scholar]
- 3.Alt FW, Oltz EM, Young F, Gorman J, Taccioli G, Chen J. VDJ recombination. Immunol Today. 1992;13:306–314. doi: 10.1016/0167-5699(92)90043-7. [DOI] [PubMed] [Google Scholar]
- 4.Karasuyama H, Rolink A, Melchers F. Surrogate light chain in B cell development. Adv Immunol. 1996;63:1–41. doi: 10.1016/s0065-2776(08)60853-6. [DOI] [PubMed] [Google Scholar]
- 5.Lassoued K, Nunez CA, Billips L, Kubagawa H, Monteiro RC, LeBlen TW, Cooper MD. Expression of surrogate light chain receptors is restricted to a late stage in pre–B cell differentiation. Cell. 1993;73:73–86. doi: 10.1016/0092-8674(93)90161-i. [DOI] [PubMed] [Google Scholar]
- 6.Karasuyama H, Rolink A, Shinkai Y, Young F, Alt FW, Melchers F. The expression of Vpre-B/λ5 surrogate light chain in early bone marrow precursor B cells of normal and B cell–deficient mutant mice. Cell. 1994;77:133–143. doi: 10.1016/0092-8674(94)90241-0. [DOI] [PubMed] [Google Scholar]
- 7.Cooper MD, Mulvaney D, Coutinho A, Cazenave P-A. A novel cell surface molecule on early B-lineage cells. Nature. 1986;321:616–618. doi: 10.1038/321616a0. [DOI] [PubMed] [Google Scholar]
- 8.Osmond DG. Population dynamics of bone marrow B lymphocytes. Immunol Rev. 1986;93:103–124. doi: 10.1111/j.1600-065x.1986.tb01504.x. [DOI] [PubMed] [Google Scholar]
- 9.Hardy RR, Carmack CE, Shinton SA, Kemp JD, Hayakawa K. Resolution and characterization of pro-B and pre–pro-B cell stages in normal mouse bone marrow. J Exp Med. 1991;173:1213–1225. doi: 10.1084/jem.173.5.1213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hardy RR, Kemp JD, Hayakawa K. Analysis of lymphoid population in scid mice; detection of a potential B lymphocyte progenitor population present at normal levels in scid mice by three color flow cytometry with B220 and S7. Curr Top Microbiol Immunol. 1989;152:19–25. doi: 10.1007/978-3-642-74974-2_3. [DOI] [PubMed] [Google Scholar]
- 11.Li YS, Hayakawa K, Hardy RR. The regulated expression of B lineage associated genes during B cell differentiation in bone marrow and fetal liver. J Exp Med. 1993;178:951–960. doi: 10.1084/jem.178.3.951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.von Boehmer H. The developmental biology of T lymphocytes. Annu Rev Immunol. 1988;6:309–326. doi: 10.1146/annurev.iy.06.040188.001521. [DOI] [PubMed] [Google Scholar]
- 13.Jameson SC, Hogquist KA, Bevan MJ. Positive selection of thymocytes. Annu Rev Immunol. 1995;13:93–126. doi: 10.1146/annurev.iy.13.040195.000521. [DOI] [PubMed] [Google Scholar]
- 14.Kisielow P, von Boehmer H. Development and selection of T cells: facts and puzzles. Adv Immunol. 1995;58:87–209. doi: 10.1016/s0065-2776(08)60620-3. [DOI] [PubMed] [Google Scholar]
- 15.Godfrey DI, Kennedy J, Suda T, Zlotnik A. A developmental pathway involving four phenotypically and functionally distinct subsets of CD3−CD4−CD8−triple-negative adult mouse thymocytes defined by CD44 and CD25 expression. J Immunol. 1993;150:4244–4252. [PubMed] [Google Scholar]
- 16.Godfrey DI, Zlotnik A. Control points in early T-cell development. Immunol Today. 1993;14:547–553. doi: 10.1016/0167-5699(93)90186-O. [DOI] [PubMed] [Google Scholar]
- 17.Levelt CN, Carsetti R, Eichmann K. Regulation of thymocyte development through CD3. II. Expression of T cell receptor β CD3 ε and maturation to the CD4+8+stage are highly correlated in individual thymocytes. J Exp Med. 1993;178:1867–1875. doi: 10.1084/jem.178.6.1867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Dudley EC, Petrie HT, Shah LM, Owen MJ, Hayday AC. T cell receptor beta chain gene rearrangement and selection during thymocyte development in adult mice. Immunity. 1994;1:83–93. doi: 10.1016/1074-7613(94)90102-3. [DOI] [PubMed] [Google Scholar]
- 19.Godfrey DI, Kennedy J, Mombaerts P, Tonegawa S, Zlotnik A. Onset of TCR-beta gene rearrangement and role of TCR-beta expression during CD3−CD4−CD8−thymocyte differentiation. J Immunol. 1994;152:4783–4792. [PubMed] [Google Scholar]
- 20.Namen AE, Lupton S, Hjerrild K, Wignall J, Mochizuki DY, Schmierer A, Mosley B, March CJ, Urdal D, Gillis S, et al. Stimulation of B-cell progenitors by cloned murine interleukin-7. Nature. 1988;333:571–573. doi: 10.1038/333571a0. [DOI] [PubMed] [Google Scholar]
- 21.Sudo T, Nishikawa S, Ohno N, Akiyama N, Tamakoshi M, Yoshida H, Nishikawa S-I. Expression and function of the interleukin 7 receptor in murine lymphocytes. Proc Natl Acad Sci USA. 1993;90:9125–9129. doi: 10.1073/pnas.90.19.9125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kincade PW. B lymphopoiesis: global factors, local control. Proc Natl Acad Sci USA. 1994;91:2888–2889. doi: 10.1073/pnas.91.8.2888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Melchers F, Haasner D, Streb M, Rolink A. B-lymphocyte lineage-committed, IL-7 and stroma cell-reactive progenitors and precursors, and their differentiation to B cells. Adv Exp Med Biol. 1992;323:111–117. doi: 10.1007/978-1-4615-3396-2_14. [DOI] [PubMed] [Google Scholar]
- 24.Kunisada T, Ogawa M, Hayashi S, Era T, Nishikawa S. Interleukin-7 and B lymphopoiesis. Chem Immunol. 1992;51:205–235. doi: 10.1159/000420760. [DOI] [PubMed] [Google Scholar]
- 25.Namen AE, Williams DE, Goodwin RG. Interleukin-7: a new hematopoietic growth factor. Prog Clin Biol Res. 1990;338:65–73. [PubMed] [Google Scholar]
- 26.Widmer, M.B., P.J. Morrissey, R.G. Goodwin, K.H. Grabstein, L.S. Park, J.D. Watson, P.W. Kincade, P.J. Conlon, and A.E. Namen. 1990. Lymphopoiesis and IL-7. Intl. J. Cell Cloning. 8 (Suppl. 1):168–172. [DOI] [PubMed]
- 27.Wang J, Lin Q, Langston H, Cooper MD. Resident bone marrow macrophages produce type 1 interferons that can selectively inhibit interleukin-7–driven growth of B lineage cells. Immunity. 1995;3:475–484. doi: 10.1016/1074-7613(95)90176-0. [DOI] [PubMed] [Google Scholar]
- 28.Su D-M, Wang J, Lin Q, Cooper MD, Watanabe T. Interferons α/β inhibit IL-7–induced proliferation of CD4−CD8−CD3−CD44+CD25+ thymocytes, but do not inhibit CD4−CD8−CD3−CD44−CD25−thymocytes. Immunology. 1997;90:543–549. doi: 10.1046/j.1365-2567.1997.00205.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.De Maeyer, E., and J. De Maeyer-Guignard. 1988. Interferons and Other Regulatory Cytokines. John Wiley and Sons, Inc. New York. 448 pp.
- 30.Borden EC, Hogan TF, Voelkel JG. Comparative antiproliferative activity in vitro of natural interferons alpha and beta for diploid and transformed human cells. Cancer Res. 1982;42:4948–4953. [PubMed] [Google Scholar]
- 31.Broxmeyer HE, Lu L, Platzer E, Feit C, Juliano L, Rubin BY. Comparative analysis of the influences of human gamma, alpha and beta interferons on human multipotential (CFU-GEMM), erythroid (BFU-E) and granulocyte-macrophage (CFU-GM) progenitor cells. J Immunol. 1983;131:1300–1305. [PubMed] [Google Scholar]
- 32.Clemens MJ, McNurlan MA. Regulation of cell proliferation and differentiation by interferons. Biochem J. 1985;226:345–360. doi: 10.1042/bj2260345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Moore RN, Larsen HS, Horohov DW, Rouse BT. Endogenous regulation of macrophage proliferative expansion by colony-stimulating factor-induced interferon. Science. 1984;223:178–181. doi: 10.1126/science.6606850. [DOI] [PubMed] [Google Scholar]
- 34.Resnitzky D, Yarden A, Zipori D, Kimchi A. Autocrine beta-related interferon controls c-myc suppression and growth arrest during hematopoietic cell differentiation. Cell. 1986;46:31–40. doi: 10.1016/0092-8674(86)90857-3. [DOI] [PubMed] [Google Scholar]
- 35.Weber H, Valenzuela D, Lujber G, Gubler M, Weissmann C. Single amino acid changes that render human IFN-alpha 2 biologically active on mouse cells. EMBO (Eur Mol Biol Organ) J. 1987;6:591–598. doi: 10.1002/j.1460-2075.1987.tb04795.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Borzillo GV, Endo K, Tsujimoto Y. Bcl-2 confers growth and survival advantage to interleukin 7–dependent early pre-B cells which become factor independent by a multistep process in culture. Oncogene. 1992;7:869–876. [PubMed] [Google Scholar]
- 37.Dumont FJ, Coker LZ. Interferon-alpha/beta enhances the expression of Ly-6 antigens on T cells in vivo and in vitro. Eur J Immunol. 1986;16:735–740. doi: 10.1002/eji.1830160704. [DOI] [PubMed] [Google Scholar]
- 38.Rolink A, ten Boekel E, Melchers F, Fearon DT, Krop I, Andersson J. A subpopulation of B220+cells in murine bone marrow does not express CD19 and contains natural killer cell progenitors. J Exp Med. 1996;183:187–194. doi: 10.1084/jem.183.1.187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Moore TA, Zlotnik A. T-cell lineage commitment and cytokine responses of thymic progenitors. Blood. 1995;86:1850–1860. [PubMed] [Google Scholar]
- 40.Wu L, Li CL, Shortman K. Thymic dendritic cell precursors: relationship to the T lymphocyte lineage and phenotype of the dendritic cell progeny. J Exp Med. 1996;184:903–911. doi: 10.1084/jem.184.3.903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ardavin C, Wu L, Li CL, Shortman K. Thymic dendritic cells and T cells develop simultaneously in the thymus from a common precursor population. Nature. 1993;362:761–763. doi: 10.1038/362761a0. [DOI] [PubMed] [Google Scholar]
- 42.Moore TA, von Freeden-Jeffry U, Murray R, Zlotnik A. Inhibition of γδ T cell development and early thymocyte maturation in IL-7 −/−mice. J Immunol. 1996;157:2366–2373. [PubMed] [Google Scholar]
- 43.Wang J, Walker H, Lin Q, Jenkins N, Copeland NG, Watanabe T, Burrows PD, Cooper MD. The mouse BP-1gene: structure, chromosomal localization, and regulation of expression by type I interferons and interleukin-7. Genomics. 1996;33:167–176. doi: 10.1006/geno.1996.0180. [DOI] [PubMed] [Google Scholar]
- 44.Akashi K, Kondo M, von Freeden-Jeffry U, Murray R, Weissman IL. Bcl-2 rescues T lymphopoiesis in interleukin-7 receptor-deficient mice. Cell. 1997;89:1033–1041. doi: 10.1016/s0092-8674(00)80291-3. [DOI] [PubMed] [Google Scholar]
- 45.Maraskovsky E, O'Reilly LA, Teepe M, Corcoran LM, Peschon JJ, Strasser A. Bcl-2 can rescue T lymphocyte development in interleukin-7 receptor-deficient mice but not in mutant rag-1 −/−mice. Cell. 1997;89:1011–1019. doi: 10.1016/s0092-8674(00)80289-5. [DOI] [PubMed] [Google Scholar]
- 46.Grabstein KH, Waldschmidt TJ, Finkelman FD, Hess BW, Alpert AR, Boiani NE, Namen AE, Morrissey PJ. Inhibition of murine B and T lymphopoiesis in vivo by an anti–interleukin 7 monoclonal antibody. J Exp Med. 1993;178:257–264. doi: 10.1084/jem.178.1.257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Watson JD, Morrissey PJ, Namen AE, Conlon PJ, Widmer MB. Effect of IL-7 on the growth of fetal thymocytes in culture. J Immunol. 1989;143:1215–1222. [PubMed] [Google Scholar]
- 48.Suda T, Zlotnik A. IL-7 maintains the T cell precursor potential of CD3−CD4−CD8−thymocytes. J Immunol. 1991;146:3068–3073. [PubMed] [Google Scholar]
- 49.Plum J, De Smedt M, Leclercq G. Exogenous IL-7 promotes the growth of CD3−CD4−CD8−CD44+ CD25+/−precursor cells and blocks the differentiation pathway of TCR-alpha beta cells in fetal thymus organ culture. J Immunol. 1993;150:2706–2716. [PubMed] [Google Scholar]
- 50.Candéias S, Muegge K, Durum SK. IL-7 receptor and VDJ recombination: trophic versus mechanistic actions. Immunity. 1997;6:501–508. doi: 10.1016/s1074-7613(00)80338-6. [DOI] [PubMed] [Google Scholar]
- 51.Bhatia SK, Tygrett LT, Grabstein KH, Waldschmidt TJ. The effect of in vivo IL-7 deprivation on T cell maturation. J Exp Med. 1995;181:1399–1409. doi: 10.1084/jem.181.4.1399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.von Freeden-Jeffry U, Vieira P, Lucian LA, McNeil T, Burdach SE, Murray R. Lymphopenia in interleukin (IL)-7 gene-deleted mice identifies IL-7 as a nonredundant cytokine. J Exp Med. 1995;181:1519–1526. doi: 10.1084/jem.181.4.1519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Peschon JJ, Morrissey PJ, Grabstein KH, Ramsdell FJ, Maraskovsky E, Gliniak BC, Park LS, Ziegler SF, Williams DE, Ware CB, Meyer JD, Davison BL. Early lymphocyte expansion is severely impaired in interleukin 7 receptor-deficient mice. J Exp Med. 1994;180:1955–1960. doi: 10.1084/jem.180.5.1955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Cao X, Shores EW, Hu-Li J, Anver MR, Kelsall BL, Russell SM, Drago J, Noguchi M, Grinberg A, Bloom ET, et al. Defective lymphoid development in mice lacking expression of the common cytokine receptor γ chain. Immunity. 1997;2:223–238. doi: 10.1016/1074-7613(95)90047-0. [DOI] [PubMed] [Google Scholar]
- 55.DiSanto JP, Muller W, Guy-Grand D, Fischer A, Rajewsky K. Lymphoid development in mice with a targeted deletion of the interleukin 2 receptor gamma chain. Proc Natl Acad Sci USA. 1995;92:377–381. doi: 10.1073/pnas.92.2.377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Nosaka T, van Deursen JM, Tripp RA, Thierfelder WE, Witthuhn BA, McMickle AP, Doherty PC, Grosveld GC, Ihle JN. Defective lymphoid development in mice lacking Jak3. Science. 1995;270:800–802. doi: 10.1126/science.270.5237.800. [DOI] [PubMed] [Google Scholar]
- 57.Park SY, Saijo K, Takahashi T, Osawa M, Arase H, Hirayama N, Miyake K, Nakauchi H, Shirasawa T, Saito T. Developmental defects of lymphoid cells in Jak3 kinase-deficient mice. Immunity. 1995;3:771–782. doi: 10.1016/1074-7613(95)90066-7. [DOI] [PubMed] [Google Scholar]
- 58.Thomis DC, Gurniak CB, Tivol E, Sharpe AH, Berg LJ. Defects in B lymphocyte maturation and T lymphocyte activation in mice lacking Jak3 defects in B lymphocyte maturation and T lymphocyte activation in mice lacking Jak3. Science. 1995;270:794–797. doi: 10.1126/science.270.5237.794. [DOI] [PubMed] [Google Scholar]
- 59.Witthuhn BA, Silvennoinen O, Miura O, Lai KS, Cwik C, Liu ET, Ihle JN. Involvement of the Jak-3 Janus kinase in signalling by interleukins 2 and 4 in lymphoid and myeloid cells. Nature. 1994;370:153–157. doi: 10.1038/370153a0. [DOI] [PubMed] [Google Scholar]
- 60.Johnston JA, Kawamura M, Kirken RA, Chen YQ, Blake TB, Shibuya K, Ortaldo JR, McVicar DW, O'Shea JJ. Phosphorylation and activation of the Jak-3 Janus kinase in response to interleukin-2. Nature. 1994;370:151–153. doi: 10.1038/370151a0. [DOI] [PubMed] [Google Scholar]
- 61.Kondo M, Takeshita T, Higuchi M, Nakamura M, Sudo T, Nishikawa S, Sugamura K. Functional participation of the IL-2 receptor gamma chain in IL-7 receptor complexes. Science. 1994;263:1453–1454. doi: 10.1126/science.8128231. [DOI] [PubMed] [Google Scholar]
- 62.Ihle JN. Cytokine receptor signalling. Nature. 1995;377:591–594. doi: 10.1038/377591a0. [DOI] [PubMed] [Google Scholar]
- 63.Sharfe N, Dadi HK, Roifman CM. JAK3 protein tyrosine kinase mediates interleukin-7–induced activation of phosphatidylinositol-3′ kinase. Blood. 1995;86:2077–2085. [PubMed] [Google Scholar]
- 64.Harada H, Fujita T, Miyamoto M, Kimura Y, Maruyama M, Furia A, Miyata T, Taniguchi T. Structurally similar but functionally distinct factors, IRF-1 and IRF-2, bind to the same regulatory elements of IFN and IFN-inducible genes. Cell. 1989;58:729–739. doi: 10.1016/0092-8674(89)90107-4. [DOI] [PubMed] [Google Scholar]
- 65.Fujita T, Reis LF, Watanabe N, Kimura Y, Taniguchi T, Vilcek J. Induction of the transcription factor IRF-1 and interferon-beta mRNAs by cytokines and activators of second-messenger pathways. Proc Natl Acad Sci USA. 1989;86:9936–9940. doi: 10.1073/pnas.86.24.9936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Miyamoto M, Fujita T, Kimura Y, Maruyama M, Harada H, Sudo Y, Miyata T, Taniguchi T. Regulated expression of a gene encoding a nuclear factor, IRF-1, that specifically binds to IFN-beta gene regulatory elements. Cell. 1988;54:903–913. doi: 10.1016/s0092-8674(88)91307-4. [DOI] [PubMed] [Google Scholar]
- 67.Yamada G, Ogawa M, Akagi K, Miyamoto H, Nakano N, Itoh S, Miyazaki J-I, Nishikawa S-I, Yamamura K-I, Taniguchi T. Specific depletion of the B-cell population induced by aberrant expression of human interferon regulatory factor 1 gene in transgenic mice. Proc Natl Acad Sci USA. 1991;88:532–536. doi: 10.1073/pnas.88.2.532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Diaz MO, Ziemin S, Le Beau MM, Pitha P, Smith SD, Chilcote RR, Rowley JD. Homozygous deletion of the alpha- and beta 1-interferon genes in human leukemia and derived cell lines. Proc Natl Acad Sci USA. 1988;85:5259–5263. doi: 10.1073/pnas.85.14.5259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Miyakoshi J, Dobler KD, Allalunis-Turner J, McKean JDS, Petruk K, Allen PBR, Aronyk KN, Weir B, Huyser-Wierenga D, Fulton D, et al. Absence of IFNA and IFNBgenes from human malignant glioma cell lines and lack of correlation with cellular sensitivity to interferons. Cancer Res. 1990;50:278–283. [PubMed] [Google Scholar]
- 70.Muller U, Steinhoff U, Reis LF, Hemmi S, Pavlovic J, Zinkernagel RM, Aguet M. Functional role of type I and type II interferons in antiviral defense. Science. 1994;264:1918–1921. doi: 10.1126/science.8009221. [DOI] [PubMed] [Google Scholar]
- 71.Grawunder U, Melchers F, Rolink A. Interferon-gamma arrests proliferation and causes apoptosis in stromal cell/interleukin-7–dependent normal murine pre-B cell lines and clones in vitro, but does not induce differentiation to surface immunoglobulin-positive B cells. Eur J Immunol. 1993;23:544–551. doi: 10.1002/eji.1830230237. [DOI] [PubMed] [Google Scholar]
- 72.Hwang SY, Hertzog PJ, Holland KA, Sumarsono SH, Tymms MJ, Hamilton JA, Whitty G, Bertoncello I, Kola I. A null mutation in the gene encoding a type I interferon receptor component eliminates antiproliferative and antiviral responses to interferons alpha and beta and alters macrophage responses. Proc Natl Acad Sci USA. 1995;92:11284–11288. doi: 10.1073/pnas.92.24.11284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Lee G, Namen AE, Gillis S, Ellingsworth LR, Kincade PW. Normal B cell precursors responsive to recombinant murine IL-7 and inhibition of IL-7 activity by transforming growth factor-beta. J Immunol. 1989;142:3875–3883. [PubMed] [Google Scholar]
- 74.Plum J, De Smedt M, Leclercq G, Vandekerckhove B. Influence of TGF-beta on murine thymocyte development in fetal thymus organ culture. J Immunol. 1995;154:5789–5798. [PubMed] [Google Scholar]
- 75.Binder D, Fehr J, Hengartner H, Zinkernagel RM. Virus-induced transient bone marrow aplasia: major role of interferon-α/β during acute infection with the noncytopathic lymphocytic choriomeningitis virus. J Exp Med. 1997;185:517–530. doi: 10.1084/jem.185.3.517. [DOI] [PMC free article] [PubMed] [Google Scholar]